
Acrylamide Neurotoxicity as a Possible Factor Responsible for Inflammation in the Cholinergic Nervous System



Abstract
:1. Introduction
2. Aim
3. Acrylamide
4. The Cholinergic System
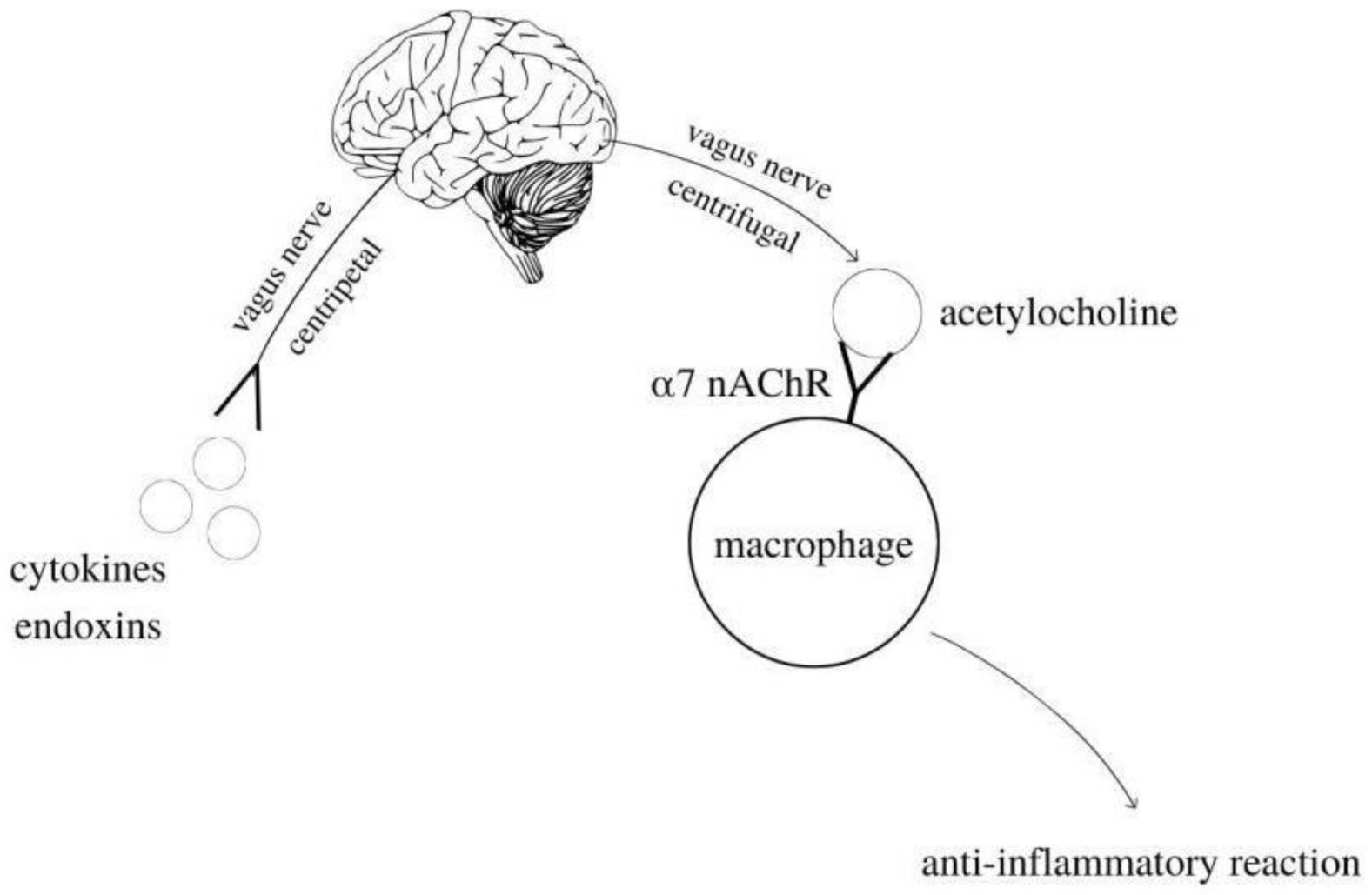
5. Cholinergic Anti-Inflammatory Pathway
6. Description of Acetylcholinesterase and the Inflammatory Response
7. Effect of Acrylamide on Acetylcholinesterase
8. Effect of ACR on the Inflammatory Response through Its Action on Acetylcholinesterase
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Koszucka, A.; Nowak, A.; Nowak, I.; Motyl, I. Acrylamide in human diet, its metabolism, toxicity, inactivation and the associated European Union legal regulations in food industry. Crit. Rev. Food Sci. Nutr. 2019, 60, 1677–1692. [Google Scholar] [CrossRef]
- Girma, K.B.; Lorenz, V.; Blaurock, S.; Edelmann, F. Coord. Chem. Rev. 2005, 249, 1283–1293. [Google Scholar]
- Oracz, J.; Nebesny, E.; Żyżelewicz, D. New trends in quantification of acrylamide in food products. Talanta 2011, 86, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Besaratinia, A.; Pfeifer, G.P. DNA adduction and mutagenic properties of acrylamide. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 580, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Galdo, V.C.; Massart, C.; Jin, L.; Vanvooren, V.; Caillet-Fauquet, P.; Andry, G.; Lothaire, P.; Dequanter, D.; Friedman, M.; Van Sande, J. Acrylamide, an in vivo thyroid carcinogenic agent, induces DNA damage in rat thyroid cell lines and primary cultures. Mol. Cell. Endocrinol. 2006, 257–258, 6–14. [Google Scholar] [CrossRef]
- Szczerbina, T.; Banach, Z.; Tylko, G.; Pyza, E. Toxic effects of acrylamide on survival, development and haemocytes of Musca domestica. Food Chem. Toxicol. 2008, 46, 2316–2319. [Google Scholar] [CrossRef]
- LoPachin, R.M. The Changing View of Acrylamide Neurotoxicity. Neurotoxicology 2004, 25, 617–630. [Google Scholar] [CrossRef]
- Yousef, M.; El-Demerdash, F. Acrylamide-induced oxidative stress and biochemical perturbations in rats. Toxicology 2006, 219, 133–141. [Google Scholar] [CrossRef]
- Marrs, T.C.; Maynard, R.L. Neurotranmission systems as targets for toxicants: A review. Cell Biol. Toxicol. 2013, 29, 381–396. [Google Scholar] [CrossRef]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondžić, A.M.; Vasic, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [Green Version]
- Kosinski, R.A.; Zaremba, M. Dynamics of the model of the Caenorhabditis Elegans neural network. Acta Phys. Pol. B 2007, 38, 2201–2210. [Google Scholar]
- Parsons, S.M.; Prior, C.; Marshall, I.G. Acetylcholine Transport, Storage, And Release. Int. Rev. Neurobiol. 1993, 35, 279–390. [Google Scholar] [PubMed]
- Bellier, J.-P.; Kimura, H. Peripheral type of choline acetyltransferase: Biological and evolutionary implications for novel mechanisms in cholinergic system. J. Chem. Neuroanat. 2011, 42, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Halder, N.; Lal, G. Cholinergic System and Its Therapeutic Importance in Inflammation and Autoimmunity. Front. Immunol. 2021, 12, 660342. [Google Scholar] [CrossRef] [PubMed]
