
IL3 Has a Detrimental Effect on Hematopoietic Stem Cell Self-Renewal in Transplantation Settings

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ex Vivo Expansion of Hematopoietic Stem Cells in Cytokine-Supplemented Medium
2.2. IL3 Reduces Human Engraftment in NSG Mice
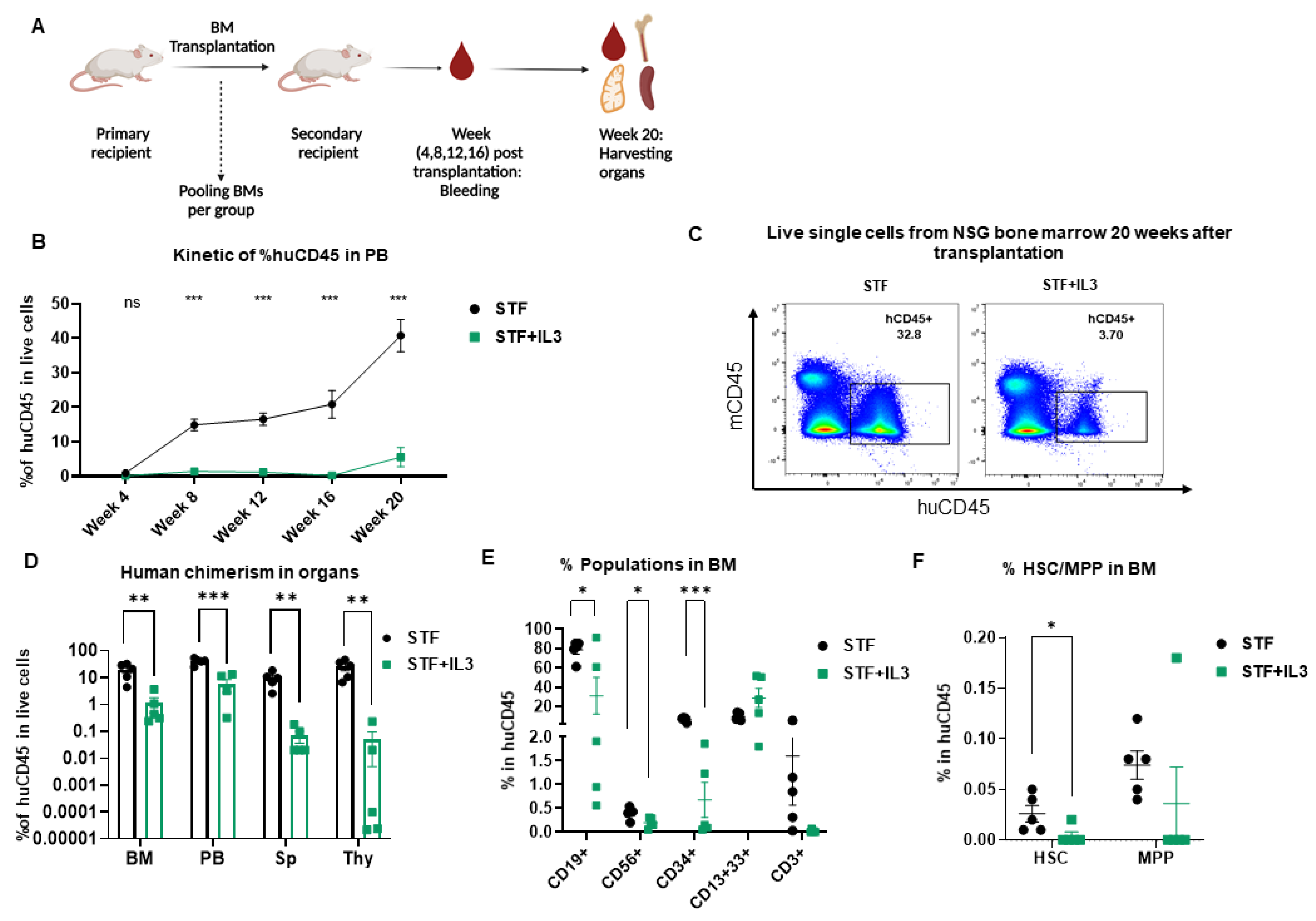
2.3. IL3 Reduces the Repopulation Capacity of Human Hematopoietic Stem Cells in Mice
3. Discussion
4. Materials and Methods
4.1. Human Cells and CD34+ Enrichment
4.2. CD34+ Cell Culture
4.3. Mice
4.4. Primary and Secondary Transplantations into NSG Mice
4.5. Flowcytometry
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Körbling, M.; Estrov, Z. Adult Stem Cells for Tissue Repair—A New Therapeutic Concept? N. Engl. J. Med. 2003, 349, 570–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copelan, E.A. Hematopoietic Stem-Cell Transplantation. N. Engl. J. Med. 2006, 354, 1813–1826. [Google Scholar] [CrossRef]
- Kondo, M.; Wagers, A.J.; Manz, M.G.; Prohaska, S.S.; Scherer, D.C.; Beilhack, G.F.; Shizuru, J.A.; Weissman, I.L. Biology of Hematopoietic Stem Cells and Progenitors: Implications for Clinical Application. Annu. Rev. Immunol. 2003, 21, 759–806. [Google Scholar] [CrossRef]
- Morrison, S.J.; Kimble, J. Asymmetric and symmetric stem-cell divisions in development and cancer. Nature 2006, 441, 1068–1074. [Google Scholar] [CrossRef]
- Bhatia, M.; Bonnet, D.; Kapp, U.; Wang, J.; Murdoch, B.; Dick, J.E. Quantitative Analysis Reveals Expansion of Human Hematopoietic Repopulating Cells After Short-term Ex Vivo Culture. J. Exp. Med. 1997, 186, 619–624. [Google Scholar] [CrossRef]
- Shpall, E.J.; Quinones, R.; Giller, R.; Zeng, C.; Baron, E.A.; Jones, R.B.; Bearman, I.S.; Nieto, Y.; Freed, B.; Madinger, N.; et al. Transplantation of ex vivo expanded cord blood. Biol. Blood Marrow Transplant. 2002, 8, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peled, T.; Mandel, J.; Goudsmid, R.; Landor, C.; Hasson, N.; Harati, D.; Austin, M.; Hasson, A.; Fibach, E.; Shpall, E.; et al. Pre-clinical development of cord blood-derived progenitor cell graft expanded ex vivo with cytokines and the polyamine copper chelator tetraethylenepentamine. Cytotherapy 2004, 6, 344–355. [Google Scholar] [CrossRef] [PubMed]
