
A Pro-Inflammatory Signature Constitutively Activated in Monogenic Autoinflammatory Diseases

 ,
,  , and
, and 
Abstract
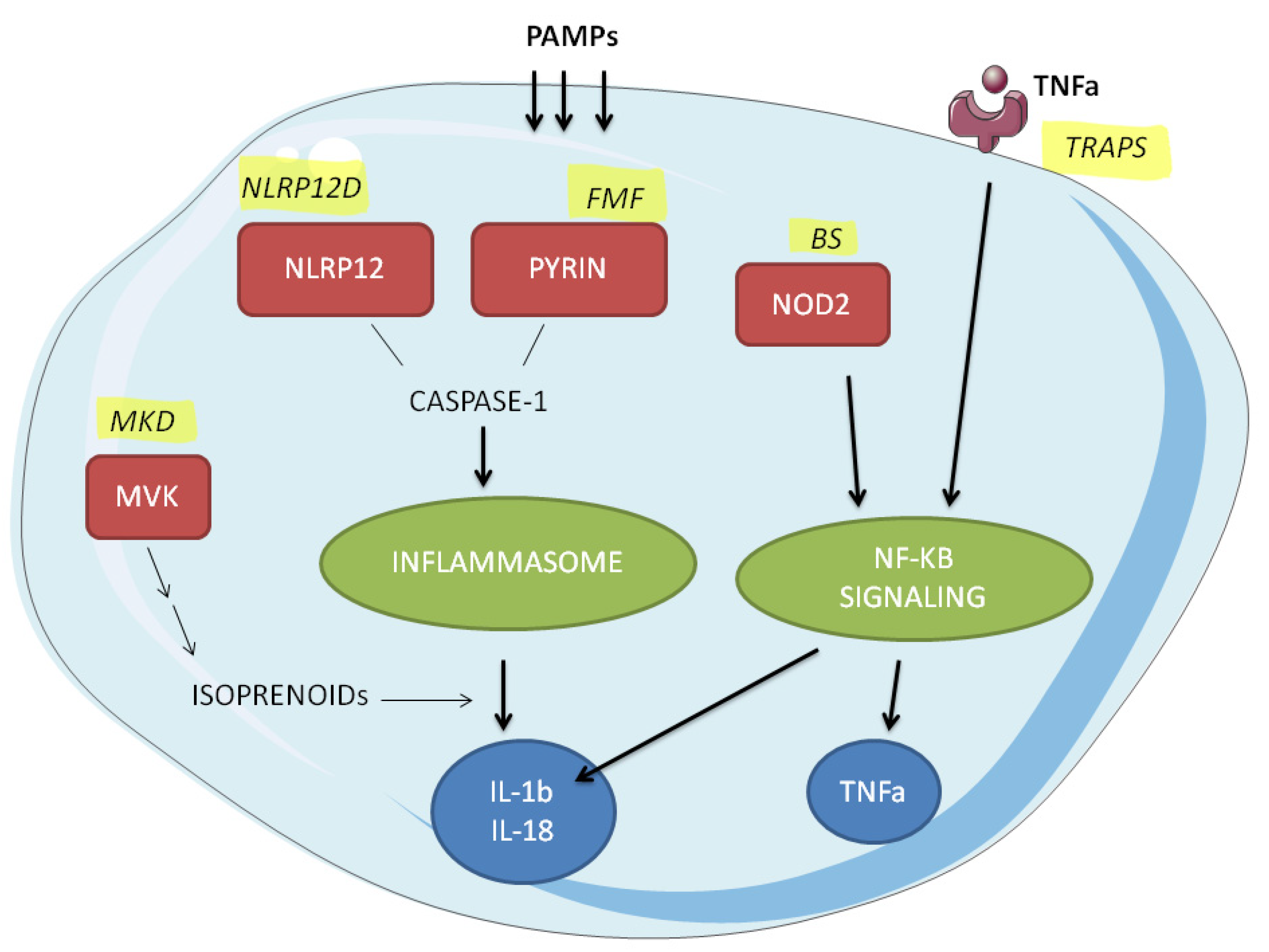
:1. Introduction
2. Results
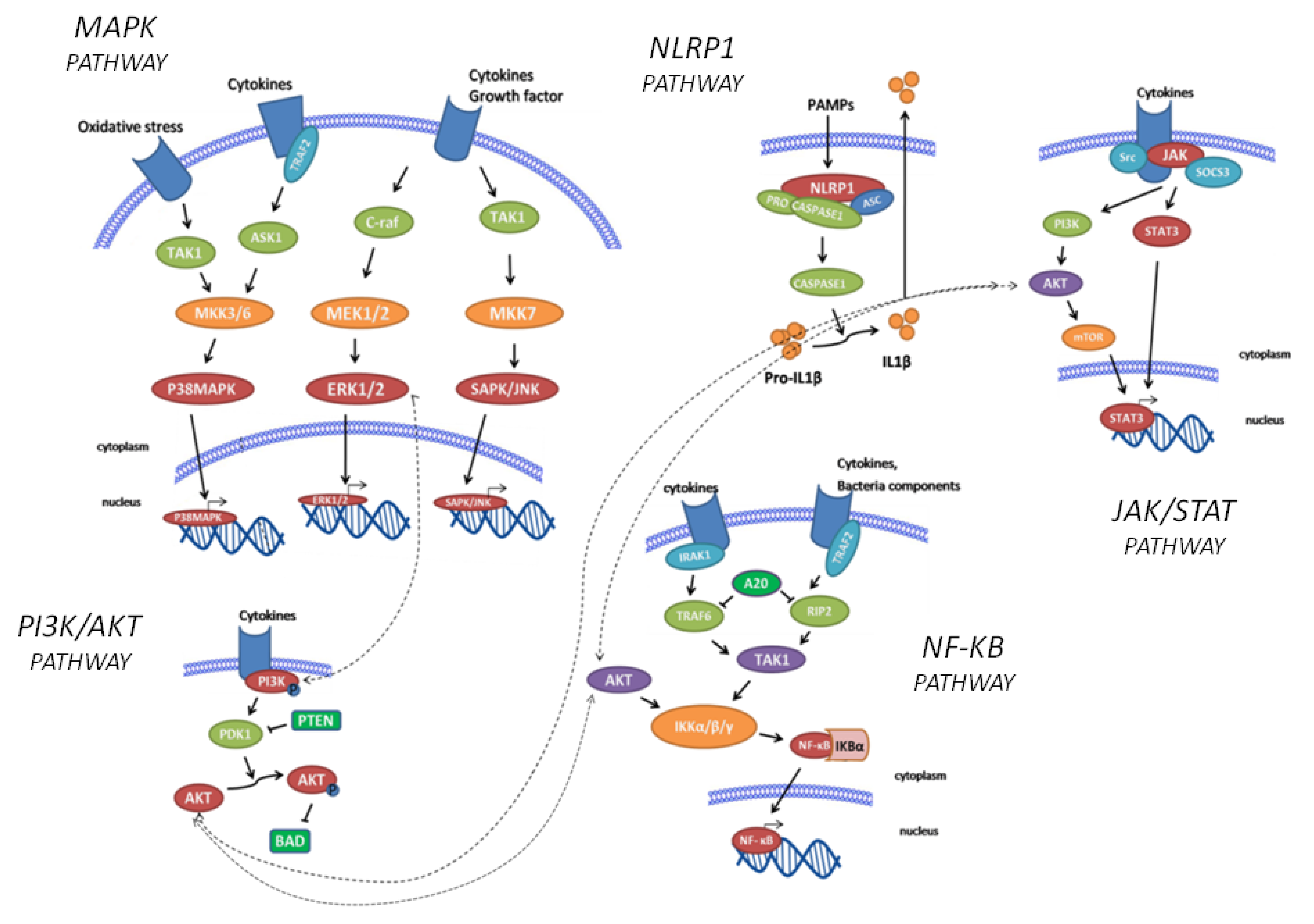
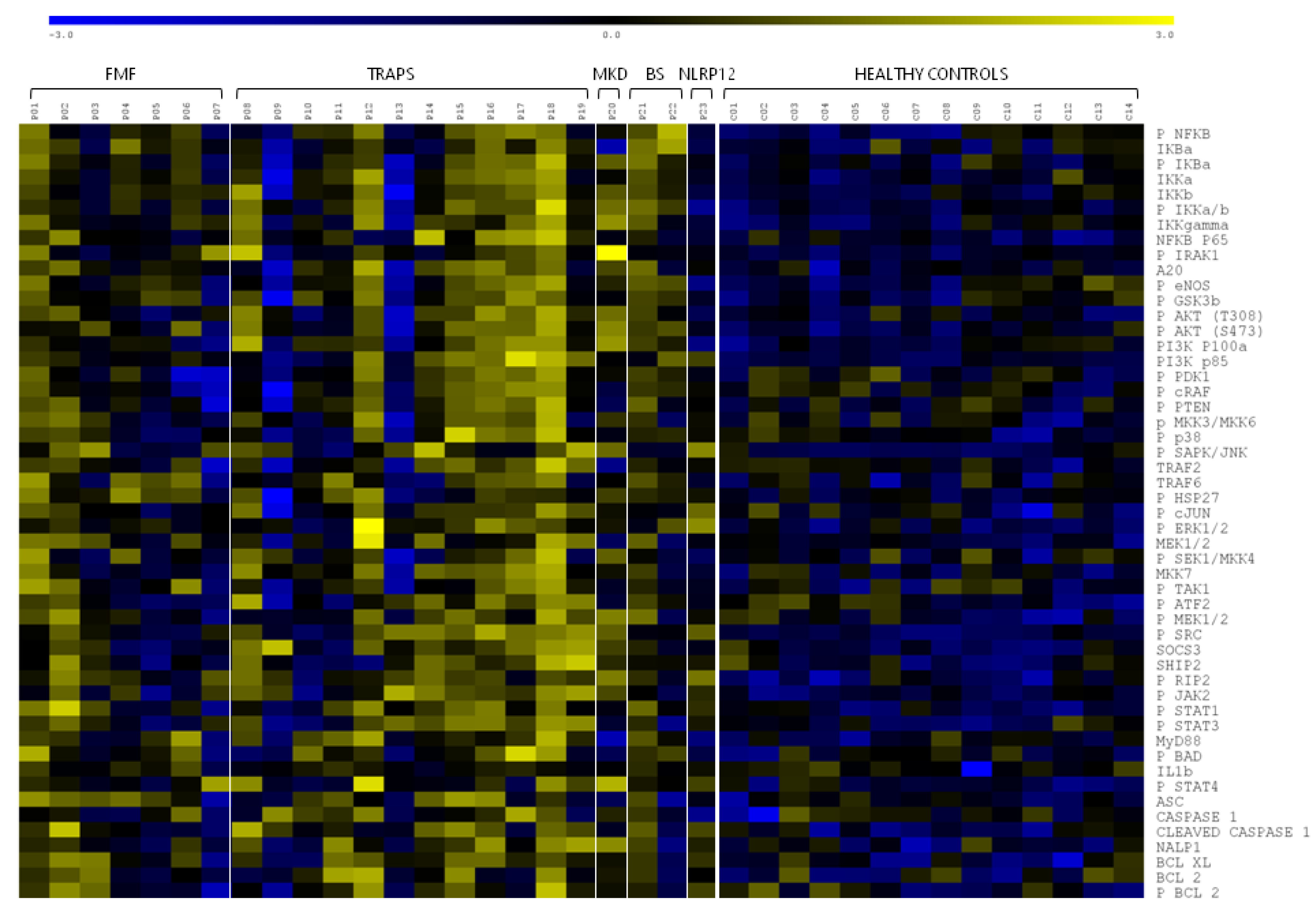
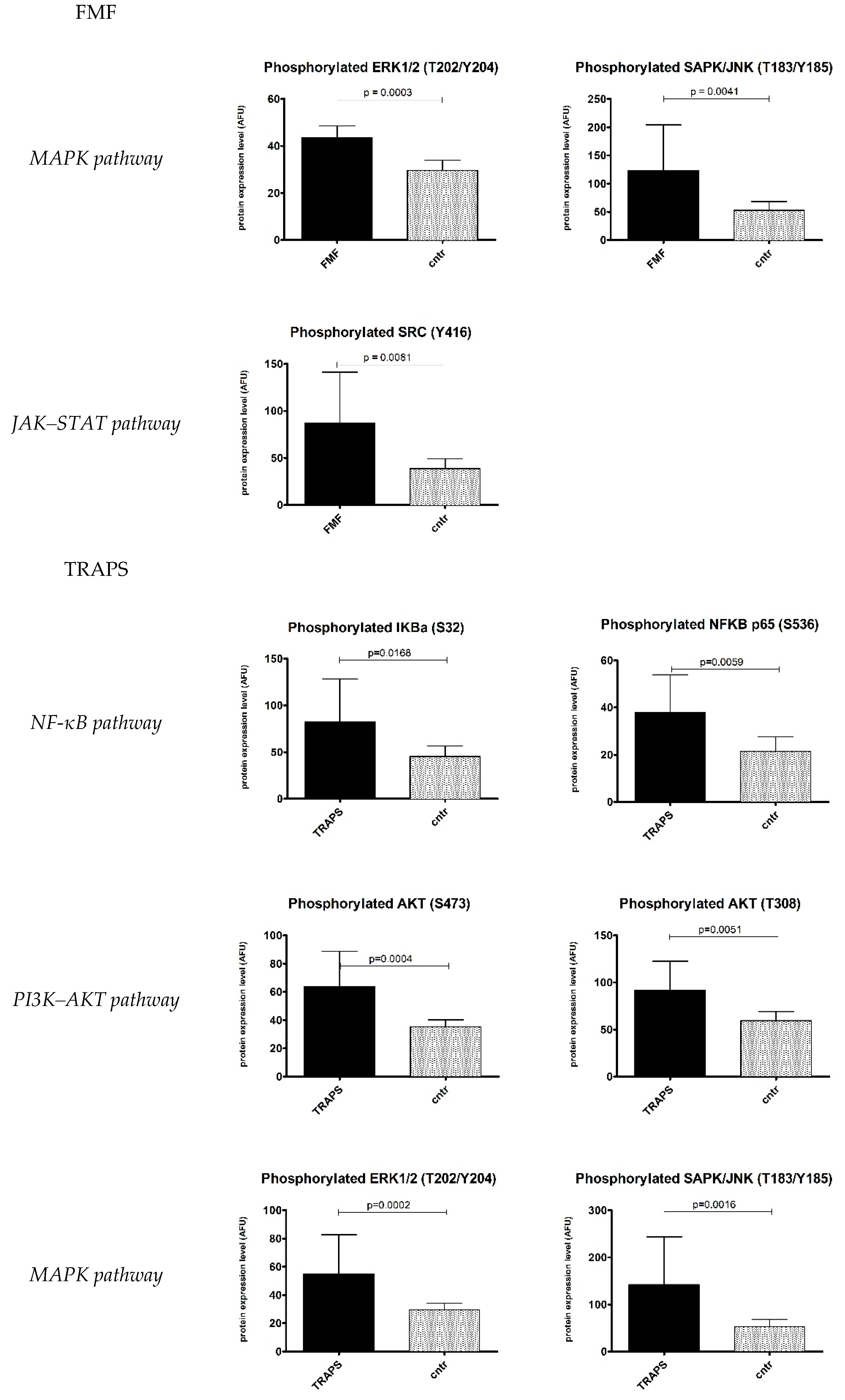
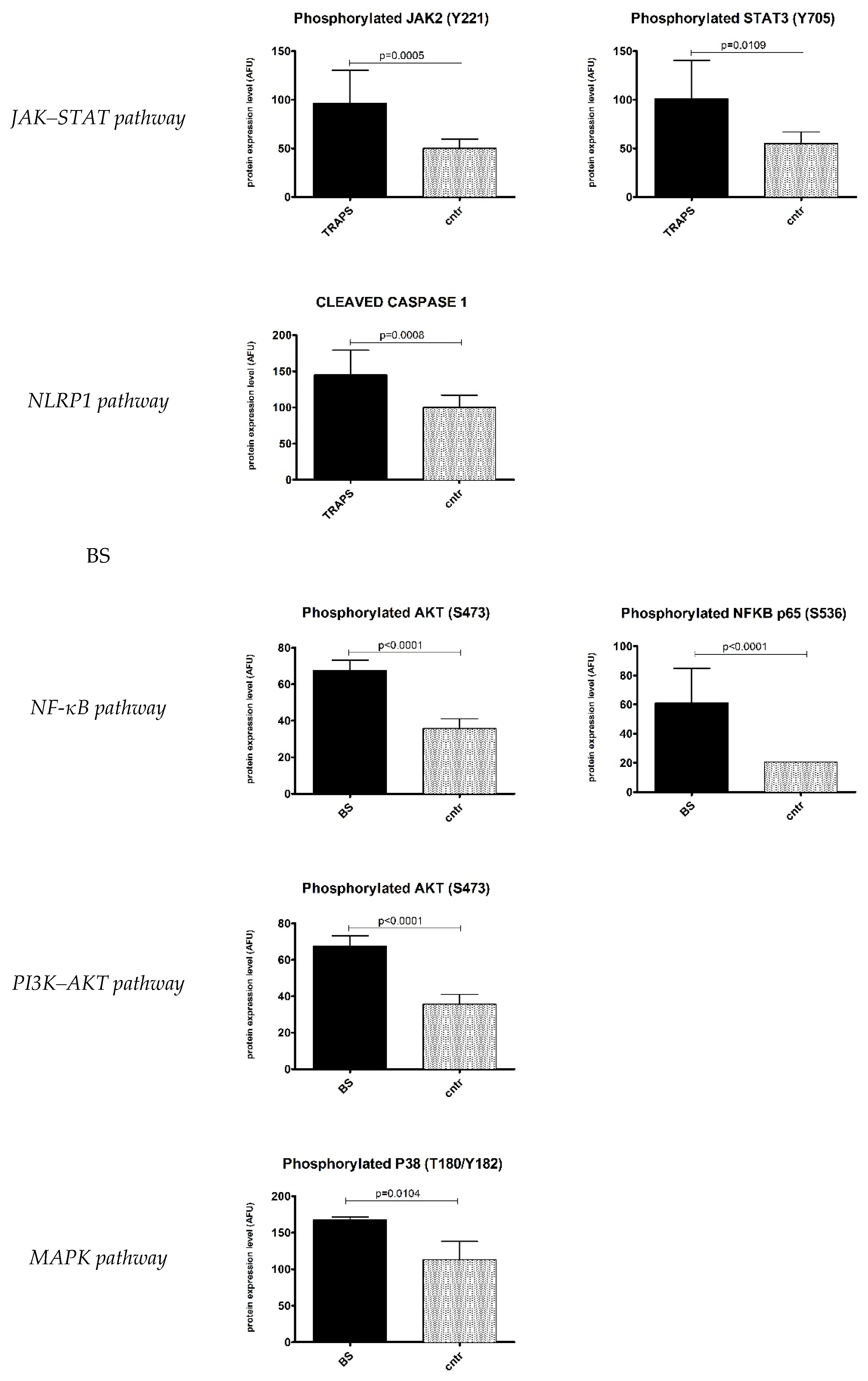
2.1. Inflammatory Signalling Pathways
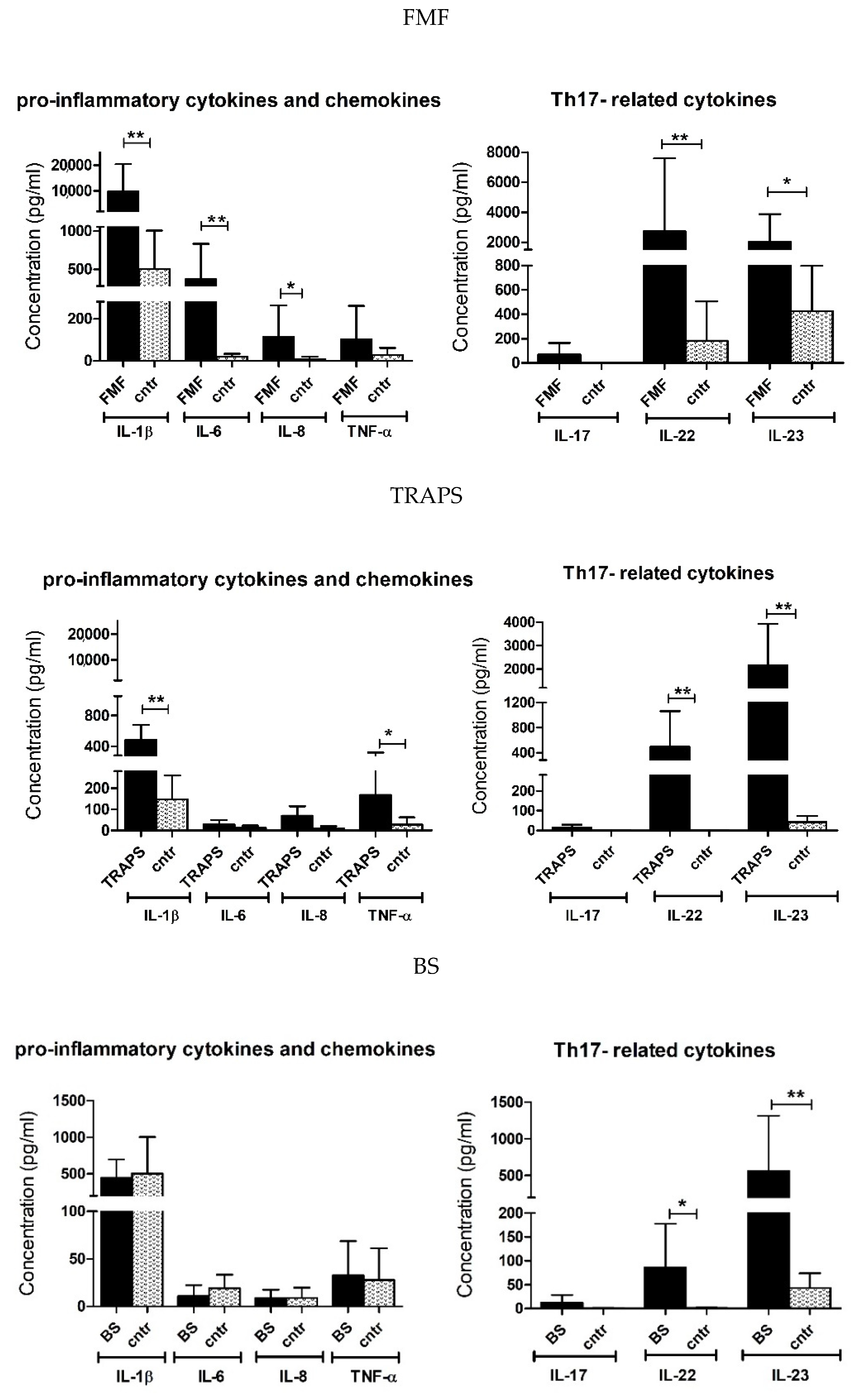
2.2. Upregulated Pro-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Serum and Peripheral Blood Mononuclear Cells Samples
4.3. Reverse Phase Protein Array (RPPA)
4.4. Antibody Microarray
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nigrovic, P.A.; Lee, P.Y.; Hoffman, H.M. Monogenic autoinflammatory disorders: Conceptual overview, phenotype, and clinical approach. J. Allergy Clin. Immunol. 2020, 146, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Beck, D.B.; Aksentijevich, I. Biochemistry of autoinflammatory diseases: Catalyzing monogenic disease. Front. Immunol. 2019, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, M.F.; Aksentijevich, I.; Galon, J.; McDermott, E.M.; Ogunkolade, B.; Centola, M.; Mansfield, E.; Gadina, M.; Karenko, L.; Pettersson, T.; et al. Germline Mutations in the Extracellular Domains of the 55 kDa TNF Receptor, TNFR1, Define a Family of Dominantly Inherited Autoinflammatory Syndromes. Cell 1999, 97, 133–144. [Google Scholar] [CrossRef]
- French FMF Consortium. A candidate gene for familial Mediterranean fever. Nat. Genet. 1997, 17, 25–31. [Google Scholar] [CrossRef]
- Aksentijevich, I.; Schnappauf, O. Molecular mechanisms of phenotypic variability in monogenic autoinflammatory diseases. Nat. Rev. Rheumatol. 2021, 17, 405–425. [Google Scholar] [CrossRef]
- Lachmann, H.J. Periodic fever syndromes. Best Pract. Res. Clin. Rheumatol. 2017, 31, 596–609. [Google Scholar] [CrossRef]
- Manthiram, K.; Zhou, Q.; Aksentijevich, I.; Kastner, D.L. The monogenic autoinflammatory diseases define new pathways in human innate immunity and inflammation. Nat. Immunol. 2017, 18, 832–842. [Google Scholar] [CrossRef]
