
Global Metabolomics Discovers Two Novel Biomarkers in Pyridoxine-Dependent Epilepsy Caused by ALDH7A1 Deficiency
 ,
,  , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
2.1. Two New PDE-ALDH7A1 Biomarkers
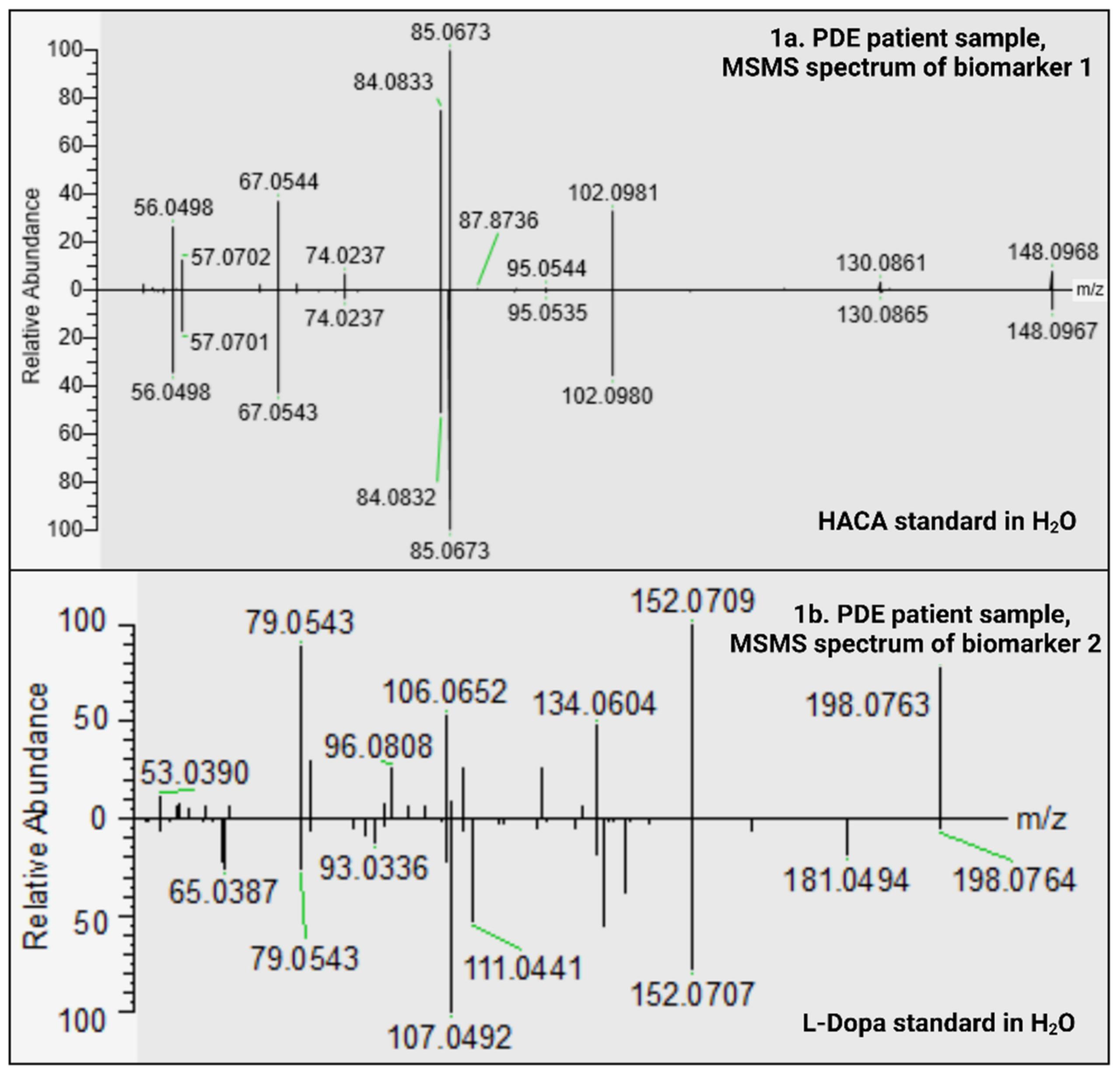
2.1.1. HACA; Level 1 Identification of Biomarker 1
2.1.2. C9H11NO4; Level 3 Identification of Biomarker 2
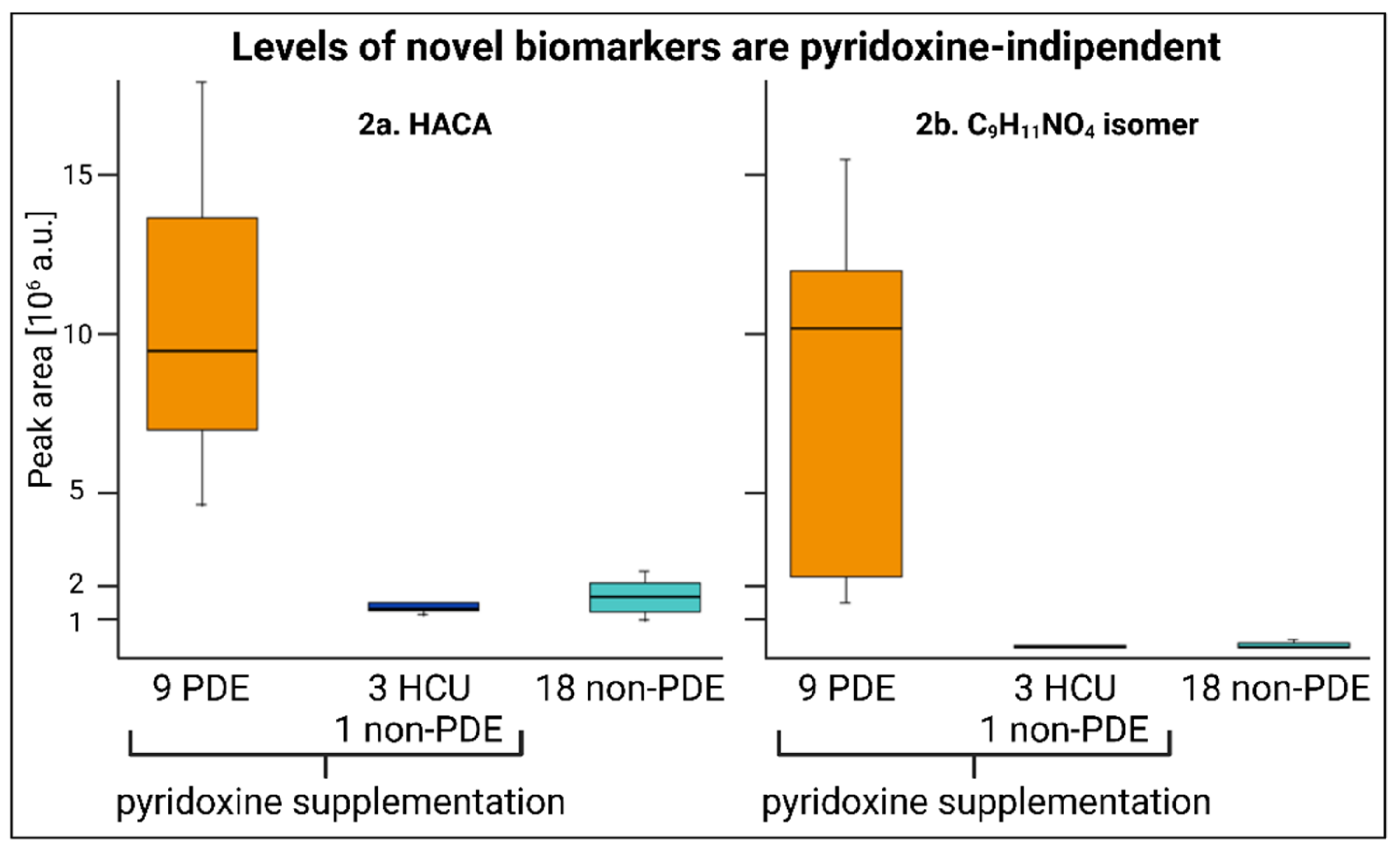
2.2. Pyridoxine-Independent Elevations
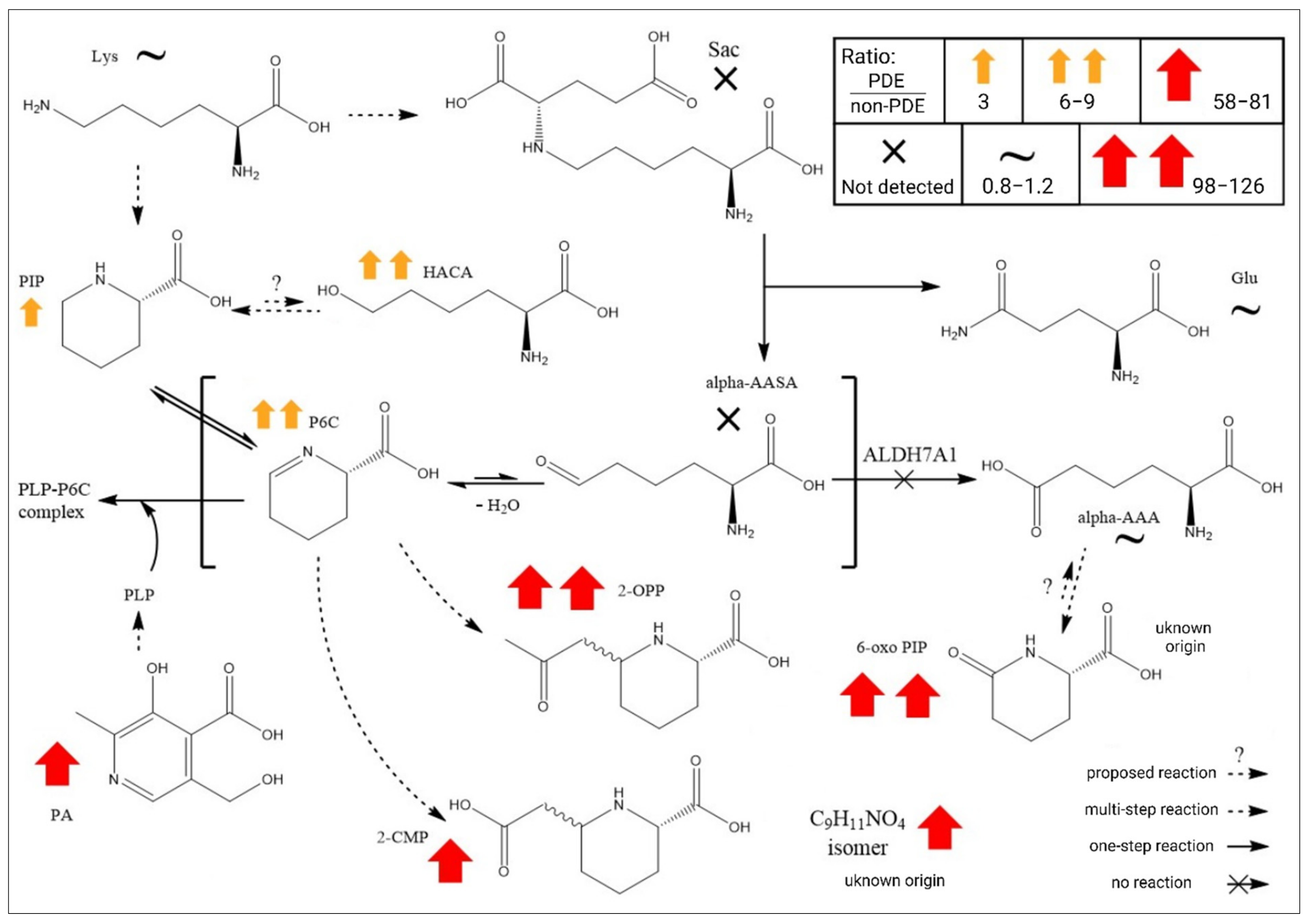
2.3. Global Metabolomics Unfolds the Biochemistry of PDE-ALDH7A1
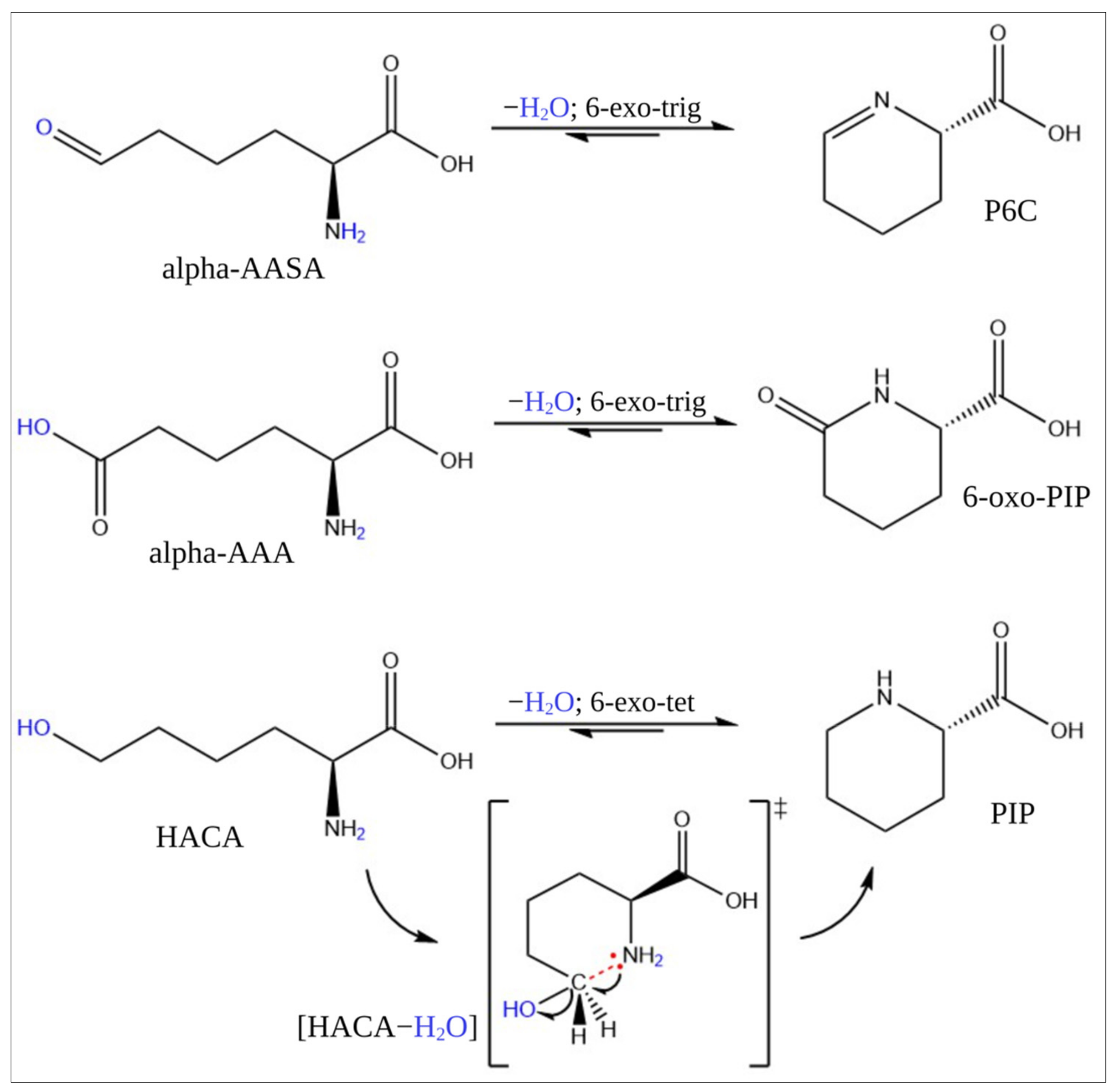
2.4. Stability Test and Conversion of HACA
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Patients and Clinical Details
4.3. Ethics Statement
4.4. Experimental Procedure
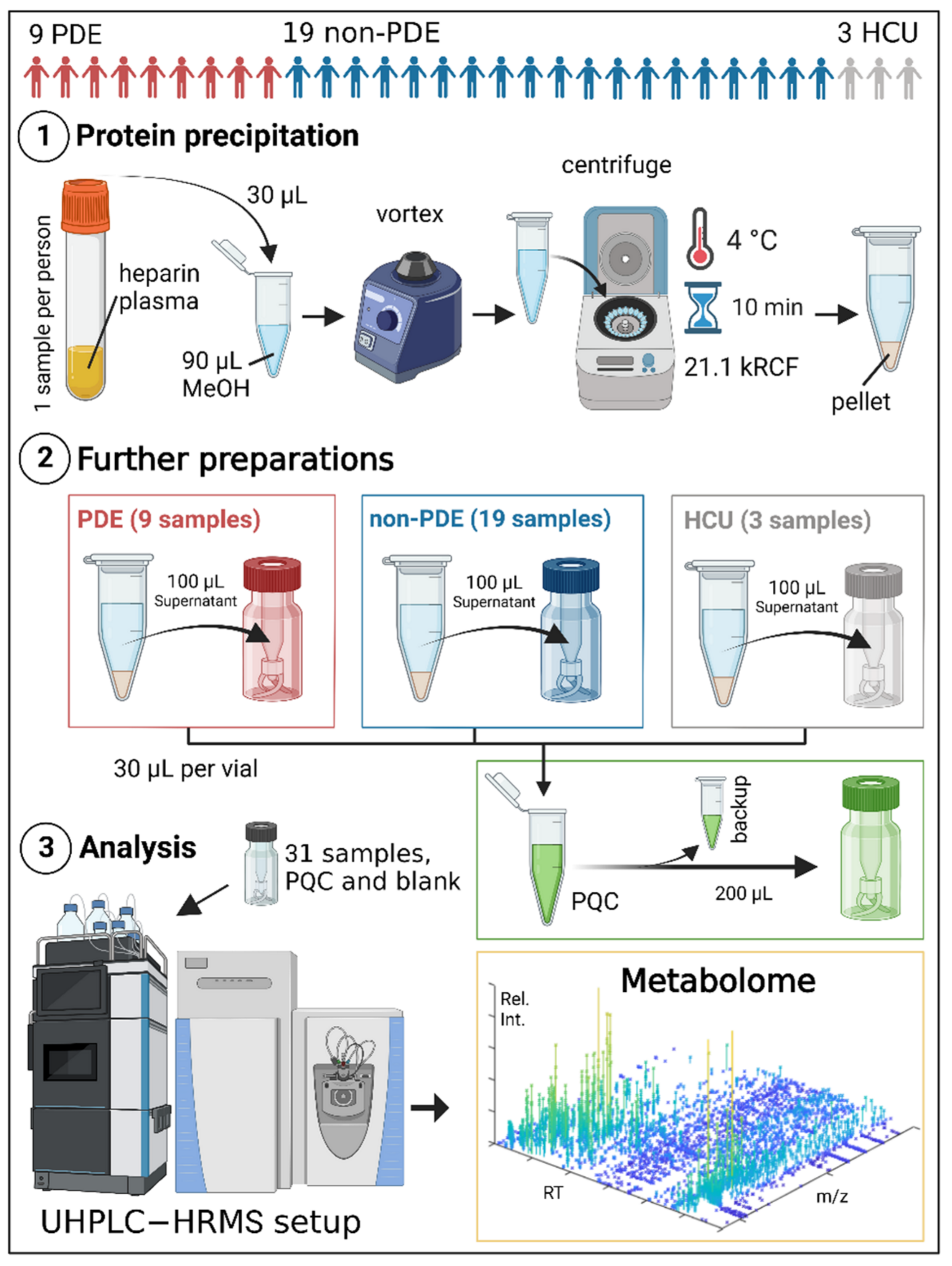
4.4.1. Patients, Controls, and Samples
4.4.2. Sample Preparations and UHPLC-HRMS Analysis
4.4.3. Computer Software
