Screening and Characterization of Shark-Derived VNARs against SARS-CoV-2 Spike RBD Protein

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Four Unique VNAR Binders Targeting SARS-CoV-2 RBD Were Identified from Immunized C. plagiosum
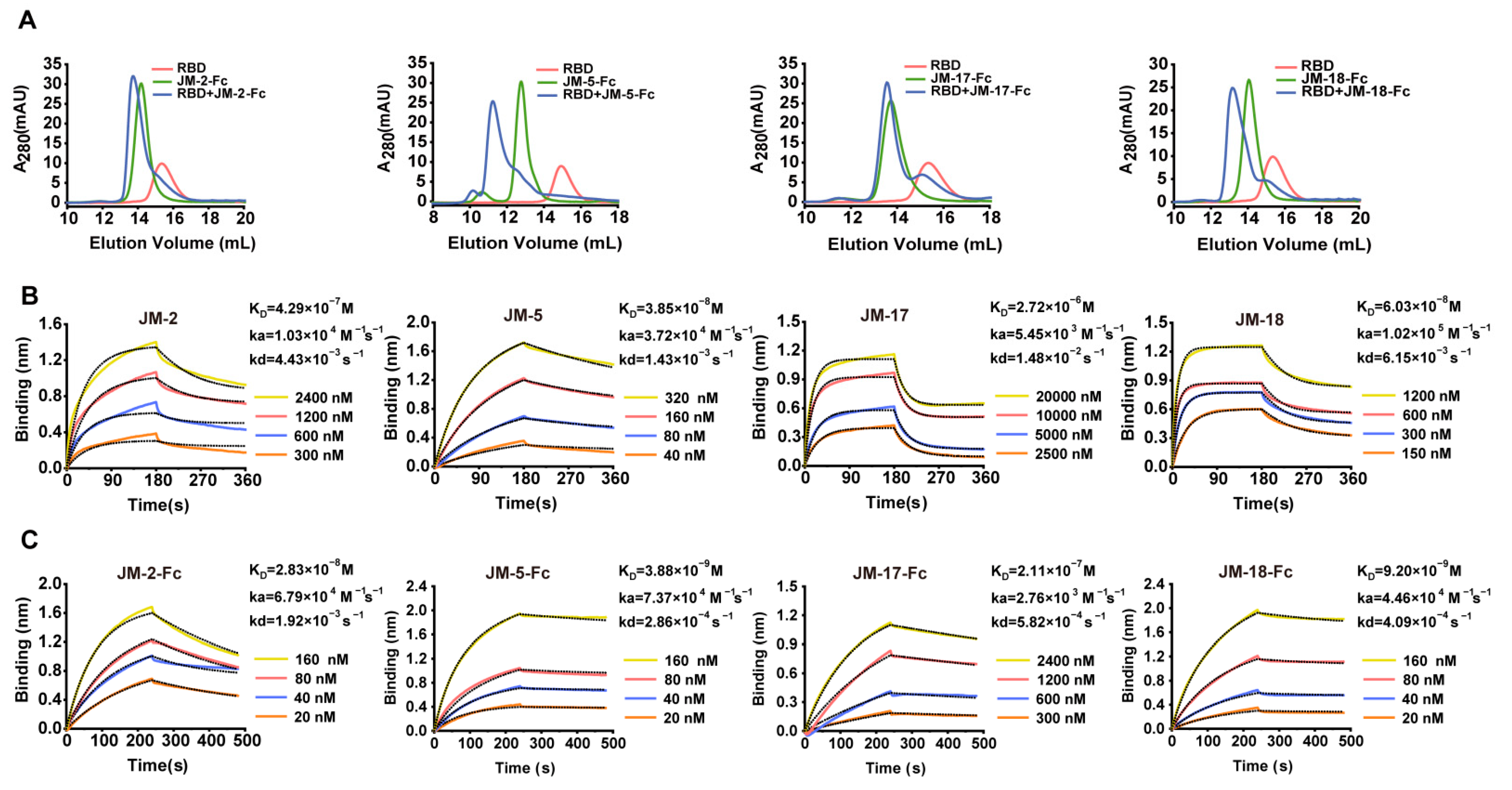
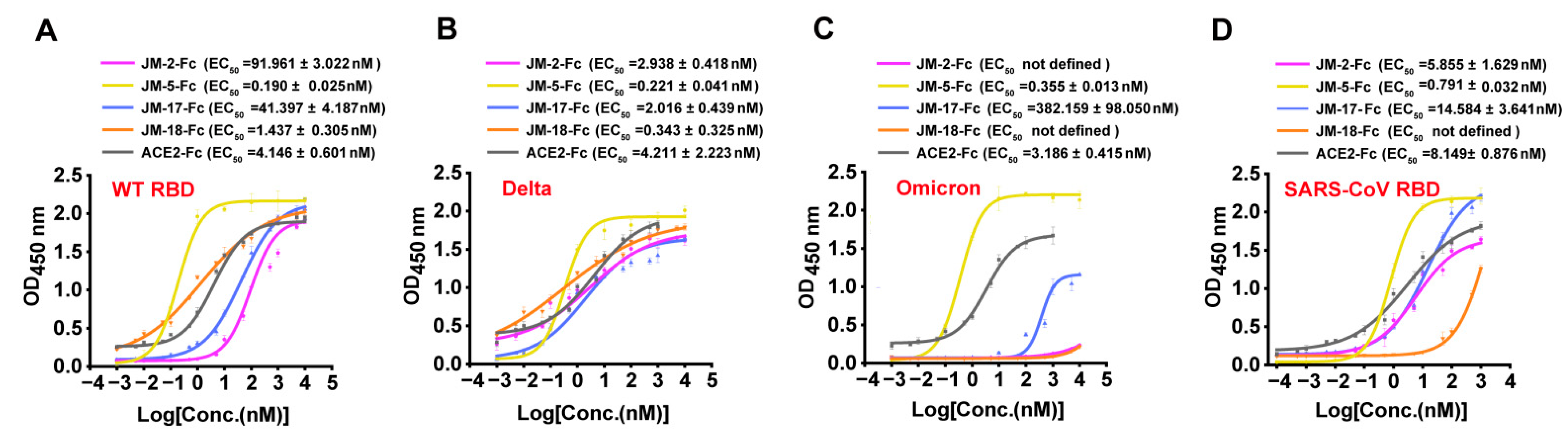
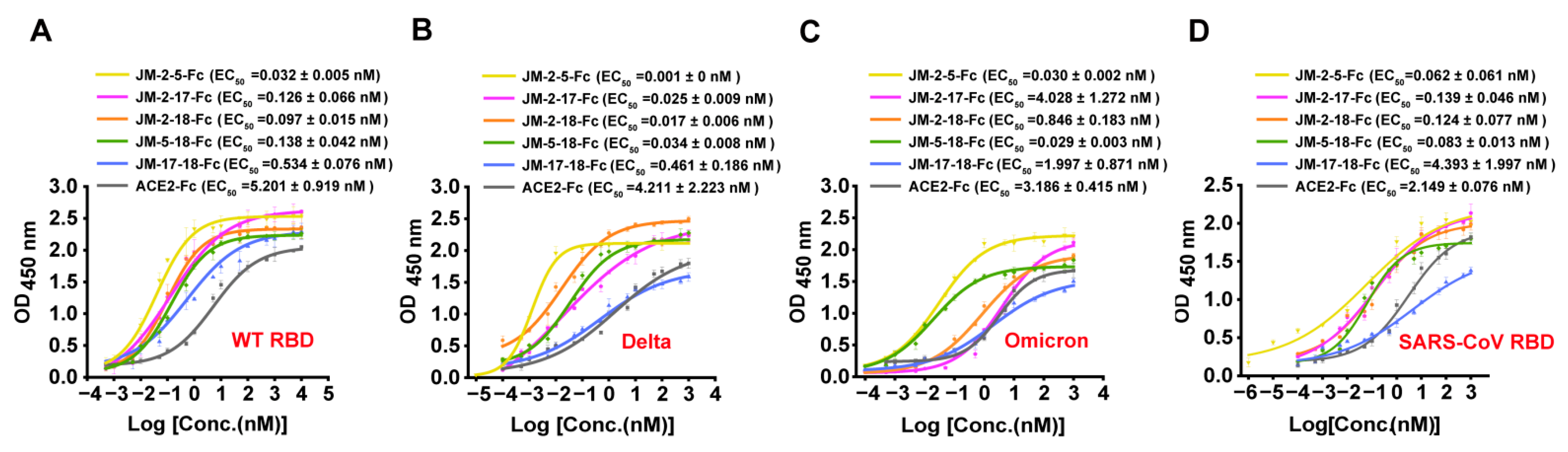
2.2. Characterization of RBD-Targeted VNARs
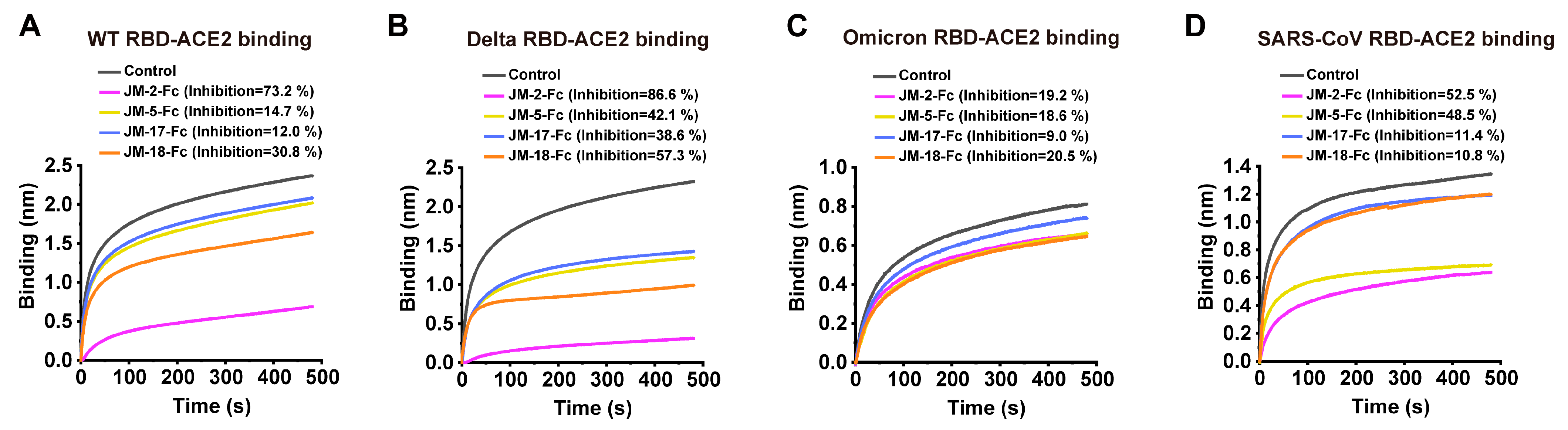
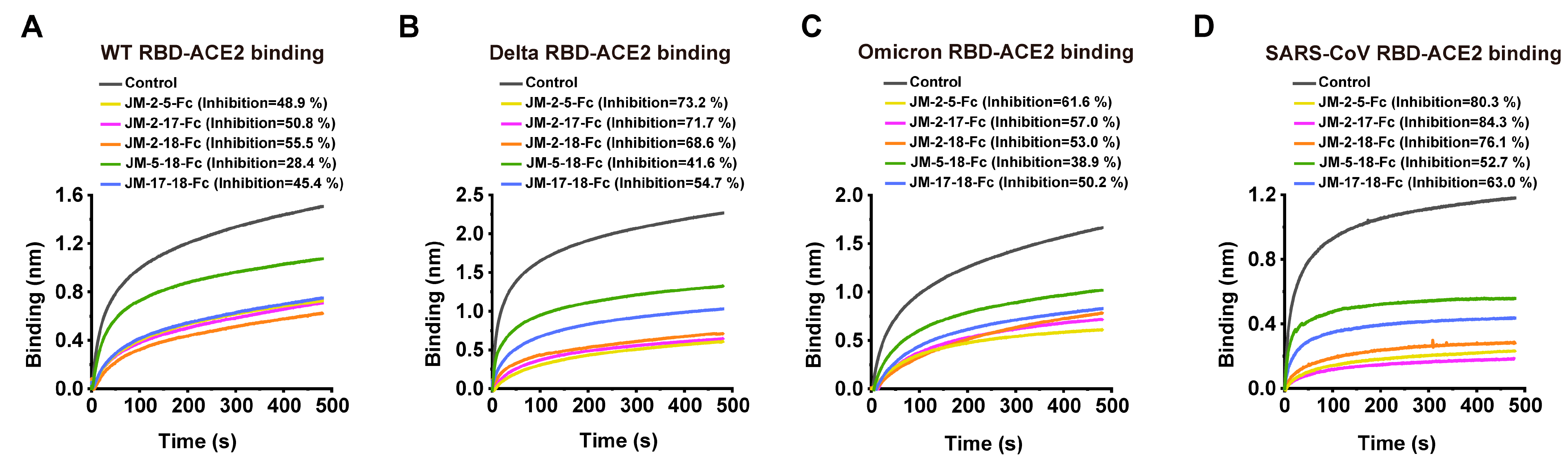
2.3. RBD-ACE2 Blockage of VNARs
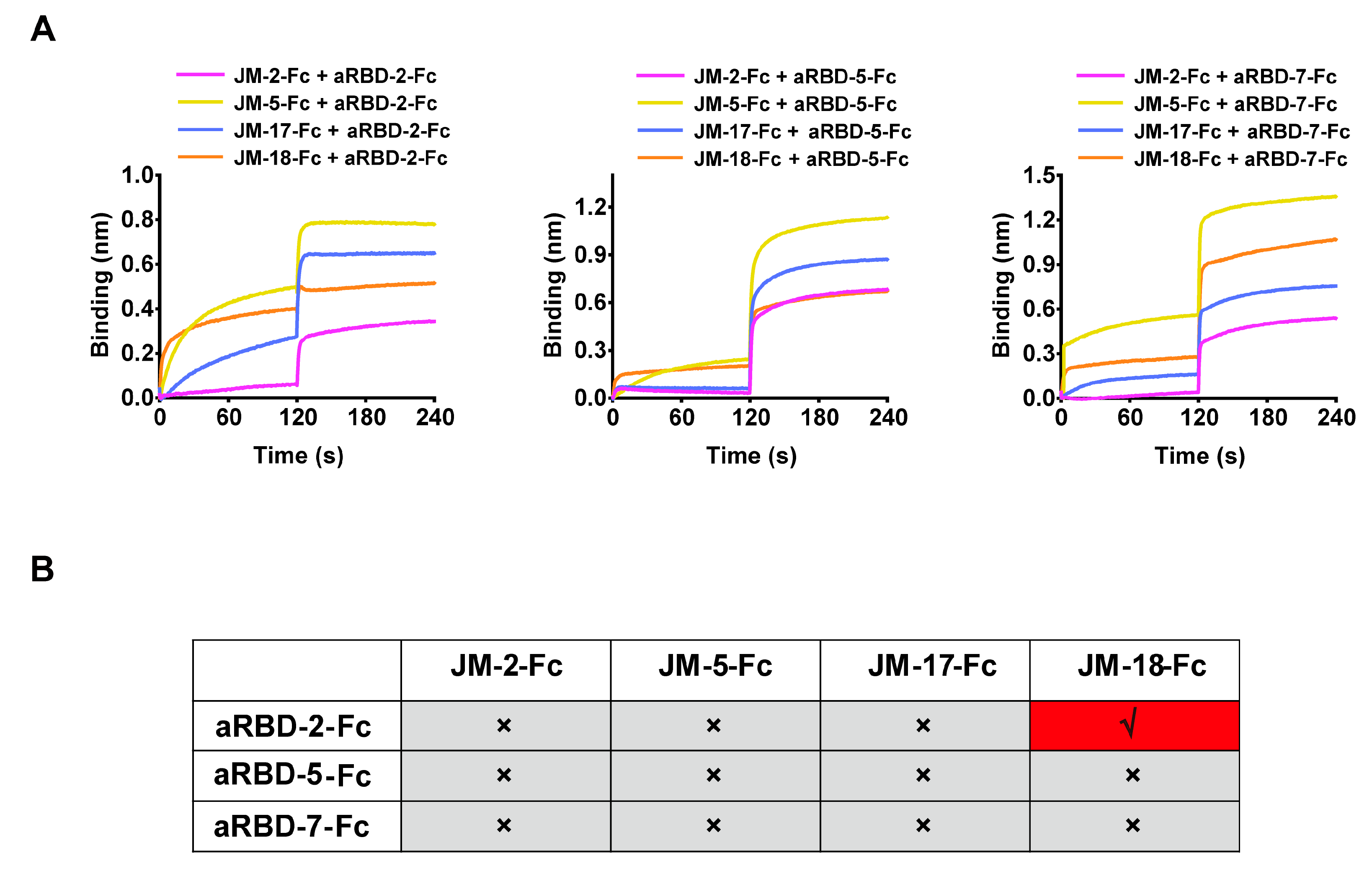
2.4. Epitope Competition of VNARs
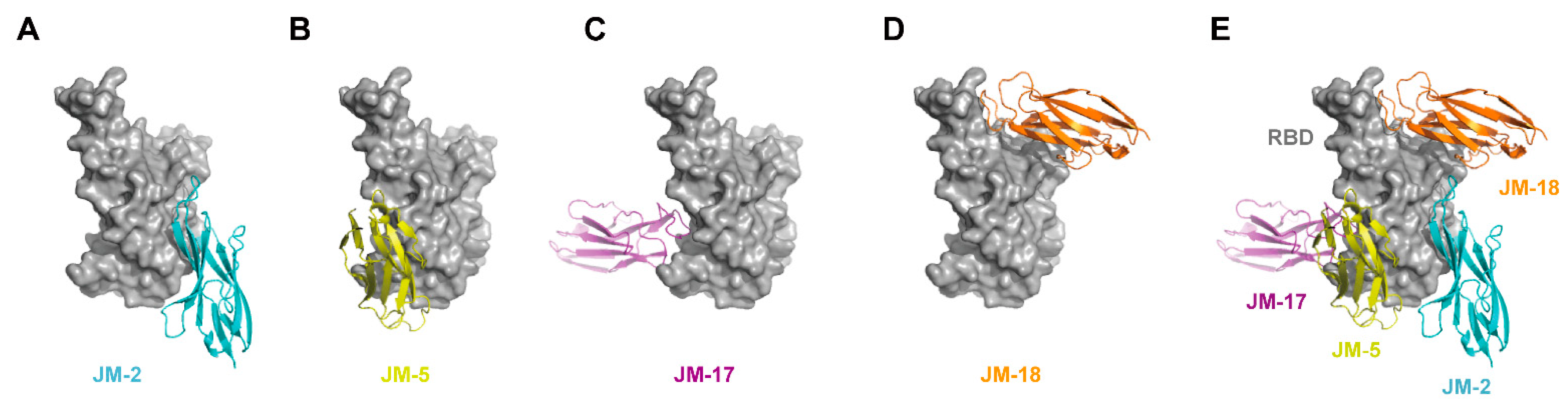
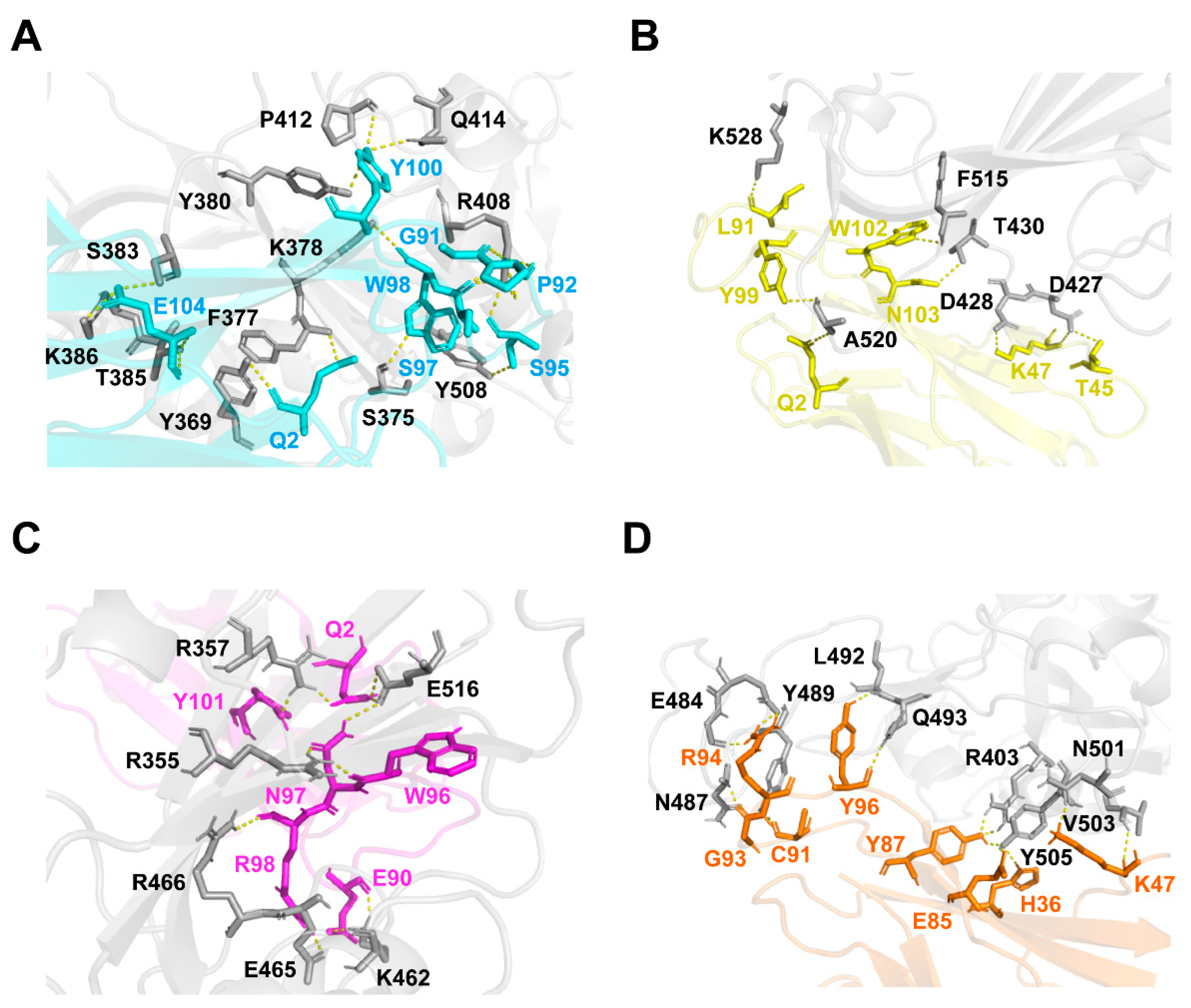
2.5. Prediction of the Binding Sites of VNARs on RBD Using Docking Simulation
2.6. Construction and Characterization of High-Affinity Bi-Paratopic Antibodies
3. Discussion
4. Materials and Methods
4.1. Plasmids and Cell Culture
4.2. Protein Expression and Purification
4.3. Biotinylation of RBD and ACE2-Fc
4.4. Phage Library Construction
4.5. Biopanning and Selection of Positive Clones
4.6. Determination of Melting Temperatures of VNARs via Thermal Shift Assay
4.7. Gel Filtration Chromatography
4.8. Affinity Determination
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. ACE2-RBD Blocking Assay
4.11. Epitope Binning
4.12. Prediction of the Binding Sites of VNARs on RBD with the ClusPro Server
4.13. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yan, R.H.; Zhang, Y.Y.; Li, Y.N.; Xia, L.; Guo, Y.Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.S.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Ghasemiyeh, P.; Mohammadi-Samani, S.; Firouzabadi, N.; Dehshahri, A.; Vazin, A. A focused review on technologies, mechanisms, safety, and efficacy of available COVID-19 vaccines. Int. Immunopharmacol. 