
Scattered Tubular Cells Markers in Macula Densa of Normal Human Adult Kidney

 ,
,  , , ,
, , , 
Abstract
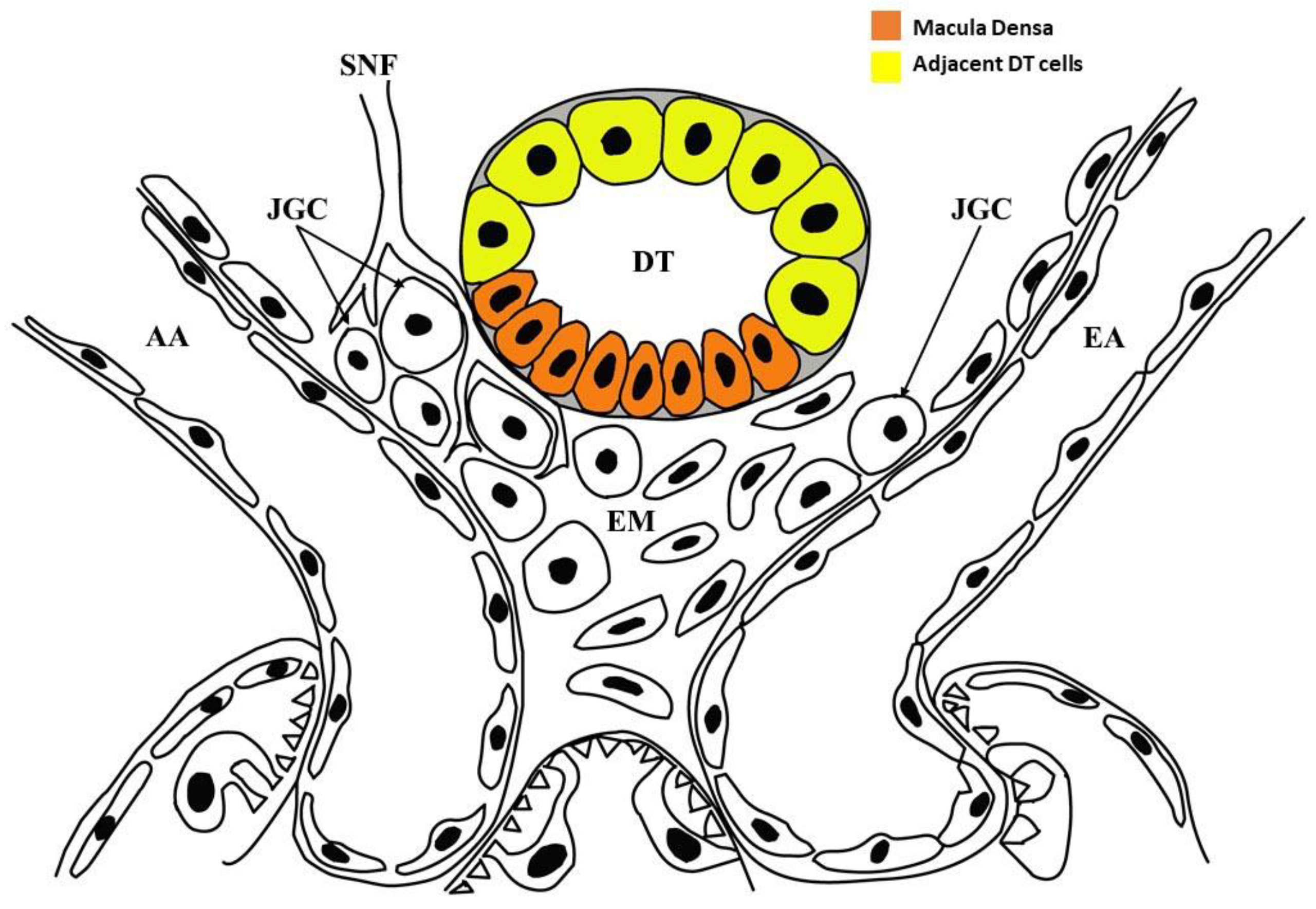
:1. Introduction
2. Results
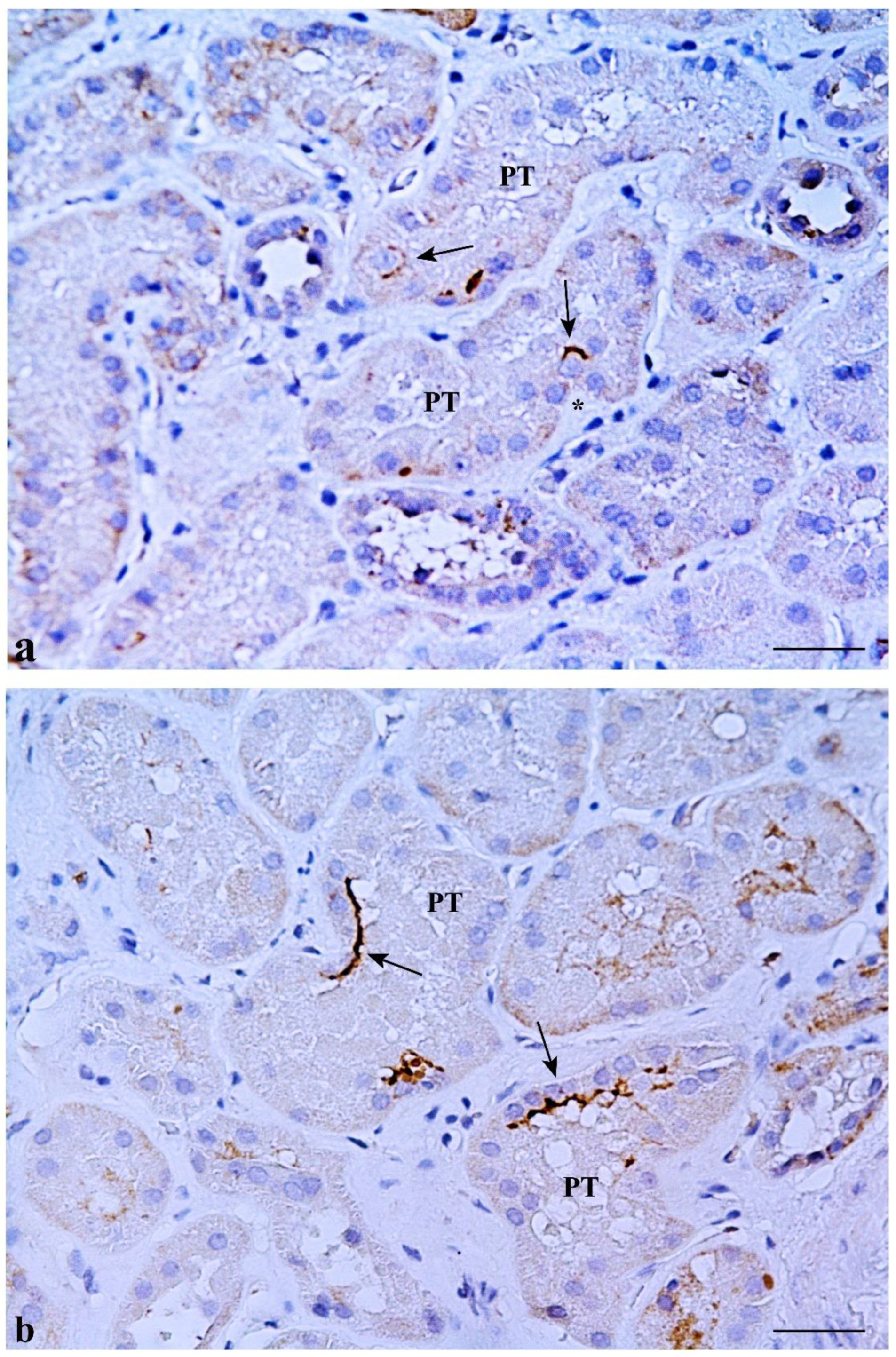
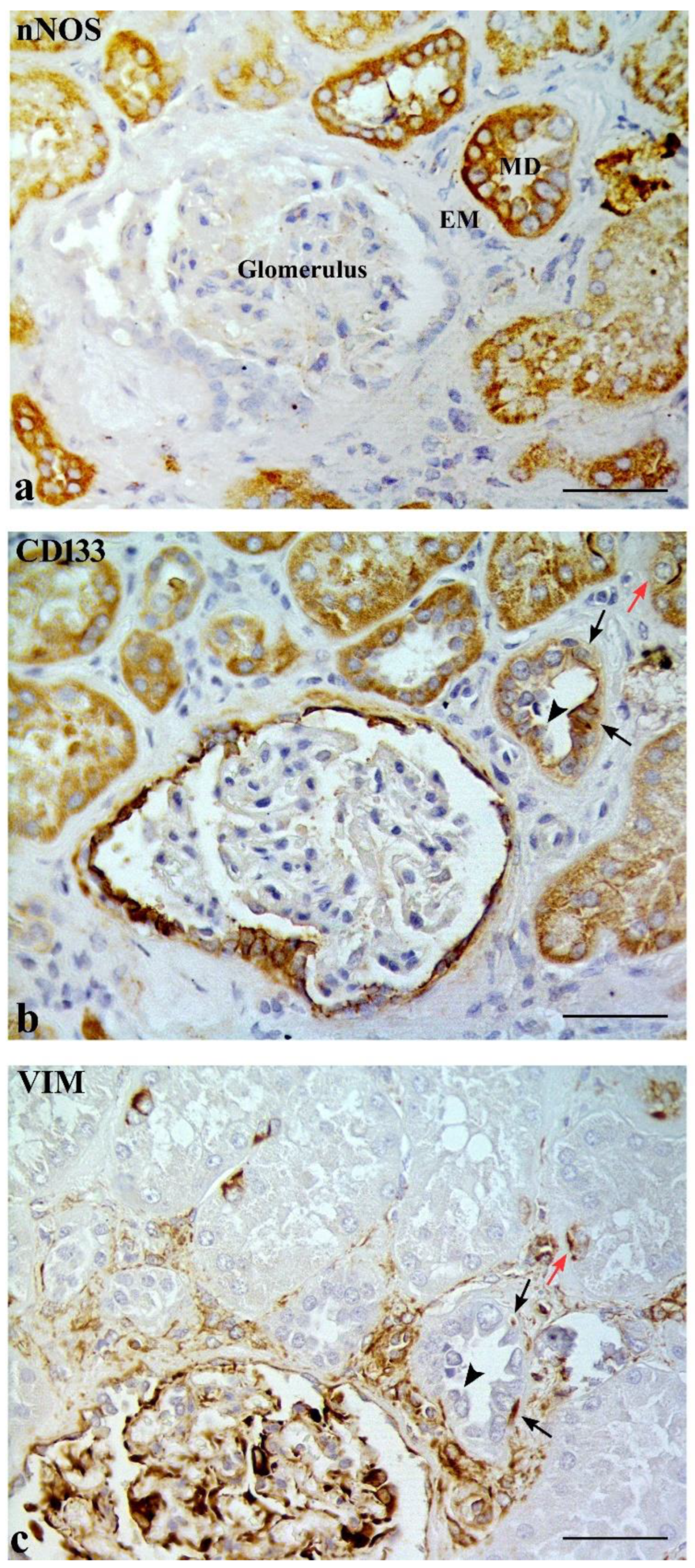
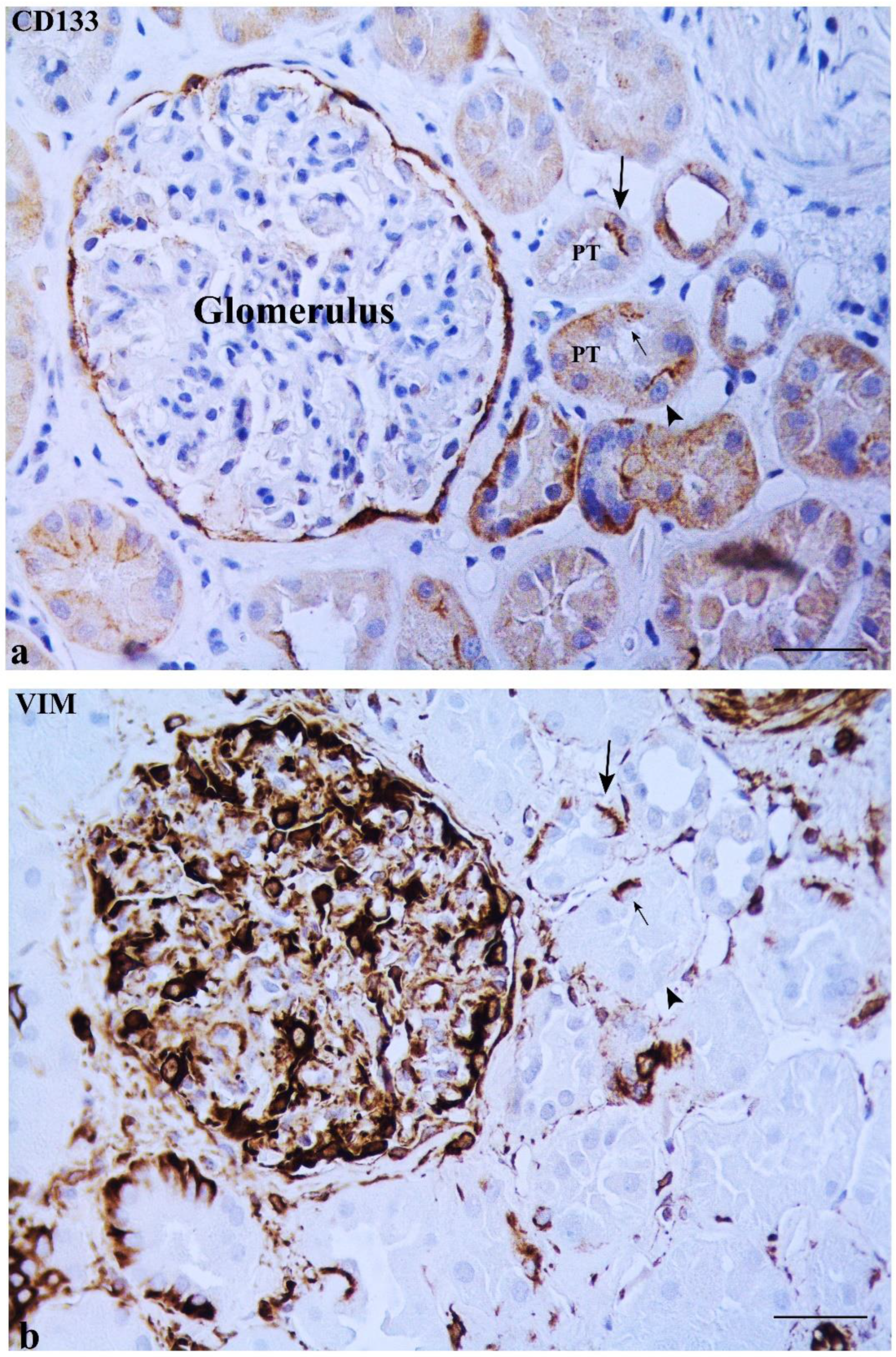
2.1. CD133 and KRT7 Expression
2.2. Some DT Cells Adjacent to MD Co-Express CD133 and VIM
3. Discussion
4. Materials and Methods
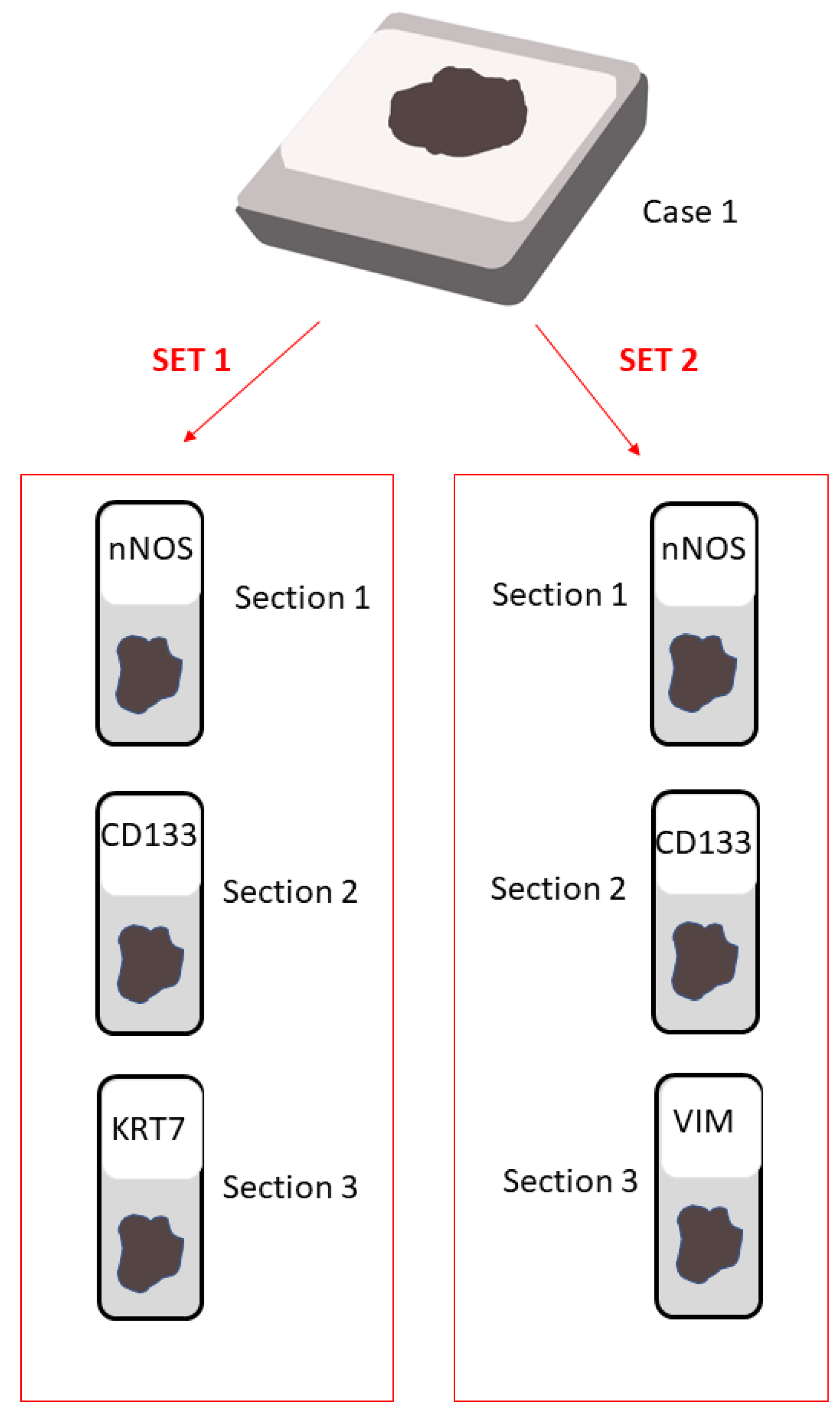
4.1. Paraffin Samples
4.2. Immunohistochemical Analysis
4.3. Transmission Electron Microscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castrop, H. Mediators of tubuloglomerular feedback regulation of glomerular filtration: ATP and adenosine. Acta Physiol. 2007, 189, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Komlosi, P.; Fintha, A.; Bell, P.D. Current mechanisms of macula densa cell signalling. Acta Physiol. Scand. 2004, 181, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Navar, L.G.; Inscho, E.W.; Majid, S.A.; Imig, J.D.; Harrison-Bernard, L.M.; Mitchell, K.D. Paracrine regulation of the renal microcirculation. Physiol. Rev. 1996, 76, 425–536. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Muhlbauer, B.; Osswald, H. Adenosine and kidney function. Physiol. Rev. 2006, 86, 901–940. [Google Scholar] [CrossRef]
- Guyton, A.C.; Langston, J.B.; Navar, G. Theory for Renal Autoregulation by Feedback at the Juxtaglomerular Apparatus. Circ. Res. 1964, 15, 187–197. [Google Scholar]
