
Impact of Leptin on the Expression Profile of Macrophages during Mechanical Strain In Vitro


 and
and
Abstract
:1. Introduction
2. Results
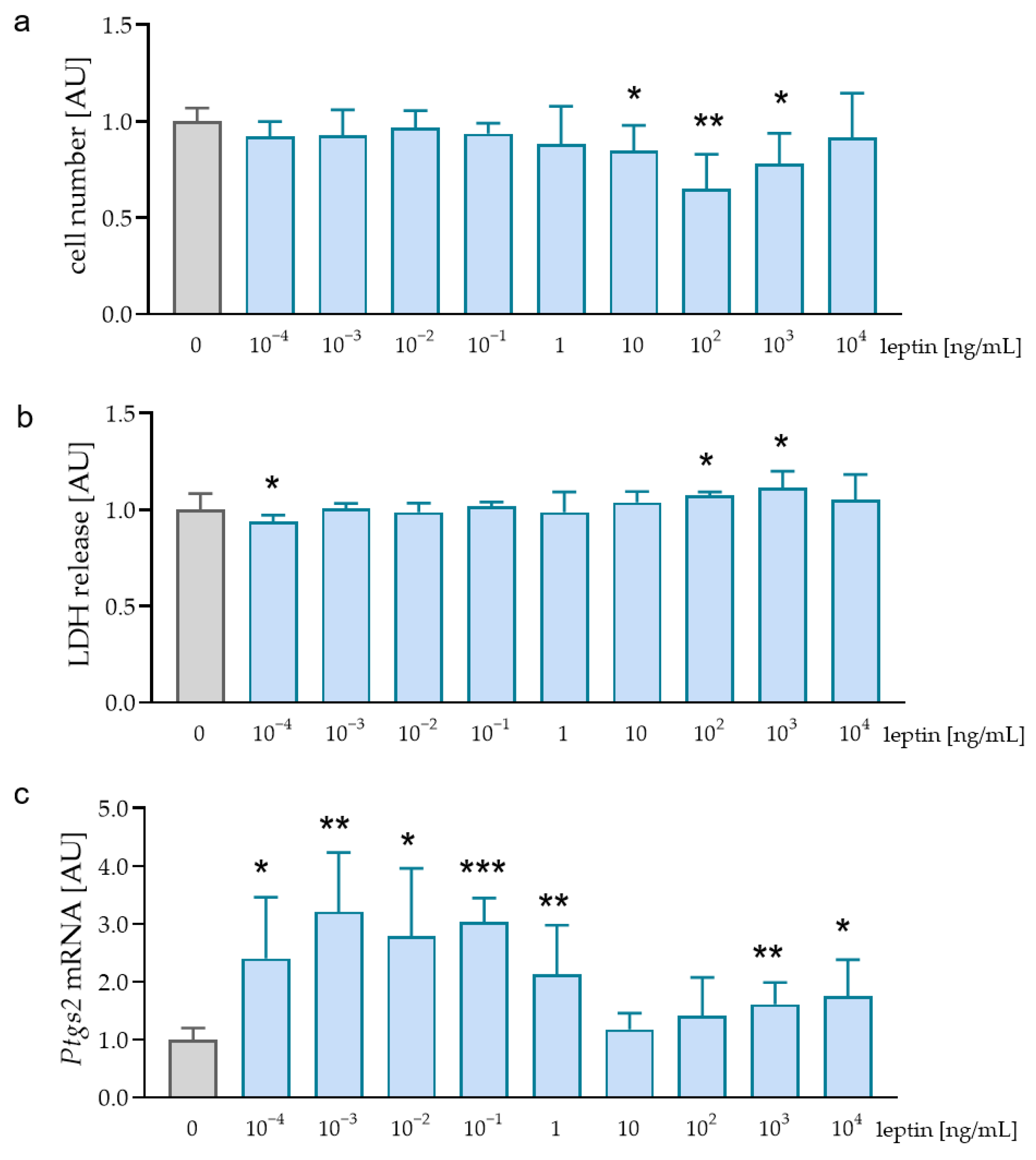
2.1. Effects of Different Leptin Concentrations without Mechanical Strain
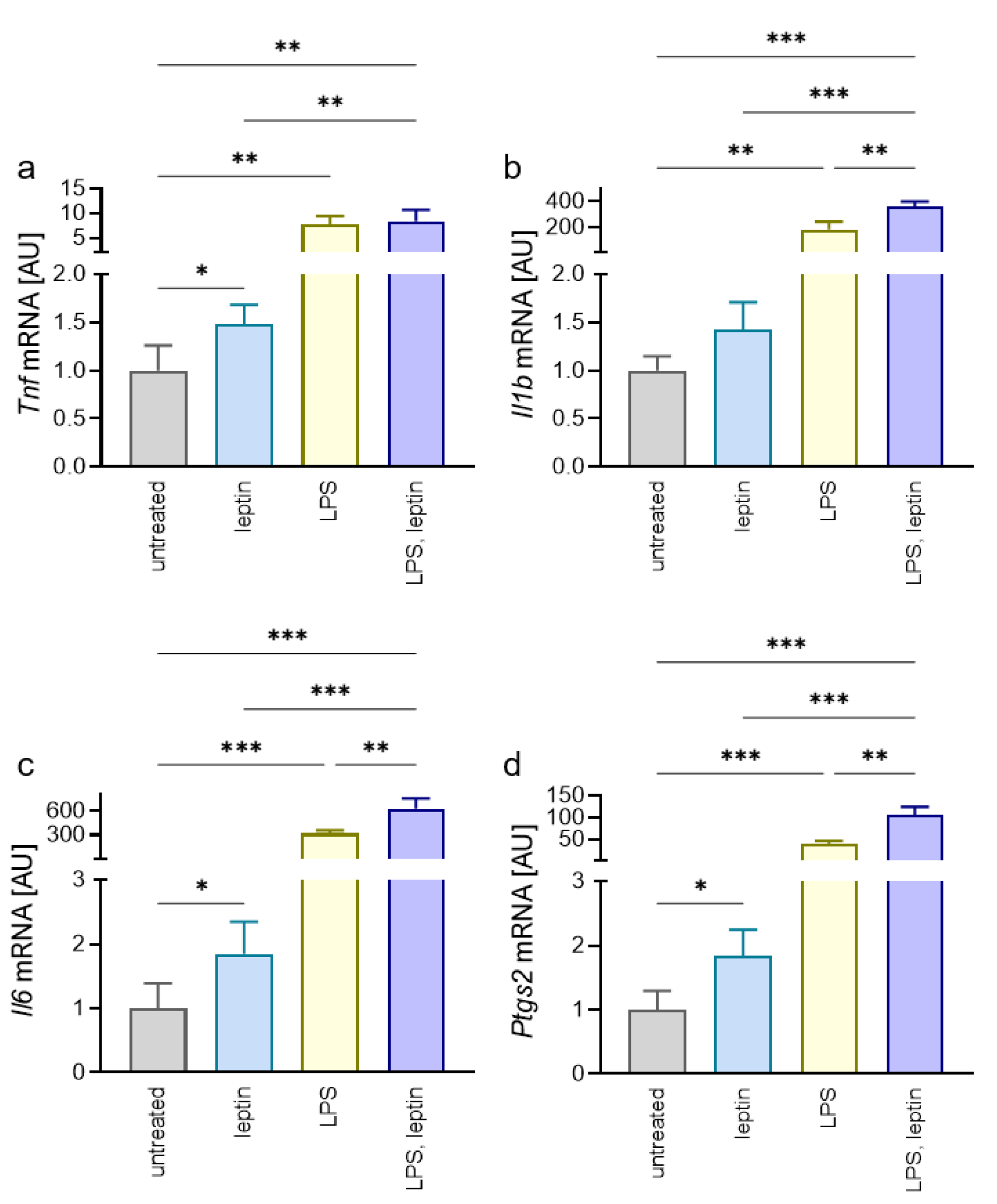
2.2. Impact of Leptin on Macrophages in Combination with Porphyromonas Gingivalis LPS
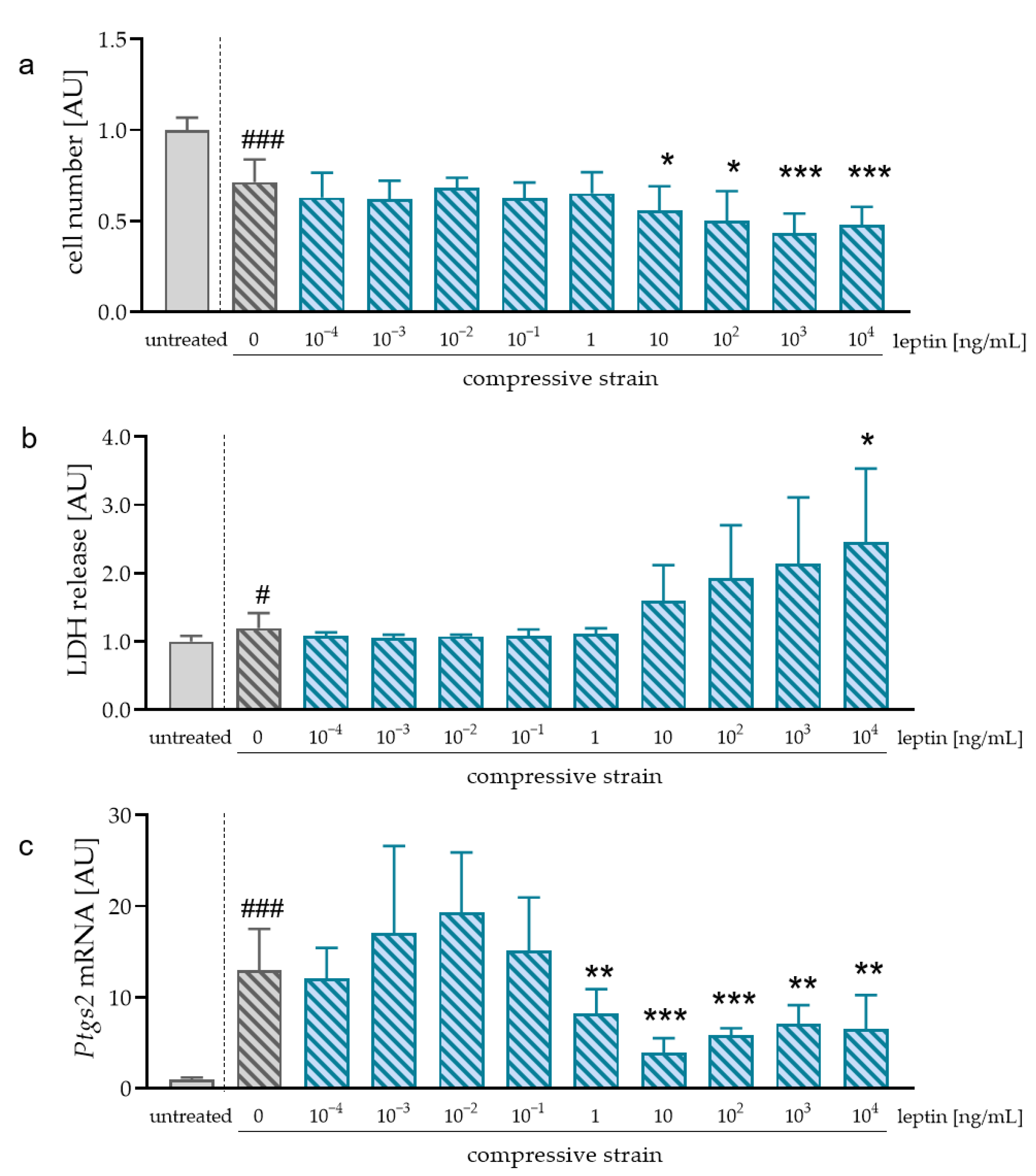
2.3. Impact of Different Leptin Concentrations during Compressive Strain
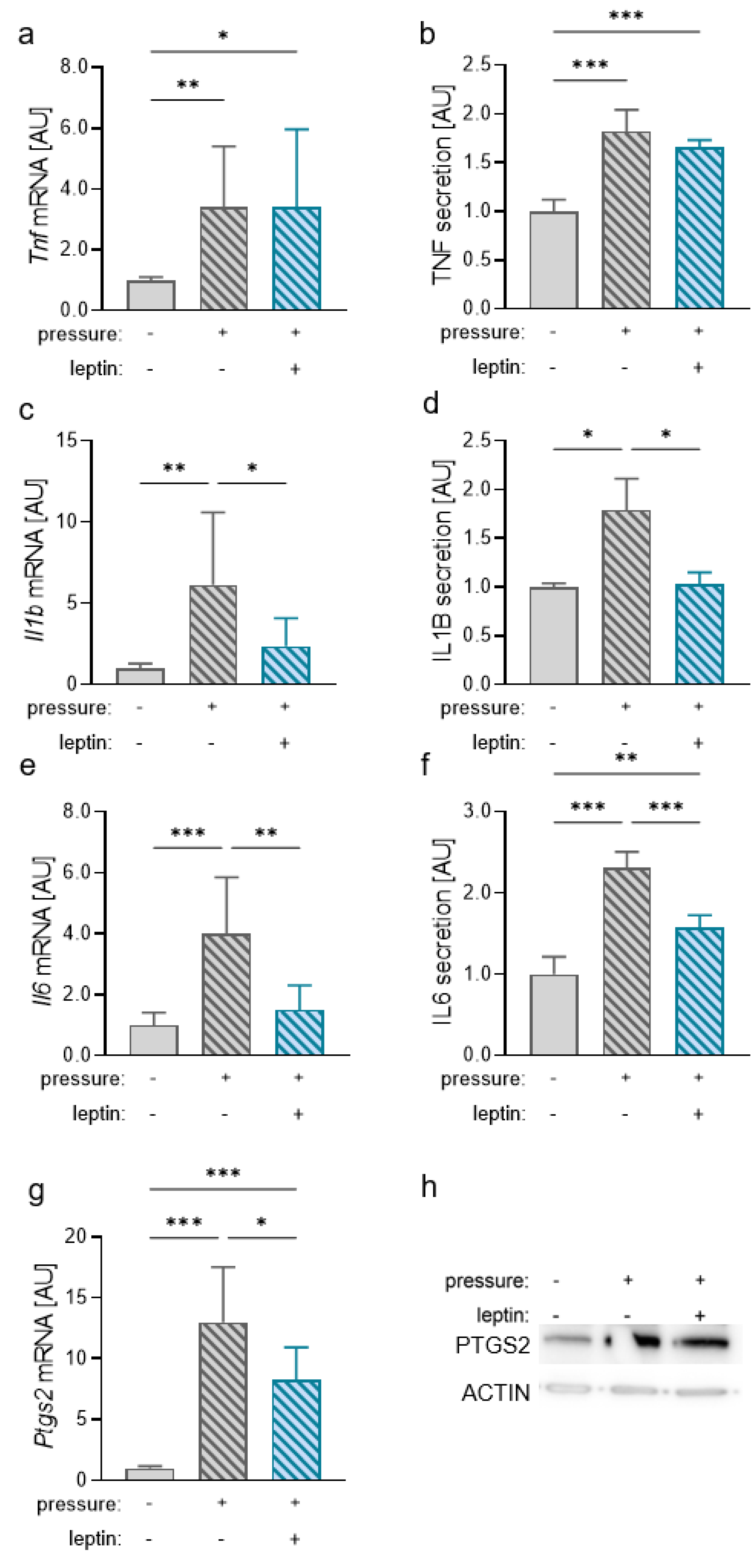
2.4. Effects of Leptin on the Expression of Proinflammatory Mediators during Compressive Strain
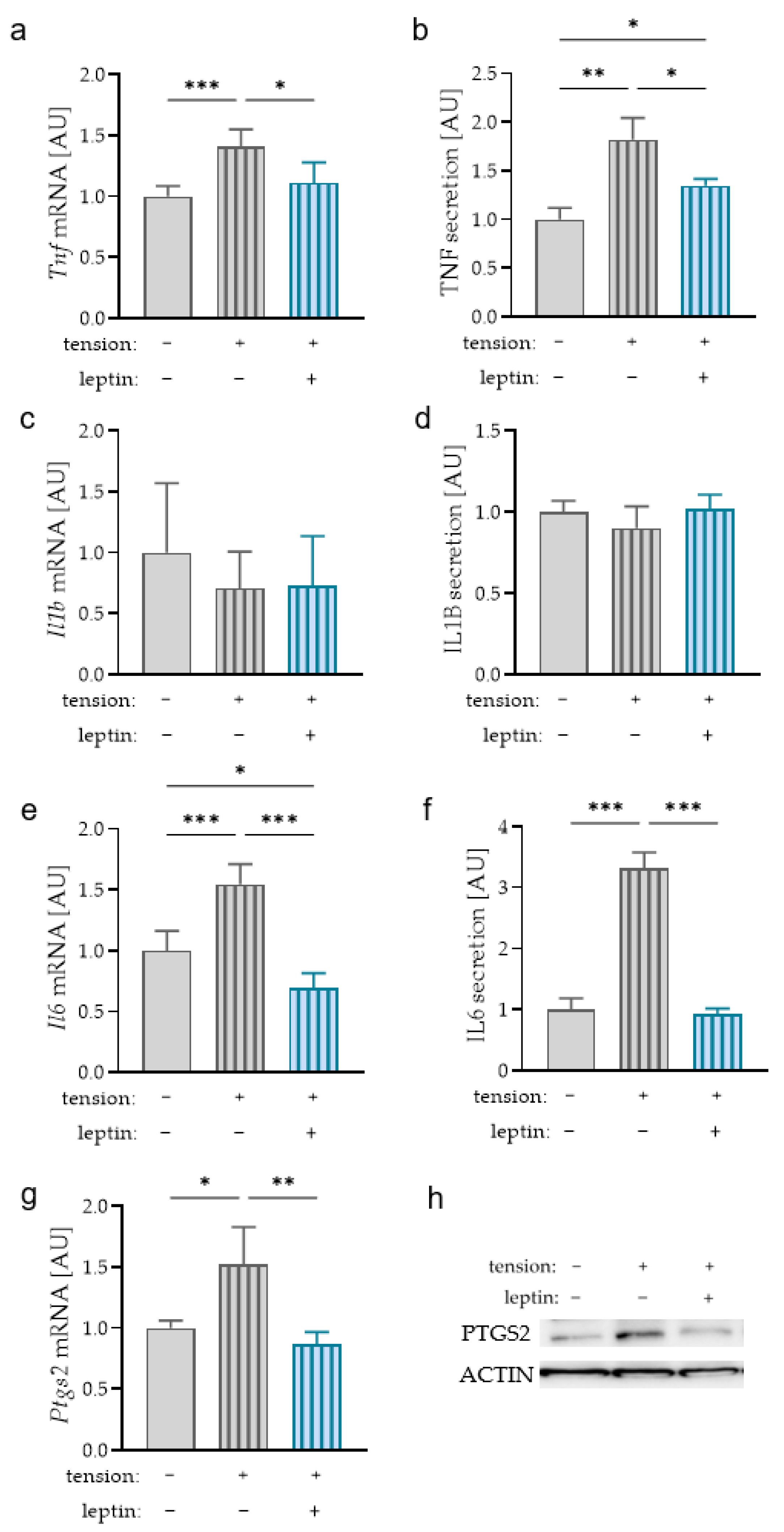
2.5. Impact of Leptin on the Expression of Proinflammatory Mediators during Tensile Strain
3. Discussion
4. Materials and Methods
4.1. Cell Culture Experiments
4.2. Determination of Cell Number
4.3. Determination of Cytotoxicity by Lactate Dehydrogenase (LDH) Assay
4.4. RNA Isolation and cDNA Synthesis
4.5. RT-qPCR Analysis
4.6. Western Blot Analysis
4.7. Enzyme-Linked Immunosorbent Assays (ELISA)
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weihrauch-Blüher, S.; Wiegand, S. Risk Factors and Implications of Childhood Obesity. Curr. Obes. Rep. 2018, 7, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Fuster, J.J.; Walsh, K. Adipokines: A link between obesity and cardiovascular disease. J. Cardiol. 2014, 63, 250–259. [Google Scholar] [CrossRef]