- Wierzbiński, P.; Florkowski, A. Cholinergis system and schiophrenia a potential therapeutic role of cholinergic agonists. Psychiatr. Psychoter. 2010, 6, 53–62. [Google Scholar]
- Okuma, Y.T.; Nomura, Y. Roles of muscarinic acetylcholine receptors in interleukin-2 synthesis in lymphocytes. Jpn. J. Clin. Pharmacol. Ther. 2001, 85, 16–19. [Google Scholar]
- Liu, E.; Xia, Y.; Kong, X.; Guo, M.; Yu, A.; Zheng, B.; Mak, S.; Xu, M.L.; Tsim, K. Interacting with α 7 nAChR is a new mechanism for AChE to enhance the inflammatory response in macrophages. Acta Pharm. Sin. B 2020, 10, 1926–1942. [Google Scholar] [CrossRef]
- Dani, J.A. Overview of nicotinic receptors and their roles in the central nervous system. Biol. Psychiatry 2001, 49, 166–174. [Google Scholar] [CrossRef]
- Hoover, D.B. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol. Ther. 2017, 179, 1–16. [Google Scholar] [CrossRef]
- Kopanska, M.; Czech, J.; Zagata, P.; Dobrek, L.; Thor, P.; Formicki, G. Effect of the different doses of acrylamide on acetylocholinoesterase activity, thiol groups, malondialdehyde concentrations in hypothalamus and selected muscles of mice. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2017, 68, 565–571. [Google Scholar]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- Wang, H.; Liao, H.; Ochani, M.; Justiniani, M.; Lin, X.; Yang, L.; Al-Abed, Y.; Wang, H.; Metz, C.; Miller, E.J.; et al. Cholinergic agonists inhibit HMGB1 release and improve survival in experimental sepsis. Nat. Med. 2004, 10, 1216–1221. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Ballina, M.; Tracey, K.J. Cholinergic control of inflammation. J. Intern. Med. 2009, 265, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Oke, S.L.; Tracey, K.J. From CNI-1493 to the immunological homunculus: Physiology of the inflammatory reflex. J. Leukoc. Biol. 2007, 83, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. Physiology and immunology of the cholinergic antiinflammatory pathway. J. Clin. Investig. 2007, 117, 289–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, V.A.; Tracey, K.J. Controlling inflammation: The cholinergic anti-inflammatory pathway. Biochem. Soc. Trans. 2006, 34, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Wang, H.; Czura, C.J.; Friedman, S.G.; Tracey, K.J. The Cholinergic Anti-inflammatory Pathway: A Missing Link in Neuroimmunomodulation. Mol. Med. 2003, 9, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Li, X.; Hao, J. The cholinergic anti-inflammatory pathway: An innovative treatment strategy for neurological diseases. Neurosci. Biobehav. Rev. 2017, 77, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Bernik, T.R.; Friedman, S.G.; Ochani, M.; DiRaimo, R.; Ulloa, L.; Yang, H.; Sudan, S.; Czura, C.J.; Ivanova, S.M.; Tracey, K.J. Pharmacological Stimulation of the Cholinergic Antiinflammatory Pathway. J. Exp. Med. 2002, 195, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Mioni, C.; Bazzani, C.; Giuliani, D.; Altavilla, D.; Leone, S.; Ferrari, A.; Minutoli, L.; Bitto, A.; Marini, H.; Zaffe, D.; et al. Activation of an efferent cholinergic pathway produces strong protection against myocardial ischemia/reperfusion injury in rats. Crit. Care Med. 2005, 33, 2621–2628. [Google Scholar] [CrossRef]
