
The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Methodology Employed for Study Selection
3. The Role of Interleukins in Embryo Implantation and Pregnancy Establishment
3.1. Basic Immunological Characteristics of the Endometrial Microenvironment during Implantation and Pregnancy
3.2. A Summary of Molecular Actions of Interleukins during Embryo Implantation
3.2.1. The Role of Interleukins with Anti-Inflammatory Properties
3.2.2. The Role of Interleukins with Pro-Inflammatory Properties
4. Clinical Data Associating Cytokine Profile with Recurrent Implantation Failure
4.1. Clinical Data Associating Leukemia Inhibitory Factor (LIF) with Recurrent Implantation Failure
4.2. Clinical Data Associating Anti and Pro Inflammatory Cytokines with Recurrent Implantation Failure
4.3. Clinical Data Associating Interleukins 12, 15 and 18 with Recurrent Implantation Failure
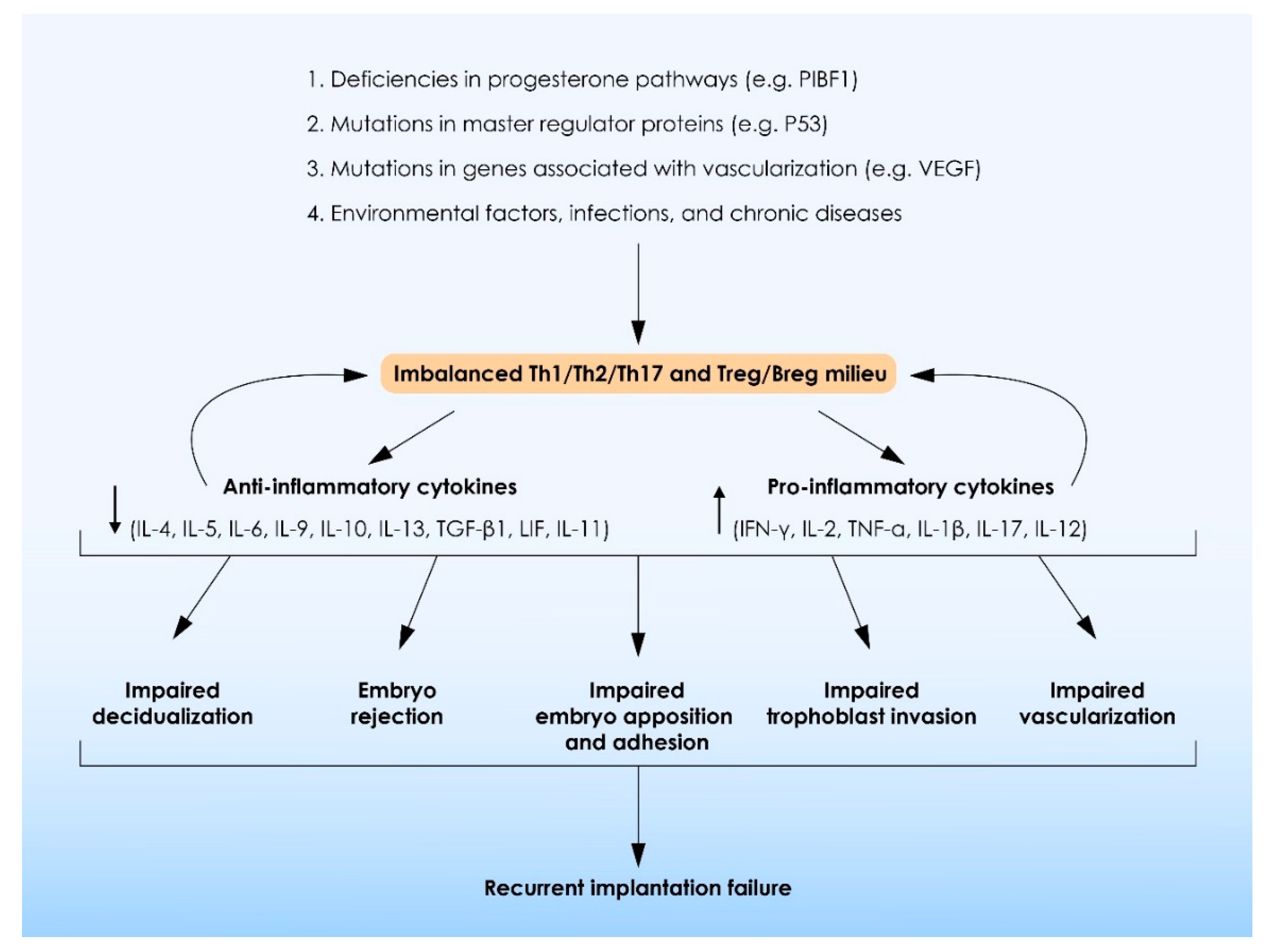
4.4. Abnormally Increased Levels of Pro-Inflammatory Cytokines Originating from Abnormal Regulation of T and B Regulatory Cell Function Are Associated with Recurrent Implantation Failure
4.5. Alterations in Upstream Pathways Regulating Interleukin Production Are Associated with Recurrent Implantation Failure
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández-Vargas, P.; Muñoz, M.; Domínguez, F. Identifying Biomarkers for Predicting Successful Embryo Implantation: Applying Single to Multi-OMICs to Improve Reproductive Outcomes. Hum. Reprod. Update 2020, 26, 264–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-M.; Kim, J.-S. A Review of Mechanisms of Implantation. Dev. Reprod. 2017, 21, 351–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Huang, X.; Mor, G.; Liao, A. Epigenetic Modifications Working in the Decidualization and Endometrial Receptivity. Cell. Mol. Life Sci. 2020, 77, 2091–2101. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.-W.; Norwitz, G.A.; Pavlicev, M.; Tilburgs, T.; Simón, C.; Norwitz, E.R. Endometrial Decidualization: The Primary Driver of Pregnancy Health. Int. J. Mol.Sci. 2020, 21, 4092. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Tsuzuki, T.; Murata, H. Decidualization of the Human Endometrium. Reprod. Med. Biol. 2018, 17, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Song, H.; Das, S.K.; Paria, B.C.; Dey, S.K. Estrogen Is a Critical Determinant That Specifies the Duration of the Window of Uterine Receptivity for Implantation. Proc. Natl. Acad. Sci. USA 2003, 100, 2963–2968. [Google Scholar] [CrossRef] [Green Version]
- Diedrich, K.; Fauser, B.C.J.M.; Devroey, P.; Griesinger, G. Evian Annual Reproduction (EVAR) Workshop Group The Role of the Endometrium and Embryo in Human Implantation. Hum. Reprod. Update 2007, 13, 365–377. [Google Scholar] [CrossRef]
- Bischof, P.; Campana, A. Trophoblast Differentiation and Invasion: Its Significance for Human Embryo Implantation. Early Pregnancy 1997, 3, 81–95. [Google Scholar]
- Ochoa-Bernal, M.A.; Fazleabas, A.T. Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 1973. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, C.; Ledger, W.; Wang, Q.; Liu, F.; Demirol, A.; Gurgan, T.; Cutting, R.; Ong, K.; Sallam, H.; Li, T.C. Recurrent Implantation Failure: Definition and Management. Reprod. Biomed. Online 2014, 28, 14–38. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhang, Y.; Ma, X.; Jia, W.; Su, Y. Determining Diagnostic Criteria of Unexplained Recurrent Implantation Failure: A Retrospective Study of Two vs Three or More Implantation Failure. Front. Endocrinol. 2021, 12, 619437. [Google Scholar] [CrossRef] [PubMed]