2021, 100, 108162. [Google Scholar] [CrossRef] [PubMed]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yuan, Q.; Wang, H.; Liu, W.; Liao, X.; Su, Y.; Wang, X.; Yuan, J.; Li, T.; Li, J.; et al. Antibody responses to SARS-CoV-2 in patients with novel coronavirus disease 2019. Clin. Infect. Dis. 2020, 71, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, H.R.; Parai, D.; Dash, G.C.; Peter, A.; Sahoo, S.K.; Pattnaik, M.; Rout, U.K.; Nanda, R.R.; Pati, S.; Bhattacharya, D. IgG antibody response against nucleocapsid and spike protein post-SARS-CoV-2 infection. Infection 2021, 49, 1045–1048. [Google Scholar] [CrossRef]
- Gallais, F.; Gantner, P.; Bruel, T.; Velay, A.; Planas, D.; Wendling, M.J.; Bayer, S.; Solis, M.; Laugel, E.; Reix, N.; et al. Evolution of antibody responses up to 13 months after SARS-CoV-2 infection and risk of reinfection. eBioMedicine 2021, 71, 103561. [Google Scholar] [CrossRef]
- Carrillo, J.; Izquierdo-Useros, N.; Ávila-Nieto, C.; Pradenas, E.; Clotet, B.; Blanco, J. Humoral immune responses and neutralizing antibodies against SARS-CoV-2; implications in pathogenesis and protective immunity. Biochem. Biophys. Res. Commun. 2021, 538, 187–191. [Google Scholar] [CrossRef]
- Taylor, P.C.; Adams, A.C.; Hufford, M.M.; de la Torre, I.; Winthrop, K.; Gottlieb, R.L. Neutralizing monoclonal antibodies for treatment of COVID-19. Nat. Rev. Immunol. 2021, 21, 382–393. [Google Scholar] [CrossRef]
- Quinti, I.; Mortari, E.P.; Salinas, A.F.; Milito, C.; Carsetti, R. IgA antibodies and IgA deficiency in SARS-CoV-2 infection. Front. Cell. Infect. Microbiol. 2021, 11, 655896. [Google Scholar] [CrossRef]
- Fenwick, C.; Turelli, P.; Perez, L.; Pellaton, C.; Esteves-Leuenberger, L.; Farina, A.; Campos, J.; Lana, E.; Fiscalini, F.; Raclot, C.; et al. A highly potent antibody effective against SARS-CoV-2 variants of concern. Cell Rep. 2021, 37, 109814. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.A.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 2020, 586, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Earle, K.A.; Ambrosino, D.M.; Fiore-Gartland, A.; Goldblatt, D.; Gilbert, P.B.; Siber, G.R.; Dull, P.; Plotkin, S.A. Evidence for antibody as a protective correlate for COVID-19 vaccines. Vaccine 2021, 39, 4423–4428. [Google Scholar] [CrossRef] [PubMed]