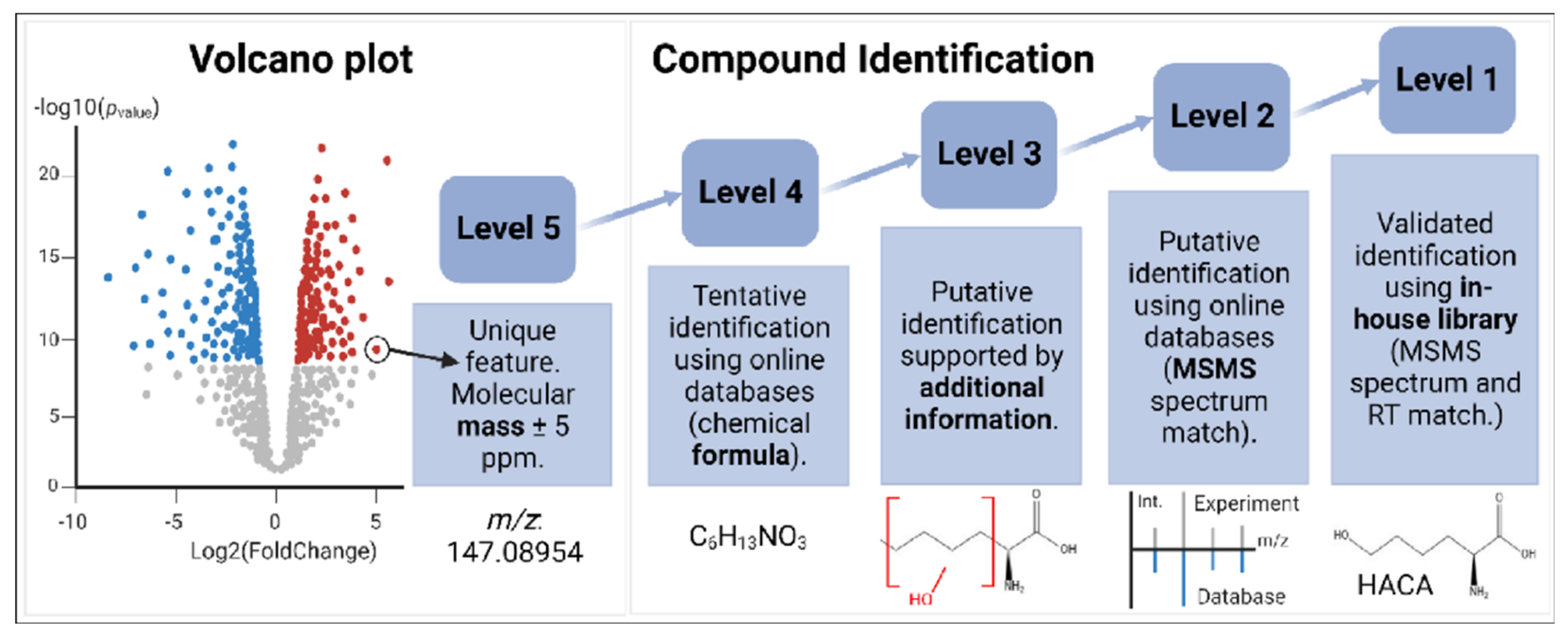
4.5. Levels of Confidence in Compound Identification

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M.P.; Plecko, B.; Mills, P.B.; Clayton, P.T. Disorders affecting vitamin B6 metabolism. J. Inherit. Metab. Dis. 2019, 42, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, M.; Elgstøen, K.B.P. Is Oxidative Stress an Overlooked Player in Pyridoxine-dependent Epilepsy? A Focused Review. Seizure 2021, 91, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Stockler, S.; Plecko, B.; Gospe, S.M., Jr.; Coulter-Mackie, M.; Connolly, M.; Van Karnebeek, C.; Mercimek-Mahmutoglu, S.; Hartmann, H.; Scharer, G.; Struijs, E. Pyridoxine dependent epilepsy and antiquitin deficiency: Clinical and molecular characteristics and recommendations for diagnosis, treatment and follow-up. Mol. Genet. Metab. 