- Galozzi, P.; Punzi, L.; Sfriso, P. Clinical Overlapping in Autoinflammatory Diseases: The Role of Gene Duplication. Front. Immunol. 2017, 8, 392. [Google Scholar] [CrossRef] [Green Version]
- Di Donato, G.; d’Angelo, D.M.; Breda, L.; Chiarelli, F. Monogenic Autoinflammatory Diseases: State of the Art and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 6360. [Google Scholar] [CrossRef]
- Tighe, P.; Negm, O.; Todd, I.; Fairclough, L. Utility, reliability and reproducibility of immunoassay multiplex kits. Methods (San Diego Calif.) 2013, 61, 23–29. [Google Scholar] [CrossRef]
- Awad, F.; Assrawi, E.; Louvrier, C.; Jumeau, C.; Georgin-Lavialle, S.; Grateau, G.; Amselem, S.; Giurgea, I.; Karabina, S.-A. Inflammasome biology, molecular pathology and therapeutic implications. Pharmacol. Ther. 2018, 187, 133–149. [Google Scholar] [CrossRef]
- Masumoto, J.; Zhou, W.; Morikawa, S.; Hosokawa, S.; Taguchi, H.; Yamamoto, T.; Kurata, M.; Kaneko, N. Molecular biology of autoinflammatory diseases. Inflamm. Regen. 2021, 41, 33. [Google Scholar] [CrossRef]
- Tartey, S.; Kanneganti, T. Inflammasomes in the pathophysiology of autoinflammatory syndromes. J. Leukoc. Biol. 2020, 107, 379–391. [Google Scholar] [CrossRef]
- Espina, V.; Liotta, L.A.; Petricoin, E.F. Reverse-phase protein microarrays for theranostics and patient tailored therapy. Methods Mol. Biol. 2009, 520, 89–105. [Google Scholar] [PubMed]
- Chae, J.J.; Wood, G.; Richard, K.; Jaffe, H.; Colburn, N.T.; Masters, S.L.; Gumucio, D.L.; Shoham, N.G.; Kastner, D.L. The familial Mediterranean fever protein, pyrin, is cleaved by caspase-1 and activates NF-kappaB through its N-terminal fragment. Blood 2008, 112, 1794–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokuta, M.A.; Cooper, K.M.; Aksentijevich, I.; Kastner, D.L.; Huttenlocher, A. Neutrophil chemotaxis in a patient with neonatal-onset multisystem inflammatory disease and Muckle-Wells syndrome. Ann. Allergy Asthma Immunol. 2005, 95, 394–399. [Google Scholar] [CrossRef]
- Karadeniz, H.; Güler, A.A.; Atas, N.; Satış, H.; Salman, R.B.; Babaoglu, H.; Tufan, A. Tofacitinib for the treatment for colchicine-resistant familial Mediterranean fever: Case-based review. Rheumatol. Int. 2020, 40, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Heilig, R.; Broz, P. Function and mechanism of the pyrin inflammasome. Eur. J. Immunol. 2018, 48, 230–238. [Google Scholar] [CrossRef]