- Favresse, J.; Gillot, C.; Di Chiaro, L.; Eucher, C.; Elsen, M.; Van Eeckhoudt, S.; David, C.; Morimont, L.; Dogné, J.M.; Douxfils, J. Neutralizing antibodies in COVID-19 patients and vaccine recipients after two doses of BNT162b2. Viruses 2021, 13, 1364. [Google Scholar] [CrossRef]
- Ali, H.; Alahmad, B.; Al-Shammari, A.A.; Alterki, A.; Hammad, M.; Cherian, P.; Alkhairi, I.; Sindhu, S.; Thanaraj, T.A.; Mohammad, A.; et al. Previous COVID-19 infection and antibody levels after vaccination. Front. Public Health 2021, 9, 778243. [Google Scholar] [CrossRef]
- Chi, X.J.; Liu, X.Y.; Wang, C.H.; Zhang, X.H.; Li, X.; Hou, J.H.; Ren, L.L.; Jin, Q.; Wang, J.W.; Yang, W. Humanized single domain antibodies neutralize SARS-CoV-2 by targeting the spike receptor binding domain. Nat. Commun. 2020, 11, 4528. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Tedla, N.; Bull, R.A. Broadly-neutralizing antibodies against emerging SARS-CoV-2 variants. Front. Immunol. 2021, 12, 752003. [Google Scholar] [CrossRef]
- Ju, B.; Zhang, Q.; Ge, J.; Wang, R.; Sun, J.; Ge, X.; Yu, J.; Shan, S.; Zhou, B.; Song, S.; et al. Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 2020, 584, 115–119. [Google Scholar] [CrossRef]
- Cao, Y.; Su, B.; Guo, X.; Sun, W.; Deng, Y.; Bao, L.; Zhu, Q.; Zhang, X.; Zheng, Y.; Geng, C.; et al. Potent neutralizing antibodies against SARS-CoV-2 identified by high-throughput single-cell sequencing of convalescent patients’ B cells. Cell 2020, 182, 73–84. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.J.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; van Haperen, R.; Osterhaus, A.D.M.E.; van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.-J. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shan, C.; Duan, X.M.; Chen, Z.H.; Liu, P.P.; Song, J.W.; Song, T.; Bi, X.S.; Han, C.; Wu, L.A.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.F.; Zhao, F.Z.; Huang, D.L.; Beutler, N.; Burns, A.; He, W.T.; Limbo, O.; Smith, C.; Song, G.; Woehl, J.; et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef]