- Thurau, K.; Schnermann, J. The Na concentration of the macula densa cells as a factor regulating glomerular filtration rate (micropuncture studies). 1965. J. Am. Soc. Nephrol. 1998, 9, 925–934. [Google Scholar] [CrossRef]
- Barajas, L.; Latta, H. A three-dimensional study of the juxtaglomerular apparatus in the rat. Light and electron microscopic observations. Lab. Investig. 1963, 12, 257–269. [Google Scholar]
- Barajas, L. Anatomy of the juxtaglomerular apparatus. Am. J. Physiol 1979, 237, F333–F343. [Google Scholar] [CrossRef]
- Latta, H.; Maunsbach, A.B. The juxtaglomerular apparatus as studied electron microscopically. J. Ultrastruct. Res. 1962, 6, 547–561. [Google Scholar] [CrossRef]
- Sikri, K.L.; Foster, C.L. Light and electron microscopical observations on the macula densa of the Syrian hamster kidney. J. Anat. 1981, 132, 57–69. [Google Scholar]
- Sottiurai, V.; Malvin, R.L. The demonstratin of cilia in canine macula densa cells. Am. J. Anat. 1972, 135, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Tisher, C.C.; Bulger, R.E.; Trump, B.F. Human renal ultrastructure. 3. The distal tubule in healthy individuals. Lab. Investig. 1968, 18, 655–668. [Google Scholar] [PubMed]
- Cangiotti, A.M.; Lorenzi, T.; Zingaretti, M.C.; Fabri, M.; Morroni, M. Polarized Ends of Human Macula Densa Cells: Ultrastructural Investigation and Morphofunctional Correlations. Anat. Rec. 2018, 301, 922–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzi, T.; Graciotti, L.; Sagrati, A.; Reguzzoni, M.; Protasoni, M.; Minardi, D.; Milanese, G.; Cremona, O.; Fabri, M.; Morroni, M. Normal human macula densa morphology and cell turnover: A histological, ultrastructural, and immunohistochemical investigation. Anat. Rec. 2020, 303, 2904–2916. [Google Scholar] [CrossRef] [PubMed]
