
Role of Innate and Adaptive Cytokines in the Survival of COVID-19 Patients

 ,
, 
 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Patients
2.2. Cytokine Results
2.3. Serum Levels of Chemokines
2.4. Serum Levels of Growth Factors Acting on Hematopoietic and Immune System Cells (Innate and Adaptive Growth Factors)
2.5. Serum Levels of Cytokines Involved in the Adaptive Immune System Response
2.6. Serum Levels of Wound-Healing and Tissue-Repair Growth Factors
2.7. Survival Predictive Value Analysis of the Serum Soluble Factor Levels in Severe COVID-19 Patients
3. Discussion
4. Materials and Methods
4.1. Study Cohorts and Inclusion and Exclusion Criteria
4.2. Study Protocol and Assays of Cytokine Serum Levels
4.3. Statistical Analysis
4.4. Ethics and Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Fact Sheet. WHO Health Emergency Dashboard. 2022. Available online: https://extranet.who.int/publicemergency (accessed on 17 July 2022).
- Sanz, J.M.; Gomez Lahoz, A.M.; Martin, R.O. Role of the immune system in SARS-CoV-2 infection: Immunopathology of COVID-19. Medicine 2021, 13, 1917–1931. [Google Scholar] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; Macary, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Rowaiye, A.B.; Okpalefe, O.A.; Onuh, A.O.; Ogidigo, J.O.; Hannah, O.O.; Ogu, A.C.; Oli, A.N.; Olofinase, S.; Onyekwere, O.; Abubakar, A.R.; et al. Attenuating the Effects of Novel COVID-19 (SARS-CoV-2) Infection-Induced Cytokine Storm and the Implications. J. Inflamm. Res. 2021, 14, 1487–1510. [Google Scholar] [CrossRef]
- Gomez-Rial, J.; Rivero-Calle, I.; Salas, A.; Martinon-Torres, F. Role of Monocytes/Macrophages in COVID-19 Pathogenesis: Implications for Therapy. Infect. Drug Resist. 2020, 13, 2485–2493. [Google Scholar] [CrossRef]
- Grom, A.A.; Horne, A.; De, B.F. Macrophage activation syndrome in the era of biologic therapy. Nat. Rev. Rheumatol. 2016, 12, 259–268. [Google Scholar] [CrossRef]
- Otsuka, R.; Seino, K.I. Macrophage activation syndrome and COVID-19. Inflamm. Regen. 2020, 40, 19. [Google Scholar] [CrossRef]
- Schulert, G.S.; Grom, A.A. Pathogenesis of macrophage activation syndrome and potential for cytokine-directed therapies. Annu. Rev. Med. 2015, 66, 145–159. [Google Scholar] [CrossRef]
- Darif, D.; Hammi, I.; Kihel, A.; El Idrissi Saik, I.; Guessous, F.; Akarid, K. The pro-inflammatory cytokines in COVID-19 pathogenesis: What goes wrong? Microb. Pathog. 2021, 153, 104799. [Google Scholar] [CrossRef]
- Farshidfar, F.; Koleini, N.; Ardehali, H. Cardiovascular complications of COVID-19. JCI Insight 2021, 6, e148980. [Google Scholar] [CrossRef]
- Foxwell, B.M.; Barrett, K.; Feldmann, M. Cytokine receptors: Structure and signal transduction. Clin. Exp. Immunol. 1992, 90, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.; Wang, J.M. Chemokines: The past, the present and the future. Cell Mol. Immunol. 2018, 15, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ye, D.; Wang, Z.; Pan, H.; Lu, X.; Wang, M.; Xu, Y.; Yu, J.; Zhang, J.; Zhao, M.; et al. The Role of Interleukin-6 Family Members in Cardiovascular Diseases. Front. Cardiovasc. Med. 2022, 9, 818890. [Google Scholar] [CrossRef] [PubMed]
- Briukhovetska, D.; Dorr, J.; Endres, S.; Libby, P.; Dinarello, C.A.; Kobold, S. Interleukins in cancer: From biology to therapy. Nat. Rev. Cancer 2021, 21, 481–499. [Google Scholar] [CrossRef]
- McInnes, I. Cytokines. In Kelley’s Textbook of Rheumatology; Elsevier Inc.: Cambridge, MA, USA, 2013; pp. 369–381. [Google Scholar]
