

Normalization of the Immunological Microenvironment and Sustained Minimal Residual Disease Negativity: Do We Need Both for Long-Term Control of Multiple Myeloma?

 and
and
Abstract
:1. Microenvironment in Multiple Myeloma: Composition and Role
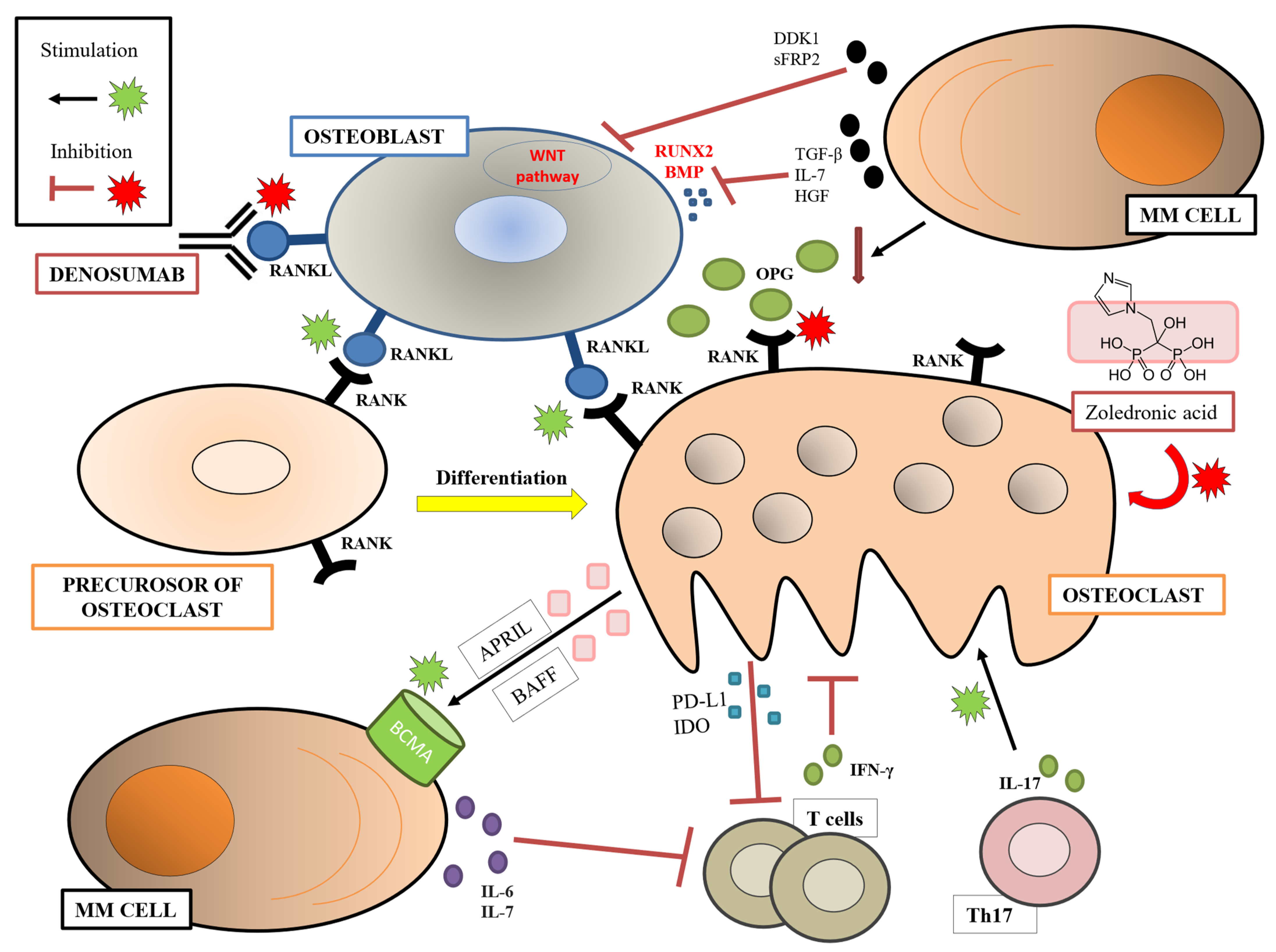
1.1. Osteoclasts, Osteoblasts, and Bone Disease
1.2. Adipocytes
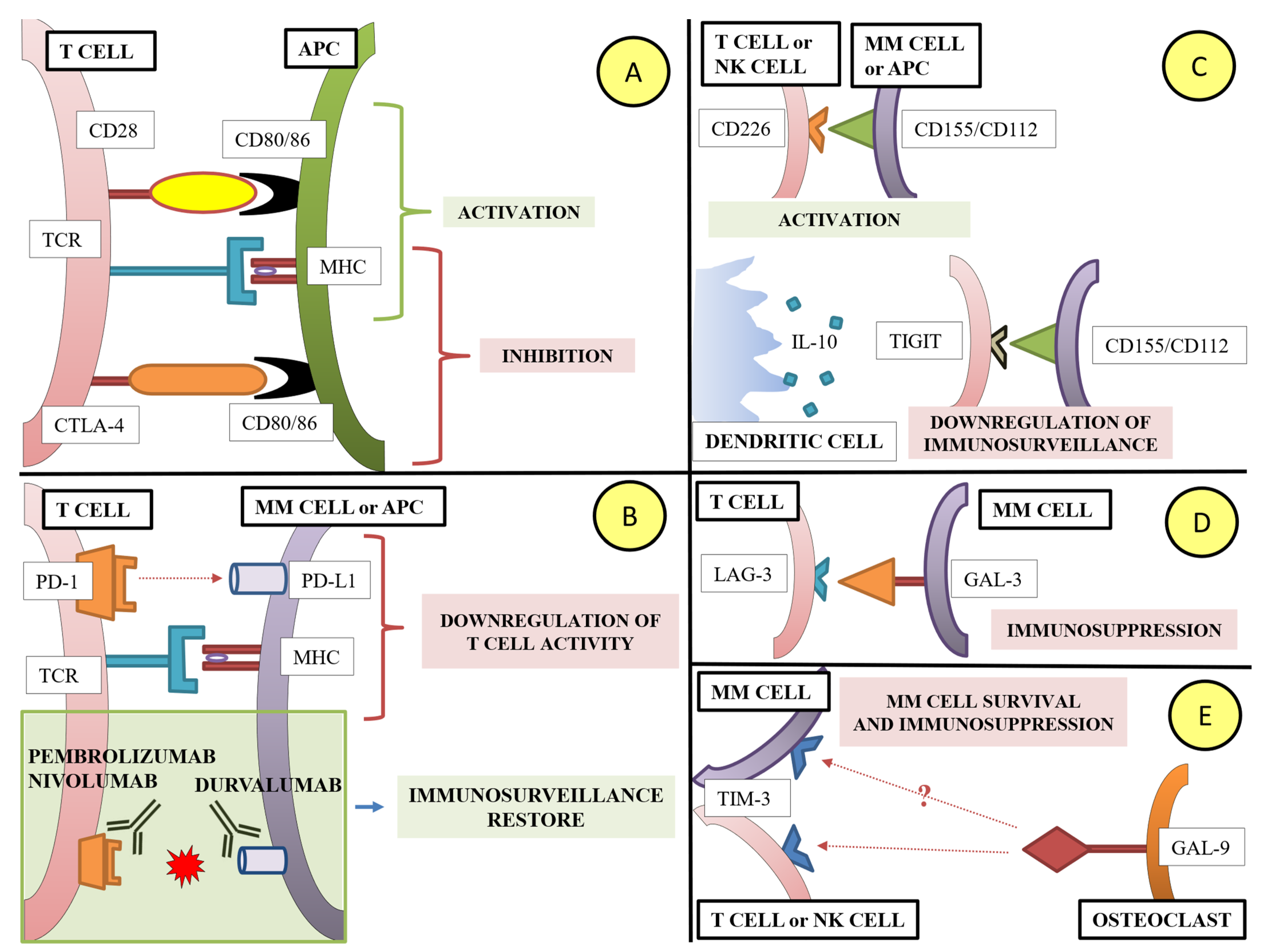
1.3. Lymphoid Cells and Immune Checkpoint Molecules
1.4. Myeloid Cells
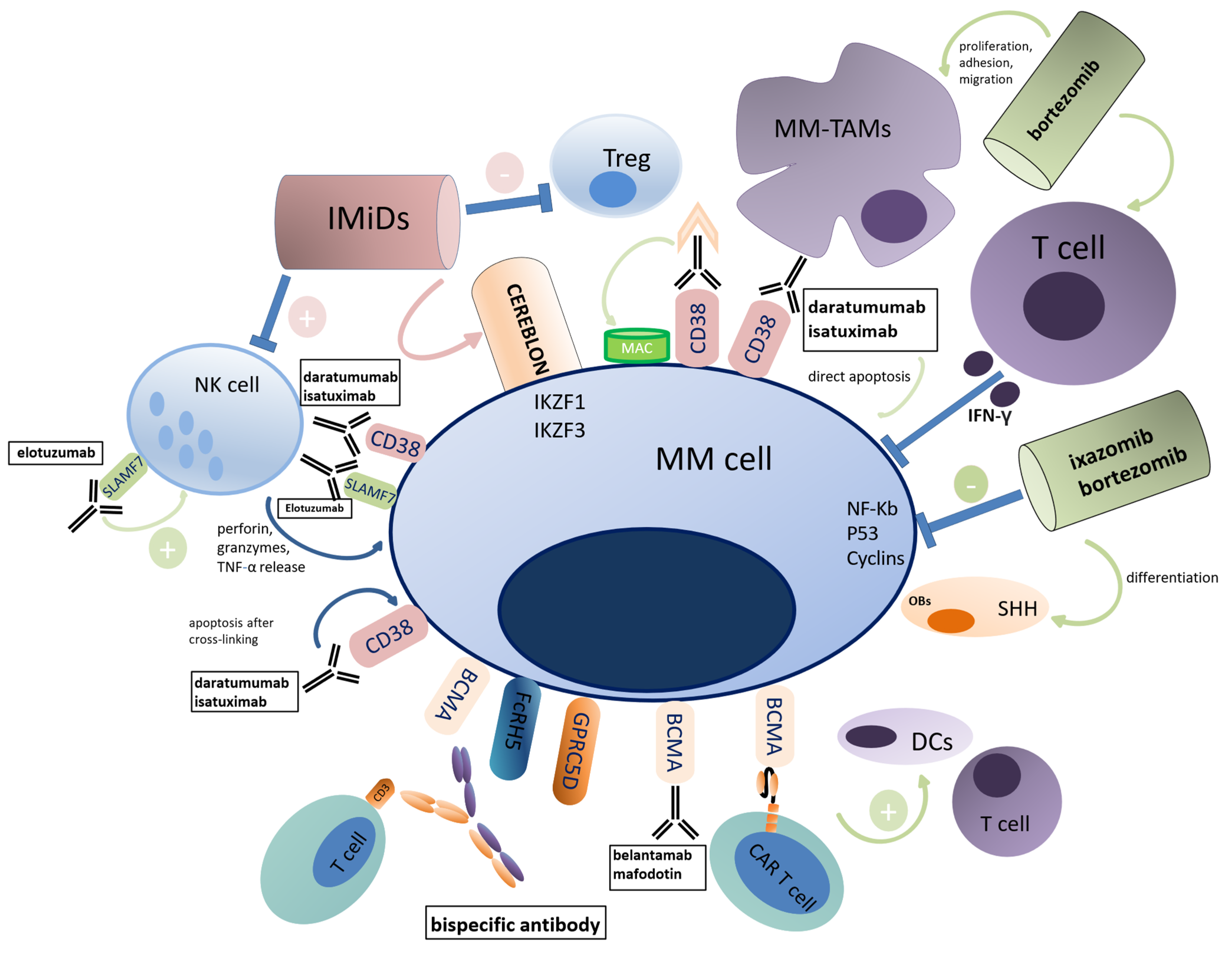
1.5. Influence of Anti-MM Drugs on the ME
2. MRD in MM: Relevance as Prognostic Factor
3. Prognostic Impact of PC Biology on Sustained MRD Negativity
4. Prognostic Impact of the BM ME on Sustained MRD Negativity
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Palumbo, A.; Anderson, K. Multiple myeloma. N. Engl. J. Med. 2011, 364, 1046–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terpos, E.; Berenson, J.; Raje, N.; Roodman, G.D. Management of bone disease in multiple myeloma. Expert Rev. Hematol. 2014, 7, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terpos, E.; Szydlo, R.; Apperley, J.F.; Hatjiharissi, E.; Politou, M.; Meletis, J.; Viniou, N.; Yataganas, X.; Goldman, J.M.; Rahemtulla, A. Soluble receptor activator of nuclear factor kappaB ligand-osteoprotegerin ratio predicts survival in multiple myeloma: Proposal for a novel prognostic index. Blood 2003, 102, 1064–1069. [Google Scholar] [CrossRef]
- Terpos, E.; Politou, M.; Szydlo, R.; Goldman, J.M.; Apperley, J.F.; Rahemtulla, A. Serum levels of macrophage inflammatory protein-1 alpha (MIP-1alpha) correlate with the extent of bone disease and survival in patients with multiple myeloma. Br. J. Haematol. 2003, 123, 106–109. [Google Scholar] [CrossRef]
- Choi, S.J.; Cruz, J.C.; Craig, F.; Chung, H.; Devlin, R.D.; Roodman, G.D.; Alsina, M. Macrophage inflammatory protein 1-alpha is a potential osteoclast stimulatory factor in multiple myeloma. Blood 2000, 96, 671–675. [Google Scholar] [CrossRef]
- Zannettino, A.C.W.; Farrugia, A.N.; Kortesidis, A.; Manavis, J.; To, L.B.; Martin, S.K.; Diamond, P.; Tamamura, H.; Lapidot, T.; Fujii, N.; et al. Elevated serum levels of stromal-derived factor-1alpha are associated with increased osteoclast activity and osteolytic bone disease in multiple myeloma patients. Cancer Res. 2005, 65, 1700–1709. [Google Scholar] [CrossRef] [Green Version]
- Niida, S.; Kaku, M.; Amano, H.; Yoshida, H.; Kataoka, H.; Nishikawa, S.; Tanne, K.; Maeda, N.; Nishikawa, S.I.; Kodama, H. Vascular endothelial growth factor can substitute for macrophage colony-stimulating factor in the support of osteoclastic bone resorption. J. Exp. Med. 1999, 190, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Abe, M.; Hiasa, M.; Oda, A.; Amou, H.; Nakano, A.; Takeuchi, K.; Kitazoe, K.; Kido, S.; Inoue, D.; et al. Myeloma cell-osteoclast interaction enhances angiogenesis together with bone resorption: A role for vascular endothelial cell growth factor and osteopontin. Clin. Cancer Res. 2007, 13, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, N.; Bataille, R.; Mancini, C.; Lazzaretti, M.; Barillé, S. Myeloma cells induce imbalance in the osteoprotegerin/osteoprotegerin ligand system in the human bone marrow environment. Blood 2001, 98, 3527–3533. [Google Scholar] [CrossRef]
- Li, J.; Sarosi, I.; Cattley, R.C.; Pretorius, J.; Asuncion, F.; Grisanti, M.; Morony, S.; Adamu, S.; Geng, Z.; Qiu, W.; et al. Dkk1-mediated inhibition of Wnt signaling in bone results in osteopenia. Bone 2006, 39, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Oyajobi, B.; Garrett, I.; Gupta, A.; Banerjee, M.; Esparza, X.; Flores, A.; Sterling, J.; Rossini, G.; Zhao, M.; Mundy, G. Role of Dickkopf 1 (DKK) in myeloma bone disease and modulation by the proteasome inhibitor velcade. J. Bone Miner. Res. 2004, 19, S4, Abstract #1011 [ASBMR 2004 26th Annual Meeting]. [Google Scholar]
- Oshima, T.; Abe, M.; Asano, J.; Hara, T.; Kitazoe, K.; Sekimoto, E.; Tanaka, Y.; Shibata, H.; Hashimoto, T.; Ozaki, S.; et al. Myeloma cells suppress bone formation by secreting a soluble Wnt inhibitor, sFRP-2. Blood 2005, 106, 3160–3165. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, N.; Colla, S.; Morandi, F.; Lazzaretti, M.; Sala, R.; Bonomini, S.; Grano, M.; Colucci, S.; Svaldi, M.; Rizzoli, V. Myeloma cells block RUNX2/CBFA1 activity in human bone marrow osteoblast progenitors and inhibit osteoblast formation and differentiation. Blood 2005, 106, 2472–2483. [Google Scholar] [CrossRef] [PubMed]
- Standal, T.; Abildgaard, N.; Fagerli, U.M.; Stordal, B.; Hjertner, Ø.; Borset, M.; Sundan, A. HGF inhibits BMP-induced osteoblastogenesis: Possible implications for the bone disease of multiple myeloma. Blood 2007, 109, 3024–3030. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Abe, M.; Oda, A.; Amou, H.; Hiasa, M.; Asano, J.; Kitazoe, K.; Kido, S.; Inoue, D.; Hashimoto, T.; et al. Enhancement of osteoblast differentiation by inhibition of TGF-beta signaling suppresses myeloma cell growth and protects from destructive bone lesions. J. Bone Miner. Res. 2006, 21, S28, Abstract #1101 [ASBMR 2006 28th Annual Meeting]. [Google Scholar]
- An, G.; Acharya, C.; Feng, X.; Wen, K.; Zhong, M.; Zhang, L.; Munshi, N.C.; Qiu, L.; Tai, Y.T.; Anderson, K.C. Osteoclasts promote immune suppressive microenvironment in multiple myeloma: Therapeutic implication. Blood 2016, 128, 1590–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukaihara, K.; D’Agostino, M.; Panaroni, C.; Fulzele, K.; Mori, T.; Siu, K.T.; Chattopadhyay, S.; Raje, N. Osteoclast-Induced Immunosuppression Occurs through Dysregulation of Immune Checkpoint Axes in Multiple Myeloma. Blood 2019, 134, 4382, [ASH 2019 61st Meeting]. [Google Scholar] [CrossRef]
- Takayanagi, H.; Ogasawara, K.; Hida, S.; Chiba, T.; Murata, S.; Sato, K.; Takaoka, A.; Yokochi, T.; Oda, H.; Tanaka, K.; et al. T-cell-mediated regulation of osteoclastogenesis by signalling cross-talk between RANKL and IFN-gamma. Nature 2000, 408, 600–605. [Google Scholar] [CrossRef]
- Giuliani, N.; Colla, S.; Sala, R.; Moroni, M.; Lazzaretti, M.; La Monica, S.; Bonomini, S.; Hojden, M.; Sammarelli, G.; Barillè, S.; et al. Human myeloma cells stimulate the receptor activator of nuclear factor-kappa B ligand (RANKL) in T lymphocytes: A potential role in multiple myeloma bone disease. Blood 2002, 100, 4615–4621. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Kido, S.; Hiasa, M.; Nakano, A.; Oda, A.; Amou, H.; Matsumoto, T. BAFF and APRIL as osteoclast-derived survival factors for myeloma cells: A rationale for TACI-Fc treatment in patients with multiple myeloma. Leukemia 2006, 20, 1313–1315. [Google Scholar] [CrossRef] [Green Version]
- Moreaux, J.; Sprynski, A.C.; Dillon, S.R.; Mahtouk, K.; Jourdan, M.; Ythier, A.; Moine, P.; Robert, N.; Jourdan, E.; Rossi, J.F.; et al. APRIL and TACI interact with syndecan-1 on the surface of multiple myeloma cells to form an essential survival loop. Eur. J. Haematol. 2009, 83, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.; Wakkach, A.; Blin-Wakkach, C. Emerging Roles of Osteoclasts in the Modulation of Bone Microenvironment and Immune Suppression in Multiple Myeloma. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, K.; Marchionni, L.; Anderson, J.; Pardoll, D.; Roodman, G.D.; Borrello, I. A novel role of IL-17-producing lymphocytes in mediating lytic bone disease in multiple myeloma. Blood 2010, 116, 3554–3563. [Google Scholar] [CrossRef] [Green Version]
- Tai, Y.T.; Acharya, C.; An, G.; Moschetta, M.; Zhong, M.Y.; Feng, X.; Cea, M.; Cagnetta, A.; Wen, K.; Van Eenennaam, H.; et al. APRIL and BCMA promote human multiple myeloma growth and immunosuppression in the bone marrow microenvironment. Blood 2016, 127, 3225–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, H.; Wang, S.; Chen, X.; Su, J. Bone Marrow Adipocytes: A Critical Player in the Bone Marrow Microenvironment. Front. Cell Dev. Biol. 2021, 9, 770705. [Google Scholar] [CrossRef]
- Panaroni, C.; Fulzele, K.; Mori, T.; Siu, K.T.; Onyewadume, C.; Maebius, A.; Raje, N. Multiple myeloma cells induce lipolysis in adipocytes and uptake fatty acids through fatty acid transporter proteins. Blood 2022, 139, 876–888. [Google Scholar] [CrossRef]
- Fairfield, H.; Costa, S.; Falank, C.; Farrell, M.; Murphy, C.S.; D’Amico, A.; Driscoll, H.; Reagan, M.R. Multiple Myeloma Cells Alter Adipogenesis, Increase Senescence-Related and Inflammatory Gene Transcript Expression, and Alter Metabolism in Preadipocytes. Front. Oncol. 2021, 10, 584683. [Google Scholar] [CrossRef]
- Jafari, A.; Fairfield, H.; Andersen, T.L.; Reagan, M.R. Myeloma-bone marrow adipocyte axis in tumour survival and treatment response. Br. J. Cancer 2021, 125, 775–777. [Google Scholar] [CrossRef]