- Nishida, N.; Tanaka, M.; Hayashi, N.; Nagata, H.; Takeshita, T.; Nakayama, K.; Morimoto, K.; Shizukuishi, S. Determination of smoking and obesity as periodontitis risks using the classification and regression tree method. J. Periodontol. 2005, 76, 923–928. [Google Scholar] [CrossRef]
- Münzberg, H.; Morrison, C.D. Structure, production and signaling of leptin. Metabolism 2015, 64, 13–23. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef]
- Morioka, T.; Emoto, M.; Yamazaki, Y.; Kurajoh, M.; Motoyama, K.; Mori, K.; Fukumoto, S.; Shioi, A.; Shoji, T.; Inaba, M. Plasma soluble leptin receptor levels are associated with pancreatic β-cell dysfunction in patients with type 2 diabetes. J. Diabetes Investig. 2018, 9, 55–62. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, Obesity, and Leptin Resistance: Where Are We 25 Years Later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef]
- Zhang, Y.; Chua, S. Leptin Function and Regulation. Compr. Physiol. 2017, 8, 351–369. [Google Scholar] [CrossRef]
- Gainsford, T.; Willson, T.A.; Metcalf, D.; Handman, E.; McFarlane, C.; Ng, A.; Nicola, N.A.; Alexander, W.S.; Hilton, D.J. Leptin can induce proliferation, differentiation, and functional activation of hemopoietic cells. Proc. Natl. Acad. Sci. USA 1996, 93, 14564–14568. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, T.; Srinivasan, B.; Padmanabhan, S. Salivary leptin levels in normal weight and overweight individuals and their correlation with orthodontic tooth movement. Angle Orthod. 2017, 87, 739–744. [Google Scholar] [CrossRef]