- Cimadomo, D.; Craciunas, L.; Vermeulen, N.; Vomstein, K.; Toth, B. Definition, Diagnostic and Therapeutic Options in Recurrent Implantation Failure: An International Survey of Clinicians and Embryologists. Hum. Reprod. 2021, 36, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Bashiri, A.; Halper, K.I.; Orvieto, R. Recurrent Implantation Failure-Update Overview on Etiology, Diagnosis, Treatment and Future Directions. Reprod. Biol. Endocrinol. 2018, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margalioth, E.J.; Ben-Chetrit, A.; Gal, M.; Eldar-Geva, T. Investigation and Treatment of Repeated Implantation Failure Following IVF-ET. Hum. Reprod. 2006, 21, 3036–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polanski, L.T.; Baumgarten, M.N.; Quenby, S.; Brosens, J.; Campbell, B.K.; Raine-Fenning, N.J. What Exactly Do We Mean by ‘Recurrent Implantation Failure’? A Systematic Review and Opinion. Reprod. Biomed. Online 2014, 28, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascarenhas, M.; Jeve, Y.; Polanski, L.; Sharpe, A.; Yasmin, E.; Bhandari, H.M. Management of Recurrent Implantation Failure: British Fertility Society Policy and Practice Guideline. Hum. Fertil. 2021, 1–25. [Google Scholar] [CrossRef]
- Chen, X.; Mariee, N.; Jiang, L.; Liu, Y.; Wang, C.C.; Li, T.C.; Laird, S. Measurement of Uterine Natural Killer Cell Percentage in the Periimplantation Endometrium from Fertile Women and Women with Recurrent Reproductive Failure: Establishment of a Reference Range. Am. J. Obstet. Gynecol. 2017, 217, 680.e1–680.e6. [Google Scholar] [CrossRef]
- Kolanska, K.; Bendifallah, S.; Cohen, J.; Placais, L.; Selleret, L.; Johanet, C.; Suner, L.; Delhommeau, F.; Chabbert-Buffet, N.; Darai, E.; et al. Unexplained Recurrent Implantation Failures: Predictive Factors of Pregnancy and Therapeutic Management from a French Multicentre Study. J. Reprod. Immunol. 2021, 145, 103313. [Google Scholar] [CrossRef]
- Lédée, N.; Petitbarat, M.; Chevrier, L.; Vitoux, D.; Vezmar, K.; Rahmati, M.; Dubanchet, S.; Gahéry, H.; Bensussan, A.; Chaouat, G. The Uterine Immune Profile May Help Women with Repeated Unexplained Embryo Implantation Failure after In Vitro Fertilization. Am. J. Reprod. Immunol. 2016, 75, 388–401. [Google Scholar] [CrossRef] [Green Version]
- Amjadi, F.; Zandieh, Z.; Mehdizadeh, M.; Aghajanpour, S.; Raoufi, E.; Aghamajidi, A.; Aflatoonian, R. The Uterine Immunological Changes May Be Responsible for Repeated Implantation Failure. J. Reprod. Immunol. 2020, 138, 103080. [Google Scholar] [CrossRef]
- Chaouat, G.; Dubanchet, S.; Ledée, N. Cytokines: Important for Implantation? J. Assist. Reprod. Genet. 2007, 24, 491–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Chaudhry, P.; Asselin, E. Bridging Endometrial Receptivity and Implantation: Network of Hormones, Cytokines, and Growth Factors. J. Endocrinol. 2011, 210, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitriadis, E.; White, C.A.; Jones, R.L.; Salamonsen, L.A. Cytokines, Chemokines and Growth Factors in Endometrium Related to Implantation. Hum. Reprod. Update 2005, 11, 613–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makrigiannakis, A.; Petsas, G.; Toth, B.; Relakis, K.; Jeschke, U. Recent Advances in Understanding Immunology of Reproductive Failure. J. Reprod. Immunol. 2011, 90, 96–104. [Google Scholar] [CrossRef]