- Trotter, T.N.; Gibson, J.T.; Sherpa, T.L.; Gowda, P.S.; Peker, D.; Yang, Y. Adipocyte-Lineage Cells Support Growth and Dissemination of Multiple Myeloma in Bone. Am. J. Pathol. 2016, 186, 3054–3063. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; He, J.; Koh, S.P.; Zhong, Y.; Liu, Z.; Wang, Z.; Zhang, Y.; Li, Z.; Tam, B.T.; Lin, P.; et al. Reprogrammed marrow adipocytes contribute to myeloma-induced bone disease. Sci. Transl. Med. 2019, 11, eaau9087. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.V.; Suchacki, K.J.; Hocking, J.; Cartwright, R.; Sowman, A.; Gamez, B.; Lea, R.; Drake, M.T.; Cawthorn, W.P.; Edwards, C.M. Myeloma Cells Down-Regulate Adiponectin in Bone Marrow Adipocytes Via TNF-Alpha. J. Bone Miner. Res. 2020, 35, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Pratt, G.; Goodyear, O.; Moss, P. Immunodeficiency and immunotherapy in multiple myeloma. Br. J. Haematol. 2007, 138, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Prabhala, R.H.; Neri, P.; Bae, J.E.; Tassone, P.; Shammas, M.A.; Allam, C.K.; Daley, J.F.; Chauhan, D.; Blanchard, E.; Thatte, H.S.; et al. Dysfunctional T regulatory cells in multiple myeloma. Blood 2006, 107, 301–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelle-Rieser, C.; Thangavadivel, S.; Biedermann, R.; Brunner, A.; Stoitzner, P.; Willenbacher, E.; Greil, R.; Jöhrer, K. T cells in multiple myeloma display features of exhaustion and senescence at the tumor site. J. Hematol. Oncol. 2016, 9, 116. [Google Scholar] [CrossRef] [Green Version]
- Miyara, M.; Yoshioka, Y.; Kitoh, A.; Shima, T.; Wing, K.; Niwa, A.; Parizot, C.; Taflin, C.; Heike, T.; Valeyre, D.; et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009, 30, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Tai, Y.T.; Lin, L.; Xing, L.; Cho, S.F.; Yu, T.; Acharya, C.; Wen, K.; Hsieh, P.A.; Dulos, J.; van Elsas, A.; et al. APRIL signaling via TACI mediates immunosuppression by T regulatory cells in multiple myeloma: Therapeutic implications. Leukemia 2019, 33, 426–438. [Google Scholar] [CrossRef]
- Bonanno, G.; Mariotti, A.; Procoli, A.; Folgiero, V.; Natale, D.; De Rosa, L.; Majolino, I.; Novarese, L.; Rocci, A.; Gambella, M.; et al. Indoleamine 2,3-dioxygenase 1 (IDO1) activity correlates with immune system abnormalities in multiple myeloma. J. Transl. Med. 2012, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.J.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef]
- Prabhala, R.H.; Pelluru, D.; Fulciniti, M.; Prabhala, H.K.; Nanjappa, P.; Song, W.; Pai, C.; Amin, S.; Tai, Y.T.; Richardson, P.G.; et al. Elevated IL-17 produced by TH17 cells promotes myeloma cell growth and inhibits immune function in multiple myeloma. Blood 2010, 115, 5385–5392. [Google Scholar] [CrossRef]
- Shen, C.J.; Yuan, Z.H.; Liu, Y.X.; Hu, G.Y. Increased numbers of T helper 17 cells and the correlation with clinicopathological characteristics in multiple myeloma. J. Int. Med. Res. 2012, 40, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.T.; Zhang, Y.C.; Zhou, X.; Xie, P.H.; Li, J. A Unique Role of T Helper 17 Cells in Different Treatment Stages of Multiple Myeloma. Clin. Lymphoma Myeloma Leuk. 2020, 20, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Favaloro, J.; Brown, R.; Aklilu, E.; Yang, S.; Suen, H.; Hart, D.; Fromm, P.; Gibson, J.; Khoo, L.; Ho, P.J.; et al. Myeloma skews regulatory T and pro-inflammatory T helper 17 cell balance in favor of a suppressive state. Leuk. Lymphoma 2014, 55, 1090–1098. [Google Scholar] [CrossRef]
- Rosser, E.C.; Mauri, C. Regulatory B cells: Origin, phenotype, and function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Tai, Y.T.; Ho, M.; Xing, L.; Chauhan, D.; Gang, A.; Qiu, L.; Anderson, K.C. Regulatory B cell-myeloma cell interaction confers immunosuppression and promotes their survival in the bone marrow milieu. Blood Cancer J. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Schütt, P.; Brandhorst, D.; Stellberg, W.; Poser, M.; Ebeling, P.; Müller, S.; Buttkereit, U.; Opalka, B.; Lindemann, M.; Grosse-Wilde, H.; et al. Immune parameters in multiple myeloma patients: Influence of treatment and correlation with opportunistic infections. Leuk. Lymphoma 2006, 47, 1570–1582. [Google Scholar] [CrossRef]
- Jurisic, V.; Srdic, T.; Konjevic, G.; Markovic, O.; Colovic, M. Clinical stage-depending decrease of NK cell activity in multiple myeloma patients. Med. Oncol. 2007, 24, 312–317. [Google Scholar] [CrossRef]
- Frassanito, M.A.; Silvestris, F.; Cafforio, P.; Silvestris, N.; Dammacco, F. IgG M-components in active myeloma patients induce a down-regulation of natural killer cell activity. Int. J. Clin. Lab. Res. 1997, 27, 48–54. [Google Scholar] [CrossRef]
- Konjević, G.; Vuletić, A.; Mirjačić Martinović, K.; Colović, N.; Čolović, M.; Jurišić, V. Decreased CD161 activating and increased CD158a inhibitory receptor expression on NK cells underlies impaired NK cell cytotoxicity in patients with multiple myeloma. J. Clin. Pathol. 2016, 69, 1009–1016. [Google Scholar] [CrossRef]
- Seymour, F.; Cavenagh, J.; Mathews, J.; Gribben, J.G. NK cells CD56bright and CD56dim subset cytokine loss and exhaustion is associated with impaired survival in myeloma. Blood Adv. 2022, 6, 5152–5159. [Google Scholar] [CrossRef]
- Benson, D.M.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Braga, W.M.T.; da Silva, B.R.; de Carvalho, A.C.; Maekawa, Y.H.; Bortoluzzo, A.B.; Rizzatti, E.G.; Atanackovic, D.; Colleoni, G.W.B. FOXP3 and CTLA4 overexpression in multiple myeloma bone marrow as a sign of accumulation of CD4(+) T regulatory cells. Cancer Immunol. Immunother. 2014, 63, 1189–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pianko, M.J.; Liu, Y.; Bagchi, S.; Lesokhin, A.M. Immune checkpoint blockade for hematologic malignancies: A review. Stem Cell Investig. 2017, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, B.; Azpilikueta, A.; Puig, N.; Ocio, E.M.; Sharma, R.; Oyajobi, B.O.; Labiano, S.; San-Segundo, L.; Rodriguez, A.; Aires-Mejia, I.; et al. PD-L1/PD-1 presence in the tumor microenvironment and activity of PD-1 blockade in multiple myeloma. Leukemia 2015, 29, 2110–2113. [Google Scholar] [CrossRef]
- Asimakopoulos, F. TIGIT checkpoint inhibition for myeloma. Blood 2018, 132, 1629–1630. [Google Scholar] [CrossRef]
- Guillerey, C.; Harjunpää, H.; Carrié, N.; Kassem, S.; Teo, T.; Miles, K.; Krumeich, S.; Weulersse, M.; Cuisinier, M.; Stannard, K.; et al. TIGIT immune checkpoint blockade restores CD8+ T-cell immunity against multiple myeloma. Blood 2018, 132, 1689–1694. [Google Scholar] [CrossRef] [Green Version]
- Neri, P.; Maity, R.; Tagoug, I.; McCulloch, S.; Duggan, P.; Jimenez-Zepeda, V.; Tay, J.; Thakurta, A.; Bahlis, N. Immunome Single Cell Profiling Reveals T Cell Exhaustion with Upregulation of Checkpoint Inhibitors LAG3 and Tigit on Marrow Infiltrating T Lymphocytes in Daratumumab and IMiDs Resistant Patients. Blood 2018, 132, 242, [ASH 2018 60th Meeting]. [Google Scholar] [CrossRef]
- Goldberg, M.V.; Drake, C.G. LAG-3 in Cancer Immunotherapy. Curr. Top. Microbiol. Immunol. 2011, 344, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Kouo, T.; Huang, L.; Pucsek, A.B.; Cao, M.; Solt, S.; Armstrong, T.; Jaffee, E. Galectin-3 Shapes Antitumor Immune Responses by Suppressing CD8+ T Cells via LAG-3 and Inhibiting Expansion of Plasmacytoid Dendritic Cells. Cancer Immunol. Res. 2015, 3, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Maruhashi, T.; Sugiura, D.; Okazaki, I.M.; Okazaki, T. LAG-3: From molecular functions to clinical applications. J. Immunother. Cancer 2020, 8, e001014. [Google Scholar] [CrossRef]
- Liu, Z.; Xiang, C.; Han, M.; Meng, N.; Luo, J.; Fu, R. Study on Tim3 Regulation of Multiple Myeloma Cell Proliferation via NF-κB Signal Pathways. Front. Oncol. 2020, 10, 584530. [Google Scholar] [CrossRef] [PubMed]
- Batorov, E.V.; Aristova, T.A.; Sergeevicheva, V.V.; Sizikova, S.A.; Ushakova, G.Y.; Pronkina, N.V.; Shishkova, I.V.; Shevela, E.Y.; Ostanin, A.A.; Chernykh, E.R. Quantitative and functional characteristics of circulating and bone marrow PD-1- and TIM-3-positive T cells in treated multiple myeloma patients. Sci. Rep. 2020, 10, 20846. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Chen, S.; Huang, J.; Chen, Y.; Yang, L.; Zhong, J.; Lu, Y.; Li, Y. High PD-1 and Tim-3 Expression Concurrent with Exhausted CD4+ and CD8+ T Cells in Bone Marrow Compared with Peripheral Blood from Patients with Multiple Myeloma. Blood 2017, 130, 4391, [ASH 2017 59th Meeting]. [Google Scholar] [CrossRef]
- Dao, T.N.; Utturkar, S.; Lanman, N.A.; Matosevic, S. TIM-3 Expression Is Downregulated on Human NK Cells in Response to Cancer Targets in Synergy with Activation. Cancers 2020, 12, 2417. [Google Scholar] [CrossRef] [PubMed]