- de Lima, M.; McMannis, J.; Gee, A.; Komanduri, K.; Couriel, D.; Andersson, B.S.; Hosing, C.; Khouri, I.; Jones, R.; Champlin, R.; et al. Transplantation of ex vivo expanded cord blood cells using the copper chelator tetraethylenepentamine: A phase I/II clinical trial. Bone Marrow Transplant. 2008, 41, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalf, D. Hematopoietic cytokines. Blood 2008, 111, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmeister, C.C.; Zhang, J.; Knight, K.L.; Le, P.; Stiff, P.J. Ex vivo expansion of umbilical cord blood stem cells for transplantation: Growing knowledge from the hematopoietic niche. Bone Marrow Transplant. 2006, 39, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.G.; van Eggermond, M.; van Roon, L.; Vloemans, S.A.; Cordes, M.; Schambach, A.; Rothe, M.; Berghuis, D.; Lagresle-Peyrou, C.; Cavazzana, M.; et al. Successful Preclinical Development of Gene Therapy for Recombinase-Activating Gene-1-Deficient SCID. Mol. Ther. Methods Clin. Dev. 2020, 17, 666–682. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Smogorzewska, E.; Hannum, C.; Crooks, G. Flt3 ligand induces proliferation of quiescent human bone marrow CD34+CD38- cells and maintains progenitor cells in vitro. Blood 1996, 87, 3563–3570. [Google Scholar] [CrossRef]
- Petzer, A.L.; Zandstra, P.W.; Piret, J.M.; Eaves, C.J. Differential cytokine effects on primitive (CD34+CD38-) human hematopoietic cells: Novel responses to Flt3-ligand and thrombopoietin. J. Exp. Med. 1996, 183, 2551–2558. [Google Scholar] [CrossRef] [Green Version]
- McKenna, H.; De Vries, P.; Brasel, K.; Lyman, S.; Williams, D. Effect of flt3 ligand on the ex vivo expansion of human CD34+ hematopoietic progenitor cells. Blood 1995, 86, 3413–3420. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, R.; Tong, J.; Brandt, J.; Traycoff, C.; Bruno, E.; Srour, E.F.; Gordon, M.S.; Mcguire, B.W.; Mcniece, I. The in vitro and in vivo effects of stem cell factor on human hematopoiesis. Stem Cells 1996, 11, 76–82. [Google Scholar] [CrossRef]
- Yonemura, Y.; Ku, H.; Hirayama, F.; Souza, L.M.; Ogawa, M. Interleukin 3 or interleukin 1 abrogates the reconstituting ability of hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 1996, 93, 4040–4044. [Google Scholar] [CrossRef] [Green Version]
- Gammaitoni, L.; Bruno, S.; Sanavio, F.; Gunetti, M.; Kollet, O.; Cavalloni, G.; Falda, M.; Fagioli, F.; Lapidot, T.; Aglietta, M.; et al. Ex vivo expansion of human adult stem cells capable of primary and secondary hemopoietic reconstitution. Exp. Hematol. 2003, 31, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Knobel, K.M.; McNally, A.M.; Berson, E.A.; Rood, D.; Chen, K.; Kilinski, L.; Tran, K.; Okarma, T.B.; Lebkowski, J.S. Long-term reconstitution of mice after ex vivo expansion of bone marrow cells: Differential activity of cultured bone marrow and enriched stem cell populations. Exp. Hematol. 1994, 22, 1227–1235. [Google Scholar]