- Chen, X.Y.; Li, R.; Pan, Z.W.; Qian, C.F.; Yang, Y.; You, R.R.; Zhao, J.; Liu, P.H.; Gao, L.Q.; Li, Z.R.; et al. Human monoclonal antibodies block the binding of SARS-CoV-2 spike protein to angiotensin converting enzyme 2 receptor. Cell. Mol. Immunol. 2020, 17, 647–649. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally-occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Avila, D.; Hughes, M.; Hughes, A.; McKinney, E.C.; Flajnik, M.F. A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature 1995, 374, 168–173. [Google Scholar] [CrossRef]
- Cheong, W.S.; Leow, C.Y.; Majeed, A.A.; Leow, C.H. Diagnostic and therapeutic potential of shark variable new antigen receptor (VNAR) single domain antibody. Int. J. Biol. Macromol. 2020, 147, 369–375. [Google Scholar] [CrossRef]
- Matz, H.; Dooley, H. Shark IgNAR-derived binding domains as potential diagnostic and therapeutic agents. Dev. Comp. Immunol. 2019, 90, 100–107. [Google Scholar] [CrossRef]
- Khalid, Z.; Chen, Y.; Yu, D.; Abbas, M.; Huan, M.; Naz, Z.; Mengist, H.M.; Cao, M.J.; Jin, T. IgNAR antibody: Structural features, diversity and applications. Fish Shellfish Immunol. 2022, 121, 467–477. [Google Scholar] [CrossRef]
- Walsh, R.; Nuttall, S.; Revill, P.; Colledge, D.; Cabuang, L.; Soppe, S.; Dolezal, O.; Griffiths, K.; Bartholomeusz, A.; Locarnini, S. Targeting the hepatitis B virus precore antigen with a novel IgNAR single variable domain intrabody. Virology 2011, 411, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Goodchild, S.A.; Dooley, H.; Schoepp, R.J.; Flajnik, M.; Lonsdale, S.G. Isolation and characterisation of Ebolavirus-specific recombinant antibody fragments from murine and shark immune libraries. Mol. Immunol. 