- Peti-Peterdi, J.; Harris, R.C. Macula densa sensing and signaling mechanisms of renin release. J. Am. Soc. Nephrol. 2010, 21, 1093–1096. [Google Scholar] [CrossRef]
- Peti-Peterdi, J. Newly stemming functions of macula densa-derived prostanoids. Hypertension 2015, 65, 987–988. [Google Scholar] [CrossRef]
- Lindgren, D.; Bostrom, A.K.; Nilsson, K.; Hansson, J.; Sjolund, J.; Moller, C.; Jirstrom, K.; Nilsson, E.; Landberg, G.; Axelson, H.; et al. Isolation and characterization of progenitor-like cells from human renal proximal tubules. Am. J. Pathol. 2011, 178, 828–837. [Google Scholar] [CrossRef]
- Romagnani, P.; Lasagni, L.; Remuzzi, G. Renal progenitors: An evolutionary conserved strategy for kidney regeneration. Nat. Rev. Nephrol. 2013, 9, 137–146. [Google Scholar] [CrossRef]
- Romagnani, P.; Remuzzi, G. CD133+ renal stem cells always co-express CD24 in adult human kidney tissue. Stem Cell Res. 2014, 12, 828–829. [Google Scholar] [CrossRef]
- Sallustio, F.; De Benedictis, L.; Castellano, G.; Zaza, G.; Loverre, A.; Costantino, V.; Grandaliano, G.; Schena, F.P. TLR2 plays a role in the activation of human resident renal stem/progenitor cells. FASEB J. 2010, 24, 514–525. [Google Scholar] [CrossRef]