2011, 104, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Gospe, S.M., Jr.; Olin, K.L.; Keen, C.L. Reduced GABA synthesis in pyridoxine-dependent seizures. Lancet 1994, 343, 1133–1134. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.B.; Struys, E.; Jakobs, C.; Plecko, B.; Baxter, P.; Baumgartner, M.; Willemsen, M.A.; Omran, H.; Tacke, U.; Uhlenberg, B. Mutations in antiquitin in individuals with pyridoxine-dependent seizures. Nat. Med. 2006, 12, 307–309. [Google Scholar] [CrossRef]
- Coughlin, C.R.; Tseng, L.A.; Abdenur, J.E.; Ashmore, C.; Boemer, F.; Bok, L.A.; Boyer, M.; Buhas, D.; Clayton, P.T.; Das, A. Consensus guidelines for the diagnosis and management of pyridoxine-dependent epilepsy due to α-aminoadipic semialdehyde dehydrogenase deficiency. J. Inherit. Metab. Dis. 2021, 44, 178–192. [Google Scholar] [CrossRef]
- van Karnebeek, C.D.; Hartmann, H.; Jaggumantri, S.; Bok, L.A.; Cheng, B.; Connolly, M.; Coughlin II, C.R.; Das, A.M.; Gospe, S.M., Jr.; Jakobs, C. Lysine restricted diet for pyridoxine-dependent epilepsy: First evidence and future trials. Mol. Genet. Metab. 2012, 107, 335–344. [Google Scholar] [CrossRef]