- Liang, P.-Y.; Diao, L.-H.; Huang, C.-Y.; Lian, R.-C.; Chen, X.; Li, G.-G.; Zhao, J.; Li, Y.-Y.; He, X.-B.; Zeng, Y. The Pro-Inflammatory and Anti-Inflammatory Cytokine Profile in Peripheral Blood of Women with Recurrent Implantation Failure. Reprod. Biomed. Online 2015, 31, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zheng, Q.; Jin, L. Dynamic Function and Composition Changes of Immune Cells during Normal and Pathological Pregnancy at the Maternal-Fetal Interface. Front. Immunol. 2019, 10, 2317. [Google Scholar] [CrossRef] [Green Version]
- Lédée, N.; Petitbarat, M.; Prat-Ellenberg, L.; Dray, G.; Cassuto, G.-N.; Chevrier, L.; Kazhalawi, A.; Vezmar, K.; Chaouat, G. The Uterine Immune Profile: A Method for Individualizing the Management of Women Who Have Failed to Implant an Embryo after IVF/ICSI. J. Reprod. Immunol. 2020, 142, 103207. [Google Scholar] [CrossRef]
- Franasiak, J.M.; Alecsandru, D.; Forman, E.J.; Gemmell, L.C.; Goldberg, J.M.; Llarena, N.; Margolis, C.; Laven, J.; Schoenmakers, S.; Seli, E. A Review of the Pathophysiology of Recurrent Implantation Failure. Fertil. Steril. 2021, 116, 1436–1448. [Google Scholar] [CrossRef]
- Franasiak, J.M.; Scott, R.T. Contribution of Immunology to Implantation Failure of Euploid Embryos. Fertil. Steril. 2017, 107, 1279–1283. [Google Scholar] [CrossRef] [Green Version]
- Aimagambetova, G.; Issanov, A.; Terzic, S.; Bapayeva, G.; Ukybassova, T.; Baikoshkarova, S.; Aldiyarova, A.; Shauyen, F.; Terzic, M. The Effect of Psychological Distress on IVF Outcomes: Reality or Speculations? PLoS ONE 2020, 15, e0242024. [Google Scholar] [CrossRef]
- Shaulov, T.; Sierra, S.; Sylvestre, C. Recurrent Implantation Failure in IVF: A Canadian Fertility and Andrology Society Clinical Practice Guideline. Reprod. Biomed. Online 2020, 41, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-X.; Lin, S.; Kong, S.-B. Psychological Stress and Functional Endometrial Disorders: Update of Mechanism Insights. Front. Endocrinol. 2021, 12, 690255. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Evans, J.; Nguyen, H.P.T.; Edgell, T.A. The Microenvironment of Human Implantation: Determinant of Reproductive Success. Am. J. Reprod. Immunol. 2016, 75, 218–225. [Google Scholar] [CrossRef]
- Sternberg, A.K.; Buck, V.U.; Classen-Linke, I.; Leube, R.E. How Mechanical Forces Change the Human Endometrium during the Menstrual Cycle in Preparation for Embryo Implantation. Cells 2021, 10, 2008. [Google Scholar] [CrossRef]
- Kelly, R.W.; King, A.E.; Critchley, H.O. Cytokine Control in Human Endometrium. Reproduction 2001, 121, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Rajaei, S.; Zarnani, A.H.; Jeddi-Tehrani, M.; Tavakoli, M.; Mohammadzadeh, A.; Dabbagh, A.; Mirahmadian, M. Cytokine Profile in the Endometrium of Normal Fertile and Women with Repeated Implantation Failure. Iran. J. Immunol. 2011, 8, 201–208. [Google Scholar]
- Saito, S. Cytokine Network at the Feto-Maternal Interface. J. Reprod. Immunol. 2000, 47, 87–103. [Google Scholar] [CrossRef]
- Liu, J.; Hao, S.; Chen, X.; Zhao, H.; Du, L.; Ren, H.; Wang, C.; Mao, H. Human Placental Trophoblast Cells Contribute to Maternal-Fetal Tolerance through Expressing IL-35 and Mediating ITR35 Conversion. Nat. Commun. 2019, 10, 4601. [Google Scholar] [CrossRef]
- Krzymowski, T.; Stefańczyk-Krzymowska, S. Advances in Understanding the Physiological Mechanism of Maternal Immune Tolerance to the Embryo. Reprod. Biol. 2012, 12, 265–270. [Google Scholar] [CrossRef]
- Warning, J.C.; McCracken, S.A.; Morris, J.M. A Balancing Act: Mechanisms by Which the Fetus Avoids Rejection by the Maternal Immune System. Reproduction 2011, 141, 715–724. [Google Scholar] [CrossRef] [Green Version]