- Ozger, H.S.; Karakus, R.; Kuscu, E.N.; Bagriacik, U.E.; Oruklu, N.; Yaman, M.; Turkoglu, M.; Erbas, G.; Atak, A.Y.; Senol, E. Serial measurement of cytokines strongly predict COVID-19 outcome. PLoS ONE 2021, 16, e0260623. [Google Scholar] [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.J.; Yu, W.C.; Peng, G.R.; Ye, C.H.; Hu, S.; Chong, P.C.T.; Yap, K.Y.; Lee, J.Y.C.; Lin, W.-C.; Yu, S.-H. The Role of Cytokines and Chemokines in Severe Acute Respiratory Syndrome Coronavirus 2 Infections. Front. Immunol. 2022, 13, 832394. [Google Scholar] [CrossRef]
- Khalil, B.A.; Elemam, N.M.; Maghazachi, A.A. Chemokines and chemokine receptors during COVID-19 infection. Comput. Struct. Biotechnol. J. 2021, 19, 976–988. [Google Scholar] [CrossRef]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Meizlish, M.L.; Pine, A.B.; Goshua, G.; Chang, C.H.; Zhang, H.; Bishai, J.; Bahel, P.; Patel, A.; Gbyli, R.; Kwan, J.; et al. Circulating Markers of Angiogenesis and Endotheliopathy in COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Petrey, A.C.; Qeadan, F.; Middleton, E.A.; Pinchuk, I.V.; Campbell, R.A.; Beswick, E.J. Cytokine release syndrome in COVID-19: Innate immune, vascular, and platelet pathogenic factors differ in severity of disease and sex. J. Leukoc. Biol. 2021, 109, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Tamayo-Velasco, A.; Martínez-Paz, P.; Peñarrubia-Ponce, M.; de la Fuente, I.; Pérez-González, S.; Fernández, I.; Dueñas, C.; Gómez-Sánchez, E.; Lorenzo-López, M.; Gómez-Pesquera, E.; et al. HGF, IL-1α, and IL-27 Are Robust Biomarkers in Early Severity Stratification of COVID-19 Patients. J. Clin. Med. 2021, 10, 2017. [Google Scholar] [CrossRef]
- Cabaro, S.; D’Esposito, V.; Di Matola, T.; Sale, S.; Cennamo, M.; Terracciano, D.; Parisi, V.; Oriente, F.; Portella, G.; Beguinot, F.; et al. Cytokine signature and COVID-19 prediction models in the two waves of pandemics. Sci. Rep. 2021, 11, 20793. [Google Scholar] [CrossRef] [PubMed]
- Moll-Bernardes, R.; de Sousa, A.S.; Macedo, A.V.S.; Lopes, R.D.; Vera, N.; Maia, L.C.R.; Feldman, A.; Arruda, G.D.A.S.; Castro, M.J.C.; Pimentel-Coelho, P.M.; et al. IL-10 and IL-12 (P70) Levels Predict the Risk of COVID-19 Progression in Hypertensive Patients: Insights From the BRACE-CORONA Trial. Front. Cardiovasc. Med. 2021, 8, 702507. [Google Scholar] [CrossRef] [PubMed]
- Mudd, P.A.; Crawford, J.C.; Turner, J.S.; Souquette, A.; Reynolds, D.; Bender, D.; Bosanquet, J.P.; Anand, N.J.; Striker, D.A.; Martin, R.S.; et al. Distinct inflammatory profiles distinguish COVID-19 from influenza with limited contributions from cytokine storm. Sci. Adv. 2020, 6, eabe3024. [Google Scholar] [CrossRef]
- Petrone, L.; Petruccioli, E.; Vanini, V.; Cuzzi, G.; Najafi, F.S.; Alonzi, T.; Castilletti, C.; Palmieri, F.; Gualano, G.; Vittozzi, P.; et al. A whole blood test to measure SARS-CoV-2-specific response in COVID-19 patients. Clin. Microbiol. Infect. 2021, 27, 286. [Google Scholar] [CrossRef]
- Tan, Y.; Tang, F. SARS-CoV-2-mediated immune system activation and potential application in immunotherapy. Med. Res. Rev. 2021, 41, 1167–1194. [Google Scholar] [CrossRef]
- Venkataraman, A.; Kumar, N.P.; Hanna, L.E.; Putlibai, S.; Karthick, M.; Rajamanikam, A.; Sadasivam, K.; Sundaram, B.; Babu, S. Plasma biomarker profiling of PIMS-TS, COVID-19 and SARS-CoV2 seropositive children—A cross-sectional observational study from southern India. EBioMedicine 2021, 66, 103317. [Google Scholar] [CrossRef]
- Youngs, J.; Provine, N.M.; Lim, N.; Sharpe, H.R.; Amini, A.; Chen, Y.-L.; Luo, J.; Edmans, M.D.; Zacharopoulou, P.; Chen, W.; et al. Identification of immune correlates of fatal outcomes in critically ill COVID-19 patients. PLoS Pathog. 2021, 17, e1009804. [Google Scholar] [CrossRef]
- World Health Organization. Director-General’s Remarks at the Media Briefing on 2019-nCoV on 11 February 2020. 2022. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-remarks-at-the-media-briefing-on-2019-ncov-on-11-february-2020 (accessed on 17 July 2022).