- Altavilla, D.; Guarini, S.; Bitto, A.; Mioni, C.; Giuliani, D.; Bigiani, A.; Squadrito, G.; Minutoli, L.; Venuti, F.S.; Messineo, F.; et al. Activation of the cholinergic anti-inflammatory pathway reduces NF-kappab activation, blunts TNF-alpha production, and protects against splanchnic artery occlusion shock. Shock 2006, 25, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Guarini, S.; Altavilla, D.; Cainazzo, M.M.; Giuliani, D.; Bigiani, A.; Marini, H.; Squadrito, G.; Minutoli, L.; Bertolini, A.; Marini, R.; et al. Efferent vagal fibre stimulation blunts nuclear factor-kappaB activation and protects against hypovolemic hemorrhagic shock. Circulation 2003, 107, 1189–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohanka, M. Inhibitors of Acetylcholinesterase and Butyrylcholinesterase Meet Immunity. Int. J. Mol. Sci. 2014, 15, 9809–9825. [Google Scholar] [CrossRef] [Green Version]
- Gnatek, Y.; Zimmerman, G.; Goll, Y.; Najami, N.; Soreq, H.; Friedman, A. Acetylcholinesterase loosens the brain’s cholinergic anti-inflammatory response and promotes epileptogenesis. Front. Mol. Neurosci. 2012, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Acetylcholinesterase and butyrylcholinesterase as possible markers of low-grade systemic inflammation. Med. Sci. Monit. 2007, 13, RA214–RA221. [Google Scholar]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Expression and Function of the Cholinergic System in Immune Cells. Front. Immunol. 2017, 8, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, V.S.; Da Silva, A.S.; Costa, M.M.; Paim, F.C.; Alves, S.H.; Lopes, S.T.; Silva, C.B.; Wolkmer, P.; Castro, J.L.C.; Cecco, B.S.; et al. Cholinergic enzymes and inflammatory markers in rats infected by Sporothrix schenckii. Microb. Pathog. 2016, 97, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Souza, C.F.; Doleski, P.H.; Moreira, K.; da Veiga, M.L.; da Rocha, M.; Santos, R.; Baldisserotto, B. Involvement of cholinergic and adenosinergic systems on the branchial immune response of experimentally infected silvercatfish with Streptococcus agalactiae. J. Fish Dis. 2018, 41, 27–32. [Google Scholar] [CrossRef]
- Gilboa-Geffen, A.; Hartmann, G.; Soreq, H. Stressing hematopoiesis and immunity: An acetylcholinesterase window into nervous and immune system interactions. Front. Mol. Neurosci. 2012, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couraud, J.Y.; Di Giamberardino, L.; Chretien, M.; Souyri, F.; Fardeau, M. Acrylamide neuropathy and changes in the axonal transport and muscular content of the molecular forms of acetylcholinesterase. Muscle Nerve 1982, 5, 302–312. [Google Scholar] [CrossRef]
- Bai, Y.; Gu, Z.; Zhang, T.; Luo, Y.; Zhang, C.; Luo, L.; Ma, Y.; Liu, J. Toxic effects of subacute exposure to acrylamide on motor endplates of the gastrocnemius in rats. Toxicology 2021, 462, 152934. [Google Scholar] [CrossRef]
- Ngo, T.; Laidler, K. Immobilized electric eel acetylcholinesterase: I. Kinetics of acetylcholinesterase trapped in polyacrylamide membranes. Biochim. et Biophys. Acta (BBA)-Enzym. 1975, 377, 303–316. [Google Scholar] [CrossRef]
