Abstract
Parkinson’s disease (PD) is a slowly progressive neurodegenerative disorder, characterized by the misfolding and aggregation of α-synuclein (α-syn) into Lewy bodies and the degeneration of dopaminergic neurons in the substantia nigra pars compacta. The urge for an early diagnosis biomarker comes from the fact that clinical manifestations of PD are estimated to appear once the substantia nigra has deteriorated and there has been a reduction of the dopamine levels from the striatum. Nowadays, extracellular vesicles (EVs) play an important role in the pathogenesis of neuro-degenerative diseases as PD. A systematic review dated August 2022 was carried out with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses with the aim to analyze the potential role of EVs as biomarkers for PD. From a total of 610 articles retrieved, 29 were eligible. This review discusses the role of EVs biochemistry and their cargo proteins, such as α-syn and DJ-1 among others, detected by a proteomic analysis as well as miRNAs and lncRNAs, as potential biomarkers that can be used to create standardized protocols for early PD diagnosis as well as to evaluate disease severity and progression.
Keywords:
Parkinson’s disease; extracellular vesicles; exosomes; biomarkers; α-synuclein; lncRNAs; miRNAs 1. Introduction
Parkinson’s disease (PD) is a slowly progressive neurodegenerative disorder, characterized by the misfolding and aggregation of α-synuclein (α-syn) into Lewy bodies and Lewy neurites and the degeneration of dopaminergic neurons in the substantia nigra pars compacta [1,2]. α-syn, the main fibrillar component of Lewy bodies, is also a protein found in the nervous system of disease-free individuals [3].
Clinical manifestations of PD are estimated to appear once the substantia nigra has deteriorated with a reduction of the dopamine levels from the striatum [4]. Clinical features include motor symptoms, such as resting tremor, muscle tone rigidity, postural and gait instability and bradykinesia [5,6]. As well, nonmotor symptoms are present, such as sleep disorders, cognitive impairment, constipation, dysphagia, salivation, hallucinations and delusions, among others [1,5].
Extracellular vesicles (EVs), as secreted membrane particles that are naturally released by all types of cells [7,8], have been of interest in different fields. They can be present in body fluids such as plasma, urine, milk, saliva, cerebrospinal fluid (CSF), bronchi alveolar lavage and synovial fluid among others [8,9,10]. Perhaps, the main role of EVs is their capacity to mediate cell-to-cell communication by transferring RNA, DNA, signaling complexes, lipids, membrane proteins, etc., in physiological but also pathophysiological conditions [2,11].
Nowadays, terms such as “microvesicles” or “ectosomes” [12], among others, have fallen into disuse, preferring terms such as “exosomes” or mainly “extracellular vesicles” as recommended by the International Society for Extracellular Vesicles in its latest statements [7].
The current knowledge of EVs in the pathogenesis of PD are starting to be elucidated. Nowadays, several research projects are focused on the study of misfolded proteins such as α-syn, DJ-1, long noncoding RNAs (lncRNAs) (which are transcribed RNA molecules with a length greater than 200 nucleotides that do not encode proteins but are involved in the regulation of gene expression through epigenetic, translation, transcription and post-transcription mechanisms) and miRNAs. In fact, the use of more purified isolation and characterization methods for EVs will help find new biomarkers for an early diagnosis of Parkinson’s disease [7,12,13,14].
The aim of this systematic review was to evaluate the viability of EVs as potential biomarkers for the detection and progression of PD. The analysis of EVs derived from different bodily fluids, their isolation methods and distinct cargos, bring us closer to understanding their implication and use as a potential tool for winning a part of the battle in this neurodegenerative disease.
2. Methods
This systematic review was developed according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement as presented in the flowchart below (Figure 1) and dated August 2022 using PubMed and Embase databases.
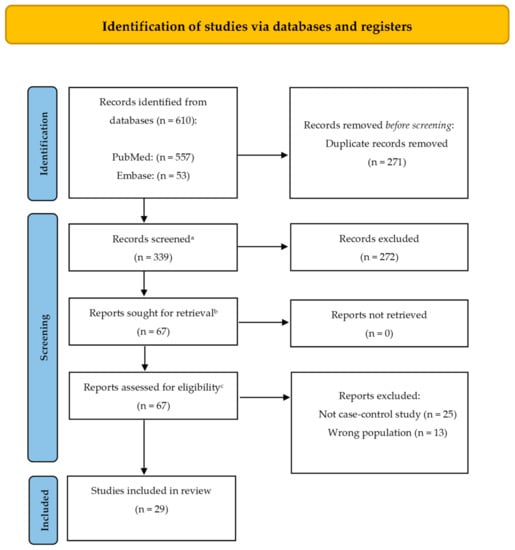
Figure 1.
PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) 2020 flow diagram representing the searching and selection process for this review [15]. a This is the number of records identified minus the number from the duplicates removed box; b This is the number of articles obtained in preparation for full text screening (subtract the number of excluded records from the total number screened (previous step) to obtain the number sought for retrieval); c This should be the number of reports sought for retrieval minus the number of reports not retrieved (previous step).
The search strategy was designed specifically for each of the search engines used in this review.
For the PubMed search, four different search strategies were developed to get the most accurate results:
- (exosomes[Title/Abstract]) AND parkinson’s disease[Title/Abstract]
- (exosomes[MeSH Terms]) AND parkinson’s disease[MeSH Terms]
- (extracellular vesicles[Title/Abstract]) AND parkinson’s disease[Title/Abstract]
- (extracellular vesicles[MeSH Terms]) AND parkinson’s disease[MeSH Terms]
For the Embase search, the strategy used was:
- exosomes:ti,ab,kw AND ‘parkinson disease’:ti,ab,kw
- extracellular vesicles:ti,ab,kw AND ‘parkinson disease’:ti,ab,kw
For inclusion criteria, only case–control studies were considered. As well, the year of publication was limited for this review. During the initial search, a significant increase in the search trends for this topic was observed in recent years (Figure 2). Thus, the year of publication was limited to the last 6 years with the purpose of analyzing the most noteworthy data available. Full texts in English were required in order to retrieve the information needed for this review: title, authors, year of publication, sample size and characteristics, inclusion/exclusion criteria, biological sample type, method of extraction of biological sample, method of isolation of exosomes and main outcomes.