- Xue, J.; Wang, J.; Gong, P.; Wu, M.; Yang, W.; Jiang, S.; Wu, Y.; Jiang, Y.; Zhang, Y.; Yuzyuk, T. Simultaneous quantification of alpha-aminoadipic semialdehyde, piperideine-6-carboxylate, pipecolic acid and alpha-aminoadipic acid in pyridoxine-dependent epilepsy. Sci. Rep. 2019, 9, 11371. [Google Scholar] [CrossRef]
- Wempe, M.F.; Kumar, A.; Kumar, V.; Choi, Y.J.; Swanson, M.A.; Friederich, M.W.; Hyland, K.; Yue, W.W.; Van Hove, J.L.; Coughlin, C.R. Identification of a novel biomarker for pyridoxine-dependent epilepsy: Implications for newborn screening. J. Inherit. Metab. Dis. 2019, 42, 565–574. [Google Scholar] [CrossRef]
- Engelke, U.F.; Van Outersterp, R.E.; Merx, J.; Van Geenen, F.A.; Van Rooij, A.; Berden, G.; Huigen, M.C.; Kluijtmans, L.A.; Peters, T.M.; Al-Shekaili, H.H. Untargeted metabolomics and infrared ion spectroscopy identify biomarkers for pyridoxine-dependent epilepsy. J. Clin. Investig. 