- Sá-Pinto, A.C.; Rego, T.M.; Marques, L.S.; Martins, C.C.; Ramos-Jorge, M.L.; Ramos-Jorge, J. Association between malocclusion and dental caries in adolescents: A systematic review and meta-analysis. Eur. Arch. Paediatr. Dent. 2018, 19, 73–82. [Google Scholar] [CrossRef]
- Alsulaiman, A.A.; Kaye, E.; Jones, J.; Cabral, H.; Leone, C.; Will, L.; Garcia, R. Incisor malalignment and the risk of periodontal disease progression. Am. J. Orthod. Dentofacial Orthop. 2018, 153, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Brook, A.H. Multilevel complex interactions between genetic, epigenetic and environmental factors in the aetiology of anomalies of dental development. Arch. Oral Biol. 2009, 54 (Suppl. S1), S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef]
- Krishnan, V.; Davidovitch, Z. Cellular, molecular, and tissue-level reactions to orthodontic force. Am. J. Orthod. Dentofacial Orthop. 2006, 129, 469.e1–469.e32. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Bauer, K.; Spanier, G.; Proff, P.; Wolf, M.; Kirschneck, C. Expression kinetics of human periodontal ligament fibroblasts in the early phases of orthodontic tooth movement. J. Orofac. Orthop. 2018, 79, 337–351. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor kappaB ligand up-regulation via prostaglandin E2 synthesis. J. