
Interleukin-22 Exerts Detrimental Effects on Salivary Gland Integrity and Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Elevation of IL-22 in Salivary Glands of NOD Mice Promotes Tissue Apoptosis and Secretory Dysfunction
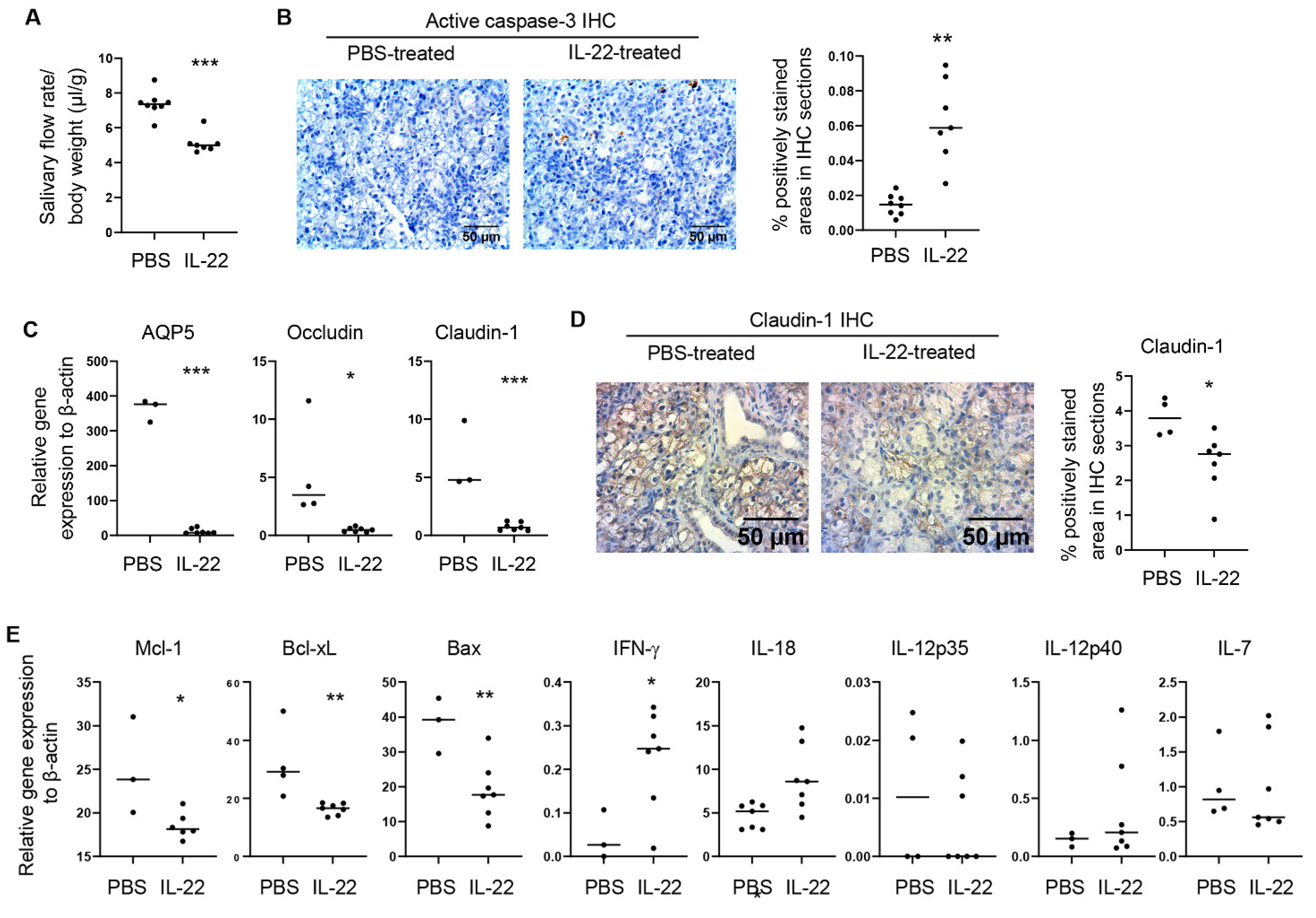
2.2. Administration of Exogenous IL-22 Induces Salivary Gland Tissue Apoptosis and Secretory Dysfunction in Normal C57BL/6 Mice
2.3. Neutralization of Endogenous IL-22 Attenuates Anti-CD3 Antibody-Induced Salivary Gland Tissue Apoptosis and Hyposalivation
2.4. IL-22 Treatment Reduces the Viable Cell Numbers of Mouse Primary Salivary Gland Epithelial Cells Cultured In Vitro
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cytokine and Antibodies
4.3. In Vivo Administration of Recombinant IL-22 and Anti-IL-22 Antibody
4.4. Measurement of Salivary Flow Rate
4.5. Histology and Immunohistochemistry
4.6. Flow Cytometry
4.7. Real-Time RT-PCR
4.8. In Vitro Culture and Treatment of Primary Mouse SMG Cells
4.9. Annexin V and 7-AAD Staining for Apoptosis Detection
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lee, B.H.; Tudares, M.A.; Nguyen, C.Q. Sjogren’s syndrome: An old tale with a new twist. Arch. Immunol. Et Ther. Exp. 2009, 57, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.; Shahane, A. The epidemiology of Sjogren’s syndrome. Clin. Epidemiol. 2014, 6, 247–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voulgarelis, M.; Tzioufas, A.G. Pathogenetic mechanisms in the initiation and perpetuation of Sjogren’s syndrome. Nat. Rev. Rheumatol. 2010, 6, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Ferrante, A.; Raimondo, S.; Giardina, A.; Dieli, F.; Campisi, G.; Alessandro, R.; Triolo, G. Potential involvement of IL-22 and IL-22-producing cells in the inflamed salivary glands of patients with Sjogren’s syndrome. Ann. Rheum. Dis. 2012, 71, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Sun, S.; Liu, Y.; Zhu, T.; Wang, K.; Ren, T.; Wu, Z.; Xu, H.; Zhu, L. Analysis of Th17-associated cytokines in tears of patients with dry eye syndrome. Eye 2014, 28, 608–613. [Google Scholar] [CrossRef]
- Kang, E.H.; Lee, Y.J.; Hyon, J.Y.; Yun, P.Y.; Song, Y.W. Salivary cytokine profiles in primary Sjogren’s syndrome differ from those in non-Sjogren sicca in terms of TNF-alpha levels and Th-1/Th-2 ratios. Clin. Exp. Rheumatol. 2011, 29, 970–976. [Google Scholar]
- Villarreal, G.M.; Alcocer-Varela, J.; Llorente, L. Differential interleukin (IL)-10 and IL-13 gene expression in vivo in salivary glands and peripheral blood mononuclear cells from patients with primary Sjogren’s syndrome. Immunol. Lett. 1996, 49, 105–109. [Google Scholar] [CrossRef]
- Lavoie, T.N.; Stewart, C.M.; Berg, K.M.; Li, Y.; Nguyen, C.Q. Expression of interleukin-22 in Sjogren’s syndrome: Significant correlation with disease parameters. Scand. J. Immunol. 2011, 74, 377–382. [Google Scholar] [CrossRef]
- Jin, J.O.; Yu, Q. T Cell-Associated Cytokines in the Pathogenesis of Sjogren’s Syndrome. J. Clin. Cell. Immunol. 2013, S! 11742. [Google Scholar] [CrossRef] [Green Version]