2021, 131, e148272. [Google Scholar] [CrossRef]
- van Outersterp, R.E.; Engelke, U.F.; Merx, J.; Berden, G.; Paul, M.; Thomulka, T.; Berkessel, A.; Huigen, M.C.; Kluijtmans, L.A.; Mecinović, J. Metabolite identification using infrared ion spectroscopy─ novel biomarkers for pyridoxine-dependent epilepsy. Anal. Chem. 2021, 93, 15340–15348. [Google Scholar] [CrossRef]
- Yazdani, M.; Elgstøen, K.B.P.; Rootwelt, H.; Shahdadfar, A.; Utheim, Ø.A.; Utheim, T.P. Tear Metabolomics in Dry Eye Disease: A Review. Int. J. Mol. Sci. 2019, 20, 3755. [Google Scholar] [CrossRef]
- Ashrafian, H.; Sounderajah, V.; Glen, R.; Ebbels, T.; Blaise, B.J.; Kalra, D.; Kultima, K.; Spjuth, O.; Tenori, L.; Salek, R.; et al. Metabolomics—The stethoscope for the 21st century. Med. Princ. Pract. 2020, 30, 301–310. [Google Scholar] [CrossRef]
- Gowda, G.N.; Zhang, S.; Gu, H.; Asiago, V.; Shanaiah, N.; Raftery, D. Metabolomics-based methods for early disease diagnostics. Expert Rev. Mol. Diagn. 2008, 8, 617–633. [Google Scholar] [CrossRef]
- Evans, E.D.; Duvallet, C.; Chu, N.D.; Oberst, M.K.; Murphy, M.A.; Rockafellow, I.; Sontag, D.; Alm, E.J. Predicting human health from biofluid-based metabolomics using machine learning. Sci. Rep. 2020, 10, 17635. [Google Scholar] [CrossRef]
- Jiménez-Chillarón, J.C.; Díaz, R.; Ramón-Krauel, M. Omics tools for the genome-wide analysis of methylation and histone modifications. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2014; Volume 63, pp. 81–110. [Google Scholar]
- Vitamin D. Fact Sheet for Health Professionals. National Institutes of Health. Office of Dietary Supplements. Available online: https://ods.od.nih.gov/factsheets/VitaminB6-HealthProfessional/ (accessed on 15 November 2022).