- Challen, G.A.; Bertoncello, I.; Deane, J.A.; Ricardo, S.D.; Little, M.H. Kidney side population reveals multilineage potential and renal functional capacity but also cellular heterogeneity. J. Am. Soc. Nephrol. 2006, 17, 1896–1912. [Google Scholar] [CrossRef]
- Kitamura, S.; Yamasaki, Y.; Kinomura, M.; Sugaya, T.; Sugiyama, H.; Maeshima, Y.; Makino, H. Establishment and characterization of renal progenitor like cells from S3 segment of nephron in rat adult kidney. FASEB J. 2005, 19, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, A.; Yamashita, S.; Nojima, Y. Identification of renal progenitor-like tubular cells that participate in the regeneration processes of the kidney. J. Am. Soc. Nephrol. 2003, 14, 3138–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, J.; Hultenby, K.; Cramnert, C.; Ponten, F.; Jansson, H.; Lindgren, D.; Axelson, H.; Johansson, M.E. Evidence for a morphologically distinct and functionally robust cell type in the proximal tubules of human kidney. Hum. Pathol. 2014, 45, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Smeets, B.; Boor, P.; Dijkman, H.; Sharma, S.V.; Jirak, P.; Mooren, F.; Berger, K.; Bornemann, J.; Gelman, I.H.; Floege, J.; et al. Proximal tubular cells contain a phenotypically distinct, scattered cell population involved in tubular regeneration. J. Pathol. 2013, 229, 645–659. [Google Scholar] [CrossRef]
- Sagrinati, C.; Netti, G.S.; Mazzinghi, B.; Lazzeri, E.; Liotta, F.; Frosali, F.; Ronconi, E.; Meini, C.; Gacci, M.; Squecco, R.; et al. Isolation and characterization of multipotent progenitor cells from the Bowman’s capsule of adult human kidneys. J. Am. Soc. Nephrol. 2006, 17, 2443–2456. [Google Scholar] [CrossRef]