- Chen, Y.; Wang, J.; Liu, C.; Su, L.; Zhang, D.; Fan, J.; Yang, Y.; Xiao, M.; Xie, J.; Xu, Y.; et al. IP-10 and MCP-1 as biomarkers associated with disease severity of COVID-19. Mol. Med. 2020, 26, 97. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Leem, A.Y.; Chung, K.S.; Ahn, J.Y.; Jung, J.Y.; Kang, Y.A.; Park, M.S.; Kim, Y.S.; Lee, S.H. Soluble IL-2R Levels Predict in-Hospital Mortality in COVID-19 Patients with Respiratory Failure. J. Clin. Med. 2021, 10, 4242. [Google Scholar] [CrossRef] [PubMed]
- Monserrat, J.; Asunsolo, A.; Gómez-Lahoz, A.; Ortega, M.A.; Gasalla, J.M.; Gasulla, Ó.; Fortuny-Profitós, J.; Mazaira-Font, F.A.; Teixidó Román, M.; Arranz, A.; et al. Impact of the Innate Inflammatory Response on ICU Admission and Death in Hospitalized Patients with COVID-19. Biomedicines 2021, 9, 1675. [Google Scholar] [CrossRef] [PubMed]
- Nagant, C.; Ponthieux, F.; Smet, J.; Dauby, N.; Doyen, V.; Besse-Hammer, T.; De Bels, D.; Maillart, E.; Corazza, F. A score combining early detection of cytokines accurately predicts COVID-19 severity and intensive care unit transfer. Int. J. Infect. Dis. 2020, 101, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shen, C.; Li, J.; Yuan, J.; Wei, J.; Huang, F.; Wang, F.; Li, G.; Xing, L.; Peng, L.; et al. Plasma IP-10 and MCP-3 levels are highly associated with disease severity and predict the progression of COVID-19. J. Allergy Clin. Immunol. 2020, 146, 119–127. [Google Scholar] [CrossRef]
- Dinarello, C.A. The IL-1 family of cytokines and receptors in rheumatic diseases. Nat. Rev. Rheumatol. 2019, 15, 612–632. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef]
- Challagundla, N.; Saha, B.; Agrawal-Rajput, R. Insights into inflammasome regulation: Cellular, molecular, and pathogenic control of inflammasome activation. Immunol. Res. 2022, 1–29. [Google Scholar] [CrossRef]
- Schaper, F.; Rose-John, S. Interleukin-6: Biology, signaling and strategies of blockade. Cytokine Growth Factor Rev. 2015, 26, 475–487. [Google Scholar] [CrossRef]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and its receptors: A highly regulated and dynamic system. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef]