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Andreev, D.; Liu, M.; Weidner, D.; Kachler, K.; Faas, M.; Grüneboom, A.; Schlötzer-Schrehardt, U.; Muñoz, L.E.; Steffen, U.; Grötsch, B.; et al. Osteocyte necrosis triggers osteoclast-mediated bone loss through macrophage-inducible C-type lectin. J. Clin. Investig. 2020, 130, 4811–4830. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Käppler, P.; Nazet, U.; Jantsch, J.; Proff, P.; Cieplik, F.; Deschner, J.; Kirschneck, C. Effects of Compressive and Tensile Strain on Macrophages during Simulated Orthodontic Tooth Movement. Mediators Inflamm. 2020, 2020, 2814015. [Google Scholar] [CrossRef]
- Hanada, R.; Hanada, T.; Penninger, J.M. Physiology and pathophysiology of the RANKL/RANK system. Biol. Chem. 2010, 391, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Schröder, A.; Meyer, A.; Spanier, G.; Damanaki, A.; Paddenberg, E.; Proff, P.; Kirschneck, C. Impact of Leptin on Periodontal Ligament Fibroblasts during Mechanical Strain. Int. J. Mol. Sci. 2021, 22, 6847. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-G.; Kim, J.-H.; Cha, J.-H.; Bak, E.-J.; Yoo, Y.-J. Induction of IL-6 and IL-8 Expression by Leptin Treatment in Periodontal Ligament Cells and Gingival Fibroblasts. Int. J. Oral Biol. 2013, 38, 73–80. [Google Scholar] [CrossRef]
- Yan, B.; Wang, L.; Li, J.; Yang, R.; Liu, Y.; Yu, T.; He, D.; Zhou, Y.; Liu, D. Effects of the multifunctional hormone leptin on orthodontic tooth movement in rats. Am. J. Transl. Res. 2020, 12, 1976–1984. [Google Scholar]