2011, 48, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhang, X.; Zheng, P.; Dube, P.H.; Zeng, W.; Chen, S.; Cheng, Q.; Yang, Y.; Wu, Y.; Zhou, J.; et al. Hetero-bivalent nanobodies provide broad-spectrum protection against SARS-CoV-2 variants of concern including Omicron. Cell. Res. 2022, 32, 831–842. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Girgis, M.; Sanda, M. Site-specific glycosylation of SARS-CoV-2: Big challenges in mass spectrometry analysis. Proteomics 2022, 22, e2100322. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, Y.; Chen, M.; Wu, G.; Yan, C.; Zhang, X.E. SARS-CoV-2 spike protein receptor-binding domain N-glycans facilitate viral internalization in respiratory epithelial cells. Biochem. Biophys. Res. Commun. 2021, 579, 69–75. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Hanke, L.; Perez, L.V.; Sheward, D.J.; Das, H.; Schulte, T.; Moliner-Morro, A.; Corcoran, M.; Achour, A.; Hedestam, G.B.K.; Hällberg, B.M.; et al. An alpaca nanobody neutralizes SARS-CoV-2 by blocking receptor interaction. Nat. Commun. 2020, 11, 4420. [Google Scholar] [CrossRef]
- Koenig, P.A.; Das, H.; Liu, H.; Kümmerer, B.M.; Gohr, F.N.; Jenster, L.M.; Schiffelers, L.D.J.; Tesfamariam, Y.M.; Uchima, M.; Wuerth, J.D.; et al. Structure-guided multivalent nanobodies block SARS-CoV-2 infection and suppress mutational escape. Science 2021, 371, eabe6230. [Google Scholar] [CrossRef]
- Li, T.; Cai, H.; Yao, H.; Zhou, B.; Zhang, N.; van Vlissingen, M.F.; Kuiken, T.; Han, W.; GeurtsvanKessel, C.H.; Gong, Y.; et al. A synthetic nanobody targeting RBD protects hamsters from SARS-CoV-2 infection. Nat. Commun. 2021, 12, 4635. [Google Scholar] [CrossRef]
- Xu, J.; Xu, K.; Jung, S.; Conte, A.; Lieberman, J.; Muecksch, F.; Lorenzi, J.C.C.; Park, S.; Schmidt, F.; Wang, Z.; et al. Nanobodies from camelid mice and llamas neutralize SARS-CoV-2 variants. Nature 2021, 595, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zeng, W.; Meng, X.; Huang, X.; Yang, Y.; Zhao, D.; Zhou, P.; Wang, X.; Zhao, C.; Sun, Y.; et al. Potent neutralization of SARS-CoV-2 by hetero-bivalent alpaca nanobodies targeting the spike receptor-binding domain. J. Virol. 2021, 95, e02438-20. [Google Scholar] [CrossRef] [PubMed]