- Storti, P.; Marchica, V.; Giuliani, N. Role of Galectins in Multiple Myeloma. Int. J. Mol. Sci. 2017, 18, 2740. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Kuroda, J.; Ashihara, E.; Oomizu, S.; Terui, Y.; Taniyama, A.; Adachi, S.; Takagi, T.; Yamamoto, M.; Sasaki, N.; et al. Galectin-9 exhibits anti-myeloma activity through JNK and p38 MAP kinase pathways. Leukemia 2010, 24, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Minnie, S.A.; Kuns, R.D.; Gartlan, K.H.; Zhang, P.; Wilkinson, A.N.; Samson, L.; Guillerey, C.; Engwerda, C.; MacDonald, K.P.A.; Smyth, M.J.; et al. Myeloma escape after stem cell transplantation is a consequence of T-cell exhaustion and is prevented by TIGIT blockade. Blood 2018, 132, 1675–1688. [Google Scholar] [CrossRef]
- Jiang, W.; Li, F.; Jiang, Y.; Li, S.; Liu, X.; Xu, Y.; Li, B.; Feng, X.; Zheng, C. Tim-3 Blockade Elicits Potent Anti-Multiple Myeloma Immunity of Natural Killer Cells. Front. Oncol. 2022, 12, 739976. [Google Scholar] [CrossRef]
- Bae, J.; Accardi, F.; Hideshima, T.; Tai, Y.-T.; Prabhala, R.; Shambley, A.; Wen, K.; Rowell, S.; Richardson, P.G.; Munshi, N.C.; et al. Targeting LAG3/GAL-3 to overcome immunosuppression and enhance anti-tumor immune responses in multiple myeloma. Leukemia 2022, 36, 138–154. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Zheng, Y.; Cai, Z.; Wang, S.; Zhang, X.; Qian, J.; Hong, S.; Li, H.; Wang, M.; Yang, J.; Yi, Q. Macrophages are an abundant component of myeloma microenvironment and protect myeloma cells from chemotherapy drug-induced apoptosis. Blood 2009, 114, 3625–3628. [Google Scholar] [CrossRef]
- Suyanı, E.; Sucak, G.T.; Akyürek, N.; Şahin, S.; Baysal, N.A.; Yağcı, M.; Haznedar, R. Tumor-associated macrophages as a prognostic parameter in multiple myeloma. Ann. Hematol. 2013, 92, 669–677. [Google Scholar] [CrossRef]
- Wang, H.; Hu, W.M.; Xia, Z.J.; Liang, Y.; Lu, Y.; Lin, S.X.; Tang, H. High numbers of CD163+ tumor-associated macrophages correlate with poor prognosis in multiple myeloma patients receiving bortezomib-based regimens. J. Cancer 2019, 10, 3239–3245. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou, K.; Tsakirakis, N.; Malandrakis, P.; Vitsos, P.; Metousis, A.; Orologas-Stavrou, N.; Ntanasis-Stathopoulos, I.; Kanellias, N.; Eleutherakis-Papaiakovou, E.; Pothos, P.; et al. Deep Phenotyping Reveals Distinct Immune Signatures Correlating with Prognostication, Treatment Responses, and MRD Status in Multiple Myeloma. Cancers 2020, 12, 3245. [Google Scholar] [CrossRef]
- Beider, K.; Bitner, H.; Leiba, M.; Gutwein, O.; Koren-Michowitz, M.; Ostrovsky, O.; Abraham, M.; Wald, H.; Galun, E.; Peled, A.; et al. Multiple myeloma cells recruit tumor-supportive macrophages through the CXCR4/CXCL12 axis and promote their polarization toward the M2 phenotype. Oncotarget 2014, 5, 11283–11296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Raimondo, F.; Azzaro, M.; Palumbo, G.; Bagnato, S.; Giustolisi, G.; Floridia, P.; Sortino, G.; Giustolisi, R. Angiogenic factors in multiple myeloma: Higher levels in bone marrow than in peripheral blood. Haematologica 2000, 85, 800–805. [Google Scholar] [PubMed]
- Scavelli, C.; Nico, B.; Cirulli, T.; Ria, R.; Di Pietro, G.; Mangieri, D.; Bacigalupo, A.; Mangialardi, G.; Coluccia, A.M.L.; Caravita, T.; et al. Vasculogenic mimicry by bone marrow macrophages in patients with multiple myeloma. Oncogene 2008, 27, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Fauskanger, M.; Haabeth, O.A.W.; Skjeldal, F.M.; Bogen, B.; Tveita, A.A. Tumor Killing by CD4+ T Cells Is Mediated via Induction of Inducible Nitric Oxide Synthase-Dependent Macrophage Cytotoxicity. Front. Immunol. 2018, 9, 1684. [Google Scholar] [CrossRef] [Green Version]
- Haabeth, O.A.W.; Lorvik, K.B.; Hammarström, C.; Donaldson, I.M.; Haraldsen, G.; Bogen, B.; Corthay, A. Inflammation driven by tumour-specific Th1 cells protects against B-cell cancer. Nat. Commun. 2011, 2, 240. [Google Scholar] [CrossRef] [Green Version]
- Tveita, A.A.; Schjesvold, F.; Haabeth, O.A.; Fauskanger, M.; Bogen, B. Tumors Escape CD4+ T-cell-Mediated Immunosurveillance by Impairing the Ability of Infiltrating Macrophages to Indirectly Present Tumor Antigens. Cancer Res. 2015, 75, 3268–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, I.R.; Martner, A.; Pisklakova, A.; Condamine, T.; Chase, T.; Vogl, T.; Roth, J.; Gabrilovich, D.; Nefedova, Y. Myeloid-derived suppressor cells regulate growth of multiple myeloma by inhibiting T cells in bone marrow. J. Immunol. 2013, 190, 3815–3823. [Google Scholar] [CrossRef]
- Van Valckenborgh, E.; Van Ginderachter, J.; Movahedi, K.; Menu, E.; Vanderkerken, K. Myeloid-Derived Suppressor Cells in Multiple Myeloma. Blood 2009, 114, 2794, [ASH 2009 51st Meeting]. [Google Scholar] [CrossRef]
- Botta, C.; Gullà, A.; Correale, P.; Tagliaferri, P.; Tassone, P. Myeloid-derived suppressor cells in multiple myeloma: Pre-clinical research and translational opportunities. Front. Oncol. 2014, 4, 348. [Google Scholar] [CrossRef] [PubMed]
- Brimnes, M.K.; Vangsted, A.J.; Knudsen, L.M.; Gimsing, P.; Gang, A.O.; Johnsen, H.E.; Svane, I.M. Increased level of both CD4+FOXP3+ regulatory T cells and CD14+HLA-DR−/low myeloid-derived suppressor cells and decreased level of dendritic cells in patients with multiple myeloma. Scand. J. Immunol. 2010, 72, 540–547. [Google Scholar] [CrossRef]
- Favaloro, J.; Liyadipitiya, T.; Brown, R.; Yang, S.; Suen, H.; Woodland, N.; Nassif, N.; Hart, D.; Fromm, P.; Weatherburn, C.; et al. Myeloid derived suppressor cells are numerically, functionally and phenotypically different in patients with multiple myeloma. Leuk. Lymphoma 2014, 55, 2893–2900. [Google Scholar] [CrossRef] [PubMed]
- Görgün, G.T.; Ohguchi, H.; Hideshima, T.; Tai, Y.-T.; Raje, N.; Munshi, N.C.; Richardson, P.G.; Laubach, J.P.; Anderson, K.C. Inhibition Of Myeloid Derived Suppressor Cells (MDSC) in The Multiple Myeloma Bone Marrow Microenvironment. Blood 2013, 122, 3089, [ASH 2013 55th Meeting]. [Google Scholar] [CrossRef]
- Leone, P.; Berardi, S.; Frassanito, M.A.; Ria, R.; De Re, V.; Cicco, S.; Battaglia, S.; Ditonno, P.; Dammacco, F.; Vacca, A.; et al. Dendritic cells accumulate in the bone marrow of myeloma patients where they protect tumor plasma cells from CD8+ T-cell killing. Blood 2015, 126, 1443–1451. [Google Scholar] [CrossRef] [Green Version]
- Do, T.H.; Johnsen, H.E.; Kjærsgaard, E.; Taaning, E.; Svane, I.M. Impaired circulating myeloid DCs from myeloma patients. Cytotherapy 2004, 6, 196–203. [Google Scholar] [CrossRef]
- Terpos, E.; Zamagni, E.; Lentzsch, S.; Drake, M.T.; García-Sanz, R.; Abildgaard, N.; Ntanasis-Stathopoulos, I.; Schjesvold, F.; de la Rubia, J.; Kyriakou, C.; et al. Treatment of multiple myeloma-related bone disease: Recommendations from the Bone Working Group of the International Myeloma Working Group. Lancet. Oncol. 2021, 22, e119–e130. [Google Scholar] [CrossRef]
- Cremers, S.; Drake, M.T.; Ebetino, F.H.; Bilezikian, J.P.; Russell, R.G.G. Pharmacology of bisphosphonates. Br. J. Clin. Pharmacol. 2019, 85, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Morgan, G.J.; Davies, F.E.; Gregory, W.M.; Cocks, K.; Bell, S.E.; Szubert, A.J.; Navarro-Coy, N.; Drayson, M.T.; Owen, R.G.; Feyler, S.; et al. First-line treatment with zoledronic acid as compared with clodronic acid in multiple myeloma (MRC Myeloma IX): A randomised controlled trial. Lancet (Lond. Engl.) 2010, 376, 1989–1999. [Google Scholar] [CrossRef]
- Hanley, D.A.; Adachi, J.D.; Bell, A.; Brown, V. Denosumab: Mechanism of action and clinical outcomes. Int. J. Clin. Pract. 2012, 66, 1139–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raje, N.; Terpos, E.; Willenbacher, W.; Shimizu, K.; García-Sanz, R.; Durie, B.; Legieć, W.; Krejčí, M.; Laribi, K.; Zhu, L.; et al. Denosumab versus zoledronic acid in bone disease treatment of newly diagnosed multiple myeloma: An international, double-blind, double-dummy, randomised, controlled, phase 3 study. Lancet Oncol. 2018, 19, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Raje, N.; Croucher, P.; Garcia-Sanz, R.; Leleu, X.; Pasteiner, W.; Wang, Y.; Glennane, A.; Canon, J.; Pawlyn, C. Denosumab compared with zoledronic acid on PFS in multiple myeloma: Exploratory results of an international phase 3 study. Blood Adv. 2021, 5, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ding, Z.-C.; Cao, Y.; Liu, C.; Habtetsion, T.; Yu, M.; Lemos, H.; Salman, H.; Xu, H.; Mellor, A.L.; et al. Alkylating Agent Melphalan Augments the Efficacy of Adoptive Immunotherapy Using Tumor-Specific CD4+ T Cells. J. Immunol. 2015, 194, 2011–2021. [Google Scholar] [CrossRef] [Green Version]