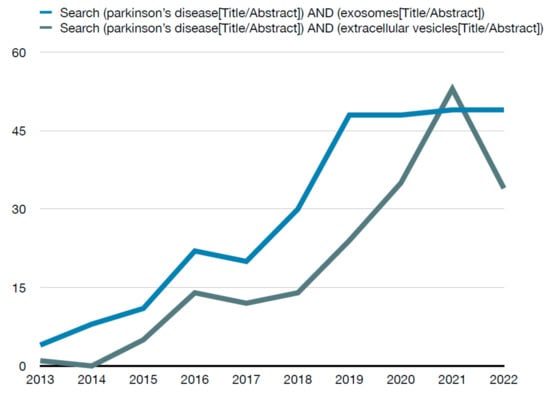
Figure 2.
Number of PubMed results per year for the search method.
As exclusion criteria, the reasons were report status as conference abstracts, review articles, case reports, letters, editorials, unpublished data, articles without full texts and articles not available in English.
Two teams of paired reviewers (J.V., M.F., J.M.S., A.M.) with expertise in medical and health evaluations and training in research methodology independently screened titles, abstracts, and full texts for eligibility, assessed generalizability, and collected data from each eligible case reports. Any disagreements were resolved by a third researcher (J.F.M.-T.).
3. Results and Discussion
A total number of 610 records were retrieved through database searching. After removing duplicates, 339 records were assessed for eligibility, and 29 articles were finally selected for this qualitative synthesis. Table 1 summarizes the records included and analyzed in this review.

Table 1.
Summary of studies analyzed and their main findings.
EVs play an imperative role in communication, transfer and delivery of information between the CNS and the circulatory system. Herein, the study of circulating EVs and their meaningful biochemistry and cargo (from prion proteins to miRNAs, LncRNAs) seems to elucidate now even more their role in PD pathogenesis.
As seen in Table 1 of all 29 studies analyzed, 22 of them used blood as a biological sample from which EVs were isolated, a clinical accessible test that still has the disadvantage of not reflecting the actual conditions of the CNS during the pathogenesis of PD [18]. The remaining studies used cerebrospinal fluid (two), urine (two), saliva (two) and blood and cerebrospinal fluid (one). In Table 2, we aimed to summarize the potential biomarkers that were analyzed in this review.
Table 2.
Summary of potential biomarkers in the analyzed studies.
3.1. α-syn and Its Derivatives
It is widely accepted that the accumulation of misfolded α-syn in PD has a primary role in the pathogenesis of the disease as the main component of Lewy bodies, along with ubiquitinated proteins, that accumulate in the surviving neurons [43]. α-syn can be found in the presynaptic terminals of neurons and participates in synaptic plasticity and vesicle trafficking [5,43]. It can be transferred neuron-to-neuron, with the capacity to form aggregates in the recipient cells but also propagate in a prion-like manner through the system [5].
The role of EVs in the carrying of α-syn as a possible pathological way of spreading for PD has been analyzed in 12 of the 29 studies. It was observed by Cerri et al. [14], Cao et al. [24] and Xia et al. [38] that the peripheral exosomal levels of α-syn in PD patients were higher than the levels in HC. EVs α-syn oligomers have garnered interest as the primary neurotoxic form of seeding healthy neurons [30].
Mutations in the α-syn-encoding gene SNCA, such as A53T, E46P, A30P, H50Q, G51D and A53E, promote the transformation of α-syn into its most toxic forms, fibrils and oligomers [44]. The oligomeric form of α-syn has been identified as the toxic variety participant in PD [30] and makes it easier for recipient cells to take up α-syn in this conformation rather than when it is in a free form [45,46]. In addition, the exosomal environment promotes α-syn aggregation as well, as a way of transportation for its propagation within the CNS and hence for the neurodegenerative process in PD [45,47,48].
Moreover, oligomeric α-syn has also been observed to be promoted by post-translational modifications (PTM) such as phosphorylation, nitration and dopamine (DA) modification. Particularly in PD, phosphorylation at Ser-129 seems to be the most prevalent PTM form of α-syn found in PD with Lewy bodies. It accelerates the formation of α-syn inclusions, as well as neuronal loss in mice [43,49].
Zhao et al. [18] studied DJ-1 and α-syn in plasma neural-derived exosomes, also using neuronal adhesion molecule L1 (L1CAM), and found significantly higher levels in the PD group than in HC. In addition, matching results, the levels of α-syn in neuronal exosomes in PD were higher compared to those in controls after electroanalyses were performed according to Fu et al. [32]. Moreover, Niu et al. [25] sustained that plasma neuronal exosomal α-syn could be involved in the pathophysiologic process of dopaminergic neurodegeneration in PD, based on the association between higher levels and increased risk of motor progression such as its ability to distinguish between early stage PD patients and HC. The expression of the neuronal adhesion molecule L1 (L1CAM) has been considered to reflect CNS status. Not only does L1 act as a cell surface receptor but it may also play a role as a ligand at a considerable distance from its origin [50]. These might explain the inconsistencies in multiple research projects that measured the same protein and why this might be increased in the analyzed samples.