- Bert, S.; Ward, E.J.; Nadkarni, S. Neutrophils in Pregnancy: New Insights into Innate and Adaptive Immune Regulation. Immunology 2021, 164, 665–676. [Google Scholar] [CrossRef] [PubMed]
- PrabhuDas, M.; Bonney, E.; Caron, K.; Dey, S.; Erlebacher, A.; Fazleabas, A.; Fisher, S.; Golos, T.; Matzuk, M.; McCune, J.M.; et al. Immune Mechanisms at the Maternal-Fetal Interface: Perspectives and Challenges. Nat. Immunol. 2015, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Lobo, S.C.; Huang, S.-T.J.; Germeyer, A.; Dosiou, C.; Vo, K.C.; Tulac, S.; Nayak, N.R.; Giudice, L.C. The Immune Environment in Human Endometrium during the Window of Implantation. Am. J. Reprod. Immunol. 2004, 52, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Bulmer, J.N.; Morrison, L.; Longfellow, M.; Ritson, A.; Pace, D. Granulated Lymphocytes in Human Endometrium: Histochemical and Immunohistochemical Studies. Hum. Reprod. 1991, 6, 791–798. [Google Scholar] [CrossRef]
- Faas, M.M.; de Vos, P. Uterine NK Cells and Macrophages in Pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Kitazawa, J.; Kimura, F.; Nakamura, A.; Morimune, A.; Takahashi, A.; Takashima, A.; Amano, T.; Tsuji, S.; Kaku, S.; Kasahara, K.; et al. Endometrial Immunity for Embryo Implantation and Pregnancy Establishment. Tohoku J. Exp. Med. 2020, 250, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopez, N.; Guilbert, L.J.; Olson, D.M. Invasion of the Leukocytes into the Fetal-Maternal Interface during Pregnancy. J. Leukoc. Biol. 2010, 88, 625–633. [Google Scholar] [CrossRef]
- Nancy, P.; Erlebacher, A. T Cell Behavior at the Maternal-Fetal Interface. Int. J. Dev. Biol. 2014, 58, 189–198. [Google Scholar] [CrossRef] [Green Version]
- von Rango, U. Fetal Tolerance in Human Pregnancy—A Crucial Balance between Acceptance and Limitation of Trophoblast Invasion. Immunol. Lett. 2008, 115, 21–32. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, M.; Lee, S.K. Role of Endometrial Immune Cells in Implantation. Clin. Exp. Reprod. Med. 2011, 38, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Timeva, T.; Shterev, A.; Kyurkchiev, S. Recurrent Implantation Failure: The Role of the Endometrium. J. Reprod. Infertil. 2014, 15, 173–183. [Google Scholar] [PubMed]
- Sfakianoudis, K.; Rapani, A.; Grigoriadis, S.; Pantou, A.; Maziotis, E.; Kokkini, G.; Tsirligkani, C.; Bolaris, S.; Nikolettos, K.; Chronopoulou, M.; et al. The Role of Uterine Natural Killer Cells on Recurrent Miscarriage and Recurrent Implantation Failure: From Pathophysiology to Treatment. Biomedicines 2021, 9, 1425. [Google Scholar] [CrossRef] [PubMed]
- Jena, M.K.; Nayak, N.; Chen, K.; Nayak, N.R. Role of Macrophages in Pregnancy and Related Complications. Arch. Immunol. Ther. Exp. 2019, 67, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Harrity, C.; Shkrobot, L.; Walsh, D.; Marron, K. ART Implantation Failure and Miscarriage in Patients with Elevated Intracellular Cytokine Ratios: Response to Immune Support Therapy. Fertil. Res. Pract. 2018, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Mor, G. Inflammation and Implantation. Am. J. Reprod. Immunol. 2010, 63, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and Pregnancy: The Role of the Immune System at the Implantation Site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-J.; Liu, Y.-L.; Sytwu, H.-K. Immunologic Regulation in Pregnancy: From Mechanism to Therapeutic Strategy for Immunomodulation. Clin. Dev. Immunol. 2012, 2012, 258391. [Google Scholar] [CrossRef]