- Chae, J.J.; Cho, Y.H.; Lee, G.S.; Cheng, J.; Liu, P.P.; Feigenbaum, L.; Katz, S.I.; Kastner, D.L. Gain-of-function Pyrin mutations induce NLRP3 protein-independent interleukin-1β activation and severe autoinflammation in mice. Immunity 2011, 34, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.; Fielding, C.A.; Williams, B.D.; Brennan, P. Mutation of the extracellular domain of tumour necrosis factor receptor 1 causes reduced NF-kappaB activation due to decreased surface expression. FEBS Lett. 2005, 579, 5193–5198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousaf, N.; Gould, D.J.; Aganna, E.; Hammond, L.; Mirakian, R.M.; Turner, M.D.; Hitman, G.A.; McDermott, M.F.; Chernajovsky, Y. Tumor necrosis factor receptor I from patients with tumor necrosis factor receptor-associated periodic syndrome interacts with wild-type tumor necrosis factor receptor I and induces ligand-independent NF-kappaB activation. Arthritis Rheumatol. 2005, 52, 2906–2916. [Google Scholar] [CrossRef] [PubMed]
- Greco, E.; Aita, A.; Galozzi, P.; Gava, A.; Sfriso, P.; Negm, O.H.; Tighe, P.; Caso, F.; Navaglia, F.; Dazzo, E.; et al. The novel S59P mutation in the TNFRSF1A gene identified in an adult onset TNF receptor associated periodic syndrome (TRAPS) constitutively activates NF-κB pathway. Arthritis Res. Ther. 2015, 17, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negm, O.H.; Mannsperger, H.A.; McDermott, E.M.; Drewe, E.; Powell, R.J.; Todd, I.; Fairclough, L.C.; Tighe, P.J. A pro-inflammatory signalome is constitutively activated by C33Y mutant TNF receptor 1 in TNF receptor-associated periodic syndrome (TRAPS). Eur. J. Immunol. 2014, 44, 2096–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, D.; Ueno, L.; Vogt, P.K. Akt-mediated regulation of NFkappaB and the essentialness of NFkappaB for the oncogenicity of PI3K and Akt. Int. J. Cancer 2009, 125, 2863–2870. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.; Park, H.; Maddipati, R.; Lobito, A.A.; Bulua, A.C.; Jackson, A.J.; Chae, J.J.; Ettinger, R.; de Koning, H.D.; Cruz, A.C.; et al. Concerted action of wild-type and mutant TNF receptors enhances inflammation in TNF receptor 1-associated periodic fever syndrome. Proc. Natl. Acad. Sci. USA 2010, 107, 9801–9806. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, S.; Muñoz-Fernández, M.Á. TNF-A may mediate inflammasome activation in the absence of bacterial infection in more than one way. PLoS ONE 2013, 8, E71477. [Google Scholar]
- Rebelo, S.L.; Bainbridge, S.E.; Amel-Kashipaz, M.R.; Radford, P.M.; Powell, R.J.; Todd, I.; Tighe, P.J. Modeling of tumor necrosis factor receptor superfamily 1A mutants associated with tumor necrosis factor receptor-associated periodic syndrome indicates misfolding consistent with abnormal function. Arthritis Rheumatol. 2006, 54, 2674–2687. [Google Scholar] [CrossRef]
- Infevers. Available online: https://infevers.umai-montpellier.fr/web/ (accessed on 10 December 2021).
- Havla, J.; Lohse, P.; Gerdes, L.A.; Hohlfeld, R.; Kümpfel, T. Symptoms related to tumor necrosis factor receptor 1-associated periodic syndrome, multiple sclerosis, and severe rheumatoid arthritis in patients carrying the TNF receptor superfamily 1A D12E/p.Asp41Glu mutation. J. Rheumatol. 2013, 40, 261–264. [Google Scholar] [CrossRef]
- Herb, M.; Gluschko, A.; Schramm, M. Reactive Oxygen Species: Not Omnipresent but Important in Many Locations. Front. Cell Dev. Biol. 2021, 9, 716406. [Google Scholar] [CrossRef]
- Bulua, A.C.; Simon, A.; Maddipati, R.; Pelletier, M.; Park, H.; Kim, K.Y.; Sack, M.N.; Kastner, D.L.; Siegel, R.M. Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and are elevated in TNFR1-associated periodic syndrome (TRAPS). J. Exp. Med. 2011, 208, 519–533. [Google Scholar] [CrossRef]
- Zhao, L.; Lee, J.Y.; Hwang, D.H. The phosphatidylinositol 3-kinase/Akt pathway negatively regulates Nod2-mediated NF-kappaB pathway. Biochem. Pharmacol. 2008, 75, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Mandey, S.H.L.; Kuijk, L.M.; Frenkel, J.; Waterham, H.R. A role for geranylgeranylation in interleukin-1β secretion. Arthritis Rheumatol. 2006, 54, 3690–3695. [Google Scholar] [CrossRef] [PubMed]