On the contrary, Stuendl et al. [30] and Si et al. [39] analyzed α-syn-containing exosomes isolated from serum (immune-captured by an antibody directed against the neural L1 cell adhesion molecule L1CAM) and CSF samples, respectively, finding that the levels of exosomal α-syn were lower in PD patients than in HC and it was inverse to the course and severity of the disease.
Furthermore, Rani et al. [36] found that a cut-off value of 3.1 × 107 particles/mL of salivary exosomes distinguished the PD vs. HC group with a sensibility and specificity of 100%; however, a larger cohort is needed to validate these results.
Although multiple studies have quantified the levels of α-syn in its multiple conformations, primarily finding an increased in PD patients vs. healthy control groups [18,25,34], the concentration of salivary exosomes should be explored in larger cohorts, as a type of noninvasive biological sample with a 100% sensibility and specificity in prior results.
In brief, the studies published to date show inconsistencies regarding the potential of α-syn to be used as a biomarker for PD, as exposed above. This may have different explanations, such as the lack of standardized protocols for sample processing, the contamination of the actual sample or the variety of conformations studied, from unfolded monomer to complex fibrils such as oligomers and the use of L1CAM as an antibody for neural-derived EVs [3,51].
3.2. PrPc, DJ-1, OxiDJ-1 and Tau Protein
Cellular prion protein (PrPc), a cell-surface glycoprotein highly expressed in central and peripheral nervous system was found increased in plasma exosomes of PD patients vs. HC and associated with cognitive decline [16]. These results sustain the role of PrPc in the physiopathological process of PD.
DJ-1 gene encodes DJ-1, a chaperone protein whose main function is to inhibit the aggregation of α-syn. It is also involved in the protection of neurons against oxidative stress and cell death [44]. It is worth mentioning that OxiDj-1 can be affected by other clinical pathologies that are undiagnosed at the time of taking the sample. Zhao et al. and Jang et al. demonstrated that plasma neural-derived DJ-1 in exosomes and OxiDJ-1 levels in urine, respectively, were increased in PD patients in comparison to healthy individuals. OxiDJ-1 showed significant differences only when ELISA was performed in comparison with a Western blot assay. Thus, OxiDJ-1 could be studied in larger cohorts as an achievable biomarker for PD detection in a near future [18,40].
Tau is a well-known protein aggregated in a hyperphosphorylated form in Alzheimer’s disease. EVs can mediate the spread of toxic forms of tau, helping it go from cell-to-cell and into different areas of the brain. Shi et al. pointed out the association between the levels of tau in neuron-derived exosomes in PD patients and the disease progression. Thus, CNS-plasma exosomal tau could be a marker for PD diagnosis [37].
3.3. RNAs and Micro RNAs
LncRNAs, as transcribed RNA molecules, are involved in the regulation of gene expression through epigenetic, translation, transcription and post-transcription mechanisms [52,53]. On the other hand, micro RNAs (miRNAs) are small noncoding RNA molecules whose length is between 19 to 22 nucleotides and are relevant because of their regulatory role in gene expression through post-transcriptional processes [33]. Some examples of dysregulated miRNAs analyzed in this work were miR-1, miR-153, miR-409-3p, miR-19b-3p, miR-10a-5p and let-7g-3p. These were identified as useful screeners to discern PD subjects from healthy controls [20]. So is the case of pure SEVs miR-34a-5p levels that were higher in PD patients even at the beginning stage of PD when the disease duration was less than 5 years [26]. Parallel to that are miR-24, miR-195 and miR-19b quantified in serum exosomes, which may be useful noninvasive biomarkers for the diagnosis of PD [31]. Caggiu et al. found that miR-155-5p was upregulated, with a significant role in the inflammatory response to α-syn in the CNS [54]. Unlike miR-155-5p, miR-146a was downregulated in the PD group, affecting the regulation of the monocyte inflammatory response. miR-125b-5p was found to be downregulated, with a direct impact on the expression of the BDNF-AS molecule, capable of promoting autophagy and apoptosis in MPTP-induced PD [55]. It is thought that lncRNAs may influence the expression of target miRNAs, therefore promoting the development of PD [56].
In this review, LncRNAs MSTRG.242001.1 and MSTRG.169261.1 were highly expressed among PD patients, while Lnc-MKRN2-42:1 could be linked to the severity of PD symptoms such as dyskinesia and dysarthria and correlated with MDS-UPDRS III score [13].
LncRNAs and miRNAs have been demonstrated to have implications in the regulation of genes that have been linked to familial PD, which comprises less than 10% of the cases, as follows: (i) Mutations in the SNCA, LRRK2, PARK2 (Parkin), PARK 6 (PINK1) and PARK7 (DJ-1) genes are involved in physiological functions such as kinase signaling, ubiquitin-mediated protein degradation and mitochondrial respiratory chain function [57]. (ii) The LRRK2 autosomal dominant mutation has been established as the most common cause of familial PD. Higher levels of exosomal Ser(P)-1292 LRRK2 have been linked to PD and the presence of nonmotor symptoms [19].; (iii) R1441C, Y1699C and G20192 LRRK2 mutations increase autophosphorylated LRRK2 protein levels at the Ser-1292 residue [19]. (iv) The R1441C mutation appears to promote the formation of abnormally large MVBs which release more exosomes, thus increasing the presence of toxic forms of α-syn in the extracellular space that promote disease spread [9]. (v) Ravanidis et al. [58] reviewed the literature regarding the dysregulation of several miRNAs in relation with their target proteins and their role in PD, finding evidence of the alterations in miR-205 (LRRK2), miR-7 and mir-153 (SNCA), miR-22 (GBA), mir-544 (DJ-1), among others. (vi) PINK1 participates in different processes regarding mitochondria, such as quality control and damage regulation and has been described as a causative gene in the pathogenesis of PD [52,53]. (vii) Unlike the mutations mentioned above, the GBA heterozygous mutation was initially linked to PD through clinical observations in populations such as the Ashkenazi Jews [59]. GBA transcribes the GCase protein whose main function is the degradation of glucocerebroside into ceramide and glucose, as well as cleaving glucosyl sphingosine. The presence of the GBA1 mutation reduces the enzymatic activity of GCase, which triggers the unfolded protein response and is linked to endoplasmic-reticulum-associated degradation [60]. (viii) The combination of GCase activity, plasma L1CAM exosomal Linc-POU3F3 and plasma L1CAM exosomal α-syn has been shown to be more reliable (AUC 0.824) in distinguishing PD vs. controls than each individually [28].