- Matsui, K.; Sano, H. T Helper 17 Cells in Primary Sjogren’s Syndrome. J. Clin. Med. 2017, 6, 65. [Google Scholar] [CrossRef] [Green Version]
- Baker, O.J.; Camden, J.M.; Redman, R.S.; Jones, J.E.; Seye, C.I.; Erb, L.; Weisman, G.A. Proinflammatory cytokines tumor necrosis factor-alpha and interferon-gamma alter tight junction structure and function in the rat parotid gland Par-C10 cell line. Am. J. Physiol. Cell Physiol. 2008, 295, C1191–C1201. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, M.; Kawakami, A.; Yamasaki, S.; Hida, A.; Nakashima, T.; Nakamura, H.; Ida, H.; Furuyama, M.; Nakashima, K.; Shibatomi, K.; et al. Regulation of apoptotic cell death by cytokines in a human salivary gland cell line: Distinct and synergistic mechanisms in apoptosis induced by tumor necrosis factor alpha and interferon gamma. J. Lab. Clin. Med. 2002, 139, 13–19. [Google Scholar] [CrossRef]
- Kulkarni, K.; Selesniemi, K.; Brown, T.L. Interferon-gamma sensitizes the human salivary gland cell line, HSG, to tumor necrosis factor-alpha induced activation of dual apoptotic pathways. Apoptosis Int. J. Program. Cell Death 2006, 11, 2205–2215. [Google Scholar] [CrossRef]
- Odusanwo, O.; Chinthamani, S.; McCall, A.; Duffey, M.E.; Baker, O.J. Resolvin D1 prevents TNF-alpha-mediated disruption of salivary epithelial formation. Am. J. Physiol. Cell Physiol. 2012, 302, C1331–C1345. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Kawai, T.; Yu, Q. Pathogenic role of endogenous TNF-alpha in the development of Sjogren’s-like sialadenitis and secretory dysfunction in non-obese diabetic mice. Lab. Investig. 2017, 97, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, G.M.; Alcocer-Varela, J.; Llorente, L. Cytokine gene and CD25 antigen expression by peripheral blood T cells from patients with primary Sjogren’s syndrome. Autoimmunity 1995, 20, 223–229. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Rutz, S.; Eidenschenk, C.; Ouyang, W. IL-22, not simply a Th17 cytokine. Immunol. Rev. 2013, 252, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Ouyang, W.; Wolk, K. Therapeutic opportunities of the IL-22-IL-22R1 system. Nat. Rev. Drug Discov. 2014, 13, 21–38. [Google Scholar] [CrossRef]
- Rutz, S.; Wang, X.; Ouyang, W. The IL-20 subfamily of cytokines--from host defence to tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 783–795. [Google Scholar] [CrossRef]
- Munoz, M.; Eidenschenk, C.; Ota, N.; Wong, K.; Lohmann, U.; Kuhl, A.A.; Wang, X.; Manzanillo, P.; Li, Y.; Rutz, S.; et al. Interleukin-22 induces interleukin-18 expression from epithelial cells during intestinal infection. Immunity 2015, 42, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Ulziibat, S.; Hardle, L.; Pfeilschifter, J.; Muhl, H. IFNalpha converts IL-22 into a cytokine efficiently activating STAT1 and its downstream targets. Biochem. Pharmacol. 2013, 85, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Muhl, H. Pro-Inflammatory Signaling by IL-10 and IL-22: Bad Habit Stirred Up by Interferons? Front. Immunol. 