- Romero, R.; Espinoza, J.; Gonçalves, L.F.; Kusanovic, J.P.; Friel, L.A.; Nien, J.K. Inflammation in Preterm and Term Labour and Delivery. Semin. Fetal Neonatal Med. 2006, 11, 317–326. [Google Scholar] [CrossRef]
- Dealtry, G.B.; O’Farrell, M.K.; Fernandez, N. The Th2 Cytokine Environment of the Placenta. Int. Arch. Allergy Immunol. 2000, 123, 107–119. [Google Scholar] [CrossRef]
- Perez-Sepulveda, A.; Torres, M.J.; Khoury, M.; Illanes, S.E. Innate Immune System and Preeclampsia. Front. Immunol. 2014, 5, 244. [Google Scholar] [CrossRef] [Green Version]
- Sykes, L.; MacIntyre, D.A.; Yap, X.J.; Teoh, T.G.; Bennett, P.R. The Th1:Th2 Dichotomy of Pregnancy and Preterm Labour. Mediat. Inflamm. 2012, 2012, 967629. [Google Scholar] [CrossRef] [Green Version]
- van Mourik, M.S.M.; Macklon, N.S.; Heijnen, C.J. Embryonic Implantation: Cytokines, Adhesion Molecules, and Immune Cells in Establishing an Implantation Environment. J. Leukoc. Biol. 2009, 85, 4–19. [Google Scholar] [CrossRef] [Green Version]
- Markert, U.R.; Morales-Prieto, D.M.; Fitzgerald, J.S. Understanding the Link between the IL-6 Cytokine Family and Pregnancy: Implications for Future Therapeutics. Expert Rev. Clin. Immunol. 2011, 7, 603–609. [Google Scholar] [CrossRef]
- Metcalfe, R.D.; Putoczki, T.L.; Griffin, M.D.W. Structural Understanding of Interleukin 6 Family Cytokine Signaling and Targeted Therapies: Focus on Interleukin 11. Front. Immunol. 2020, 11, 1424. [Google Scholar] [CrossRef]
- Murakami, M.; Kamimura, D.; Hirano, T. Pleiotropy and Specificity: Insights from the Interleukin 6 Family of Cytokines. Immunity 2019, 50, 812–831. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Chávez, F.; López-Portales, Ó.H.; Baeza-Martínez, D.A.; Cancino-Díaz, J.C.; Murrieta-Coxca, J.M.; Cancino-Díaz, M.E.; Pérez-Tapia, S.M.; Rodríguez-Martínez, S. IκBNS and IL-6 Expression Is Differentially Established in the Uterus of Pregnant Healthy and Infected Mice. Heliyon 2020, 6, e04122. [Google Scholar] [CrossRef]
- Aghajanova, L. Leukemia Inhibitory Factor and Human Embryo Implantation. Ann. N. Y. Acad. Sci. 2004, 1034, 176–183. [Google Scholar] [CrossRef]
- Huang, C.; Sun, H.; Wang, Z.; Liu, Y.; Cheng, X.; Liu, J.; Jiang, R.; Zhang, X.; Zhen, X.; Zhou, J.; et al. Increased Krüppel-like Factor 12 Impairs Embryo Attachment via Downregulation of Leukemia Inhibitory Factor in Women with Recurrent Implantation Failure. Cell Death Discov. 2018, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, R.; Terakawa, J.; Kato, Y.; Azimi, S.; Inoue, N.; Ohmori, Y.; Hondo, E. The Contribution of Leukemia Inhibitory Factor (LIF) for Embryo Implantation Differs among Strains of Mice. Immunobiology 2014, 219, 512–521. [Google Scholar] [CrossRef]
- Salleh, N.; Giribabu, N. Leukemia Inhibitory Factor: Roles in Embryo Implantation and in Nonhormonal Contraception. Sci. World J. 2014, 2014, 201514. [Google Scholar] [CrossRef] [Green Version]
- Kondera-Anasz, Z.; Sikora, J.; Mielczarek-Palacz, A. Leukemia Inhibitory Factor: An Important Regulator of Endometrial Function. Am. J. Reprod. Immunol. 2004, 52, 97–105. [Google Scholar] [CrossRef]
- Sun, X.; Bartos, A.; Whitsett, J.A.; Dey, S.K. Uterine Deletion of Gp130 or Stat3 Shows Implantation Failure with Increased Estrogenic Responses. Mol. Endocrinol. 2013, 27, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Catalano, R.D.; Johnson, M.H.; Campbell, E.A.; Charnock-Jones, D.S.; Smith, S.K.; Sharkey, A.M. Inhibition of Stat3 Activation in the Endometrium Prevents Implantation: A Nonsteroidal Approach to Contraception. Proc. Natl. Acad. Sci. USA 2005, 102, 8585–8590. [Google Scholar] [CrossRef] [Green Version]