- de Magalhães, R.J.P.; Vidriales, M.B.; Paiva, B.; Fernandez-Gimenez, C.; García-Sanz, R.; Mateos, M.V.; Gutierrez, N.C.; Lecrevisse, Q.; Blanco, J.F.; Hernández, J.; et al. Analysis of the immune system of multiple myeloma patients achieving long-term disease control by multidimensional flow cytometry. Haematologica 2013, 98, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteche-López, A.; Kreutzman, A.; Alegre, A.; Sanz Martín, P.; Aguado, B.; González-Pardo, M.; Espiño, M.; Villar, L.M.; García Belmonte, D.; De La Cámara, R.; et al. Multiple myeloma patients in long-term complete response after autologous stem cell transplantation express a particular immune signature with potential prognostic implication. Bone Marrow Transpl. 2017, 52, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Swan, D.; Gurney, M.; Krawczyk, J.; Ryan, A.E.; O’Dwyer, M. Beyond DNA Damage: Exploring the Immunomodulatory Effects of Cyclophosphamide in Multiple Myeloma. HemaSphere 2020, 4, e350. [Google Scholar] [CrossRef]
- Barilà, G.; Pavan, L.; Vedovato, S.; Berno, T.; Lo Schirico, M.; Arangio Febbo, M.; Teramo, A.; Calabretto, G.; Vicenzetto, C.; Gasparini, V.R.; et al. Treatment Induced Cytotoxic T-Cell Modulation in Multiple Myeloma Patients. Front. Oncol. 2021, 11, 682658. [Google Scholar] [CrossRef]
- Fricker, L.D. Proteasome Inhibitor Drugs. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 457–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-L.; Hsu, Y.-T.; Wu, C.-C.; Yang, Y.-C.; Wang, C.; Wu, T.-C.; Hung, C.-F. Immune mechanism of the antitumor effects generated by bortezomib. J. Immunol. 2012, 189, 3209–3220. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, M.; Di Pietro, G.; Ria, R.; Gnoni, A.; Mangialardi, G.; Guarini, A.; Ditonno, P.; Musto, P.; D’Auria, F.; Ricciardi, M.R.; et al. Bortezomib and zoledronic acid on angiogenic and vasculogenic activities of bone marrow macrophages in patients with multiple myeloma. Eur. J. Cancer 2010, 46, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Schwestermann, J.; Besse, A.; Driessen, C.; Besse, L. Contribution of the Tumor Microenvironment to Metabolic Changes Triggering Resistance of Multiple Myeloma to Proteasome Inhibitors. Front. Oncol. 2022, 12, 899272. [Google Scholar] [CrossRef]
- Zhou, Q.; Liang, J.; Yang, T.; Liu, J.; Li, B.; Li, Y.; Fan, Z.; Wang, W.; Chen, W.; Yuan, S.; et al. Carfilzomib modulates tumor microenvironment to potentiate immune checkpoint therapy for cancer. EMBO Mol. Med. 2022, 14, e14502. [Google Scholar] [CrossRef]
- Muz, B.; Ghazarian, R.N.; Ou, M.; Luderer, M.J.; Kusdono, H.D.; Azab, A.K. Spotlight on ixazomib: Potential in the treatment of multiple myeloma. Drug Des. Devel. Ther. 2016, 10, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krönke, J.; Udeshi, N.D.; Narla, A.; Grauman, P.; Hurst, S.N.; McConkey, M.; Svinkina, T.; Heckl, D.; Comer, E.; Li, X.; et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science 2014, 343, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Bjorklund, C.C.; Lu, L.; Kang, J.; Hagner, P.R.; Havens, C.G.; Amatangelo, M.; Wang, M.; Ren, Y.; Couto, S.; Breider, M.; et al. Rate of CRL4(CRBN) substrate Ikaros and Aiolos degradation underlies differential activity of lenalidomide and pomalidomide in multiple myeloma cells by regulation of c-Myc and IRF4. Blood Cancer J. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, J.Y.; Labarthe, M.C.; Meyer, B.; Dasgupta, P.; Dalgleish, A.G.; Galustian, C. Enhanced cross-priming of naive CD8+ T cells by dendritic cells treated by the IMiDs® immunomodulatory compounds lenalidomide and pomalidomide. Immunology 2013, 139, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Vo, M.C.; Anh-NguyenThi, T.; Lee, H.J.; Nguyen-Pham, T.N.; Jaya Lakshmi, T.; Jung, S.H.; Kim, H.J.; Lee, J.J. Lenalidomide enhances the function of dendritic cells generated from patients with multiple myeloma. Exp. Hematol. 2017, 46, 48–55. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kitawaki, T.; Sugimoto, N.; Fujita, H.; Kawase, Y.; Takaori-Kondo, A.; Kadowaki, N. Anti-inflammatory modulation of human myeloid-derived dendritic cell subsets by lenalidomide. Immunol. Lett. 2019, 211, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Vescovini, R.; Bolzoni, M.; Marchica, V.; Storti, P.; Toscani, D.; Accardi, F.; Notarfranchi, L.; Palma, B.D.; Manferdini, C.; et al. Lenalidomide increases human dendritic cell maturation in multiple myeloma patients targeting monocyte differentiation and modulating mesenchymal stromal cell inhibitory properties. Oncotarget 2017, 8, 53053–53067. [Google Scholar] [CrossRef] [PubMed]
- Cytlak, U.; Resteu, A.; Bogaert, D.; Kuehn, H.S.; Altmann, T.; Gennery, A.; Jackson, G.; Kumanovics, A.; Voelkerding, K.V.; Prader, S.; et al. Ikaros family zinc finger 1 regulates dendritic cell development and function in humans. Nat. Commun. 2018, 9, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, R.M.; Chunder, N.; Chen, C.; Umetsu, S.E.; Winandy, S.; Wells, A.D. Ikaros enforces the costimulatory requirement for IL2 gene expression and is required for anergy induction in CD4+ T lymphocytes. J. Immunol. 2007, 179, 7305–7315. [Google Scholar] [CrossRef] [Green Version]
- Galustian, C.; Meyer, B.; Labarthe, M.C.; Dredge, K.; Klaschka, D.; Henry, J.; Todryk, S.; Chen, R.; Muller, G.; Stirling, D.; et al. The anti-cancer agents lenalidomide and pomalidomide inhibit the proliferation and function of T regulatory cells. Cancer Immunol. Immunother. 2009, 58, 1033–1045. [Google Scholar] [CrossRef]
- Kuwahara-Ota, S.; Shimura, Y.; Steinebach, C.; Isa, R.; Yamaguchi, J.; Nishiyama, D.; Fujibayashi, Y.; Takimoto-Shimomura, T.; Mizuno, Y.; Matsumura-Kimoto, Y.; et al. Lenalidomide and pomalidomide potently interfere with induction of myeloid-derived suppressor cells in multiple myeloma. Br. J. Haematol. 2020, 191, 784–795. [Google Scholar] [CrossRef]
- Davies, F.E.; Raje, N.; Hideshima, T.; Lentzsch, S.; Young, G.; Tai, Y.T.; Lin, B.; Podar, K.; Gupta, D.; Chauhan, D.; et al. Thalidomide and immunomodulatory derivatives augment natural killer cell cytotoxicity in multiple myeloma. Blood 2001, 98, 210–216. [Google Scholar] [CrossRef]
- Pierceall, W.E.; Amatangelo, M.D.; Bahlis, N.J.; Siegel, D.S.; Rahman, A.; van Oekelen, O.; Neri, P.; Young, M.; Chung, W.; Serbina, N.; et al. Immunomodulation in Pomalidomide, Dexamethasone, and Daratumumab-Treated Patients with Relapsed/Refractory Multiple Myeloma. Clin. Cancer Res. 2020, 26, 5895–5902. [Google Scholar] [CrossRef]
- Perumal, D.; Imai, N.; Lagana, A.; Finnigan, J.; Melnekoff, D.; Leshchenko, V.V.; Solovyov, A.; Madduri, D.; Chari, A.; Cho, H.J.; et al. Mutation-derived Neoantigen-specific T-cell Responses in Multiple Myeloma. Clin. Cancer Res. 2020, 26, 450–464. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, M.; Innorcia, S.; Boccadoro, M.; Bringhen, S. Monoclonal antibodies to treat multiple myeloma: A dream come true. Int. J. Mol. Sci. 2020, 21, 8192. [Google Scholar] [CrossRef]
- Morandi, F.; Horenstein, A.L.; Costa, F.; Giuliani, N.; Pistoia, V.; Malavasi, F. CD38: A Target for Immunotherapeutic Approaches in Multiple Myeloma. Front. Immunol. 2018, 9, 2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, P.; van de Donk, N.W.C.J.; Miguel, J.S.; Lokhorst, H.; Nahi, H.; Ben-Yehuda, D.; Cavo, M.; Cook, G.; Delforge, M.; Einsele, H.; et al. Practical Considerations for the Use of Daratumumab, a Novel CD38 Monoclonal Antibody, in Myeloma. Drugs 2016, 76, 853–867. [Google Scholar] [CrossRef]
- Zhu, C.; Song, Z.; Wang, A.; Srinivasan, S.; Yang, G.; Greco, R.; Theilhaber, J.; Shehu, E.; Wu, L.; Yang, Z.Y.; et al. Isatuximab Acts Through Fc-Dependent, Independent, and Direct Pathways to Kill Multiple Myeloma Cells. Front. Immunol. 2020, 11, 1771. [Google Scholar] [CrossRef] [PubMed]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.C.J.; Weiss, B.M.; et al. Daratumumab depletes CD38+ immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, H.C.; Stevenaert, F.; Krejcik, J.; Van der Borght, K.; Smets, T.; Bald, J.; Abraham, Y.; Ceulemans, H.; Chiu, C.; Vanhoof, G.; et al. High-Parameter Mass Cytometry Evaluation of Relapsed/Refractory Multiple Myeloma Patients Treated with Daratumumab Demonstrates Immune Modulation as a Novel Mechanism of Action. Cytom. Part A 2019, 95A, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Casneuf, T.; Xu, X.S.; Adams, H.C.; Axel, A.E.; Chiu, C.; Khan, I.; Ahmadi, T.; Yan, X.; Lonial, S.; Plesner, T.; et al. Effects of daratumumab on natural killer cells and impact on clinical outcomes in relapsed or refractory multiple myeloma. Blood Adv. 2017, 1, 2105–2114. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Y.; Hughes, T.; Zhang, J.; Caligiuri, M.A.; Benson, D.M.; Yu, J. Fratricide of NK Cells in Daratumumab Therapy for Multiple Myeloma Overcome by Ex Vivo-Expanded Autologous NK Cells. Clin. Cancer Res. 2018, 24, 4006–4017. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Zhang, L.; Acharya, C.; An, G.; Wen, K.; Qiu, L.; Munshi, N.C.; Tai, Y.T.; Anderson, K.C. Targeting CD38 Suppresses Induction and Function of T Regulatory Cells to Mitigate Immunosuppression in Multiple Myeloma. Clin. Cancer Res. 2017, 23, 4290–4300. [Google Scholar] [CrossRef] [Green Version]