- Yan, B.; Liu, D.; Zhang, C.; Zhang, T.; Wang, X.; Yang, R.; Liu, Y.; He, D.; Zhou, Y. Obesity attenuates force-induced tooth movement in mice with the elevation of leptin level: A preliminary translational study. Am. J. Transl. Res. 2018, 10, 4107–4118. [Google Scholar]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef]
- Karthikeyan, B.V.; Pradeep, A.R. Leptin levels in gingival crevicular fluid in periodontal health and disease. J. Periodontal Res. 2007, 42, 300–304. [Google Scholar] [CrossRef]
- Lago, F.; Dieguez, C.; Gómez-Reino, J.; Gualillo, O. Adipokines as emerging mediators of immune response and inflammation. Nat. Clin. Pract. Rheumatol. 2007, 3, 716–724. [Google Scholar] [CrossRef]
- Issa, R.I.; Griffin, T.M. Pathobiology of obesity and osteoarthritis: Integrating biomechanics and inflammation. Pathobiol. Aging Age Relat. Dis. 2012, 2, 17470. [Google Scholar] [CrossRef] [PubMed]
- Naylor, C.; Petri, W.A. Leptin Regulation of Immune Responses. Trends Mol. Med. 2016, 22, 88–98. [Google Scholar] [CrossRef]
- Loffreda, S.; Yang, S.Q.; Lin, H.Z.; Karp, C.L.; Brengman, M.L.; Wang, D.J.; Klein, A.S.; Bulkley, G.B.; Bao, C.; Noble, P.W.; et al. Leptin regulates proinflammatory immune responses. FASEB J. 1998, 12, 57–65. [Google Scholar] [CrossRef]