- Moberg, C.; Bourlev, V.; Ilyasova, N.; Olovsson, M. Endometrial Expression of LIF and Its Receptor and Peritoneal Fluid Levels of IL-1α and IL-6 in Women with Endometriosis Are Associated with the Probability of Pregnancy. Arch. Gynecol. Obstet. 2015, 292, 429–437. [Google Scholar] [CrossRef]
- Massimiani, M.; Lacconi, V.; La Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular Signaling Regulating Endometrium-Blastocyst Crosstalk. Int. J. Mol. Sci. 2019, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, E.; Stoikos, C.; Tan, Y.-L.; Salamonsen, L.A. Interleukin 11 Signaling Components Signal Transducer and Activator of Transcription 3 (STAT3) and Suppressor of Cytokine Signaling 3 (SOCS3) Regulate Human Endometrial Stromal Cell Differentiation. Endocrinology 2006, 147, 3809–3817. [Google Scholar] [CrossRef]
- Rosario, G.X.; Hondo, E.; Jeong, J.-W.; Mutalif, R.; Ye, X.; Yee, L.X.; Stewart, C.L. The LIF-Mediated Molecular Signature Regulating Murine Embryo Implantation. Biol. Reprod. 2014, 91, 66. [Google Scholar] [CrossRef]
- Rosario, G.X.; Stewart, C.L. The Multifaceted Actions of Leukaemia Inhibitory Factor in Mediating Uterine Receptivity and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Fedorcsák, P.; Storeng, R. Effects of Leptin and Leukemia Inhibitory Factor on Preimplantation Development and STAT3 Signaling of Mouse Embryos In Vitro. Biol. Reprod. 2003, 69, 1531–1538. [Google Scholar] [CrossRef]
- Tabibzadeh, S.; Kong, Q.F.; Babaknia, A.; May, L.T. Progressive Rise in the Expression of Interleukin-6 in Human Endometrium during Menstrual Cycle Is Initiated during the Implantation Window. Hum. Reprod. 1995, 10, 2793–2799. [Google Scholar] [CrossRef]
- Champion, H.; Innes, B.A.; Robson, S.C.; Lash, G.E.; Bulmer, J.N. Effects of Interleukin-6 on Extravillous Trophoblast Invasion in Early Human Pregnancy. Mol. Hum. Reprod. 2012, 18, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Perrier d’Hauterive, S.; Charlet-Renard, C.; Berndt, S.; Dubois, M.; Munaut, C.; Goffin, F.; Hagelstein, M.-T.; Noël, A.; Hazout, A.; Foidart, J.-M.; et al. Human Chorionic Gonadotropin and Growth Factors at the Embryonic-Endometrial Interface Control Leukemia Inhibitory Factor (LIF) and Interleukin 6 (IL-6) Secretion by Human Endometrial Epithelium. Hum. Reprod. 2004, 19, 2633–2643. [Google Scholar] [CrossRef] [Green Version]
- Gridelet, V.; Perrier d’Hauterive, S.; Polese, B.; Foidart, J.-M.; Nisolle, M.; Geenen, V. Human Chorionic Gonadotrophin: New Pleiotropic Functions for an “Old” Hormone During Pregnancy. Front. Immunol. 2020, 11, 343. [Google Scholar] [CrossRef] [Green Version]
- Habbeddine, M.; Verbeke, P.; Karaz, S.; Bobé, P.; Kanellopoulos-Langevin, C. Leukocyte Population Dynamics and Detection of IL-9 as a Major Cytokine at the Mouse Fetal-Maternal Interface. PLoS ONE 2014, 9, e107267. [Google Scholar] [CrossRef]
- Piccinni, M.-P.; Raghupathy, R.; Saito, S.; Szekeres-Bartho, J. Cytokines, Hormones and Cellular Regulatory Mechanisms Favoring Successful Reproduction. Front. Immunol. 2021, 12, 717808. [Google Scholar] [CrossRef]
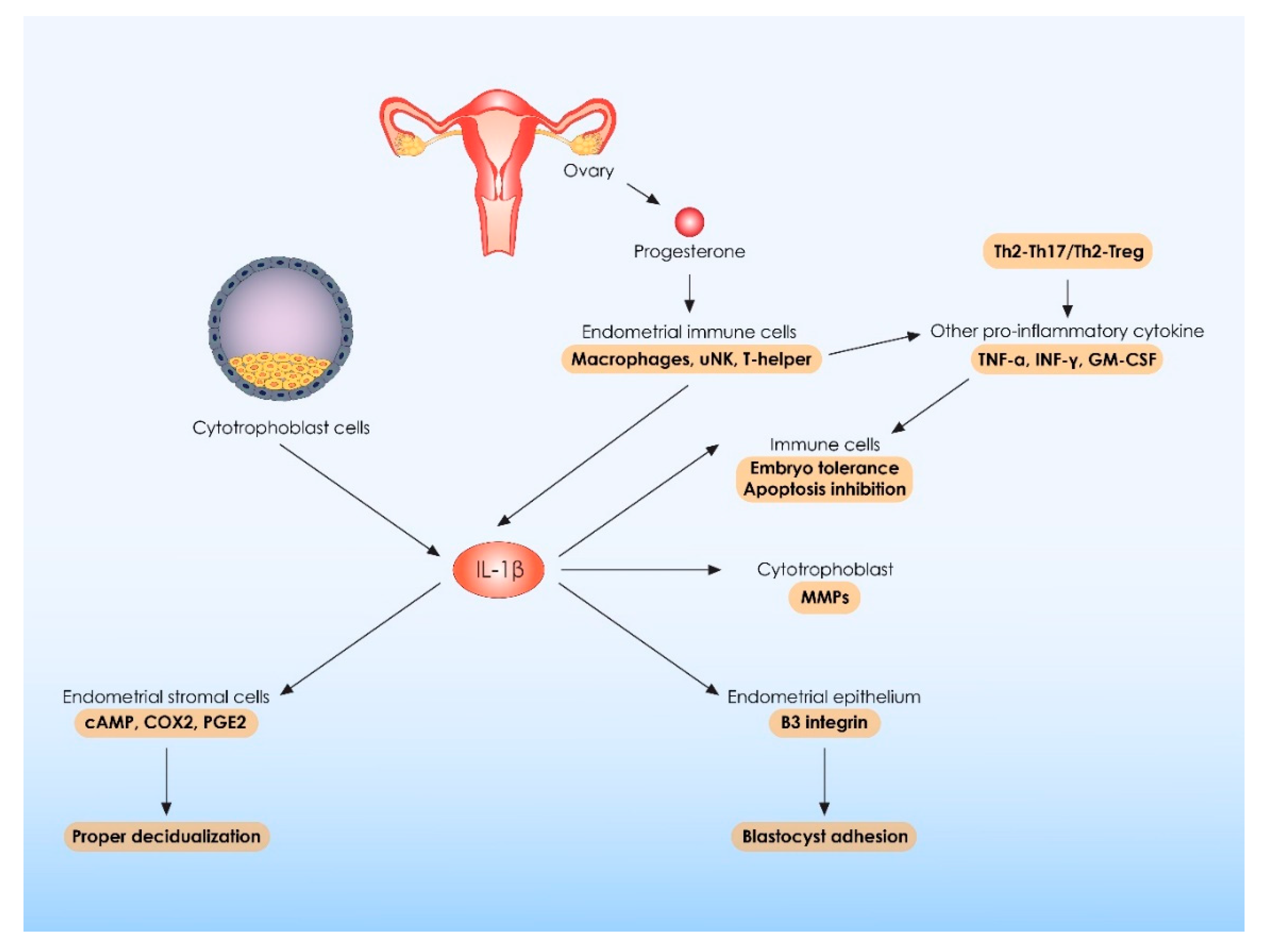
- White, C.A.; Dimitriadis, E.; Sharkey, A.M.; Stoikos, C.J.; Salamonsen, L.A. Interleukin 1 Beta Is Induced by Interleukin 11 during Decidualization of Human Endometrial Stromal Cells, but Is Not Released in a Bioactive Form. J. Reprod. Immunol. 2007, 73, 28–38. [Google Scholar] [CrossRef]