- Casneuf, T.; Adams, H.C.; van de Donk, N.W.C.J.; Abraham, Y.; Bald, J.; Vanhoof, G.; Van der Borght, K.; Smets, T.; Foulk, B.; Nielsen, K.C.; et al. Deep immune profiling of patients treated with lenalidomide and dexamethasone with or without daratumumab. Leukemia 2021, 35, 573–584. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Oriol, A.; Nahi, H.; San-Miguel, J.; Bahlis, N.J.; Usmani, S.Z.; Rabin, N.; Orlowski, R.Z.; Komarnicki, M.; Suzuki, K.; et al. Daratumumab, Lenalidomide, and Dexamethasone for Multiple Myeloma. N. Engl. J. Med. 2016, 375, 1319–1331. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Toscani, D.; Chillemi, A.; Quarona, V.; Bolzoni, M.; Marchica, V.; Vescovini, R.; Mancini, C.; Martella, E.; Campanini, N.; et al. Expression of CD38 in myeloma bone niche: A rational basis for the use of anti-CD38 immunotherapy to inhibit osteoclast formation. Oncotarget 2017, 8, 56598–56611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Iqbal, J.; Dolgilevich, S.; Yuen, T.; Wu, X.; Moonga, B.S.; Adebanjo, O.A.; Bevis, P.J.R.; Lund, F.; Huang, C.L.-H.; et al. Disordered osteoclast formation and function in a CD38 (ADP-ribosyl cyclase)-deficient mouse establishes an essential role for CD38 in bone resorption. FASEB J. 2003, 17, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Bolzoni, M.; Toscani, D.; Costa, F.; Vicario, E.; Aversa, F.; Giuliani, N. The link between bone microenvironment and immune cells in multiple myeloma: Emerging role of CD38. Immunol. Lett. 2019, 205, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Saltarella, I.; Desantis, V.; Melaccio, A.; Solimando, A.G.; Lamanuzzi, A.; Ria, R.; Storlazzi, C.T.; Mariggiò, M.A.; Vacca, A.; Frassanito, M.A. Mechanisms of Resistance to Anti-CD38 Daratumumab in Multiple Myeloma. Cells 2020, 9, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambhampati, S.; Wen, K.W.; Sung, V.; Wong, S.W.K.; Martin, T.G.; Wolf, J.L.; Shah, N.; Wiita, A. Investigating macrophage function as a mechanism of resistance to daratumumab in relapsed refractory multiple myeloma patients. J. Clin. Oncol. 2020, 38, e20547, [ASCO 2020 Annual Meeting]. [Google Scholar] [CrossRef]
- Fotiou, D.; Gavriatopoulou, M.; Ntanasis-Stathopoulos, I.; Migkou, M.; Kanellias, N.; Malandrakis, P.; Theodorakakou, F.; Dialoupi, I.; Eleutherakis-Papaiakovou, E.; Roussou, M.; et al. The Addition of IMiDs for Patients with Daratumumab-Refractory Multiple Myeloma Can Overcome Refractoriness to Both Agents. Blood 2020, 136, 21, [ASH 2020 62nd Meeting]. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Lonial, S.; White, D.; Moreau, P.; Weisel, K.; San-Miguel, J.; Shpilberg, O.; Grosicki, S.; Špička, I.; Walter-Croneck, A.; et al. Elotuzumab, lenalidomide, and dexamethasone in RRMM: Final overall survival results from the phase 3 randomized ELOQUENT-2 study. Blood Cancer J. 2020, 10, 91. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Dytfeld, D.; Grosicki, S.; Moreau, P.; Takezako, N.; Hori, M.; Leleu, X.; LeBlanc, R.; Suzuki, K.; Raab, M.S.; et al. Elotuzumab Plus Pomalidomide and Dexamethasone for Relapsed/Refractory Multiple Myeloma: Final Overall Survival Analysis From the Randomized Phase II ELOQUENT-3 Trial. J. Clin. Oncol. 2022, 379, 1811–1822. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Richardson, P.G.; Bahlis, N.J.; Grosicki, S.; Cavo, M.; Beksaç, M.; Legieć, W.; Liberati, A.M.; Goldschmidt, H.; Belch, A.; et al. Addition of elotuzumab to lenalidomide and dexamethasone for patients with newly diagnosed, transplantation ineligible multiple myeloma (ELOQUENT-1): An open-label, multicentre, randomised, phase 3 trial. Lancet Haematol. 2022, 9, e403–e414. [Google Scholar] [CrossRef]
- Collins, S.M.; Bakan, C.E.; Swartzel, G.D.; Hofmeister, C.C.; Efebera, Y.A.; Kwon, H.; Starling, G.C.; Ciarlariello, D.; Bhaskar, S.; Briercheck, E.L.; et al. Elotuzumab directly enhances NK cell cytotoxicity against myeloma via CS1 ligation: Evidence for augmented NK cell function complementing ADCC. Cancer Immunol. Immunother. 2013, 62, 1841–1849. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.S.; Cohen, A.D.; Pazina, T. Mechanisms of NK Cell Activation and Clinical Activity of the Therapeutic SLAMF7 Antibody, Elotuzumab in Multiple Myeloma. Front. Immunol. 2018, 9, 2551. [Google Scholar] [CrossRef] [Green Version]
- Awwad, M.H.S.; Mahmoud, A.; Bruns, H.; Echchannaoui, H.; Kriegsmann, K.; Lutz, R.; Raab, M.S.; Bertsch, U.; Munder, M.; Jauch, A.; et al. Selective elimination of immunosuppressive T cells in patients with multiple myeloma. Leukemia 2021, 35, 2602–2615. [Google Scholar] [CrossRef] [PubMed]
- Azuma, Y.; Ito, T.; Inaba, M.; Imai, K.; Hotta, M.; Satake, A.; Nomura, S. Elotuzumab Enhances the Th2-Mediated Immune Response of Dendritic Cell Induced by Immunomodulatory Drugs (IMiDs). Blood 2019, 134, 4342, [ASH 2019 61st Meeting]. [Google Scholar] [CrossRef]
- Schlaweck, S.; Strauss, L.; Daecke, S.; Brossart, P.; Heine, A. Elotuzumab spares dendritic cell integrity and functionality. J. Cancer Res. Clin. Oncol. 2021, 147, 2167–2170. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.V.; Blacklock, H.; Schjesvold, F.; Oriol, A.; Simpson, D.; George, A.; Goldschmidt, H.; Larocca, A.; Chanan-Khan, A.; Sherbenou, D.; et al. Pembrolizumab plus pomalidomide and dexamethasone for patients with relapsed or refractory multiple myeloma (KEYNOTE-183): A randomised, open-label, phase 3 trial. Lancet Haematol. 2019, 6, e459–e469. [Google Scholar] [CrossRef]
- Usmani, S.Z.; Schjesvold, F.; Oriol, A.; Karlin, L.; Cavo, M.; Rifkin, R.M.; Yimer, H.A.; LeBlanc, R.; Takezako, N.; McCroskey, R.D.; et al. Pembrolizumab plus lenalidomide and dexamethasone for patients with treatment-naive multiple myeloma (KEYNOTE-185): A randomised, open-label, phase 3 trial. Lancet Haematol. 2019, 6, e448–e458. [Google Scholar] [CrossRef]
- Fares, C.M.; Van Allen, E.M.; Drake, C.G.; Allison, J.P.; Hu-Lieskovan, S. Mechanisms of Resistance to Immune Checkpoint Blockade: Why Does Checkpoint Inhibitor Immunotherapy Not Work for All Patients? Am. Soc. Clin. Oncol. Educ. Book 2019, 39, 147–164. [Google Scholar] [CrossRef]
- Costello, C. The future of checkpoint inhibition in multiple myeloma? Lancet Haematol. 2019, 6, e439–e440. [Google Scholar] [CrossRef]
- Oliva, S.; Troia, R.; D’Agostino, M.; Boccadoro, M.; Gay, F. Promises and Pitfalls in the Use of PD-1/PD-L1 Inhibitors in Multiple Myeloma. Front. Immunol. 2018, 9, 2749. [Google Scholar] [CrossRef] [Green Version]
- Le Calvez, B.; Moreau, P.; Touzeau, C. Immune checkpoint inhibitors for the treatment of myeloma: Novel investigational options. Expert Opin. Investig. Drugs 2021, 30, 965–973. [Google Scholar] [CrossRef]
- Kleber, M.; Ntanasis-Stathopoulos, I.; Terpos, E. BCMA in Multiple Myeloma-A Promising Key to Therapy. J. Clin. Med. 2021, 10, 4088. [Google Scholar] [CrossRef] [PubMed]
- Lonial, S.; Lee, H.C.; Badros, A.; Trudel, S.; Nooka, A.K.; Chari, A.; Abdallah, A.O.; Callander, N.; Lendvai, N.; Sborov, D.; et al. Belantamab mafodotin for relapsed or refractory multiple myeloma (DREAMM-2): A two-arm, randomised, open-label, phase 2 study. Lancet Oncol. 2020, 21, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Trudel, S.; Lendvai, N.; Popat, R.; Voorhees, P.M.; Reeves, B.; Libby, E.N.; Richardson, P.G.; Anderson, L.D.; Sutherland, H.J.; Yong, K.; et al. Targeting B-cell maturation antigen with GSK2857916 antibody–drug conjugate in relapsed or refractory multiple myeloma (BMA117159): A dose escalation and expansion phase 1 trial. Lancet Oncol. 2018, 19, 1641–1653. [Google Scholar] [CrossRef]
- Laurent, S.A.; Hoffmann, F.S.; Kuhn, P.H.; Cheng, Q.; Chu, Y.; Schmidt-Supprian, M.; Hauck, S.M.; Schuh, E.; Krumbholz, M.; Rübsamen, H.; et al. γ-Secretase directly sheds the survival receptor BCMA from plasma cells. Nat. Commun. 2015, 6, 7333. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, E.; Gillespie, A.; Tang, G.; Ferros, M.; Harutyunyan, N.M.; Vardanyan, S.; Gottlieb, J.; Li, M.; Wang, C.S.; Chen, H.; et al. Soluble B-Cell Maturation Antigen Mediates Tumor-Induced Immune Deficiency in Multiple Myeloma. Clin. Cancer Res. 2016, 22, 3383–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonial, S.; Grosicki, S.; Hus, M.; Song, K.W.; Facon, T.; Callander, N.S.; Ribrag, V.; Uttervall, K.; Quach, H.; Vorobyev, V.I.; et al. Synergistic effects of low-dose belantamab mafodotin in combination with a gamma-secretase inhibitor (nirogacestat) in patients with relapsed/refractory multiple myeloma (RRMM): DREAMM-5 study. J. Clin. Oncol. 2022, 40, 8019, [ASCO 2022 Annual Meeting]. [Google Scholar] [CrossRef]