- Tsiotra, P.C.; Boutati, E.; Dimitriadis, G.; Raptis, S.A. High insulin and leptin increase resistin and inflammatory cytokine production from human mononuclear cells. Biomed Res. Int. 2013, 2013, 487081. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Da Park, J.; Kim, Y.H.; Kim, Y.; Kim, S.G.; Shon, K.J.; Choi, Y.-W.; Lee, S.-J. Upregulation of heme oxygenase-1 via PI3K/Akt and Nrf-2 signaling pathways mediates the anti-inflammatory activity of Schisandrin in Porphyromonas gingivalis LPS-stimulated macrophages. Immunol. Lett. 2011, 139, 93–101. [Google Scholar] [CrossRef]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 2014, 476068. [Google Scholar] [CrossRef]
- Diya, Z.; Lili, C.; Shenglai, L.; Zhiyuan, G.; Jie, Y. Lipopolysaccharide (LPS) of Porphyromonas gingivalis induces IL-1beta, TNF-alpha and IL-6 production by THP-1 cells in a way different from that of Escherichia coli LPS. Innate Immun. 2008, 14, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Sakaida, I.; Uchida, K.; Terai, S.; Okita, K. Leptin enhances TNF-alpha production via p38 and JNK MAPK in LPS-stimulated Kupffer cells. Life Sci. 2005, 77, 1502–1515. [Google Scholar] [CrossRef]
- Landgraf, M.A.; Silva, R.C.; Corrêa-Costa, M.; Hiyane, M.I.; Carvalho, M.H.C.; Landgraf, R.G.; Câmara, N.O.S. Leptin downregulates LPS-induced lung injury: Role of corticosterone and insulin. Cell. Physiol. Biochem. 2014, 33, 835–846. [Google Scholar] [CrossRef]