The increasing study of RNAs related to PD elucidates their stability characteristics and although numerous factors are involved in their expression profile, they can be correctly quantified by achievable, reproducible methods, leading these to be potential biomarkers for early diagnosis and disease stage [61].
3.4. Neural-Derived Extracellular Vesicles
Neural-derived extracellular vesicles (NDEVs) and their cargo could be the most representative image of the actual state of a neurodegenerative disease such as PD, hence the constant search for specific markers.
EVs are secreted by all types of cells including neurons. NDEVs seem to reflect the brain status in neurodegenerative diseases and also potentially mediate the seeding of pathogenic forms of prone proteins such as α-syn in healthy neurons [62].
Numerous researchers have isolated exosomes containing L1CAM from plasma of PD patients to analyze their cargo in order to find a reliable biomarker with a noninvasive sample. From levels of α-syn in plasma neuronal exosomes, tau, LncRNA, GCase activity and DJ-1 are useful for the quantification of neuron-derived exosomes and oligodendrocyte-derived via ELISA, which could help its use as a biomarker for the diagnosis and progression of PD [18,28,32,37,39]. However, recently Norman et al. evaluated the use of L1CAM for the isolation of NDEV in plasma and CSF and advised against its use as a reliable marker due to its behavior as a soluble protein and not as a specific marker for EVs, explaining its nonspecific binding with soluble proteins such as α-syn [63]. In addition, the MISEV 2018 (Minimal Information for Studies of EVs 2018 guidelines) does not propose any biological marker to differentiate subtypes of EVs or to know their cell of origin [7].
3.5. Other Potential Biomarkers
The identification of specific proteins that are involved in PD pathogenesis has brought a new insight in the search for new biomarkers for early PD diagnosis and prognosis. Proteomic technology allows the study of the protein signature of EVs whether in normal conditions or in pathologic scenarios such as PD via different biofluids.
The expression of several proteins in EVs has been studied and analyzed. Jiang et al. found the C1q complex decreased PD patients’ serum exosomes. C1q is the recognition molecule key which contributes to the innate immune defense and regulates the adaptive immune response for the neuroprotection of the CNS and mediation of the formation of central synapses. Although linked to PD, it is still very difficult to elucidate the total role of the expression of C1q in the development of this disease [17,64].
As mentioned above, proteomics of EV derived from the CNS, erythrocytes or different biofluids have thrown hints for the study of new proteins that are involved in neurological mechanisms.
Lamontagne-Proulx et al. [22] performed a proteomic analysis of EVs derived from erythrocytes. In total, 8 out of the 818 proteins identified in the proteome of EVs had expressions that were significantly different in PD patients with various stages. Among these, QDPR is a key catalyzer for the recycling of BH4 (tetrahydrobiopterin), an essential cofactor in the biosynthesis of serotonin and precursors of L-dopa and 5-hydroxy-L-tryptophan (5-HTP) [65]. On the other hand, genetic variations in the USP24 protein coding gene, a member of the ubiquitin-specific protease family, is associated with the risk for late-onset PD [66].
Gualerzi et al. [23] studied the biochemistry of EVs through Raman spectroscopy and found there were biochemical differences between circulating EVs involving proteins, lipids and saccharides, which made it possible to discern between PD patients from HC with an accuracy of 71%. These findings could be useful to evaluate the effectiveness of rehabilitation and pharmacological treatments of PD in the future.
Blood-derived exosomal clusterin, complement C1r subcomponent, afamin, angiotensinogen variant, apolipoprotein D (ApoD), gelsolin and PEDF were progressively upregulated from mild to severe PD [17]. It is of major interest to analyze the molecular function of each protein and their interaction with upregulated pathways related to PD disease.
Apolipoprotein A1 (Apo A1), clusterin, complement C1r subcomponent and fibrinogen gamma chain exosomal expression levels in plasma of PD subjects may serve as a biomarker for the diagnosis of PD while Apo A1 could be of use in the future to measure the progression of the disease [27,35].
3.6. Neuroinflammation and Neurodegeneration
Neuroinflammation has been recognized as a key mediator in PD progression. There appears to be an alteration in the normal morphology of glial cells, including astrocytes and microglia, as well as an increase in inflammatory mediators in the parenchyma. This glial reaction is thought to happen due to neuronal death or dysfunction [67,68]. Exosomes containing pathologic forms of α-syn may activate microglia cells, promoting the accumulation and transmission of α-syn, while contributing to neuroinflammation by releasing inflammatory mediators [38].
The autophagy–lysosomal pathway (ALP) is capable of degrading aggregated misfolded proteins, such as α-syn, which in pathologic forms cause a neurodegenerative event when taken by microglia [69]. The dysfunction of ALP may come from an activation of microglia cells by exosomes containing pathologic forms of α-syn, leading to an accumulation and transmission of this protein into the system [38].
In addition to the mechanisms mentioned above, it has been hypothesized that the elevation of phosphorylated Ser-1292 in PD patients with advanced cognitive impairment is related to higher levels of plasma C-reactive protein levels as well as other inflammatory cytokines that eventually exacerbate neurodegeneration [5,14].