- Marc, F.; Moldovan, C.M.; Hoza, A.; Magheru, S.; Ciavoi, G.; Farcas, D.M.; Sachelarie, L.; Calin, G.; Romila, L.; Damir, D.; et al. Comparative Study of Cytokine Storm Treatment in Patients with COVID-19 Pneumonia Using Immunomodulators. J. Clin. Med. 2022, 11, 2945. [Google Scholar] [CrossRef]
- Gupta, R. Could anakinra outmatch dexamethasone/tocilizumab in COVID-19? Bull Natl. Res. Cent. 2022, 46, 100. [Google Scholar] [CrossRef] [PubMed]
- Naveed, Z.; Sarwar, M.; Ali, Z.; Saeed, D.; Choudhry, K.; Sarfraz, A.; Sarfraz, Z.; Felix, M.; Cherrez-Ojeda, I. Anakinra treatment efficacy in reduction of inflammatory biomarkers in COVID-19 patients: A meta-analysis. J. Clin. Lab. Anal. 2022, 36, e24434. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Mizoguchi, I.; Chiba, Y.; Ohashi, M.; Xu, M.; Yoshimoto, T. Expanding Diversity in Molecular Structures and Functions of the IL-6/IL-12 Heterodimeric Cytokine Family. Front. Immunol. 2016, 7, 479. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tian, Z.; Wei, H. Developmental and Functional Control of Natural Killer Cells by Cytokines. Front. Immunol. 2017, 8, 930. [Google Scholar] [CrossRef] [PubMed]
- Pillet, A.H.; Theze, J.; Rose, T. Interleukin (IL)-2 and IL-15 have different effects on human natural killer lymphocytes. Hum. Immunol. 2011, 72, 1013–1017. [Google Scholar] [CrossRef]
- David, B.A.; Kubes, P. Exploring the complex role of chemokines and chemoattractants in vivo on leukocyte dynamics. Immunol. Rev. 2019, 289, 9–30. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef]
- Teixeira, A.L.; Gama, C.S.; Rocha, N.P.; Teixeira, M.M. Revisiting the Role of Eotaxin-1/CCL11 in Psychiatric Disorders. Front. Psychiatry 2018, 9, 241. [Google Scholar] [CrossRef]
- Chen, X.; Wei, Q.; Hu, Y.; Wang, C. Role of Fractalkine in promoting inflammation in sepsis-induced multiple organ dysfunction. Infect. Genet. Evol. 2020, 85, 104569. [Google Scholar] [CrossRef]
- Rivas-Fuentes, S.; Valdes, V.J.; Espinosa, B.; Gorocica-Rosete, P.; Salgado-Aguayo, A. Could SARS-CoV-2 blocking of ACE2 in endothelial cells result in upregulation of CX3CL1, promoting thrombosis in COVID-19 patients? Med. Hypotheses 2021, 151, 110570. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [PubMed]