- Abd Alkhaleq, H.; Kornowski, R.; Waldman, M.; Zemel, R.; Lev, D.L.; Shainberg, A.; Miskin, R.; Hochhauser, E. Leptin modulates gene expression in the heart, cardiomyocytes and the adipose tissue thus mitigating LPS-induced damage. Exp. Cell Res. 2021, 404, 112647. [Google Scholar] [CrossRef]
- Schröder, A.; Barschkies, L.; Jantsch, J.; Proff, P.; Gölz, L.; Deschner, J.; Kirschneck, C. Role of Oxygen Supply in Macrophages in a Model of Simulated Orthodontic Tooth Movement. Mediators Inflamm. 2020, 2020, 5802435. [Google Scholar] [CrossRef] [PubMed]
- Nazet, U.; Schröder, A.; Spanier, G.; Wolf, M.; Proff, P.; Kirschneck, C. Simplified method for applying static isotropic tensile strain in cell culture experiments with identification of valid RT-qPCR reference genes for PDL fibroblasts. Eur. J. Orthod. 2020, 42, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | 5′-Forward Primer-3′ | 5′-Reverse Primer-3′ |
|---|---|---|---|
| Eef1a1 | Eukaryotic Translation Elongation Factor-1-α-1 | AAAACATGATTACAGGCACATCCC | GCCCGTTCTTGGAGATACCAG |
| Gapdh | Glyceraldehyde-3-phosphate dehydrogenase | GTCATCCCAGAGCTGAACGG | ATGCCTGCTTCACCACCTTC |
| Il1b | Interleukin-1-beta | GTGTAATGAAAGACGGCACACC | ACCAGTTGGGGAACTCTGC |
| Il6 | Interleukin-6 | ACAAAGCCAGAGTCCTTCAGAG | GAGCATTGGAAATTGGGGTAGG |
| Ptgs2 | prostaglandin-endoperoxide synthase-2 | TCCCTGAAGCCGTACACATC | TCCCCAAAGATAGCATCTGGAC |
| Sdha | Succinate Dehydrogenase Complex Flavoprotein Subunit A | AACACTGGAGGAAGCACACC | AGTAGGAGCGGATAGCAGGAG |