- Bussolati, B.; Bruno, S.; Grange, C.; Buttiglieri, S.; Deregibus, M.C.; Cantino, D.; Camussi, G. Isolation of renal progenitor cells from adult human kidney. Am. J. Pathol. 2005, 166, 545–555. [Google Scholar] [CrossRef]
- Eymael, J.; Smeets, B. Origin and fate of the regenerating cells of the kidney. Eur. J. Pharmacol. 2016, 790, 62–73. [Google Scholar] [CrossRef]
- Stamellou, E.; Leuchtle, K.; Moeller, M.J. Regenerating tubular epithelial cells of the kidney. Nephrol. Dial. Transplant. 2021, 36, 1968–1975. [Google Scholar] [CrossRef]
- Razga, Z.; Nyengaard, J.R. The effect of angiotensin II on the number of macula densa cells through the AT1 receptor. Nephron Physiol. 2009, 112, p37–p43. [Google Scholar] [CrossRef]
- Razga, Z. Functional Relevancies of Trans-Differentiation in the Juxtaglomerular Apparatus of Rat Kidney. Int. J. Nephrol. Renov. Dis. 2020, 13, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Lieberthal, W.; Nigam, S.K. Acute renal failure. I. Relative importance of proximal vs. distal tubular injury. Am. J. Physiol. 1998, 275, F623–F631. [Google Scholar] [CrossRef] [PubMed]
- Sekine, M.; Monkawa, T.; Morizane, R.; Matsuoka, K.; Taya, C.; Akita, Y.; Joh, K.; Itoh, H.; Hayashi, M.; Kikkawa, Y.; et al. Selective depletion of mouse kidney proximal straight tubule cells causes acute kidney injury. Transgenic Res. 2012, 21, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, L.; Hiatt, M.J.; Yoder, M.C.; Tarantal, A.F.; Matsell, D.G. Ontogeny of CD24 in the human kidney. Kidney Int. 2010, 77, 1123–1131. [Google Scholar] [CrossRef]