- Berdeja, J.G.; Madduri, D.; Usmani, S.Z.; Jakubowiak, A.; Agha, M.; Cohen, A.D.; Stewart, A.K.; Hari, P.; Htut, M.; Lesokhin, A.; et al. Ciltacabtagene autoleucel, a B-cell maturation antigen-directed chimeric antigen receptor T-cell therapy in patients with relapsed or refractory multiple myeloma (CARTITUDE-1): A phase 1b/2 open-label study. Lancet (Lond. Engl.) 2021, 398, 314–324. [Google Scholar] [CrossRef]
- Martin, T.; Usmani, S.Z.; Berdeja, J.G.; Jakubowiak, A.; Agha, M.; Cohen, A.D.; Hari, P.; Avigan, D.; Deol, A.; Htut, M.; et al. Updated Results from CARTITUDE-1: Phase 1b/2Study of Ciltacabtagene Autoleucel, a B-Cell Maturation Antigen-Directed Chimeric Antigen Receptor T Cell Therapy, in Patients With Relapsed/Refractory Multiple Myeloma. Blood 2021, 138, 549, Abstract #549 [ASH 2021 63rd Meeting]. [Google Scholar] [CrossRef]
- Munshi, N.C.; Anderson, L.D.; Shah, N.; Madduri, D.; Berdeja, J.; Lonial, S.; Raje, N.; Lin, Y.; Siegel, D.; Oriol, A.; et al. Idecabtagene Vicleucel in Relapsed and Refractory Multiple Myeloma. N. Engl. J. Med. 2021, 384, 705–716. [Google Scholar] [CrossRef]
- Dhodapkar, K.M.; Cohen, A.D.; Kaushal, A.; Garfall, A.L.; Manalo, R.J.; Carr, A.R.; McCachren, S.S.; Stadtmauer, E.A.; Lacey, S.F.; Melenhorst, J.J.; et al. Changes in Bone Marrow Tumor and Immune Cells Correlate with Durability of Remissions Following BCMA CAR T Therapy in Myeloma. Blood Cancer Discov. 2022, OF1–OF12. [Google Scholar] [CrossRef]
- D’Agostino, M.; Raje, N. Anti-BCMA CAR T-cell therapy in multiple myeloma: Can we do better? Leukemia 2020, 34, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Liu, M.; Zhang, Y.; Wang, X. Bispecific T cell engagers: An emerging therapy for management of hematologic malignancies. J. Hematol. Oncol. 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Nooka, A.K.; Moreau, P.; Usmani, S.Z.; Garfall, A.L.; van de Donk, N.W.C.J.; San-Miguel, J.F.; Rocafiguera, A.O.; Chari, A.; Karlin, L.; Mateos, M.-V.; et al. Teclistamab, a B-cell maturation antigen (BCMA) x CD3 bispecific antibody, in patients with relapsed/refractory multiple myeloma (RRMM): Updated efficacy and safety results from MajesTEC-1. J. Clin. Oncol. 2022, 40, 8007, [ASCO 2022 Annual Meeting]. [Google Scholar] [CrossRef]
- Meermeier, E.W.; Welsh, S.J.; Sharik, M.E.; Du, M.T.; Garbitt, V.M.; Riggs, D.L.; Shi, C.X.; Stein, C.K.; Bergsagel, M.; Chau, B.; et al. Tumor burden limits bispecific antibody efficacy through T cell exhaustion averted by concurrent cytotoxic therapy. Blood Cancer Discov. 2021, 2, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.Y.; Minnema, M.C.; Berdeja, J.G.; Oriol, A.; van de Donk, N.W.C.J.; Rodriguez-Otero, P.; Askari, E.; Mateos, M.-V.; Costa, L.J.; Verona, R.I.; et al. Updated Phase 1 Results from MonumenTAL-1: First-in-Human Study of Talquetamab, a G Protein-Coupled Receptor Family C Group 5 Member D x CD3 Bispecific Antibody, in Patients with Relapsed/Refractory Multiple Myeloma. Blood 2021, 138, 158, [ASH 2021 63rd Meeting]. [Google Scholar] [CrossRef]
- Trudel, S.; Cohen, A.D.; Krishnan, A.Y.; Fonseca, R.; Spencer, A.; Berdeja, J.G.; Lesokhin, A.; Forsberg, P.A.; Laubach, J.P.; Costa, L.J.; et al. Cevostamab Monotherapy Continues to Show Clinically Meaningful Activity and Manageable Safety in Patients with Heavily Pre-Treated Relapsed/Refractory Multiple Myeloma (RRMM): Updated Results from an Ongoing Phase I Study. Blood 2021, 138, 157, [ASH 2021 63rd Meeting]. [Google Scholar] [CrossRef]
- Buccione, C.; Fragale, A.; Polverino, F.; Ziccheddu, G.; Aricò, E.; Belardelli, F.; Proietti, E.; Battistini, A.; Moschella, F. Role of interferon regulatory factor 1 in governing Treg depletion, Th1 polarization, inflammasome activation and antitumor efficacy of cyclophosphamide. Int. J. Cancer 2018, 142, 976–987. [Google Scholar] [CrossRef] [Green Version]
- Schiavoni, G.; Sistigu, A.; Valentini, M.; Mattei, F.; Sestili, P.; Spadaro, F.; Sanchez, M.; Lorenzi, S.; D’Urso, M.T.; Belardelli, F.; et al. Cyclophosphamide synergizes with type I interferons through systemic dendritic cell reactivation and induction of immunogenic tumor apoptosis. Cancer Res. 2011, 71, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Okazuka, K.; Ishida, T. Proteasome inhibitors for multiple myeloma. Jpn. J. Clin. Oncol. 2018, 48, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Holstein, S.A.; McCarthy, P.L. Immunomodulatory Drugs in Multiple Myeloma: Mechanisms of Action and Clinical Experience. Drugs 2017, 77, 505–520. [Google Scholar] [CrossRef]
- Haslett, P.A.J.; Corral, L.G.; Albert, M.; Kaplan, G. Thalidomide costimulates primary human t lymphocytes, preferentially inducing proliferation, cytokine production, and cytotoxic responses in the CD8+ subset. J. Exp. Med. 1998, 187, 1885–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magen, H.; Muchtar, E. Elotuzumab: The first approved monoclonal antibody for multiple myeloma treatment. Ther. Adv. Hematol. 2016, 7, 187–195. [Google Scholar] [CrossRef]
- Sun, J.; Park, C.; Guenthner, N.; Gurley, S.; Zhang, L.; Lubben, B.; Adebayo, O.; Bash, H.; Chen, Y.; Maksimos, M.; et al. Tumor-associated macrophages in multiple myeloma: Advances in biology and therapy. J. Immunother. Cancer 2022, 10, e003975. [Google Scholar] [CrossRef] [PubMed]
- Lesokhin, A.M.; Ansell, S.M.; Armand, P.; Scott, E.C.; Halwani, A.; Gutierrez, M.; Millenson, M.M.; Cohen, A.D.; Schuster, S.J.; Lebovic, D.; et al. Nivolumab in Patients With Relapsed or Refractory Hematologic Malignancy: Preliminary Results of a Phase Ib Study. J. Clin. Oncol. 2016, 34, 2698–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateos, M.V.; Orlowski, R.Z.; Ocio, E.M.; Rodríguez-Otero, P.; Reece, D.; Moreau, P.; Munshi, N.; Avigan, D.E.; Siegel, D.S.; Ghori, R.; et al. Pembrolizumab combined with lenalidomide and low-dose dexamethasone for relapsed or refractory multiple myeloma: Phase I KEYNOTE-023 study. Br. J. Haematol. 2019, 186, e117–e121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badros, A.Z.; Hyjek, E.; Ma, N.; Lesokhin, A.M.; Rapoport, A.P.; Kocoglu, M.H.; Lederer, E.; Philip, S.; Lesho, P.; Johnson, A.; et al. Pembrolizumab in Combination with Pomalidomide and Dexamethasone for Relapsed/Refractory Multiple Myeloma (RRMM). Blood 2016, 128, 490, [ASH 2016 58th Meeting]. [Google Scholar] [CrossRef]
- Benson, D.M.; Cohen, A.D.; Jagannath, S.; Munshi, N.C.; Spitzer, G.; Hofmeister, C.C.; Efebera, Y.A.; Andre, P.; Zerbib, R.; Caligiuri, M.A. A Phase I Trial of the Anti-KIR Antibody IPH2101 and Lenalidomide in Patients with Relapsed/Refractory Multiple Myeloma. Clin. Cancer Res. 2015, 21, 4055–4061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, P.; Attal, M.; Hulin, C.; Arnulf, B.; Belhadj, K.; Benboubker, L.; Béné, M.C.; Broijl, A.; Caillon, H.; Caillot, D.; et al. Bortezomib, thalidomide, and dexamethasone with or without daratumumab before and after autologous stem-cell transplantation for newly diagnosed multiple myeloma (CASSIOPEIA): A randomised, open-label, phase 3 study. Lancet 2019, 394, 29–38. [Google Scholar] [CrossRef]
- Voorhees, P.M.; Kaufman, J.L.; Laubach, J.; Sborov, D.W.; Reeves, B.; Rodriguez, C.; Chari, A.; Silbermann, R.; Costa, L.J.; Anderson, L.D.; et al. Daratumumab, lenalidomide, bortezomib, and dexamethasone for transplant-eligible newly diagnosed multiple myeloma: The GRIFFIN trial. Blood 2020, 136, 936–945. [Google Scholar] [CrossRef]
- Mateos, M.-V.; Dimopoulos, M.A.; Cavo, M.; Suzuki, K.; Jakubowiak, A.; Knop, S.; Doyen, C.; Lucio, P.; Nagy, Z.; Kaplan, P.; et al. Daratumumab plus Bortezomib, Melphalan, and Prednisone for Untreated Myeloma. N. Engl. J. Med. 2018, 378, 518–528. [Google Scholar] [CrossRef]
- Facon, T.; Kumar, S.; Plesner, T.; Orlowski, R.Z.; Moreau, P.; Bahlis, N.; Basu, S.; Nahi, H.; Hulin, C.; Quach, H.; et al. Daratumumab plus Lenalidomide and Dexamethasone for Untreated Myeloma. N. Engl. J. Med. 2019, 380, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Chanan-Khan, A.A.; Giralt, S. Importance of achieving a complete response in multiple myeloma, and the impact of novel agents. J. Clin. Oncol. 2010, 28, 2612–2624. [Google Scholar] [CrossRef] [PubMed]
- Munshi, N.C.; Avet-Loiseau, H.; Rawstron, A.C.; Owen, R.G.; Child, J.A.; Thakurta, A.; Sherrington, P.; Samur, M.K.; Georgieva, A.; Anderson, K.C.; et al. Association of Minimal Residual Disease With Superior Survival Outcomes in Patients With Multiple Myeloma: A Meta-analysis. JAMA Oncol. 2017, 3, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Munshi, N.C.; Avet-Loiseau, H.; Anderson, K.C.; Neri, P.; Paiva, B.; Samur, M.; Dimopoulos, M.; Kulakova, M.; Lam, A.; Hashim, M.; et al. A large meta-analysis establishes the role of MRD negativity in long-term survival outcomes in patients with multiple myeloma. Blood Adv. 2020, 4, 5988–5999. [Google Scholar] [CrossRef]