- Dang, E.V.; Cyster, J.G. Loss of sterol metabolic homeostasis triggers inflammasomes—How and why. Curr. Opin. Immunol. 2019, 56, 1–9. [Google Scholar] [CrossRef]
- Carapito, R.; Carapito, C.; Morlon, A.; Paul, N.; Jacome, A.S.V.; Alsaleh, G.; Rolli, V.; Tahar, O.; Aouadi, I.; Rompais, M.; et al. Multi-OMICS analyses unveil STAT1 as a potential modifier gene in mevalonate kinase deficiency. Ann. Rheum. Dis. 2018, 77, 1675–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jéru, I.; Duquesnoy, P.; Fernandes-Alnemri, T.; Cochet, E.; Yu, J.W.; Lackmy-Port-Lis, M.; Grimprel, E.; Landman-Parker, J.; Hentgen, V.; Marlin, S.; et al. Mutations in NALP12 cause hereditary periodic fever syndromes. Proc. Natl. Acad. Sci. USA 2008, 105, 1614–1619. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Ghilardi, N.; Xie, M.H.; de Sauvage, F.J.; Gurney, A.L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 2003, 278, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Qu, N.; Xu, M.; Mizoguchi, I.; Furusawa, J.; Kaneko, K.; Watanabe, K.; Mizuguchi, J.; Itoh, M.; Kawakami, Y.; Yoshimoto, T. Pivotal roles of T-helper 17-related cytokines, IL-17, IL-22, and IL-23, in inflammatory diseases. Clin. Dev. Immunol. 2013, 2013, 968549. [Google Scholar] [CrossRef] [Green Version]
- Tahmasebinia, F.; Pourgholaminejad, A. The role of Th17 cells in auto-inflammatory neurological disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 408–416. [Google Scholar] [CrossRef]
- Meng, G.; Zhang, F.; Fuss, I.; Kitani, A.; Strober, W. A mutation in the Nlrp3 gene causing inflammasome hyperactivation potentiates Th17 cell-dominant immune responses. Immunity 2009, 30, 860–874. [Google Scholar] [CrossRef] [Green Version]
- Ovadia, A.; Livneh, A.; Feld, O.; Ben-Zvi, I.; Kukuy, E.; Kivity, S.; Lidar, M.; Barda-Saad, M.; Shinar, Y. T helper 17 polarization in familial Mediterranean fever. Genes Immune. 2013, 14, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, M.; Yousefi, M.; Abbaspour-Aghdam, S.; Dolati, S.; Aghebati-Maleki, L.; Eghbal-Fard, S.; Khabbazi, A.; Rostamzadeh, D.; Alipour, S.; Shabani, M. Disturbed Th17/Treg balance, cytokines, and miRNAs in peripheral blood of patients with Behcet’s disease. J. Cell. Physiol. 2019, 234, 3985–3994. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.C.; Skokos, D. Th17 response and inflammatory autoimmune diseases. Int. J. Inflamm. 2012, 2012, 819467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannsperger, H.A.; Gade, S.; Henjes, F.; Beissbarth, T.; Korf, U. RPPanalyzer: Analysis of reverse-phase protein array data. Bioinformatics 2010, 26, 2202–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvarajah, S.; Negm, O.H.; Hamed, M.R.; Tubby, C.; Todd, I.; Tighe, P.J.; Harrison, T.; Fairclough, L.C. Development and validation of protein microarray technology for simultaneous inflammatory mediator detection in human sera. Mediat. Inflamm. 2014, 2014, 820304. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age | Sex | Disease | Associated Gene | Mutations or Polymorphisms | Clinical | Treatments |
|---|---|---|---|---|---|---|---|
| P1 | 30 y | F | FMF | MEFV | E148Q he | Absence of febrile episodes and arthralgia; CRP and SAA slightly above the normal limits | None; (Colchicine 1mg/day discontinued for remission) |
| P2 | 48 y | F | FMF | MEFV | E148Q he + M680I he | Absence of febrile episodes, arthralgia, and abdominal pain; CRP and SAA normal | Colchicine 1 mg/day |
| P3 | 38 y | M | FMF | MEFV | R202 Q he | Occasional febrile episodes not associated with other symptoms; CRP and SAA slightly above the normal limits | Colchicine 1 mg/day |
| P4 | 19 y | F | FMF | MEFV | none | Occasional febrile episodes not associated with other symptoms | Colchicine 1 mg/day |
| P5 | 34 y | F | FMF | MEFV | E148Q he + P369S he | Accentuation of joint symptoms in the ankles and stabbing pain in the thoracic area; CRP and SAA normal; a slight reduction in neutrophils | Colchicine 1 mg/day |
| P6 | 57 y | M | FMF | MEFV | R202Q he | Abdominal pain; occasional febrile episodes | Colchicine 1mg/day |
| P7 | 36 y | M | FMF | MEFV | M641I he | Absence of febrile episodes and arthralgia | none |
| P8 | 70 y | M | TRAPS | TNFRSF1A | S59P he | Episodes of arthralgia | Steroids 4mg |
| P9 | 46 y | F | TRAPS | TNFRSF1A | R92Q he | Pain in the cervical spine; occasional febrile episodes; arthralgia | Infliximab 5 mg/kg every 6 weeks |