- Polari, L.; Alam, C.M.; Nystrom, J.H.; Heikkila, T.; Tayyab, M.; Baghestani, S.; Toivola, D.M. Keratin intermediate filaments in the colon: Guardians of epithelial homeostasis. Int. J. Biochem. Cell Biol. 2020, 129, 105878. [Google Scholar] [CrossRef]
- Toivola, D.M.; Strnad, P.; Habtezion, A.; Omary, M.B. Intermediate filaments take the heat as stress proteins. Trends Cell Biol. 2010, 20, 79–91. [Google Scholar] [CrossRef]
- Djudjaj, S.; Papasotiriou, M.; Bulow, R.D.; Wagnerova, A.; Lindenmeyer, M.T.; Cohen, C.D.; Strnad, P.; Goumenos, D.S.; Floege, J.; Boor, P. Keratins are novel markers of renal epithelial cell injury. Kidney Int. 2016, 89, 792–808. [Google Scholar] [CrossRef]
- Ivaska, J.; Pallari, H.M.; Nevo, J.; Eriksson, J.E. Novel functions of vimentin in cell adhesion, migration, and signaling. Exp. Cell Res. 2007, 313, 2050–2062. [Google Scholar] [CrossRef]
- Witzgall, R.; Brown, D.; Schwarz, C.; Bonventre, J.V. Localization of proliferating cell nuclear antigen, vimentin, c-Fos, and clusterin in the postischemic kidney. Evidence for a heterogenous genetic response among nephron segments, and a large pool of mitotically active and dedifferentiated cells. J. Clin. Investig. 1994, 93, 2175–2188. [Google Scholar] [CrossRef]
- Mundel, P.; Bachmann, S.; Bader, M.; Fischer, A.; Kummer, W.; Mayer, B.; Kriz, W. Expression of nitric oxide synthase in kidney macula densa cells. Kidney Int. 1992, 42, 1017–1019. [Google Scholar] [CrossRef]