2013, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenberg, G.F.; Nair, M.G.; Kirn, T.J.; Zaph, C.; Fouser, L.A.; Artis, D. Pathological versus protective functions of IL-22 in airway inflammation are regulated by IL-17A. J. Exp. Med. 2010, 207, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Powell, N.; Pantazi, E.; Pavlidis, P.; Tsakmaki, A.; Li, K.; Yang, F.; Parker, A.; Pin, C.; Cozzetto, D.; Minns, D.; et al. Interleukin-22 orchestrates a pathological endoplasmic reticulum stress response transcriptional programme in colonic epithelial cells. Gut 2020, 69, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.W.; Mittal, S.K.; Hwang, H.S.; Chang, E.J.; Lee, J.H.; Seo, Y.; Yeo, A.; Noh, H.; Lee, H.S.; Chauhan, S.K.; et al. Lacrimal gland-derived IL-22 regulates IL-17-mediated ocular mucosal inflammation. Mucosal Immunol. 2017, 10, 1202–1210. [Google Scholar] [CrossRef] [Green Version]
- Barone, F.; Nayar, S.; Campos, J.; Cloake, T.; Withers, D.R.; Toellner, K.M.; Zhang, Y.; Fouser, L.; Fisher, B.; Bowman, S.; et al. IL-22 regulates lymphoid chemokine production and assembly of tertiary lymphoid organs. Proc. Natl. Acad. Sci. USA 2015, 112, 11024–11029. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jin, J.O.; Kawai, T.; Yu, Q. Endogenous programmed death ligand-1 restrains the development and onset of Sjgren’s syndrome in non-obese diabetic mice. Sci. Rep. 2016, 6, 39105. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yu, Q. Disruption of CXCR3 function impedes the development of Sjogren’s syndrome-like xerostomia in non-obese diabetic mice. Lab. Investig. 2018, 98, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Wolk, K.; Witte, E.; Hoffmann, U.; Doecke, W.D.; Endesfelder, S.; Asadullah, K.; Sterry, W.; Volk, H.D.; Wittig, B.M.; Sabat, R. IL-22 induces lipopolysaccharide-binding protein in hepatocytes: A potential systemic role of IL-22 in Crohn’s disease. J. Immunol. 2007, 178, 5973–5981. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Yi, P.; Ru, W.; Jie, Z.; Wang, H.; Ghanayem, T.; Wang, X.; Alamer, E.; Liu, J.; Hu, H.; et al. IL-22 hinders antiviral T cell responses and exacerbates ZIKV encephalitis in immunocompetent neonatal mice. J. Neuroinflammat. 2020, 17, 249. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jin, J.O.; Patel, E.S.; Yu, Q. Interleukin-6 inhibits apoptosis of exocrine gland tissues under inflammatory conditions. Cytokine 2015, 76, 244–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, O.J. Current trends in salivary gland tight junctions. Tissue Barriers 2016, 4, e1162348. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Song, Y.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A.S. Defective secretion of saliva in transgenic mice lacking aquaporin-5 water channels. J. Biol. Chem. 1999, 274, 20071–20074. [Google Scholar] [CrossRef] [Green Version]
- Lai, Z.; Yin, H.; Cabrera-Perez, J.; Guimaro, M.C.; Afione, S.; Michael, D.G.; Glenton, P.; Patel, A.; Swaim, W.D.; Zheng, C.; et al. Aquaporin gene therapy corrects Sjogren’s syndrome phenotype in mice. Proc. Natl. Acad. Sci. USA 2016, 113, 5694–5699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.; Brayer, J.; Gao, J.; Brown, V.; Killedar, S.; Yasunari, U.; Peck, A.B. A dual role for interferon-gamma in the pathogenesis of Sjogren’s syndrome-like autoimmune exocrinopathy in the nonobese diabetic mouse. Scand. J. Immunol. 2004, 60, 552–565. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, Q. Anti-IL-7 receptor-alpha treatment ameliorates newly established Sjogren’s-like exocrinopathy in non-obese diabetic mice. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 2438–2447. [Google Scholar] [CrossRef]