- Kumar, S.; Paiva, B.; Anderson, K.C.; Durie, B.; Landgren, O.; Moreau, P.; Munshi, N.; Lonial, S.; Bladé, J.; Mateos, M.-V.; et al. International Myeloma Working Group consensus criteria for response and minimal residual disease assessment in multiple myeloma. Lancet Oncol. 2016, 17, e328–e346. [Google Scholar] [CrossRef]
- Flores-Montero, J.; Sanoja-Flores, L.; Paiva, B.; Puig, N.; García-Sánchez, O.; Böttcher, S.; Van Der Velden, V.H.J.; Pérez-Morán, J.J.; Vidriales, M.B.; García-Sanz, R.; et al. Next Generation Flow for highly sensitive and standardized detection of minimal residual disease in multiple myeloma. Leukemia 2017, 31, 2094–2103. [Google Scholar] [CrossRef] [Green Version]
- Oliva, S.; Genuardi, E.; Petrucci, M.T.; D’Agostino, M.; Auclair, D.; Spadano, A.; Jacob, A.P.; Cea, M.; De Rosa, L.; Gozzetti, A.; et al. Impact of Minimal Residual Disease (MRD) By Multiparameter Flow Cytometry (MFC) and Next-Generation Sequencing (NGS) on Outcome: Results of Newly Diagnosed Transplant-Eligible Multiple Myeloma (MM) Patients Enrolled in the Forte Trial. Blood 2020, 136, 44–45, [Abstract #491, ASH 2020 62nd Meeting]. [Google Scholar] [CrossRef]
- Avet-Loiseau, H.; Bene, M.C.; Wuilleme, S.; Corre, J.; Attal, M.; Arnulf, B.; Garderet, L.; Macro, M.; Stoppa, A.-M.; Delforge, M.; et al. Concordance of Post-consolidation Minimal Residual Disease Rates by Multiparametric Flow Cytometry and Next-generation Sequencing in CASSIOPEIA. Clin. Lymphoma Myeloma Leuk. 2019, 19, e3–e4, [Abstract #OAB–004, 17th IMW 2019]. [Google Scholar] [CrossRef]
- Medina, A.; Puig, N.; Flores-Montero, J.; Jimenez, C.; Sarasquete, M.E.; Garcia-Alvarez, M.; Prieto-Conde, I.; Chillon, C.; Alcoceba, M.; Gutierrez, N.C.; et al. Comparison of next-generation sequencing (NGS) and next-generation flow (NGF) for minimal residual disease (MRD) assessment in multiple myeloma. Blood Cancer J. 2020, 10, 108. [Google Scholar] [CrossRef]
- Martinez-Lopez, J.; Lahuerta, J.J.; Pepin, F.; González, M.; Barrio, S.; Ayala, R.; Puig, N.; Montalban, M.A.; Paiva, B.; Weng, L.; et al. Prognostic value of deep sequencing method for minimal residual disease detection in multiple myeloma. Blood 2014, 123, 3073–3079. [Google Scholar] [CrossRef] [Green Version]
- Perrot, A.; Lauwers-Cances, V.; Corre, J.; Robillard, N.; Hulin, C.; Chretien, M.L.; Dejoie, T.; Maheo, S.; Stoppa, A.M.; Pegourie, B.; et al. Minimal residual disease negativity using deep sequencing is a major prognostic factor in multiple myeloma. Blood 2018, 132, 2456–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San-Miguel, J.; Avet-Loiseau, H.; Paiva, B.; Kumar, S.; Dimopoulos, M.A.; Facon, T.; Mateos, M.V.; Touzeau, C.; Jakubowiak, A.; Usmani, S.Z.; et al. Sustained minimal residual disease negativity in newly diagnosed multiple myeloma and the impact of daratumumab in MAIA and ALCYONE. Blood 2022, 139, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Gay, F.; Musto, P.; Rota Scalabrini, D.; Galli, M.; Belotti, A.; Zamagni, E.; Bertamini, L.; Zambello, R.; Quaresima, M.; De Sabbata, G.; et al. Survival Analysis of Newly Diagnosed Transplant-Eligible Multiple Myeloma Patients in the Randomized Forte Trial. Blood 2020, 136, 35–37, [Abstract #141, ASH 2020 62nd Meeting]. [Google Scholar] [CrossRef]
- Costa, L.J.; Chhabra, S.; Medvedova, E.; Dholaria, B.R.; Schmidt, T.M.; Godby, K.N.; Silbermann, R.; Dhakal, B.; Bal, S.; Giri, S.; et al. Daratumumab, Carfilzomib, Lenalidomide, and Dexamethasone With Minimal Residual Disease Response-Adapted Therapy in Newly Diagnosed Multiple Myeloma. J. Clin. Oncol. 2022, 40, 2901–2912. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, M.; Oliva, S.; Rota-Scalabrini, D.; Petrucci, M.T.; Zambello, R.; De Sabbata, G.; Liberati, A.M.; Pietrantuono, G.; Tosi, P.; Pisani, F.; et al. Predictors of Unsustained Negativity in Minimal Residual Disease (MRD)-Negative Transplant-Eligible Newly Diagnosed Multiple Myeloma (MM) Patients Enrolled in the FORTE Trial. Clin. Lymphoma Myeloma Leuk. 2022, 22, S6–S7, [Abstract #OAB-011, 19th IMS 2022 Meeting]. [Google Scholar] [CrossRef]
- Goicoechea, I.; Puig, N.; Cedena, M.T.; Burgos, L.; Cordón, L.; Vidriales, M.B.; Flores-Montero, J.; Gutierrez, N.C.; Calasanz, M.J.; Ramos, M.L.M.; et al. Deep MRD profiling defines outcome and unveils different modes of treatment resistance in standard- And high-risk myeloma. Blood 2021, 137, 49–60. [Google Scholar] [CrossRef]
- Paiva, B.; Cedena, M.T.; Puig, N.; Arana, P.; Vidriales, M.B.; Cordon, L.; Flores-Montero, J.; Gutierrez, N.C.; Martín-Ramos, M.L.; Martinez-Lopez, J.; et al. Minimal residual disease monitoring and immune profiling in multiple myeloma in elderly patients. Blood 2016, 127, 3165–3174. [Google Scholar] [CrossRef]
- Barlogie, B.; Mitchell, A.; Van Rhee, F.; Epstein, J.; Morgan, G.J.; Crowley, J. Curing myeloma at last: Defining criteria and providing the evidence. Blood 2014, 124, 3043–3051. [Google Scholar] [CrossRef]
- Coffey, D.G.; Maura, F.; Gonzalez-Kozlova, E.; Diaz-Mejia3, J.; Luo, P.; Zhang, Y.; Xu, Y.; Warren, E.H.; Smith, E.L.; Cho, H.J.; et al. Normalization of the Immune Microenvironment during Lenalidomide Maintenance Is Associated with Sustained MRD Negativity in Patients with Multiple Myeloma. Blood 2021, 138, 329, [ASH 2021 63rd Meeting]. [Google Scholar] [CrossRef]
- Gowda, S.; Silbermann, R.; Schmidt, T.; Dhakal, B.; Bal, S.; Biltibo, E.; Chhabra, S.; Dholaria, B.; Giri, S.; Godby, K.; et al. Humoral Immune Reconstitution After Quadruplet Therapy, Autologous Hematopoietic Cell Transplant (AHCT) and Measurable Residual Disease Adapted Treatment Cessation in Newly Diagnosed Myeloma (NDMM). Clin. Lymphoma Myeloma Leuk. 2022, 22, S15–S16, [Abstract #OAB-026, 19th IMS 2022 Meeting]. [Google Scholar] [CrossRef]
- Maura, F.; Boyle, E.M.; Diamond, B.; Blaney, P.; Ghamlouch, H.; Ziccheddu, B.; Wang, Y.; Maclachlan, K.H.; Hoffman, J.E.; Hassoun, H.; et al. Genomic and Immune Signatures Predict Sustained MRD Negativity in Newly Diagnosed Multiple Myeloma Patients Treated with Daratumumab, Carfilzomib, Lenalidomide, and Dexamethasone (D-KRd). Blood 2021, 138, 325, [ASH 2021 63rd Meeting]. [Google Scholar] [CrossRef]
- Bertamini, L.; Bertuglia, G.; Oliva, S. Beyond Clinical Trials in Patients With Multiple Myeloma: A Critical Review of Real-World Results. Front. Oncol. 2022, 12, 844779. [Google Scholar] [CrossRef] [PubMed]
- Mina, R.; Lonial, S. Is there still a role for stem cell transplantation in multiple myeloma? Cancer 2019, 125, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.K.; Harrison, S.J.; Cavo, M.; de la Rubia, J.; Popat, R.; Gasparetto, C.; Hungria, V.; Salwender, H.; Suzuki, K.; Kim, I.; et al. Venetoclax or placebo in combination with bortezomib and dexamethasone in patients with relapsed or refractory multiple myeloma (BELLINI): A randomised, double-blind, multicentre, phase 3 trial. Lancet Oncol. 2020, 21, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Bonello, F.; Cani, L.; D’Agostino, M. Risk Stratification Before and During Treatment in Newly Diagnosed Multiple Myeloma: From Clinical Trials to the Real-World Setting. Front. Oncol. 2022, 12, 830922. [Google Scholar] [CrossRef]
- Mina, R.; Bonello, F.; Oliva, S. Minimal Residual Disease in Multiple Myeloma: Ready for Prime Time? Cancer J. (United States) 2021, 27, 247–255. [Google Scholar] [CrossRef]
- Bai, R.; Cui, J. Development of Immunotherapy Strategies Targeting Tumor Microenvironment Is Fiercely Ongoing. Front. Immunol. 2022, 13, 890166. [Google Scholar] [CrossRef]
- Guerrero, C.; Puig, N.; Cedena, M.T.; Goicoechea, I.; Perez, C.; Garcés, J.J.; Botta, C.; Calasanz, M.J.; Gutierrez, N.C.; Martin-Ramos, M.L.; et al. A Machine Learning Model Based on Tumor and Immune Biomarkers to Predict Undetectable MRD and Survival Outcomes in Multiple Myeloma. Clin. Cancer Res. 2022, 28, 2598–2609. [Google Scholar] [CrossRef]
- Pawlyn, C.; Davies, F.E. Predicting the Future: Machine-Based Learning for MRD Prognostication. Clin. Cancer Res. 2022, 28, 2482–2484. [Google Scholar] [CrossRef]
- Tahri, S.; de Jong, M.; Fokkema, C.; Papazian, N.; Kellermayer, Z.; Den Hollander, C.; Vermeulen, M.; van Duin, M.; van de Woestijne, P.; Nasserinejad, K.; et al. Single-cell transcriptomic analysis reveals reduction of cytotoxic NK cells in a subset of newly diagnosed multiple myeloma patients impacting outcome after daratumumab therapy. Clin. Lymphoma Myeloma Leuk. 2022, 22, S17–S18, [Abstract #OAB-029, 19th IMS 2022 Meeting]. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Drug Group | Drug Name | Drug Mechanism | Effect on the ME | ||
|---|---|---|---|---|---|
| Bisphosphonates | Zoledronic acid | Inhibition of FPPS | - Inhibition of OC-mediated bone resorption [91] - Proliferation of M1 phenotype TAMs [103] | ||