- Equils, O.; Kellogg, C.; McGregor, J.; Gravett, M.; Neal-Perry, G.; Gabay, C. The Role of the IL-1 System in Pregnancy and the Use of IL-1 System Markers to Identify Women at Risk for Pregnancy Complications. Biol. Reprod. 2020, 103, 684–694. [Google Scholar] [CrossRef]
- Fazleabas, A.T.; Kim, J.J.; Strakova, Z. Implantation: Embryonic Signals and the Modulation of the Uterine Environment--A Review. Placenta 2004, 25, S26–S31. [Google Scholar] [CrossRef]
- Townson, D.H.; Pate, J.L. Regulation of Prostaglandin Synthesis by Interleukin-1 Beta in Cultured Bovine Luteal Cells. Biol. Reprod. 1994, 51, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Miceli, F.; Tropea, A.; Minici, F.; Navarra, P.; Lanzone, A.; Apa, R. Interleukin-1β Stimulates Progesterone Production by in Vitro Human Luteal Cells: Evidence of a Mediatory Role of Prostaglandins. J. Clin. Endocrinol. Metab. 2003, 88, 2690–2694. [Google Scholar] [CrossRef] [Green Version]
- Keir, M.; Yi, T.; Lu, T.; Ghilardi, N. The Role of IL-22 in Intestinal Health and Disease. J. Exp. Med. 2020, 217, e20192195. [Google Scholar] [CrossRef]
- Pavlov, O.; Selutin, A.; Pavlova, O.; Selkov, S. Macrophages Are a Source of IL-17 in the Human Placenta. Am. J. Reprod. Immunol. 2018, 80, e13016. [Google Scholar] [CrossRef]
- Ruiz de Morales, J.M.G.; Puig, L.; Daudén, E.; Cañete, J.D.; Pablos, J.L.; Martín, A.O.; Juanatey, C.G.; Adán, A.; Montalbán, X.; Borruel, N.; et al. Critical Role of Interleukin (IL)-17 in Inflammatory and Immune Disorders: An Updated Review of the Evidence Focusing in Controversies. Autoimmun. Rev. 2020, 19, 102429. [Google Scholar] [CrossRef]
- Sabat, R.; Ouyang, W.; Wolk, K. Therapeutic Opportunities of the IL-22-IL-22R1 System. Nat. Rev. Drug Discov. 2014, 13, 21–38. [Google Scholar] [CrossRef]
- Annunziato, F.; Cosmi, L.; Santarlasci, V.; Maggi, L.; Liotta, F.; Mazzinghi, B.; Parente, E.; Filì, L.; Ferri, S.; Frosali, F.; et al. Phenotypic and Functional Features of Human Th17 Cells. J. Exp. Med. 2007, 204, 1849–1861. [Google Scholar] [CrossRef] [Green Version]
- Cosmi, L.; De Palma, R.; Santarlasci, V.; Maggi, L.; Capone, M.; Frosali, F.; Rodolico, G.; Querci, V.; Abbate, G.; Angeli, R.; et al. Human Interleukin 17-Producing Cells Originate from a CD161+CD4+ T Cell Precursor. J. Exp. Med. 2008, 205, 1903–1916. [Google Scholar] [CrossRef]
- Saito, S.; Nakashima, A.; Shima, T.; Ito, M. Th1/Th2/Th17 and Regulatory T-Cell Paradigm in Pregnancy. Am. J. Reprod. Immunol. 2010, 63, 601–610. [Google Scholar] [CrossRef]
- Wang, W.-J.; Hao, C.-F.; Lin, Y.; Yin, G.-J.; Bao, S.-H.; Qiu, L.-H.; Lin, Q.-D. Increased Prevalence of T Helper 17 (Th17) Cells in Peripheral Blood and Decidua in Unexplained Recurrent Spontaneous Abortion Patients. J. Reprod. Immunol. 2010, 84, 164–170. [Google Scholar] [CrossRef]
- de Lima Kaminski, V.; Ellwanger, J.H.; Matte, M.C.C.; Savaris, R.F.; Vianna, P.; Chies, J.A.B. IL-17 Blood Levels Increase in Healthy Pregnancy but Not in Spontaneous Abortion. Mol. Biol. Rep. 2018, 45, 1565–1568. [Google Scholar] [CrossRef]
- Wu, H.-X.; Jin, L.-P.; Xu, B.; Liang, S.-S.; Li, D.-J. Decidual Stromal Cells Recruit Th17 Cells into Decidua to Promote Proliferation and Invasion of Human Trophoblast Cells by Secreting IL-17. Cell. Mol. Immunol. 2014, 11, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Hambartsoumian, E. Endometrial Leukemia Inhibitory Factor (LIF) as a Possible Cause of Unexplained Infertility and Multiple Failures of Implantation. Am. J. Reprod. Immunol. 1998, 39, 137–143. [Google Scholar] [CrossRef]