- Kolios, A.G.A.; Tsokos, G.C.; Klatzmann, D. Interleukin-2 and regulatory T cells in rheumatic diseases. Nat. Rev. Rheumatol. 2021, 17, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, H.M.; Gale, R.P. G-CSF and GM-CSF Are Different. Which One Is Better for COVID-19? Acta Haematol. 2021, 144, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.X.; Leonard, W.J. The Common Cytokine Receptor gamma Chain Family of Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028449. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Jia, J.A.; Li, H.; Wan, N.; Zhang, S.; Ma, X. Lymphocyte-monocyte-neutrophil index: A predictor of severity of coronavirus disease 2019 patients produced by sparse principal component analysis. Virol. J. 2021, 18, 115. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar] [CrossRef]
- Boehlen, F.; Clemetson, K.J. Platelet chemokines and their receptors: What is their relevance to platelet storage and transfusion practice? Transfus. Med. 2001, 11, 403–417. [Google Scholar] [CrossRef]
- Blair, P.; Flaumenhaft, R. Platelet α-granules: Basic biology and clinical correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef] [PubMed]
 ) surviving COVID-19 patients, (
) surviving COVID-19 patients, ( ) non-surviving COVID-19 patients and (
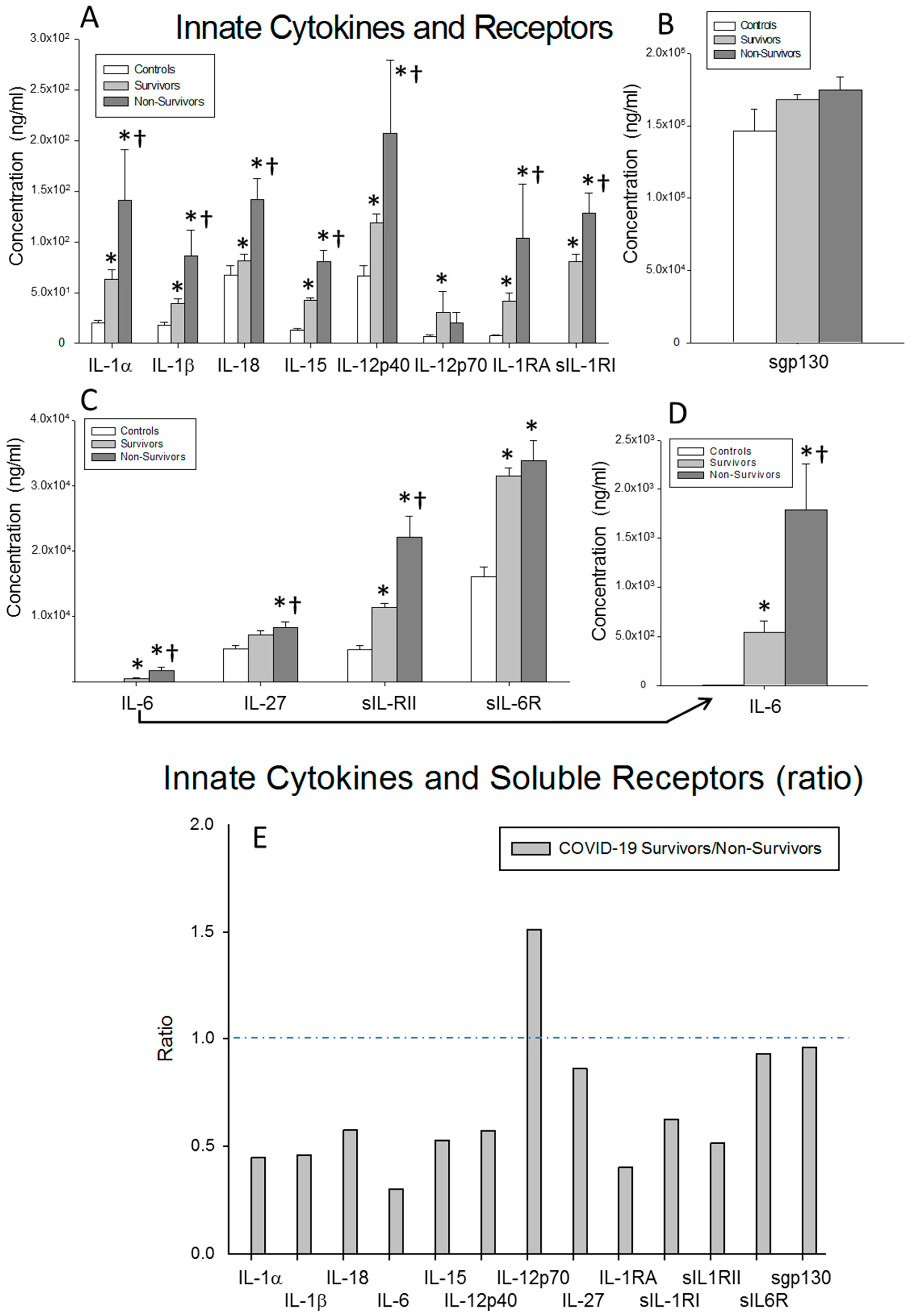
) non-surviving COVID-19 patients and ( ) healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (E) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (E) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (E) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
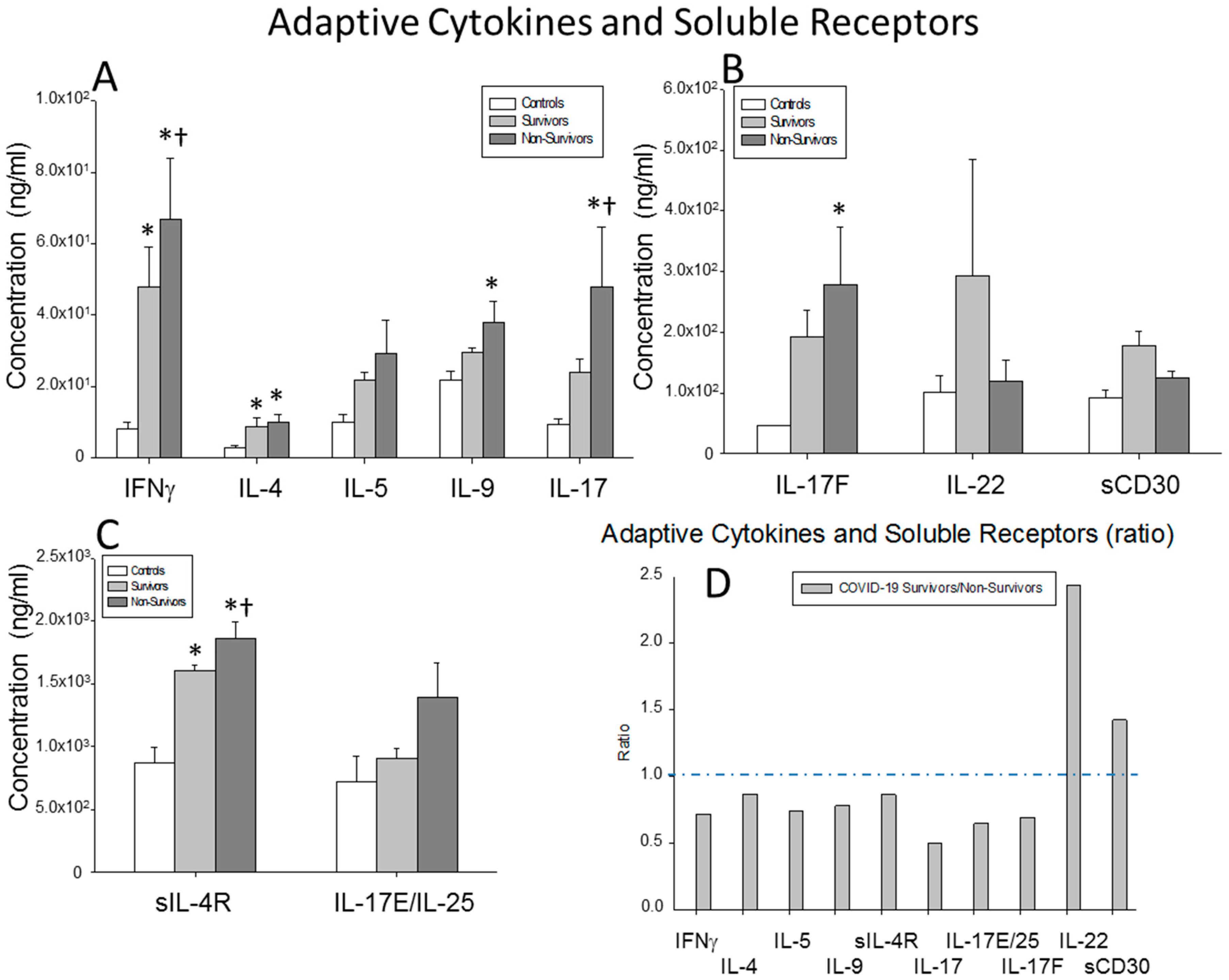
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (E) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each soluble factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each chemokine analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each chemokine analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each chemokine analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
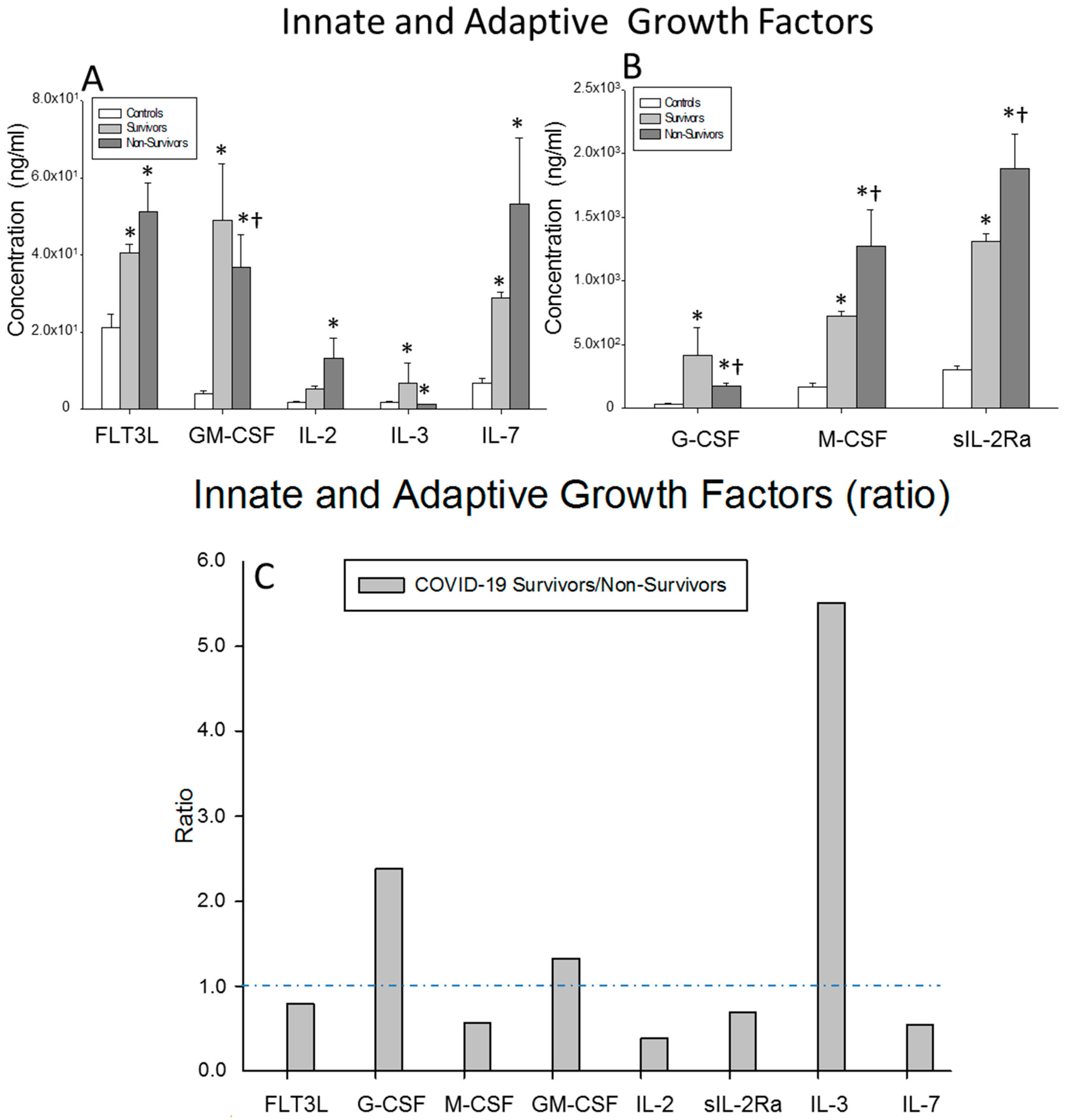