| Alkylating agents | Melphalan | Alkylation of DNA at the N7 position of guanine and induction of DNA inter-strand cross-linkages | - Leucodepletion [96] - Myelodepletion - Activation of CD8+ T cells [96] | ||
| Cyclophosphamide | DNA damage (DNA strand breaks) | - Polarization of Th1 [99,167] - Depletion of Tregs [99,167] - Activation of DCs [99,168] - Polarization of the M1 phenotype TAMs and enhancement of ADCP [99] | |||
| Proteasome inhibitors (PIs) | Bortezomib | It binds reversibly to the β 5 subunit of the proteasome [169] | Inhibition of NF-κB pathway [101] SHH signaling pathway activation * | - Recruitment of CD8+-T lymphocytes and IFN-γ+ lymphocytes [103] - Proliferation of M1 phenotype TAMs [103] | |
| Carfilzomib | It binds irreversibly to the β 5 subunit of the proteasome [169] | - Reprogramming of TAMs into M1-like macrophages [105] | |||
| Ixazomib | It binds reversibly to β 5 subunit and β at high concentration of the proteasome [169] | - Decrease in vasculogenesis [106] - Inhibition of OC-mediated bone resorption [106] - Induction of OB activity [106] | |||
| Immunomodulatory drugs (IMiDs) | Thalidomide | Degradation of transcription factors Ikaros and Aiolos through cereblon stabilization [170] | - Induction of CD8+ T-cell cytotoxicity [171] - Increase in NK-cell cytotoxicity [116] | ||
| Lenalidomide | - Induction of CD8+ T-cell cytotoxicity [171] - Inhibition of Treg proliferation [114] - Inhibition of MDSCs [115] - Induction of DC activation [109] | ||||
| Pomalidomide | - Induction of CD8+ T-cell cytotoxicity [171] - Inhibition of Treg proliferation [114] - Inhibition of MDSCs [115] - Induction of DC activation [109] | ||||
| Monoclonal antibodies (mAbs) | Denosumab | Anti-RANKL [93] | - Inhibition of OC activity [94,95] | ||
| Daratumumab | Anti-CD38 | ADCC ADCP CDC [123] | - Inhibition of CD38+ Tregs [124] - Inhibition of CD38+ Bregs [124] - Inhibition of CD38+ MDSCs [124] - Reduction of CD38+ NK cells [124,126,127] - Increase in CD8+ T-cell cytotoxicity [124] - Inhibition of osteoclastogenesis [131,132] | ||
| Isatuximab | Anti-CD38 | ADCC ADCP CDC [124] | - Inhibition of CD38+ Tregs [128] - Inhibition of CD38+ Bregs [128] - Inhibition of CD38+ MDSCs [128] - Reduction of CD38+ NK cells [128] - Increase in CD8+ T-cell cytotoxicity [128] | ||
| Elotuzumab | Anti-SLAMF7 | ADCC ADCP [172] | - Activation of NK cells [141] - Reduction of SLAMF7+ Treg cells [142] - Enhancement of DC activity [143,144] | ||
| Pembrolizumab | Anti-PD-1 | - Activation of Th1 and their cytokine secretion [149] - Inhibition and killing of Tregs [149] | |||
| Antibody–drug conjugates (ADC) | Belantamab mafodotin | Anti-BCMA combined with mafodotin (tubulin inhibitor) | - Disruption of the interaction between the BM ME and MM cells, depending on BCMA [21] | ||
| Chimeric antigen receptor (CAR) T cells | - Idecabtagene vicleucel (ide-cel) - Ciltacabtagene autoleucel (Cilta-cel) | Anti-BCMA | - Activation of cytotoxic T cells [160] - Activation of DCs [160] - Reduction of MDSCs [160] | ||
| Bispecific antibodies (bsAbs) | Teclistamab | Anti-BCMA and CD3 | - Activation of CD3+CD8+ T cells [162] | ||
| Talquetamab | Anti-GPRC5D and CD3 | - Activation of CD3+CD8+ T cells [162] | |||
| Cevostamab | Anti-FcRH5 and CD3 | - Activation of CD3+CD8+ T cells [162] | |||
| Drug | Drug Class and Mechanism | ClinicalTrials.gov Identifier | Study Phase | Description | Status | Results |
|---|---|---|---|---|---|---|
| BI-505 | MAb directed against intercellular adhesion molecule 1 (ICAM-1) | NCT02756728 | I/II | BI-505 in conjunction with ASCT in patients with MM | Terminated due to high risk of cardiopulmonary events | - |
| Pasireotide (SOM230 LAR) | Small somatostatin (SST) analog, interaction with PI3K/MAPK pathway, RANKL, and IGF-1 | NCT04603872 | II | SOM230 LAR in combination with bortezomib and dexamethasone in RRMM patients | Withdrawn before participants enrolled in the trial. | - |
| Sorafenib | Small molecules anti-VEGFR | NCT00536575 | I/II | sorafenib and bortezomib in RRMM patients | Terminated | 15% ORR, lower than preplanned |
| Tremelimumab | MAb, anti-CTLA-4 | NCT02716805 | I | Tremelimumab, durvalumab, and HDC-ASCT in TE patients with MM | Early termination due to safety signals in other studies investigating combination regimens including similar drugs | - |
| Durvalumab | MAb, anti-PD-L1 | |||||
| Durvalumab | MAb, anti-PD-L1 | NCT02685826 | I/II | Durvalumab in combination with lenalidomide with and without dexamethasone in adults with NDMM | Completed | Not available |
| Nivolumab | MAb anti PD-1 | NCT01592370 [174] | 1I/II | Nivolumab as monotherapy or in combination with ipilimumab/lirilumab vs. daratumumab plus pomalidomide and dexamethasone in RRMM patients with ≥2 prior lines of therapy | Completed | Nivolumab monotherapy ORR: 4% (1/27) |
| Nivolumab | MAb anti-PD-1 | NCT02726581 | III | Nivolumab plus pomalidomide and dexamethasone or pomalidomide and dexamethasone or nivolumab plus elotuzumab, pomalidomide, and dexamethasone in RRMM patients with ≥2 prior lines of therapy | Completed | Not available |
| Pembrolizumab | MAb anti-PD-1 | KEYNOTE-023 [175], NCT02036502 | I | Pembrolizumab in combination with lenalidomide and low-dose dexamethasone in RRMM patients | Completed | ORR: 44% (22/50) |
| Pembrolizumab | MAb anti-PD-1 | KEYNOTE-185 [146], NCT02579863 | III | Lenalidomide and dexamethasone plus pembrolizumab vs. lenalidomide and dexamethasone alone | Completed | ORR: 64% (96/151) |
| Pembrolizumab | MAb anti-PD-1 | HP-00061522 [176] NCT02289222 | II | Pembrolizumab plus pomalidomide and dexamethasone in RRMM patients | Completed | ORR: 60% (29/48) |
| Pembrolizumab | MAb anti-PD-1 | KEYNOTE-183 [145] NCT02576977 | III | Pomalidomide plus dexamethasone and pembrolizumab in RRMM patients | Completed | ORR: 34% (43/125) |
| Atezolizumab | MAb, anti-PD-L1 | NCT02431208 | I | Atezolizumab alone or in combination with an IMiD (pomalidomide/lenalidomide) and/or daratumumab in MM patients | Completed | Not available |
| Lirilumab (BMS-986015) | MAb, anti-KIR | NCT02252263 | I | Elotuzumab in combination with either lirilumab (BMS-986015) or urelumab (BMS-663513) in MM patients | Completed | Not available |
| Urelumab (BMS-663513) | MAb, anti-CD137 | |||||
| IPH2101 | MAb, anti-KIR | KIRIMID [177] NCT01217203 | I | IPH2101 and lenalidomide in RRMM patients | Completed | ORR: 33% (5/15) |
| Relatlimab | MAb, anti-LAG-3 | NCT04150965 | I/II | Immuno-oncology drugs elotuzumab, anti-LAG-3, and anti-TIGIT in RRMM patients | Recruiting, ongoing | Not available |
| BMS-986207 | MAb, anti-TIGIT | |||||
| EOS-448 | MAb, anti-TIGIT | NCT05289492 | I/II | EOS884448 alone and in combination with iberdomide with or without dexamethasone in RRMM patients | Recruiting, ongoing | Not available |
| TTI-621 | Fusion protein consisting of the N-terminal domain of human SIRPα linked to a human IgG1 Fc region and of a CD47-blocking innate immune checkpoint | NCT02663518 | Ia/Ib | TTI-621 in patients with RR hematologic malignancies (including MM) and selected solid tumors | Active, not recruiting | Not available |
| TTI-622 | Fusion protein consisting of the N-terminal domain of human SIRPα linked to a human IgG1 Fc region and of a CD47-blocking innate immune checkpoint | NCT03530683 | Ia/Ib | TTI-622 in patients with RRMM, RR lymphoma, and ND acute myeloid leukemia (AML) | Recruiting, not active | Not available |
| SRF231 | MAb, anti-CD47 | NCT03512340 | I/Ib | SRF231 in patients with advanced solid and hematologic cancers (including MM) | Completed | Not Available |
| AO-176 | MAb inhibiting the CD47-SIRPα checkpoint | NCT04445701 | I/II | AO-176 as monotherapy and in combination with bortezomib-dexamethasone in RRMM patients | Active, not recruiting | Not available |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertuglia, G.; Cani, L.; Larocca, A.; Gay, F.; D’Agostino, M. Normalization of the Immunological Microenvironment and Sustained Minimal Residual Disease Negativity: Do We Need Both for Long-Term Control of Multiple Myeloma? Int. J. Mol. Sci. 2022, 23, 15879. https://doi.org/10.3390/ijms232415879
Bertuglia G, Cani L, Larocca A, Gay F, D’Agostino M. Normalization of the Immunological Microenvironment and Sustained Minimal Residual Disease Negativity: Do We Need Both for Long-Term Control of Multiple Myeloma? International Journal of Molecular Sciences. 2022; 23(24):15879. https://doi.org/10.3390/ijms232415879
Chicago/Turabian StyleBertuglia, Giuseppe, Lorenzo Cani, Alessandra Larocca, Francesca Gay, and Mattia D’Agostino. 2022. "Normalization of the Immunological Microenvironment and Sustained Minimal Residual Disease Negativity: Do We Need Both for Long-Term Control of Multiple Myeloma?" International Journal of Molecular Sciences 23, no. 24: 15879. https://doi.org/10.3390/ijms232415879