4. Conclusions
The interest in EVs has recently increased as it has been demonstrated that they play an important role in the multiple pathways that lead to neurodegenerative disorders such as PD. Their neuroprotective and neurotoxic functions have been meticulously studied. They intervene in the clearing of misfolded proteins by releasing detoxifying substances but also participate in the accumulation and transport of the origin cell cargo to specific cells, including misfolded proteins, nucleic acids and other cell constituents [8,12].
It is worth emphasizing that of all studies in this review, the largest total sample size was of 303 participants [37]. Future research projects should be performed in larger and independent cohorts with longitudinal approaches.
EVs isolation methods were varied, using centrifugation, differential centrifugation, ultracentrifugation, precipitation, or a combination of these, magnetic bead separation, size exclusion chromatography and fluorescence-activated cell sorting. Subsequent validation of the isolated exosomes in these studies was done using a variety of techniques such as Western blot, ELISA, TEM or NTA. The variation in the results found in this field clearly manifests the urgent need of more standardized protocols in EVs experimentation but also manifests the fantastic advances that have emerged with it.
Current data shed light on exosomal proteins, such as α-syn and DJ-1, as well as miRNAs and lncRNAs, as potential biomarkers that could be used to create standardized protocols for early PD diagnosis as well as tools to evaluate the severity and progression of the disease. Meaningful contributions such as the proteomic analysis of EVs and the study of its biochemistry as a fingerprint signature have marked a path for new biomarkers in this complex neurodegenerative disease. However, the future should be focused on finding reliable surface markers, using standardized procedures for the correct isolation of NDEVs from noninvasive biological samples such as saliva, which has demonstrated a good performance in early PD diagnosis, through the quantification of phospho-α-syn from neural-derived exosomes. Knowing the origin of EVs remains a challenge in research hence more studies are needed so we can resolve the puzzle correctly and understand every piece and pathway in PD.
In fact, miRNAs isolated from exosomal CSF (miR-1, miR-153, miR-409-3p, miR-19b-3p, miR-10a-5p and let-7g-3p) have shown great performance in distinguishing between PD patients and HC subjects. Even though it is an invasive procedure, lumbar puncture remains the most approachable way to study CNS. However, miR-24, miR-195 and miR-19b, all measured together in serum exosomes of PD patients, presented an AUC of 0.946 (95% CI, 0.910–0.981), the specificity was 90.0% and the sensitivity was 85.3%, so these may be useful noninvasive biomarkers for the diagnosis of PD. Future research is needed to identify the biological function of miR-24, miR-195 and miR-19b but for now, they shed light into this journey and create a new basis for new approaches.
The future should be focused on finding future biomarkers that not only allow us to make an early diagnosis of the disease, but also to distinguish PD from other neurodegenerative disorders.
Author Contributions
J.V., J.M.S., M.F., A.M. and J.F.M.-T. were responsible for the conceptualization, design, developed methodology, validation, data curation, and writing—review and editing; J.V., M.F. and J.M.S. were primarily responsible for the investigation and writing—original draft preparation; J.V., J.M.S., M.F., A.M. and J.F.M.-T. were responsible for the review. All authors have read and agreed to the published version of the manuscript.
Funding
Supported by the Agencia Estatal de Investigación, Ministerio de Ciencia e Innovación, Spain (grant number PID2019-105713GB-I00/AEI/10.13039/501100011033), and Conselleria d’Educació, Cultura i Esports, Generalitat Valenciana, Valencia, Spain (grant PROMETEO/2020/071).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kalia, L.V.; Kalia, S.K.; Lang, A.E. Disease-modifying strategies for Parkinson’s disease. Mov. Disord. 2015, 30, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Vandendriessche, C.; Bruggeman, A.; van Cauwenberghe, C.; Vandenbroucke, R.E. Extracellular vesicles in Alzheimer’s and Parkinson’s Disease: Small entities with large consequences. Cells 2020, 9, 2485. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Kalia, S.K. α-Synuclein and Lewy pathology in Parkinson’s disease. Curr. Opin. Neurol. 2015, 28, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.U.; Akram, M.; Daniyal, M.; Zainab, R. Awareness and current knowledge of Parkinson’s Disease: A neurodegenerative disorder. Int. J. Neurosci. 2019, 129, 55–93. [Google Scholar] [CrossRef]
- Russo, I.; Bubacco, L.; Greggio, E. Exosomes-associated neurodegeneration and progression of Parkinson’s disease. Am. J. Neurodegener. Dis. 2012, 1, 217. [Google Scholar]