- Altobelli, E.; Latella, G.; Morroni, M.; Licini, C.; Tossetta, G.; Mazzucchelli, R.; Profeta, V.F.; Coletti, G.; Leocata, P.; Castellucci, M.; et al. Low HtrA1 expression in patients with longstanding ulcerative colitis and colorectal cancer. Oncol. Rep. 2017, 38, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Marinelli Busilacchi, E.; Costantini, A.; Mancini, G.; Tossetta, G.; Olivieri, J.; Poloni, A.; Viola, N.; Butini, L.; Campanati, A.; Goteri, G.; et al. Nilotinib Treatment of Patients Affected by Chronic Graft-versus-Host Disease Reduces Collagen Production and Skin Fibrosis by Downmodulating the TGF-beta and p-SMAD Pathway. Biol. Blood Marrow Transpl. 2020, 26, 823–834. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases | Gender | Age | Tumor Histopathology |
|---|---|---|---|
| Case 1 | Female | 78 | Clear cell carcinoma |
| Case 2 | Male | 51 | Clear cell carcinoma |
| Case 3 | Male | 85 | Clear cell carcinoma |
| Case 4 | Female | 57 | Oncocytoma |
| Case 5 | Male | 47 | Clear cell carcinoma |
| Case 6 | Female | 37 | Chromophobe renal cell carcinoma |
| Antibodies | Dilution | Company |
|---|---|---|
| Rabbit polyclonal nNOS (#160870) | 1:500 | Cayman Chem., Ann Arbor, MI, USA |
| Rabbit polyclonal CD133 (#HPA004922) | 1:650 | Sigma, St. Louis, MO, USA |
| Mouse monoclonal Cytokeratin 7 (#M7018) | 1:50 | Dako, Glostrup, DM, USA |
| Mouse monoclonal Vimentin (#GA630) | 1:1 | Dako, Glostrup, DM, USA |
| Isotype mouse IgG (#I-2000-1) | 1:50 | Vector lab., Burlingame, CA, USA |