- Kuhara, T.; Akiyama, T.; Ohse, M.; Koike, T.; Shibasaki, J.; Imai, K.; Cooper, A.L. Identification of new biomarkers of pyridoxine-dependent epilepsy by GC/MS-based urine metabolomics. Anal. Biochem. 2020, 604, 113739. [Google Scholar] [CrossRef]
- Vinayavekhin, N.; Saghatelian, A. Untargeted metabolomics. Curr. Protoc. Mol. Biol. 2010, 90, 30.1. 1–30.1. 24. [Google Scholar]
- Dunn, W.B.; Broadhurst, D.I.; Atherton, H.J.; Goodacre, R.; Griffin, J.L. Systems level studies of mammalian metabolomes: The roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 2011, 40, 387–426. [Google Scholar] [CrossRef]
- Wishart, D.S. Advances in metabolite identification. Bioanalysis 2011, 3, 1769–1782. [Google Scholar] [CrossRef]
- Dührkop, K.; Nothias, L.-F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C. Systematic classification of unknown metabolites using high-resolution fragmentation mass spectra. Nat. Biotechnol. 2021, 39, 462–471. [Google Scholar] [CrossRef]
- Böcker, S.; Dührkop, K. Fragmentation trees reloaded. J. Cheminformatics 2016, 8, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.E.; Thomas, R.C.; Kruse, L.I.; Silberman, L. Rules for ring closure: Ring formation by conjugate addition of oxygen nucleophiles. J. Org. Chem. 1977, 42, 3846–3852. [Google Scholar] [CrossRef]
- Zschocke, J.; Hoffmann, G.F. Vademecum Metabolicum: Diagnosis and Treatment of Inborn Errors of Metabolism, 5th ed.; Schattauer verlag: Stuttgart, Germany, 2021. [Google Scholar]
- Gilmore, K.; Mohamed, R.K.; Alabugin, I.V. The Baldwin rules: Revised and extended. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2016, 6, 487–514. [Google Scholar] [CrossRef]
- Brundidge, S.P.; Gaeta, F.C.; Hook, D.J.; Sapino, C., Jr.; Elander, R.P.; Morin, R.B. Association of 6-oxo-piperidine-2-carboxylic acid with penicillin V production in Penicillium chrysogenum fermentations. J. Antibiot. 1980, 33, 1348–1351. [Google Scholar] [CrossRef]
- Henriksen, C.M.; Nielsen, J.; Villadsen, J. Cyclization of α-aminoadipic acid into the δ-lactam 6-oxo-piperidine-2-carboxylic acid by Penicillium chrysogenum. J. Antibiot. 1998, 51, 99–106. [Google Scholar] [CrossRef]
- de la Fuente, J.L.; Rumbero, A.; Martin, J.F.; Lirsa, P. Δ-1-Piperideine-6-carboxylate dehydrogenase, a new enzyme that forms α-aminoadipate in Streptomyces clavuligerus and other cephamycin C-producing actinomycetes. Biochem. J. 1997, 327, 59–64. [Google Scholar] [CrossRef]
- Zierer, J.; Jackson, M.A.; Kastenmüller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J. The fecal metabolome as a functional readout of the gut microbiome. Nat. Genet. 2018, 50, 790–795. [Google Scholar] [CrossRef]
- Brocker, C.; Cantore, M.; Failli, P.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) attenuates reactive aldehyde and oxidative stress induced cytotoxicity. Chem.-Biol. Interact. 2011, 191, 269–277. [Google Scholar] [CrossRef]
- Brocker, C.; Lassen, N.; Estey, T.; Pappa, A.; Cantore, M.; Orlova, V.V.; Chavakis, T.; Kavanagh, K.L.; Oppermann, U.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress. J. Biol. Chem. 2010, 285, 18452–18463. [Google Scholar] [CrossRef]
- Requena, J.R.; Chao, C.-C.; Levine, R.L.; Stadtman, E.R. Glutamic and aminoadipic semialdehydes are the main carbonyl products of metal-catalyzed oxidation of proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 69–74. [Google Scholar] [CrossRef]
- Pietzsch, J.; Bergmann, R. Analysis of 6-hydroxy-2-aminocaproic acid (HACA) as a specific marker of protein oxidation: The use of N (O, S)-ethoxycarbonyl trifluoroethyl ester derivatives and gas chromatography/mass spectrometry. Amino Acids 2004, 26, 45–51. [Google Scholar] [CrossRef]