- Ferran, C.; Sheehan, K.; Dy, M.; Schreiber, R.; Merite, S.; Landais, P.; Noel, L.H.; Grau, G.; Bluestone, J.; Bach, J.F.; et al. Cytokine-related syndrome following injection of anti-CD3 monoclonal antibody: Further evidence for transient in vivo T cell activation. Eur. J. Immunol. 1990, 20, 509–515. [Google Scholar] [CrossRef]
- Matthys, P.; Dillen, C.; Proost, P.; Heremans, H.; Van Damme, J.; Billiau, A. Modification of the anti-CD3-induced cytokine release syndrome by anti-interferon-gamma or anti-interleukin-6 antibody treatment: Protective effects and biphasic changes in blood cytokine levels. Eur. J. Immunol. 1993, 23, 2209–2216. [Google Scholar] [CrossRef]
- Ikeura, K.; Kawakita, T.; Tsunoda, K.; Nakagawa, T.; Tsubota, K. Characterization of Long-Term Cultured Murine Submandibular Gland Epithelial Cells. PLoS ONE 2016, 11, e0147407. [Google Scholar] [CrossRef] [Green Version]
- Roescher, N.; Tak, P.P.; Illei, G.G. Cytokines in Sjogren’s syndrome. Oral Dis. 2009, 15, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Schinocca, C.; Rizzo, C.; Fasano, S.; Grasso, G.; La Barbera, L.; Ciccia, F.; Guggino, G. Role of the IL-23/IL-17 Pathway in Rheumatic Diseases: An Overview. Front. Immunol. 2021, 12, 637829. [Google Scholar] [CrossRef]
- Jin, J.O.; Han, X.; Yu, Q. Interleukin-6 induces the generation of IL-10-producing Tr1 cells and suppresses autoimmune tissue inflammation. J. Autoimmun. 2013, 40, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Huber, S.; Gagliani, N.; Esplugues, E.; O’Connor, W., Jr.; Huber, F.J.; Chaudhry, A.; Kamanaka, M.; Kobayashi, Y.; Booth, C.J.; Rudensky, A.Y.; et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3(-) and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity 2011, 34, 554–565. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Onodera, S.; Hu, Y.; Yu, Q. Interleukin-22 Exerts Detrimental Effects on Salivary Gland Integrity and Function. Int. J. Mol. Sci. 2022, 23, 12997. https://doi.org/10.3390/ijms232112997
Zhou J, Onodera S, Hu Y, Yu Q. Interleukin-22 Exerts Detrimental Effects on Salivary Gland Integrity and Function. International Journal of Molecular Sciences. 2022; 23(21):12997. https://doi.org/10.3390/ijms232112997
Chicago/Turabian StyleZhou, Jing, Shoko Onodera, Yang Hu, and Qing Yu. 2022. "Interleukin-22 Exerts Detrimental Effects on Salivary Gland Integrity and Function" International Journal of Molecular Sciences 23, no. 21: 12997. https://doi.org/10.3390/ijms232112997
APA StyleZhou, J., Onodera, S., Hu, Y., & Yu, Q. (2022). Interleukin-22 Exerts Detrimental Effects on Salivary Gland Integrity and Function. International Journal of Molecular Sciences, 23(21), 12997. https://doi.org/10.3390/ijms232112997