- Zhang, T.M.; Yu, S.Y.; Guo, P.; Du, Y.; Hu, Y.; Piao, Y.S.; Zuo, L.J.; Lian, T.H.; Wang, R.D.; Yu, Q.J.; et al. Nonmotor symptoms in patients with Parkinson disease: A cross-sectional observational study. Medicine 2016, 95, e5400. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Porro, C.; Panaro, M.A.; Lofrumento, D.D.; Hasalla, E.; Trotta, T. The multiple roles of exosomes in Parkinson’s disease: An overview. Immunopharmacol. Immunotoxicol. 2019, 41, 469–476. [Google Scholar] [CrossRef]
- De la Torre Gomez, C.; Goreham, R.V.; Bech Serra, J.J.; Nann, T.; Kussmann, M. “Exosomics”-A review of biophysics, biology and biochemistry of exosomes with a focus on human breast milk. Front. Genet. 2018, 9, 92. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.F. Extracellular vesicles and neurodegenerative diseases. J. Neurosci. 2019, 39, 9269–9273. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Han, C.L.; Wang, K.L.; Sui, Y.P.; Li, Z.B.; Chen, N.; Fan, S.Y.; Shimabukuro, M.; Wang, F.; Meng, F.G. Integrated analysis of exosomal LncRNA and MRNA expression profiles reveals the involvement of Lnc-MKRN2-42:1 in the pathogenesis of Parkinson’s disease. CNS Neurosci. Ther. 2020, 26, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Cerri, S.; Ghezzi, C.; Sampieri, M.; Siani, F.; Avenali, M.; Dornini, G.; Zangaglia, R.; Minafra, B.; Blandini, F. The Exosomal/Total α-Synuclein ratio in plasma is associated with glucocerebrosidase activity and correlates with measures of disease severity in PD patients. Front. Cell. Neurosci. 2018, 12, 125. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Leng, B.; Sun, H.; Zhao, J.; Liu, Y.; Shen, T.; Liu, W.; Liu, X.; Tan, M.; Li, F.; Zhang, J.; et al. Plasma exosomal prion protein levels are correlated with cognitive decline in PD patients. Neurosci. Lett. 2020, 723, 134866. [Google Scholar] [CrossRef]
- Jiang, R.; Rong, C.; Ke, R.; Meng, S.; Yan, X.; Ke, H.; Wu, S.; Azim, A. Differential proteomic analysis of serum exosomes reveals alterations in progression of Parkinson disease. Medicine 2019, 98, e17478. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Chen, Z.T.; Zhou, R.L.; Zhang, X.; Ye, Q.Y.; Wang, Y.Z. Increased DJ-1 and α-Synuclein in plasma neural-derived exosomes as potential markers for Parkinson’s disease. Front. Aging Neurosci. 2019, 10, 438. [Google Scholar] [CrossRef]
- Fraser, K.B.; Rawlins, A.B.; Clark, R.G.; Alcalay, R.N.; Standaert, D.G.; Liu, N.; West, A.B. Ser(P)-1292 LRRK2 in urinary exosomes is elevated in idiopathic Parkinson’s disease. Mov. Disord. 2016, 31, 1543–1550. [Google Scholar] [CrossRef]
- Gui, Y.X.; Liu, H.; Zhang, L.S.; Lv, W.; Hu, X.Y. Altered MicroRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6, 37043–37053. [Google Scholar] [CrossRef]
- Vacchi, E.; Burrello, J.; di Silvestre, D.; Burrello, A.; Bolis, S.; Mauri, P.; Vassalli, G.; Cereda, C.W.; Farina, C.; Barile, L.; et al. Immune profiling of plasma-derived extracellular vesicles identifies Parkinson disease. Neurol-Neuroimmunol. 2020, 7, e866. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne-Proulx, J.; St-Amour, I.; Labib, R.; Pilon, J.; Denis, H.L.; Cloutier, N.; Roux-Dalvai, F.; Vincent, A.T.; Mason, S.L.; Williams-Gray, C.; et al. Portrait of blood-derived extracellular vesicles in patients with Parkinson’s disease. Neurobiol. Dis. 2019, 124, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Gualerzi, A.; Picciolini, S.; Carlomagno, C.; Terenzi, F.; Ramat, S.; Sorbi, S.; Bedoni, M. Raman profiling of circulating extracellular vesicles for the stratification of Parkinson’s patients. Nanomedicine 2019, 22, 102097. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Wu, Y.; Liu, G.; Jiang, Y.; Wang, X.; Wang, Z.; Zhang, J.; Feng, T. α-Synuclein in salivary extracellular vesicles as a potential biomarker of Parkinson’s disease. Neurosci. Lett. 2019, 696, 114–120. [Google Scholar] [CrossRef]
- Niu, M.; Li, Y.; Li, G.; Zhou, L.; Luo, N.; Yao, M.; Kang, W.; Liu, J. A longitudinal study on α-synuclein in plasma neuronal exosomes as a biomarker for Parkinson’s disease development and progression. Eur. J. Neurol. 2020, 27, 967–974. [Google Scholar] [CrossRef]
- Grossi, I.; Radeghieri, A.; Paolini, L.; Porrini, V.; Pilotto, A.; Padovani, A.; Marengoni, A.; Barbon, A.; Bellucci, A.; Pizzi, M.; et al. MicroRNA-34a-5p expression in the plasma and in its extracellular vesicle fractions in subjects with Parkinson’s disease: An exploratory study. Int. J. Mol. Med. 2021, 47, 533–546. [Google Scholar] [CrossRef]
- Ohmichi, T.; Mitsuhashi, M.; Tatebe, H.; Kasai, T.; Ali El-Agnaf, O.M.; Tokuda, T. Quantification of brain-derived extracellular vesicles in plasma as a biomarker to diagnose Parkinson’s and related diseases. Parkinsonism Relat. Disord. 2019, 61, 82–87. [Google Scholar] [CrossRef]
- Zou, J.; Guo, Y.; Wei, L.; Yu, F.; Yu, B.; Xu, A. Long noncoding RNA POU3F3 and α-synuclein in plasma L1CAM exosomes combined with β-glucocerebrosidase activity: Potential predictors of Parkinson’s disease. Neurotherapeutics 2020, 17, 1104–1119. [Google Scholar] [CrossRef]