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each chemokine analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (C) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between circulating levels of surviving and non-surviving COVID-19 patients. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between circulating levels of surviving and non-surviving COVID-19 patients. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between circulating levels of surviving and non-surviving COVID-19 patients. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
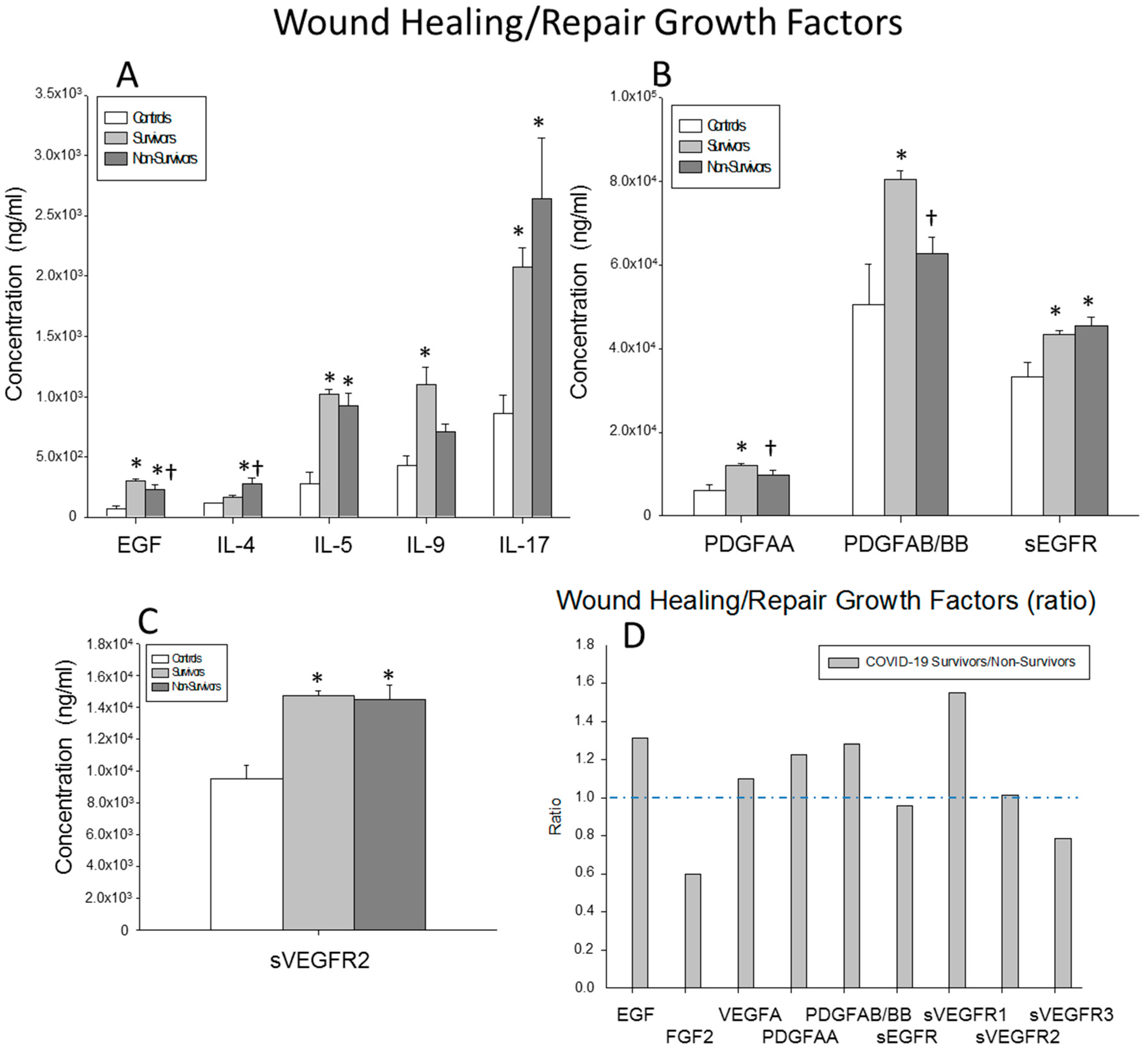
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between circulating levels of surviving and non-surviving COVID-19 patients. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each wound-healing and repair growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each wound-healing and repair growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each wound-healing and repair growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors.
) surviving COVID-19 patients, () non-surviving COVID-19 patients and () healthy controls. All the values are expressed as the means ± S.E.M.s *, p < 0.05 for surviving or non-surviving COVID-19 patients versus healthy controls. †, p < 0.05 for surviving versus non-surviving COVID-19 patients. (D) Data represent the ratios between the circulating levels in surviving and non-surviving COVID-19 patients of each wound-healing and repair growth factor analyzed. The dotted line represents a reference line for the ratio equal to 1 between survivors and non-survivors. ), are ordered from the best to worst predictive values obtained (ROC values) from the first 34 soluble factors studied.
), are ordered from the best to worst predictive values obtained (ROC values) from the first 34 soluble factors studied.
), are ordered from the best to worst predictive values obtained (ROC values) from the first 34 soluble factors studied.
), are ordered from the best to worst predictive values obtained (ROC values) from the first 34 soluble factors studied.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survivors (249) | Non-Survivors (37) | Total | p-Value | |
|---|---|---|---|---|
| Age ( years), mean (SD) | 62.95 (12.35) | 70.32 (12.19) | 63.90 (12.55) | <0.01 |
| Gender (male) | 63.86% | 72.97% | 65.03% | 0.27 |
| Oxygen saturation on Hospital admission | <0.01 | |||
| Extremely low (<80%) | 6.02% | 18.92% | 7.69% | |