- Ohmori, K.; Luo, Y.; Jia, Y.; Nishida, J.; Wang, Z.; Bunting, K.D.; Wang, D.; Huang, H. IL-3 Induces Basophil Expansion In Vivo by Directing Granulocyte-Monocyte Progenitors to Differentiate into Basophil Lineage-Restricted Progenitors in the Bone Marrow and by Increasing the Number of Basophil/Mast Cell Progenitors in the Spleen. J. Immunol. 2009, 182, 2835–2841. [Google Scholar] [CrossRef] [Green Version]
- Piacibello, W.; Gammaitoni, L.; Bruno, S.; Gunetti, M.; Fagioli, F.; Cavalloni, G.; Aglietta, M. Negative Influence of IL3 on the Expansion of Human Cord Blood In Vivo Long-Term Repopulating Stem Cells. J. Hematotherapy 2000, 9, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Peters, S.O.; Kittler, E.L.; Ramshaw, H.S.; Quesenberry, P.J. Ex vivo expansion of murine marrow cells with interleukin-3 (IL-3), IL-6, IL-11, and stem cell factor leads to impaired engraftment in irradiated hosts. Blood 1996, 87, 30–37. [Google Scholar] [CrossRef]
- Piacibello, W.; Sanavio, F.; Garetto, L.; Severino, A.; Dané, A.; Gammaitoni, L.; Aglietta, M. Differential growth factor requirement of primitive cord blood hematopoietic stem cell for self-renewal and amplification vs proliferation and differentiation. Leukemia 1998, 12, 718–727. [Google Scholar] [CrossRef] [Green Version]
- Tajer, P.; Pike-Overzet, K.; Arias, S.; Havenga, M.; Staal, F.J. Ex Vivo Expansion of Hematopoietic Stem Cells for Therapeutic Purposes: Lessons from Development and the Niche. Cells 2019, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef]
- Haylock, D.; To, L.; Dowse, T.; Juttner, C.; Simmons, P. Ex vivo expansion and maturation of peripheral blood CD34+ cells into the myeloid lineage. Blood 1992, 80, 1405–1412. [Google Scholar] [CrossRef]
- Broxmeyer, E.H.; Hangoc, G.; Cooper, S.; Ribeiro, R.C.; Graves, V.; Yoder, M.; Wagner, J.; Vadhan-Raj, S.; Benninger, L.; Rubinstein, P. Growth characteristics and expansion of human umbilical cord blood and estimation of its potential for transplantation in adults. Proc. Natl. Acad. Sci. USA 1992, 89, 4109–4113. [Google Scholar] [CrossRef] [Green Version]
- Fares, I.; Chagraoui, J.; Gareau, Y.; Gingras, S.; Ruel, R.; Mayotte, N.; Csaszar, E.; Knapp, D.J.; Miller, P.; Ngom, M.; et al. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science 2014, 345, 1509–1512. [Google Scholar] [CrossRef] [Green Version]