- Prasad, S.N. Muralidhara Evidence of acrylamide induced oxidative stress and neurotoxicity in Drosophila melanogaster—Its amelioration with spice active enrichment: Relevance to neuropathy. Neurotoxicology 2012, 33, 1254–1264. [Google Scholar] [CrossRef]
- Kopańska, M.; Lukáč, N.; Kapusta, E.; Formicki, G. Acrylamide Influence on Activity of Acetylcholinesterase, Thiol Groups, and Malondialdehyde Content in the Brain of Swiss Mice. J. Biochem. Mol. Toxicol. 2015, 29, 472–478. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, F.S.; Hu, X.; Chen, F.; Chan, H.M. Acrylamide-induced neurotoxicity in primary astrocytes and microglia: Roles of the Nrf2-ARE and NF-κB pathways. Food Chem. Toxicol. 2017, 106 Pt A, 25–35. [Google Scholar] [CrossRef]
- Liu, Z.; Song, G.; Zou, C.; Liu, G.; Wu, W.; Yuan, T.; Liu, X. Acrylamide induces mitochondrial dysfunction and apoptosis in BV-2 microglial cells. Free Radic. Biol. Med. 2015, 84, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Zong, C.; Hasegawa, R.; Urushitani, M.; Zhang, L.; Nagashima, D.; Sakurai, T.; Ichihara, S.; Ohsako, S.; Ichihara, G. Role of microglial activation and neuroinflammation in neurotoxicity of acrylamide in vivo and in vitro. Arch. Toxicol. 2019, 93, 2007–2019. [Google Scholar] [CrossRef] [PubMed]
- Ekuban, F.A.; Zong, C.; Takikawa, M.; Morikawa, K.; Sakurai, T.; Ichihara, S.; Itoh, K.; Yamamoto, M.; Ohsako, S.; Ichihara, G. Genetic ablation of Nrf2 exacerbates neurotoxic effects of acrylamide in mice. Toxicology 2021, 456, 152785. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acaroz, U.; Ince, S.; Arslan-Acaroz, D.; Gurler, Z.; Kucukkurt, I.; Demirel, H.H.; Arslan, H.O.; Varol, N.; Zhu, K. The ameliorative effects of boron against acrylamide-induced oxidative stress, inflammatory response, and metabolic changes in rats. Food Chem. Toxicol. 2018, 118, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Bo, N.; Yilin, H.; Chaoyue, Y.; Lu, L.; Yuan, Y. Acrylamide induces NLRP3 inflammasome activation via oxidative stress- and endoplasmic reticulum stress-mediated MAPK pathway in HepG2 cells. Food Chem. Toxicol. 2020, 145, 111679. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Yang, J.; Zhang, G.; Yuan, X.; Li, W.; Long, J.; Luo, Y.; Li, Y.; Wang, Y. NLRP3 inflammasome inhibition attenuates subacute neurotoxicity induced by acrylamide in vitro and in vivo. Toxicology 2020, 432, 152392. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| The Products Type | The Acrylamide Content (µg/kg) |
|---|---|
| Bread (rolls, bread, bagels) | 70–430 |
| Potato chips | <50–3500 |
| Potato fries | 200–2287 |
| Boiled potatoes | 48 |
| Cookies, crackers, biscuits | <30–3200 |
| Rusks | 80–1200 |
| Cereals | 30–1400 |
| Gingerbread cookies | <50–100 |
| Chocolate (powder) | 64–457 |
| Nuts, peanut butter | 64–457 |
| Meat, poultry | 30–64 |
| Baked asparagus | 143 |
| Authors | Aim | Methods | Results | Conclusions |
|---|---|---|---|---|