- Chung, C.C.; Chan, L.; Chen, J.H.; Hung, Y.C.; Hong, C.T. Plasma extracellular vesicle α-synuclein level in patients with Parkinson’s disease. Biomolecules 2021, 11, 744. [Google Scholar] [CrossRef]
- Stuendl, A.; Kunadt, M.; Kruse, N.; Bartels, C.; Moebius, W.; Danzer, K.M.; Mollenhauer, B.; Schneider, A. Induction of α-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain 2016, 139, 481–494. [Google Scholar] [CrossRef]
- Cao, X.Y.; Lu, J.M.; Zhao, Z.Q.; Li, M.C.; Lu, T.; An, X.S.; Xue, L.J. MicroRNA biomarkers of Parkinson’s disease in serum exosome-like microvesicles. Neurosci. Lett. 2017, 644, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Jiang, C.; Tofaris, G.K.; Davis, J.J. Facile Impedimetric analysis of neuronal exosome markers in Parkinson’s disease diagnostics. Anal. Chem. 2020, 92, 13647–13651. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.F.; Qu, M.W.; Li, G.C.; Zhang, F.B.; Rui, H.C. Circulating exosomal MiRNAs as diagnostic biomarkers in Parkinson’s disease. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5278–5283. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xie, Z.; Zhang, X.; Mao, J.; Wang, M.; Wei, S.; Fu, Y.; Zheng, H.; He, Y.; Chen, H.; et al. Investigation of α-synuclein species in plasma exosomes and the oligomeric and phosphorylated α-synuclein as potential peripheral biomarker of Parkinson’s disease. Neuroscience 2021, 469, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Kojima, M.; Kurosawa, T.; Sasaki, R.; Ichihara, S.; Hiraku, Y.; Tomimoto, H.; Murata, M.; Oikawa, S. Proteomic profiling of exosomal proteins for blood-based biomarkers in Parkinson’s disease. Neuroscience 2018, 392, 121–128. [Google Scholar] [CrossRef]
- Rani, K.; Mukherjee, R.; Singh, E.; Kumar, S.; Sharma, V.; Vishwakarma, P.; Bharti, P.S.; Nikolajeff, F.; Dinda, A.K.; Goyal, V.; et al. Neuronal exosomes in saliva of Parkinson’s disease patients: A pilot study. Parkinsonism Relat. Disord. 2019, 67, 21–23. [Google Scholar] [CrossRef]
- Shi, M.; Kovac, A.; Korff, A.; Cook, T.J.; Ginghina, C.; Bullock, K.M.; Yang, L.; Stewart, T.; Zheng, D.; Aro, P.; et al. CNS Tau efflux via exosomes is likely increased in Parkinson’s disease but not in Alzheimer’s disease. Alzheimer Dement. 2016, 12, 1125–1131. [Google Scholar] [CrossRef]
- Xia, Y.; Zhang, G.; Han, C.; Ma, K.; Guo, X.; Wan, F.; Kou, L.; Yin, S.; Liu, L.; Huang, J.; et al. Microglia as modulators of exosomal alpha-synuclein transmission. Cell Death Dis. 2019, 10, 174. [Google Scholar] [CrossRef]
- Si, X.; Tian, J.; Chen, Y.; Yan, Y.; Pu, J.; Zhang, B. Central nervous system-derived exosomal alpha-synuclein in serum may be a biomarker in Parkinson’s disease. Neuroscience 2019, 413, 308–316. [Google Scholar] [CrossRef]
- Jang, J.; Jeong, S.; Lee, S.I.; Seol, W.; Seo, H.; Son, I.; Ho, D.H. Oxidized DJ-1 levels in urine samples as a putative biomarker for Parkinson’s disease. Parkinsons Dis. 2018, 2018, 1241757. [Google Scholar] [CrossRef]
- Lucien, F.; Benarroch, E.E.; Mullan, A.; Ali, F.; Boeve, B.F.; Mielke, M.M.; Petersen, R.C.; Kim, Y.; Stang, C.; Camerucci, E.; et al. Poly (ADP-Ribose) and α-synuclein extracellular vesicles in patients with Parkinson disease: A possible biomarker of disease severity. PLoS ONE 2022, 17, e0264446. [Google Scholar] [CrossRef] [PubMed]
- Kluge, A.; Bunk, J.; Schaeffer, E.; Drobny, A.; Xiang, W.; Knacke, H.; Bub, S.; Lückstädt, W.; Arnold, P.; Lucius, R.; et al. Detection of neuron-derived pathological α-synuclein in blood. Brain 2022, 145, 3058–3071. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.M.; de Miranda, B.; Sanders, L.H. Alpha-synuclein: Pathology, mitochondrial dysfunction and neuroinflammation in Parkinson’s DISEase. Neurobiol. Dis. 2018, 109, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Delamarre, A.; Meissner, W.G. Epidemiology, Environmental risk factors and genetics of Parkinson’s disease. Presse Medicale 2017, 46, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Sun, T.; An, J.; Wen, L.; Liu, F.; Bu, Z.; Cui, Y.; Feng, J. Potential roles of exosomes in Parkinson’s Disease: From pathogenesis, diagnosis, and treatment to prognosis. Front. Cell. Dev. Biol. 2020, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Danzer, K.M.; Kranich, L.R.; Ruf, W.P.; Cagsal-Getkin, O.; Winslow, A.R.; Zhu, L.; Vanderburg, C.R.; McLean, P.J. Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol. Neurodegener. 2012, 7, 42. [Google Scholar] [CrossRef]
- Li, J.Y.; Englund, E.; Holton, J.L.; Soulet, D.; Hagell, P.; Lees, A.J.; Lashley, T.; Quinn, N.P.; Rehncrona, S.; Björklund, A.; et al. Lewy bodies in grafted neurons in subjects with Parkinson’s Disease suggest host-to-graft disease propagation. Nat. Med. 2008, 14, 501–503. [Google Scholar] [CrossRef]
- Emmanouilidou, E.; Melachroinou, K.; Roumeliotis, T.; Garbis, S.D.; Ntzouni, M.; Margaritis, L.H.; Stefanis, L.; Vekrellis, K. Cell-Produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J. Neurosci. 2010, 30, 6838–6851. [Google Scholar] [CrossRef]