- Schoof, M.; Faust, B.; Saunders, R.A.; Sangwan, S.; Rezelj, V.; Hoppe, N.; Boone, M.; Billesbolle, C.B.; Puchades, C.; Azumaya, C.M.; et al. An ultrapotent synthetic nanobody neutralizes SARS-CoV-2 by stabilizing inactive Spike. Science 2020, 370, 1473–1479. [Google Scholar] [CrossRef]
- Dong, J.B.; Huang, B.; Wang, B.; Titong, A.; Kankanamalage, S.G.; Jia, Z.J.; Wright, M.; Parthasarathy, P.; Liu, Y. Development of humanized tri-specific nanobodies with potent neutralization for SARS-CoV-2. Sci. Rep. 2021, 10, 17806. [Google Scholar] [CrossRef]
- Dong, J.; Huang, B.; Jia, Z.; Wang, B.; Kankanamalage, S.G.; Titong, A.; Liu, Y. Development of multi-specific humanized llama antibodies blocking SARS-CoV-2/ACE2 interaction with high affinity and avidity. Emerg. Microbes Infect. 2020, 9, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.F.; Nambulli, S.; Xiao, Z.Y.; Liu, H.; Sang, Z.; Duprex, W.P.; Schneidman-Duhovny, D.; Zhang, C.; Shi, Y. Versatile and multivalent nanobodies efficiently neutralize SARS-CoV-2. Science 2020, 370, 1479–1484. [Google Scholar] [CrossRef]
- Gauhar, A.; Privezentzev, C.V.; Demydchuk, M.; Gerlza, T.; Rieger, J.; Kungl, A.J.; Walsh, F.S.; Rutkowski, J.L.; Stocki, P. Single domain shark VNAR antibodies neutralize SARS-CoV-2 infection in vitro. FASEB J. 2021, 35, e21970. [Google Scholar] [CrossRef]
- Ubah, O.C.; Lake, E.W.; Gunaratne, G.S.; Gallant, J.P.; Fernie, M.; Robertson, A.J.; Marchant, J.S.; Bold, T.D.; Langlois, R.A.; Matchett, W.E.; et al. Mechanisms of SARS-CoV-2 neutralization by shark variable new antigen receptors elucidated through X-ray crystallography. Nat. Commun. 2021, 12, 7325. [Google Scholar] [CrossRef]