| Gnatek et al. [36] | Changes in the brain cholinergic gene expression and related immune responses during pilocarpine-induced epilepsy. | AChE levels and inflammatory markers were measured during pilocarpine-induced seizures in mice. | the action of pilocarpine leads to an increase in the level of AChE | Increased AChE levels are associated with an increased immune response. The cholinergic system is a potential area that can be influenced in order to reduce epileptic seizures. |
| Castro et al. [39] | AChE activity in serum, blood and lymphocytes and its relationship to the immune response after induced inflammation | The rats were divided into three groups that were infected by different routes. AChE activity in blood and lymphocytes was assessed at 15, 30 and 40 days after infection. | A significant increase in AChE activity was observed, especially in the intraperitoneally infected group. | AChE plays an important role in modulating the immune response. |
| Baldissera et al. [40] | Influence of the cholinergic system on the immune response and inflammation in the gills. | Ach levels in the gills of silver catfish were compared in fish with/without induced inflammation | Reduced acetylcholinesterase activity and increased acetylcholine levels in the gills of infected animals were observed. | the inflammatory process alters the cholinergic system, suggesting a contribution of AChE activity to the immune and inflammatory response by regulating ACh levels. The reduction in AChE activity exerts an anti-inflammatory profile. |
| Authors | Aim | Description of the Study | Results | Conclusions |
|---|---|---|---|---|
| Kopańska et al. [46] | Assessment of the effect of Acrylamide on the activity of Acetylcholinesterase in the hypothalamus, myocardium, skeletal muscles of the thigh and smooth muscles of the small intestine. | The effect of Acrylamide on the activity of Acetylcholinesterase in the hypothalamus, myocardium, skeletal muscles of the thigh and smooth muscles of the small intestine was measured depending on the thiol groups and the concentration of malondialdehyde. Swiss mice were used for the study. ACR was injected intraperitoneally at various doses. | AChE activity was significantly lower in all structures studied. The greatest decrease of 75.09% was observed in the hypothalamus 24 hours after uptake. This was accompanied by a statistically significant increase in malondialdehyde levels in most of the structures studied and at ACR doses.The highest increase in MDA was observed after 48 hours (45.12) and 192 hours (46.43) of exposure to ACR (40 mg/kg). | The assessment of Acetylcholinesterase activity in the muscles and hypothalamus of mice was very important due to the fact that many scientific reports indicate a direct effect of Acrylamide on peripheral nerves. It causes both their structural damage and negative physiological changes. After intraperitoneal injection of acrylamide, oxidative stress occurs in the body. |