- Suda, T.; Suda, J.; Ogawa, M.; Ihle, J.N. Permissive role of interleukin 3 (IL-3) in proliferation and differentiation of multipotential hemopoietic progenitors in culture. J. Cell. Physiol. 1985, 124, 182–190. [Google Scholar] [CrossRef]
- Ihle, J.N.; Keller, J.; Oroszlan, S.; Henderson, E.L.; Copeland, T.D.; Fitch, F.; Prystowsky, M.B.; Goldwasser, E.; Schrader, J.W.; Palaszynski, E.; et al. Biologic properties of homogeneous interleukin 3. I. Demonstration of WEHI-3 growth factor activity, mast cell growth factor activity, p cell-stimulating factor activity, colony-stimulating factor activity, and histamine-producing cell-stimulating factor activity. J. Immunol. 1983, 131, 282–287. [Google Scholar]
- Matsunaga, T.; Hirayama, F.; Yonemura, Y.; Murray, R.; Ogawa, M. Negative Regulation by Interleukin-3 (IL-3) of Mouse Early B-Cell Progenitors and Stem Cells in Culture: Transduction of the Negative Signals by βc and βIL-3 Proteins of IL-3 Receptor and Absence of Negative Regulation by Granulocyte-Macrophage Colony-Stimulating Factor. Blood 1998, 92, 901–907. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajer, P.; Canté-Barrett, K.; Naber, B.A.E.; Vloemans, S.A.; van Eggermond, M.C.J.A.; van der Hoorn, M.-L.; Pike-Overzet, K.; Staal, F.J.T. IL3 Has a Detrimental Effect on Hematopoietic Stem Cell Self-Renewal in Transplantation Settings. Int. J. Mol. Sci. 2022, 23, 12736. https://doi.org/10.3390/ijms232112736
Tajer P, Canté-Barrett K, Naber BAE, Vloemans SA, van Eggermond MCJA, van der Hoorn M-L, Pike-Overzet K, Staal FJT. IL3 Has a Detrimental Effect on Hematopoietic Stem Cell Self-Renewal in Transplantation Settings. International Journal of Molecular Sciences. 2022; 23(21):12736. https://doi.org/10.3390/ijms232112736
Chicago/Turabian StyleTajer, Parisa, Kirsten Canté-Barrett, Brigitta A. E. Naber, Sandra A. Vloemans, Marja C. J. A. van Eggermond, Marie-Louise van der Hoorn, Karin Pike-Overzet, and Frank J. T. Staal. 2022. "IL3 Has a Detrimental Effect on Hematopoietic Stem Cell Self-Renewal in Transplantation Settings" International Journal of Molecular Sciences 23, no. 21: 12736. https://doi.org/10.3390/ijms232112736
APA StyleTajer, P., Canté-Barrett, K., Naber, B. A. E., Vloemans, S. A., van Eggermond, M. C. J. A., van der Hoorn, M.-L., Pike-Overzet, K., & Staal, F. J. T. (2022). IL3 Has a Detrimental Effect on Hematopoietic Stem Cell Self-Renewal in Transplantation Settings. International Journal of Molecular Sciences, 23(21), 12736. https://doi.org/10.3390/ijms232112736