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.G.; Sharaf, N.G.; Huey-Tubman, K.E.; Lee, Y.E.; et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A noncompeting pair of human neutralizing antibodies block COVID-19 virus binding to its receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Mohammed, A.; Zeng, W.; Mengist, H.M.; Kombe, A.J.K.; Ou, H.; Yang, Y.; Dan, Z.; Xu, Z.; Ma, H.; Jin, T. Generation, biochemical characterizations and validation of potent nanobodies derived from alpaca specific for human receptor of advanced glycation end product. Biochem. Biophys. Res. Commun. 2021, 581, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Chen, Z.L.; Sun, J.; Xu, T.T.; Wang, Q.; Yi, H.S.; Niu, X.F.; Zhu, J.B.; Fan, M.Z.; Hou, R.T.; et al. A class of shark-derived single-domain antibodies can broadly neutralize SARS-related coronaviruses and the structural basis of neutralization and Omicron escape. Small Methods 2022, 6, 2200387. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Schafer, A.; Kulkarni, S.S.; Liu, X.L.; Martinez, D.R.; Chen, C.; Sun, Z.H.; Leist, S.R.; Drelich, A.; Zhang, L.Y.; et al. High potency of a bivalent human VH domain in SARS-CoV-2 animal models. Cell 2020, 183, 429–441.e16. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Ma, H.; Ding, C.; Yang, Y.; Sun, Y.; Huang, X.; He, W.; Xiang, Y.; Gao, Y.; Jin, T. Characterization of SARS-CoV-2-specific antibodies in COVID-19 patients reveals highly potent neutralizing IgA. Signal Transduct. Target. Ther. 2021, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zeng, W.; He, H.; Zhao, D.; Jiang, D.; Zhou, P.; Cheng, L.; Li, Y.; Ma, X.; Jin, T. Serum IgA, IgM, and IgG responses in COVID-19. Cell. Mol. Immunol. 2020, 17, 773–775. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VNAR | KD (M) | ka (M−1s−1) | kd (s−1) | Fold of KD Increase |
|---|---|---|---|---|
| JM-2 | 4.29 × 10−7 | 1.03 × 104 | 4.43 × 10−3 | N.A. |
| JM-5 | 3.85 × 10−8 | 3.72 × 104 | 1.43 × 10−3 | N.A. |
| JM-17 | 2.72 × 10−6 | 5.45 × 104 | 1.48 × 10−2 | N.A. |