- Danke, S.L. Hexahomoserine as a Competitive Inhibitor for the Incorporation of Lysine into Protein Molecules. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 1963. [Google Scholar]
- Jansen, L.A.; Hevner, R.F.; Roden, W.H.; Hahn, S.H.; Jung, S.; Gospe Jr, S.M. Glial localization of antiquitin: Implications for pyridoxine-dependent epilepsy. Ann. Neurol. 2014, 75, 22–32. [Google Scholar] [CrossRef]
- Al-Shekaili, H.H.; Petkau, T.L.; Pena, I.; Lengyell, T.C.; Verhoeven-Duif, N.M.; Ciapaite, J.; Bosma, M.; van Faassen, M.; Kema, I.P.; Horvath, G. A novel mouse model for pyridoxine-dependent epilepsy due to antiquitin deficiency. Hum. Mol. Genet. 2020, 29, 3266–3284. [Google Scholar] [CrossRef]
- Bok, L.A.; Halbertsma, F.J.; Houterman, S.; Wevers, R.A.; Vreeswijk, C.; Jakobs, C.; Struys, E.; Van der Hoeven, J.H.; Sival, D.A.; Willemsen, M.A. Long-term outcome in pyridoxine-dependent epilepsy. Dev. Med. Child Neurol. 2012, 54, 849–854. [Google Scholar] [CrossRef]
- Coughlin, C.R.; Tseng, L.A.; Bok, L.A.; Hartmann, H.; Footitt, E.; Striano, P.; Tabarki, B.M.; Lunsing, R.J.; Stockler-Ipsiroglu, S.; Gordon, S. Association Between Lysine Reduction Therapies and Cognitive Outcomes in Patients With Pyridoxine-Dependent Epilepsy. Neurology 2022, 99, e2627–e2636. [Google Scholar] [CrossRef]
- Skogvold, H.B.; Sandås, E.M.; Østeby, A.; Løkken, C.; Rootwelt, H.; Rønning, P.O.; Wilson, S.R.; Elgstøen, K.B.P. Bridging the polar and hydrophobic metabolome in single-run untargeted liquid chromatography-mass spectrometry dried blood spot metabolomics for clinical purposes. J. Proteome Res. 2021, 20, 4010–4021. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef]
- Dührkop, K.; Shen, H.; Meusel, M.; Rousu, J.; Böcker, S. Searching molecular structure databases with tandem mass spectra using CSI: FingerID. Proc. Natl. Acad. Sci. USA 2015, 112, 12580–12585. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted metabolomics strategies—Challenges and emerging directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | m/z | RT (min) | Molecular Formula (Compound Discoverer) | Fold Change (Sample/Control) | p-Value |
|---|---|---|---|---|---|
| Biomarker 1 | 148.09668 | 2.442 | C6H13NO3 | 5.852 | 3.9 E-7 |
| Biomarker 2 | 198.07602 | 6.526 | C9H11NO4 | 84.162 | 1.6 E-6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Böhm, H.-O.; Yazdani, M.; Sandås, E.M.; Østeby Vassli, A.; Kristensen, E.; Rootwelt, H.; Skogvold, H.B.; Brodtkorb, E.; Elgstøen, K.B.P. Global Metabolomics Discovers Two Novel Biomarkers in Pyridoxine-Dependent Epilepsy Caused by ALDH7A1 Deficiency. Int. J. Mol. Sci. 2022, 23, 16061. https://doi.org/10.3390/ijms232416061
Böhm H-O, Yazdani M, Sandås EM, Østeby Vassli A, Kristensen E, Rootwelt H, Skogvold HB, Brodtkorb E, Elgstøen KBP. Global Metabolomics Discovers Two Novel Biomarkers in Pyridoxine-Dependent Epilepsy Caused by ALDH7A1 Deficiency. International Journal of Molecular Sciences. 2022; 23(24):16061. https://doi.org/10.3390/ijms232416061
Chicago/Turabian StyleBöhm, Hans-Otto, Mazyar Yazdani, Elise Mørk Sandås, Anja Østeby Vassli, Erle Kristensen, Helge Rootwelt, Hanne Bendiksen Skogvold, Eylert Brodtkorb, and Katja Benedikte Prestø Elgstøen. 2022. "Global Metabolomics Discovers Two Novel Biomarkers in Pyridoxine-Dependent Epilepsy Caused by ALDH7A1 Deficiency" International Journal of Molecular Sciences 23, no. 24: 16061. https://doi.org/10.3390/ijms232416061
APA StyleBöhm, H.-O., Yazdani, M., Sandås, E. M., Østeby Vassli, A., Kristensen, E., Rootwelt, H., Skogvold, H. B., Brodtkorb, E., & Elgstøen, K. B. P. (2022). Global Metabolomics Discovers Two Novel Biomarkers in Pyridoxine-Dependent Epilepsy Caused by ALDH7A1 Deficiency. International Journal of Molecular Sciences, 23(24), 16061. https://doi.org/10.3390/ijms232416061