| Isotype rabbit IgG (#I-1000-5) | 1:500 or 1:650 | Vector lab., Burlingame, CA, USA |
| Secondary goat anti-rabbit antibody (#BA-1000-1.5) | 1:200 | Vector lab., Burlingame, CA, USA |
| Secondary goat anti-mouse antibody (#BA-9200-1.5) | 1:200 | Vector lab., Burlingame, CA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tossetta, G.; Fantone, S.; Lorenzi, T.; Galosi, A.B.; Sagrati, A.; Fabri, M.; Marzioni, D.; Morroni, M. Scattered Tubular Cells Markers in Macula Densa of Normal Human Adult Kidney. Int. J. Mol. Sci. 2022, 23, 10504. https://doi.org/10.3390/ijms231810504
Tossetta G, Fantone S, Lorenzi T, Galosi AB, Sagrati A, Fabri M, Marzioni D, Morroni M. Scattered Tubular Cells Markers in Macula Densa of Normal Human Adult Kidney. International Journal of Molecular Sciences. 2022; 23(18):10504. https://doi.org/10.3390/ijms231810504
Chicago/Turabian StyleTossetta, Giovanni, Sonia Fantone, Teresa Lorenzi, Andrea Benedetto Galosi, Andrea Sagrati, Mara Fabri, Daniela Marzioni, and Manrico Morroni. 2022. "Scattered Tubular Cells Markers in Macula Densa of Normal Human Adult Kidney" International Journal of Molecular Sciences 23, no. 18: 10504. https://doi.org/10.3390/ijms231810504
APA StyleTossetta, G., Fantone, S., Lorenzi, T., Galosi, A. B., Sagrati, A., Fabri, M., Marzioni, D., & Morroni, M. (2022). Scattered Tubular Cells Markers in Macula Densa of Normal Human Adult Kidney. International Journal of Molecular Sciences, 23(18), 10504. https://doi.org/10.3390/ijms231810504