| P10 | 22 y | M | TRAPS | TNFRSF1A | c.626-32G > T | Abdominal pain; occasional febrile episodes, arthralgia; CRP slightly above the normal limit | Steroids as needed |
| P11 | 27 y | F | TRAPS | TNFRSF1A | c.625+10A > G | Hand dermatitis; no episodes of hyperpyrexia; no skin rash; no oral or genital ulcers; not arthralgia. | Colchicine 1mg/day; NSAIDs |
| P12 | 36 y | F | TRAPS | TNFRSF1A | V95M he | Occasional febrile episodes not associated with other symptoms | Canakinumab 150mg/8 weeks |
| P13 | 34 y | M | TRAPS | TNFRSF1A | delY103-R104 | Controlled articular and cutaneous manifestations | Etanercept 50mg/week |
| P14 | 32 y | M | TRAPS | TNFRSF1A | T50M he | Controlled disease manifestations | none |
| P15 | 54 y | F | TRAPS | TNFRSF1A | R104Q he | Occasional febrile episodes | Anakinra 100mg/day |
| P16 | 41 y | M | TRAPS | TNFRSF1A | T50M he | Controlled articular and cutaneous manifestations | Anakinra 100mg/day |
| P17 | 34 y | M | TRAPS | TNFRSF1A | R92Q he | Absence of febrile episodes and arthralgia | none |
| P18 | 18 y | F | TRAPS | TNFRSF1A | D12E he | Controlled disease manifestations | Anakinra 100mg/day |
| P19 | 56 y | F | TRAPS | TNFRSF1A | R92Q he | Controlled disease manifestations | Anakinra 100mg/day |
| P20 | 14 y | M | MKD | MVK | V377I ho | Controlled articular and cutaneous manifestations | none |
| P21 | 59 y | F | BS | NOD2 | E383K he | Controlled articular and cutaneous manifestations; severe uveitis | Steroids 4mg; Humira 40mg/14days |
| P22 | 34 y | F | BS | NOD2 | E383K he | Controlled articular and cutaneous manifestations; progressive uveitis | none; (Steroids 4mg discontinued for remission) |
| P23 | 38 y | M | NLRP12D | NLRP12 | G448A he | Absence of febrile episodes and arthralgia | none |
| NF-κb Pathway | PI3/AKT Pathway | MAPK Pathways | |||
|---|---|---|---|---|---|
| Antibody | Dilution | Antibody | Dilution | Antibody | Dilution |
| NF-κB | 1:250 | p eNOS (Ser1177) | 1:500 | p MKK3/MKK6 (Ser189/Ser207) | 1:100 |
| p NF-κB p65 (Ser536) | 1:100 | p BAD (Ser136) | 1:500 | p P38 MAPK (Thr180/Tyr182) | 1:1000 |
| IΚBα | 1:100 | p AKT (Thr308) | 1:50 | p SAPK/JNK (Thr183/Tyr185) | 1:1000 |
| p IΚBα (Ser32) | 1:500 | p AKT (Ser473) | 1:25 | TRAF2 | 1:500 |