| Low (80–89%) | 12.45% | 32.43% | 15.03% | |
| Medium (90–94%) | 57.83% | 48.65% | 56.64% | |
| Normal (>94%) | 23.69% | 0% | 20.63 | |
| No. Leucocytes, mean (SD), (n°/μL) | 8258.94 (4822.31) | 13,272.58 (11980.6) | 8910.26 (6429.82) | <0.01 |
| Ratio Lymphocytes/Leucocytes, mean (SD) | 0.18(0.11) | 0.12(0.15) | 0.17(0.12) | <0.01 |
| D-Dimer, mean (SD), (ng/mL) | 3296.63 (13,408.49) | 9500.24(11,480.64) | 4086.18 (13,324.76) | <0.01 |
| Ferritin, mean (SD), (ng/mL) | 787.89 (644.57) | 1405.95(1136.21) | 865.63 (751.05) | <0.01 |
| Charlson Index | 0.93 (1.40) | 1.72 (1.40) | 1.03 (1.43) | <0.01 |
| Elixhauser Index | 1.89 (1.69) | 2.91 (1.86) | 2.02 (1.74) | <0.01 |
| Commorbidities | ||||
| Acute Kidney failure | 8.43% | 27.03% | 10.84% | <0.01 |
| Dementia | 0.80% | 8.11% | 1.75% | <0.01 |
| 1. Proinflammatory and Anti-Inflammatory Cytokines of the Innate and Innate/Adaptive Immune Systems | |||||
| Innate | Criterion | Sensitivity | Specificity | Area under the ROC Curve | Significance Value (p) |
| IL-15 | ≤44.7 | 71.67 | 76.74 | 0.765 | 0.0001 |
| sIL-1RI | ≤129.2 | 90.24 | 62.5 | 0.764 | 0.0112 |
| sIL-1RII | ≤12544 | 75.77 | 65.12 | 0.733 | 0.0001 |
| IL-6 | ≤158.73 | 74.74 | 67.44 | 0.726 | 0.0001 |
| IL-18 | ≤68.02 | 61.38 | 79.07 | 0.708 | 0.0001 |
| IL-27 | ≤5701 | 58.16 | 69.77 | 0.649 | 0.0019 |
| IL-1β | ≤42.24 | 81.44 | 41.86 | 0.634 | 0.0055 |
| IL-1RA | ≤14.7 | 55.94 | 67.44 | 0.625 | 0.0095 |
| IL-12p70 | ≤4.94 | 42.17 | 81.82 | 0.609 | 0.048 |
| Innate/adaptive | |||||
| IL-10 | ≤19.54 | 53.28 | 86.05 | 0.726 | 0.0001 |
| sTNF-RII | ≤11487 | 58.7 | 74.42 | 0.696 | 0.0001 |
| sRAGE | ≤99.58 | 66.8 | 63.41 | 0.673 | 0.0004 |
| sTNF-RI | ≤2476 | 82.07 | 48.84 | 0.661 | 0.0008 |
| TNFα | ≤102.75 | 67.69 | 58.14 | 0.651 | 0.0016 |
| sCD40L | >3781 | 53.58 | 76.74 | 0.646 | 0.0004 |
| TNFβ | ≤21.52 | 66.78 | 55.81 | 0.611 | 0.0214 |
| TGFα | ≤20.29 | 53.42 | 69.77 | 0.602 | 0.0347 |
| 2. Chemokines | |||||
| MCP3 | ≤71.67 | 68.26 | 76.74 | 0.769 | 0.0001 |
| IL-8 | ≤77.12 | 70.99 | 74.42 | 0.737 | 0.0001 |
| MIG | ≤6689 | 47.96 | 86.05 | 0.707 | 0.0001 |
| IP-10 | ≤433.19 | 48.81 | 83.72 | 0.691 | 0.0001 |
| MDC | >628 | 76.19 | 48.84 | 0.647 | 0.0004 |
| MCP1 | ≤2752 | 78.23 | 53.49 | 0.637 | 0.0044 |