| Prasad et al. [45] | Assessment of the potential of acrylamide to induce oxidative stress and neurotoxic effects in Drosophila melanogaster flies. | Adult male Drosophilia melanogaster flies have been exposed to dietary acrylamide. | ACR exposure resulted in dose-related and time-related mortality. The higher dose and the longer of the exposure to acrylamide, the worse results. Exposure to ACR caused locomotor deficits, severe oxidative stress, mitochondrial function, increased activity of acetylcholinesterase and decreased dopamine levels. | ACR-induced neurotoxicity may be mediated through an oxidative stress mechanism. ACR causes numerous physiological disorders, depending on the dose and time, such as abnormal modulation of nerve transmission by affecting the activity of AChE. |
| Couraud et al. [42] | To investigate the axonal transport of proteins in the proximal axons of the sciatic nerve in chickens as a result of acrylamide poisoning. | AChE occurs in nervous tissues of healthy chickens in four molecular forms (G1, G2, G4, A12). Three-day-old chickens were exposed to acrylamide at a dose of 100 mg/kg for 12 days. In the sciatic nerve of chickens poisoned with acrylamide, the action of individual forms of AChE was investigated. | Axonal transport of A12 was reduced by 60% and G4 by 21% due to ACR treatment. | ACR poisoning causes pathology in the action of some molecular forms of AChE. These forms can be considered sensitive markers of axonal transport and innervation phases. |
| Bai et al. [43] | To study the effect of exposure of the gastrocnemius motor plate in rats to ACR. | The rats were divided into groups. Different doses of ACR were administered to each group for 21 days. All rats were randomized into control groups, 9, 18 and 36 mg/kg. | As the dose of ACR exposure increased, more severe changes in the structure of muscle fibers and nerve endings were observed. Changes were observed in the structure of muscle fibers and nerve endings and a decrease in AChE content in motor plates. | ACR is toxic to the motor plate by altering the AChE content. |
| Ngo et al. [44] | To evaluate the effect of acrylamide on acetylcholinesterase. | The research was carried out on an electric eel. The study concerned the trapping of acetylcholinesterase from electric eel in a polyacrylamide gel. | AChE immobilized in the polyacrylamide gel showed a significantly reduced activity. | Acrylamide inhibits the activity of acetylcholinesterase. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopańska, M.; Łagowska, A.; Kuduk, B.; Banaś-Ząbczyk, A. Acrylamide Neurotoxicity as a Possible Factor Responsible for Inflammation in the Cholinergic Nervous System. Int. J. Mol. Sci. 2022, 23, 2030. https://doi.org/10.3390/ijms23042030
Kopańska M, Łagowska A, Kuduk B, Banaś-Ząbczyk A. Acrylamide Neurotoxicity as a Possible Factor Responsible for Inflammation in the Cholinergic Nervous System. International Journal of Molecular Sciences. 2022; 23(4):2030. https://doi.org/10.3390/ijms23042030
Chicago/Turabian StyleKopańska, Marta, Anna Łagowska, Barbara Kuduk, and Agnieszka Banaś-Ząbczyk. 2022. "Acrylamide Neurotoxicity as a Possible Factor Responsible for Inflammation in the Cholinergic Nervous System" International Journal of Molecular Sciences 23, no. 4: 2030. https://doi.org/10.3390/ijms23042030