- Stefanis, L.; Emmanouilidou, E.; Pantazopoulou, M.; Kirik, D.; Vekrellis, K.; Tofaris, G.K. How Is alpha-synuclein cleared from the cell? J. Neurochem. 2019, 150, 577–590. [Google Scholar] [CrossRef]
- Kenwrick, S.; Watkins, A.; de Angelis, E. Neural cell recognition molecule L1: Relating biological complexity to human disease mutations. Hum. Mol. Genet. 2000, 9, 879–886. [Google Scholar] [CrossRef]
- Mollenhauer, B.; Batrla, R.; El-Agnaf, O.; Galasko, D.R.; Lashuel, H.A.; Merchant, K.M.; Shaw, L.M.; Selkoe, D.J.; Umek, R.; Vanderstichele, H.; et al. A user’s guide for a-synuclein biomarker studies in biological fluids: Perianalytical considerations. Mov. Dis. 2017, 32, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Elkouris, M.; Kouroupi, G.; Vourvoukelis, A.; Papagiannakis, N.; Kaltezioti, V.; Matsas, R.; Stefanis, L.; Xilouri, M.; Politis, P.K. Long non-coding rnas associated with neurodegeneration-linked genes are reduced in Parkinson’s disease patients. Front. Cell. Neurosci. 2019, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Riva, P.; Ratti, A.; Venturin, M. The long non-coding RNAs in neurodegenerative diseases: Novel mechanisms of pathogenesis. Curr. Alzheimer Res. 2016, 13, 1219–1231. [Google Scholar] [CrossRef]
- Caggiu, E.; Paulus, K.; Mameli, G.; Arru, G.; Sechi, G.P.; Sechi, L.A. Differential expression of MiRNA 155 and MiRNA 146a in Parkinson’s disease patients. eNeurologicalSci 2018, 13, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhao, X.; Lu, K.; Cheng, G. LncRNA BDNF-AS Promotes autophagy and apoptosis in MPTP-induced Parkinson’s disease via ablating MicroRNA-125b-5p. Brain Res. Bull. 2020, 157, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Huang, X.; Li, R. LncRNA MALAT1/MiR-205-5p axis regulates MPP+-induced cell apoptosis in MN9D cells by directly targeting LRRK2. Am. J. Transl. Res. 2018, 10, 563. [Google Scholar] [PubMed]
- Calabrese, V.; Santoro, A.; Monti, D.; Crupi, R.; di Paola, R.; Latteri, S.; Cuzzocrea, S.; Zappia, M.; Giordano, J.; Calabrese, E.J.; et al. Aging and Parkinson’s Disease: Inflammaging, neuroinflammation and biological remodeling as key factors in pathogenesis. Free Radic. Biol. Med. 2018, 115, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Ravanidis, S.; Bougea, A.; Papagiannakis, N.; Maniati, M.; Koros, C.; Simitsi, A.M.; Bozi, M.; Pachi, I.; Stamelou, M.; Paraskevas, G.P.; et al. Circulating brain-enriched MicroRNAs for detection and discrimination of idiopathic and genetic Parkinson’s disease. Mov. Disord. 2020, 35, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Gan-Or, Z.; Liong, C.; Alcalay, R.N. GBA-associated Parkinson’s disease and other synucleinopathies. Curr. Neurol. Neurosci. Rep. 2017, 18, 1–10. [Google Scholar] [CrossRef]
- Schapira, A.H.V. Glucocerebrosidase and Parkinson disease: Recent advances. Mol. Cell. Neurosci. 2015, 66, 37–42. [Google Scholar] [CrossRef]
- Manna, I.; Quattrone, A.; de Benedittis, S.; Iaccino, E.; Quattrone, A. Roles of non-coding RNAs as novel diagnostic biomarkers in Parkinson’s disease. J. Parkinsons Dis. 2021, 11, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Ngolab, J.; Trinh, I.; Rockenstein, E.; Mante, M.; Florio, J.; Trejo, M.; Masliah, D.; Adame, A.; Masliah, E.; Rissman, R.A. Brain-derived exosomes from dementia with Lewy bodies propagate α-synuclein pathology. Acta Neuropathol. Commun. 2017, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.; Ter-Ovanesyan, D.; Trieu, W.; Lazarovits, R.; Kowal, E.J.K.; Lee, J.H.; Chen-Plotkin, A.S.; Regev, A.; Church, G.M.; Walt, D.R. L1CAM is not associated with extracellular vesicles in human cerebrospinal fluid or plasma. Nat. Methods 2021, 18, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, F.S.; Dijkstra, D.J.; van Kooten, C.; Gelderman, K.A.; Trouw, L.A. Circulating C1q levels in health and disease, more than just a biomarker. Mol. Immunol. 2021, 140, 206–216. [Google Scholar] [CrossRef]
- Breuer, M.; Guglielmi, L.; Zielonka, M.; Hemberger, V.; Kölker, S.; Okun, J.G.; Hoffmann, G.F.; Carl, M.; Sauer, S.W.; Opladen, T. QDPR homologues in danio rerio regulate melanin synthesis, early gliogenesis, and glutamine homeostasis. PLoS ONE 2019, 14, e0215162. [Google Scholar] [CrossRef]
- Li, Y.; Schrodi, S.; Rowland, C.; Tacey, K.; Catanese, J.; Grupe, A. Genetic evidence for ubiquitin-specific proteases USP24 and USP40 as candidate genes for late-onset Parkinson disease. Hum. Mut. 2006, 27, 1017–1023. [Google Scholar] [CrossRef]
- Ransohoff, R.M. How neuroinflammation contributes to neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef]
- Console, L.; Scalise, M.; Indiveri, C. Exosomes in inflammation and role as biomarkers. Clin. Chim. Acta 2019, 488, 165–171. [Google Scholar] [CrossRef]
- Finkbeiner, S. The autophagy lysosomal pathway and neurodegeneration. Cold Spring Harb. Perspect. Biol. 2020, 12, a033993. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).