| RANTES | >3371 | 94.88 | 32.56 | 0.605 | 0.0153 |
| 3. Innate and Adaptive Growth Factors | |||||
| G-CSF | ≤58.68 | 54.47 | 89.74 | 0.715 | 0.0001 |
| GM-CSF | ≤9.81 | 45.1 | 100 | 0.694 | 0.0502 |
| M-CSF | ≤927.52 | 72.4 | 53.66 | 0.655 | 0.0015 |
| sIL-2Ra | ≤979.22 | 44.86 | 79.07 | 0.646 | 0.0023 |
| 4. Adaptive Cytokines | |||||
| IFNγ | ≤25.15 | 70.98 | 52.38 | 0.611 | 0.023 |
| IL-17A | ≤10.42 | 48.24 | 74.42 | 0.603 | 0.0327 |
| 5. Wound-Healing/Repair Growth Factors | |||||
| PDGFABBB | >63788 | 64.29 | 65.12 | 0.664 | 0.0001 |
| FGF2 | ≤112.23 | 45.55 | 83.72 | 0.661 | 0.0007 |
| EGF | >169.49 | 65.64 | 65.12 | 0.642 | 0.0006 |
| PDGFAA | >8732 | 65.99 | 58.14 | 0.626 | 0.0029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monserrat, J.; Gómez-Lahoz, A.; Ortega, M.A.; Sanz, J.; Muñoz, B.; Arévalo-Serrano, J.; Rodríguez, J.M.; Gasalla, J.M.; Gasulla, Ó.; Arranz, A.; et al. Role of Innate and Adaptive Cytokines in the Survival of COVID-19 Patients. Int. J. Mol. Sci. 2022, 23, 10344. https://doi.org/10.3390/ijms231810344
Monserrat J, Gómez-Lahoz A, Ortega MA, Sanz J, Muñoz B, Arévalo-Serrano J, Rodríguez JM, Gasalla JM, Gasulla Ó, Arranz A, et al. Role of Innate and Adaptive Cytokines in the Survival of COVID-19 Patients. International Journal of Molecular Sciences. 2022; 23(18):10344. https://doi.org/10.3390/ijms231810344
Chicago/Turabian StyleMonserrat, Jorge, Ana Gómez-Lahoz, Miguel A. Ortega, José Sanz, Benjamin Muñoz, Juan Arévalo-Serrano, José Miguel Rodríguez, Jose Maria Gasalla, Óscar Gasulla, Alberto Arranz, and et al. 2022. "Role of Innate and Adaptive Cytokines in the Survival of COVID-19 Patients" International Journal of Molecular Sciences 23, no. 18: 10344. https://doi.org/10.3390/ijms231810344
APA StyleMonserrat, J., Gómez-Lahoz, A., Ortega, M. A., Sanz, J., Muñoz, B., Arévalo-Serrano, J., Rodríguez, J. M., Gasalla, J. M., Gasulla, Ó., Arranz, A., Fortuny-Profitós, J., Mazaira-Font, F. A., Teixidó Román, M., Martínez-A, C., Balomenos, D., Asunsolo, A., Álvarez-Mon, M., & on behalf of the COVID-19 HUPA Group. (2022). Role of Innate and Adaptive Cytokines in the Survival of COVID-19 Patients. International Journal of Molecular Sciences, 23(18), 10344. https://doi.org/10.3390/ijms231810344