| IKKα | 1:250 | PI3K p100α | 1:250 | TRAF6 | 1:500 |
| IKKβ | 1:250 | PI3K p85 | 1:250 | p HSP27 (Ser82) | 1:50 |
| p IKKα/β (Ser176/Ser177) | 1:250 | p PDK1 (Ser259) | 1:1000 | p c-JUN (Ser63) | 1:200 |
| IKKγ | 1:250 | p GSK 3β (Ser9) | 1:500 | p ERK1/2 (Thr202/Tyr204) | 1:2000 |
| p IRAK1 (Thr209) | 1:3000 | p c-RAF (Ser259) | 1:1000 | MEK1/2 | 1:250 |
| A20 | 1:150 | p PTEN (Ser380) | 1:1000 | p MEK1/2 (Ser217/Ser221) | 1:250 |
| p SEK1/MKK4 (Ser257/Thr261) | 1:250 | ||||
| MKK7 | 1:100 | ||||
| p TAK1 (Ser412) | 1:100 | ||||
| p ATF2 (Thr71) | 1:250 | ||||
| SRC–JAK–STAT3 pathway | INFLAMMASOME pathway | others | |||
| Antibody | Dilution | Antibody | Dilution | Antibody | Dilution |
| p STAT3 (Tyr705) | 1:100 | IL-1β (*) | 1:100 | MyD88 | 1:50 |
| p c-SRC (Tyr416) | 1:1,0000 | ASC (TMS1) | 1:150 | Actin (Mouse) | 1:1000 |
| SOCS3 | 1:500 | CASPASE1 | 1:250 | Actin (Rabbit) | 1:1000 |
| SHIP2 | 1:2000 | CLEAVED CASPASE1 | 1:50 | ||
| p RIP2 (Ser176) | 1:500 | NALP1 | 1:150 | ||
| p JAK2 (Tyr1007) | 1:1000 | p BAD (Ser136) | 1:500 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galozzi, P.; Negm, O.; Bindoli, S.; Tighe, P.; Sfriso, P.; Punzi, L. A Pro-Inflammatory Signature Constitutively Activated in Monogenic Autoinflammatory Diseases. Int. J. Mol. Sci. 2022, 23, 1828. https://doi.org/10.3390/ijms23031828
Galozzi P, Negm O, Bindoli S, Tighe P, Sfriso P, Punzi L. A Pro-Inflammatory Signature Constitutively Activated in Monogenic Autoinflammatory Diseases. International Journal of Molecular Sciences. 2022; 23(3):1828. https://doi.org/10.3390/ijms23031828
Chicago/Turabian StyleGalozzi, Paola, Ola Negm, Sara Bindoli, Patrick Tighe, Paolo Sfriso, and Leonardo Punzi. 2022. "A Pro-Inflammatory Signature Constitutively Activated in Monogenic Autoinflammatory Diseases" International Journal of Molecular Sciences 23, no. 3: 1828. https://doi.org/10.3390/ijms23031828
APA StyleGalozzi, P., Negm, O., Bindoli, S., Tighe, P., Sfriso, P., & Punzi, L. (2022). A Pro-Inflammatory Signature Constitutively Activated in Monogenic Autoinflammatory Diseases. International Journal of Molecular Sciences, 23(3), 1828. https://doi.org/10.3390/ijms23031828