| Tbp | TATA-box-binding protein | CTATCACTCCTGCCACACCAG | CACGAAGTGCAATGGTCTTTAGG |
| Tnf | Tumor necrosis factor | ACAAGCCTGTAGCCCACGTC | TTGTTGTCTTTGAGATCCATGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paddenberg, E.; Osterloh, H.; Jantsch, J.; Nogueira, A.; Proff, P.; Kirschneck, C.; Schröder, A. Impact of Leptin on the Expression Profile of Macrophages during Mechanical Strain In Vitro. Int. J. Mol. Sci. 2022, 23, 10727. https://doi.org/10.3390/ijms231810727
Paddenberg E, Osterloh H, Jantsch J, Nogueira A, Proff P, Kirschneck C, Schröder A. Impact of Leptin on the Expression Profile of Macrophages during Mechanical Strain In Vitro. International Journal of Molecular Sciences. 2022; 23(18):10727. https://doi.org/10.3390/ijms231810727
Chicago/Turabian StylePaddenberg, Eva, Hannah Osterloh, Jonathan Jantsch, Andressa Nogueira, Peter Proff, Christian Kirschneck, and Agnes Schröder. 2022. "Impact of Leptin on the Expression Profile of Macrophages during Mechanical Strain In Vitro" International Journal of Molecular Sciences 23, no. 18: 10727. https://doi.org/10.3390/ijms231810727
APA StylePaddenberg, E., Osterloh, H., Jantsch, J., Nogueira, A., Proff, P., Kirschneck, C., & Schröder, A. (2022). Impact of Leptin on the Expression Profile of Macrophages during Mechanical Strain In Vitro. International Journal of Molecular Sciences, 23(18), 10727. https://doi.org/10.3390/ijms231810727