| JM-18 | 6.03 × 10−8 | 1.02 × 105 | 6.15 × 10−3 | N.A. |
| JM-2-Fc | 2.83 × 10−8 | 6.79 × 104 | 1.92 × 10−3 | vs. JM-2:15 |
| JM-5-Fc | 3.88 × 10−9 | 7.37 × 104 | 2.86 × 10−4 | vs. JM-5:10 |
| JM-17-Fc | 2.11 × 10−7 | 2.76 × 103 | 5.82 × 10−4 | vs. JM-17:13 |
| JM-18-Fc | 9.20 × 10−9 | 4.46 × 104 | 4.09 × 10−4 | vs. JM-18:6.5 |
| JM-2-5 | 6.39 × 10−9 | 3.73 × 104 | 2.38 × 10−4 | vs. JM-2:149; vs. JM-5:6 |
| JM-2-17 | 3.21 × 10−8 | 1.69 × 104 | 5.44 × 10−4 | vs. JM-2:13; vs. JM-17:86 |
| JM-2-18 | 3.07 × 10−8 | 5.43 × 104 | 1.67 × 10−3 | vs. JM-2:14; vs. JM-18:2 |
| JM-5-18 | 3.40 × 10−7 | 4.75 × 104 | 1.62 × 10−2 | N.A. |
| JM-17-18 | 3.57 × 10−8 | 5.61 × 104 | 2.00 × 10−3 | vs. JM-17:76; vs. JM-18:1.7 |
| JM-2-5-Fc | <1 × 10−12 | 4.14 × 104 | <1 × 10−7 | >1000 |
| JM-2-17-Fc | <1 × 10−12 | 2.73 × 104 | <1 × 10−7 | >1000 |
| JM-2-18-Fc | <1 × 10−12 | 4.54 × 104 | <1 × 10−7 | >1000 |
| JM-5-18-Fc | <1 × 10−12 | 4.67 × 104 | <1 × 10−7 | >1000 |
| JM-17-18-Fc | 3.26 × 10−10 | 9.11 × 104 | 2.97 × 10−5 | vs. JM-17-18:109 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-L.; Lin, J.-J.; Ma, H.; Zhong, N.; Xie, X.-X.; Yang, Y.; Zheng, P.; Zhang, L.-J.; Jin, T.; Cao, M.-J. Screening and Characterization of Shark-Derived VNARs against SARS-CoV-2 Spike RBD Protein. Int. J. Mol. Sci. 2022, 23, 10904. https://doi.org/10.3390/ijms231810904
Chen Y-L, Lin J-J, Ma H, Zhong N, Xie X-X, Yang Y, Zheng P, Zhang L-J, Jin T, Cao M-J. Screening and Characterization of Shark-Derived VNARs against SARS-CoV-2 Spike RBD Protein. International Journal of Molecular Sciences. 2022; 23(18):10904. https://doi.org/10.3390/ijms231810904
Chicago/Turabian StyleChen, Yu-Lei, Jin-Jin Lin, Huan Ma, Ning Zhong, Xin-Xin Xie, Yunru Yang, Peiyi Zheng, Ling-Jing Zhang, Tengchuan Jin, and Min-Jie Cao. 2022. "Screening and Characterization of Shark-Derived VNARs against SARS-CoV-2 Spike RBD Protein" International Journal of Molecular Sciences 23, no. 18: 10904. https://doi.org/10.3390/ijms231810904