- Steck, T.; Giess, R.; Suetterlin, M.W.; Bolland, M.; Wiest, S.; Poehls, U.G.; Dietl, J. Leukaemia Inhibitory Factor (LIF) Gene Mutations in Women with Unexplained Infertility and Recurrent Failure of Implantation after IVF and Embryo Transfer. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 112, 69–73. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, H.-R.; Lim, E.J.; Park, M.; Yoon, J.A.; Kim, Y.S.; Kim, E.-K.; Shin, J.-E.; Kim, J.H.; Kwon, H.; et al. Integrative Analyses of Uterine Transcriptome and MicroRNAome Reveal Compromised LIF-STAT3 Signaling and Progesterone Response in the Endometrium of Patients with Recurrent/Repeated Implantation Failure (RIF). PLoS ONE 2016, 11, e0157696. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, N.; Stern, C.; McBain, J.; Lopata, A.; Kornman, L.; Wilkinson, D. Analysis of Intra-Uterine Cytokine Concentration and Matrix-Metalloproteinase Activity in Women with Recurrent Failed Embryo Transfer. Hum. Reprod. 2003, 18, 608–615. [Google Scholar] [CrossRef]
- Boomsma, C.M.; Kavelaars, A.; Eijkemans, M.J.C.; Lentjes, E.G.; Fauser, B.C.J.M.; Heijnen, C.J.; Macklon, N.S. Endometrial Secretion Analysis Identifies a Cytokine Profile Predictive of Pregnancy in IVF. Hum. Reprod. 2009, 24, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, K.; Yamaguchi, T.; Honjo, H. Central Role of Interleukin-15 in Postovulatory Recruitment of Peripheral Blood CD16(-) Natural Killer Cells into Human Endometrium. J. Clin. Endocrinol. Metab. 2005, 90, 2932–2940. [Google Scholar] [CrossRef] [Green Version]
- Murata, H.; Tanaka, S.; Okada, H. Immune Tolerance of the Human Decidua. J. Clin. Med. 2021, 10, 351. [Google Scholar] [CrossRef]
- Mariee, N.; Li, T.C.; Laird, S.M. Expression of Leukaemia Inhibitory Factor and Interleukin 15 in Endometrium of Women with Recurrent Implantation Failure after IVF.; Correlation with the Number of Endometrial Natural Killer Cells. Hum. Reprod. 2012, 27, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Ida, A.; Tsuji, Y.; Muranaka, J.; Kanazawa, R.; Nakata, Y.; Adachi, S.; Okamura, H.; Koyama, K. IL-18 in Pregnancy; the Elevation of IL-18 in Maternal Peripheral Blood during Labour and Complicated Pregnancies. J. Reprod. Immunol. 2000, 47, 65–74. [Google Scholar] [CrossRef]
- Löb, S.; Ochmann, B.; Ma, Z.; Vilsmaier, T.; Kuhn, C.; Schmoeckel, E.; Herbert, S.-L.; Kolben, T.; Wöckel, A.; Mahner, S.; et al. The Role of Interleukin-18 in Recurrent Early Pregnancy Loss. J. Reprod. Immunol. 2021, 148, 103432. [Google Scholar] [CrossRef] [PubMed]
- French, A.R.; Holroyd, E.B.; Yang, L.; Kim, S.; Yokoyama, W.M. IL-18 Acts Synergistically with IL-15 in Stimulating Natural Killer Cell Proliferation. Cytokine 2006, 35, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, S.; Nagai, N.; Kanaeda, T.; Karasaki-Suzuki, M.; Ishii, M.; Chishima, F.; Satoh, K. Interleukin-12 Augments Cytolytic Activity of Peripheral and Decidual Lymphocytes against Choriocarcinoma Cell Lines and Primary Culture Human Placental Trophoblasts. Am. J. Reprod. Immunol. 1999, 41, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Lédée-Bataille, N.; Dubanchet, S.; Coulomb-L’hermine, A.; Durand-Gasselin, I.; Frydman, R.; Chaouat, G. A New Role for Natural Killer Cells, Interleukin (IL)-12, and IL-18 in Repeated Implantation Failure after in Vitro Fertilization. Fertil. Steril. 2004, 81, 59–65. [Google Scholar] [CrossRef]
- Ma, X.; Yan, W.; Zheng, H.; Du, Q.; Zhang, L.; Ban, Y.; Li, N.; Wei, F. Regulation of IL-10 and IL-12 Production and Function in Macrophages and Dendritic Cells. F1000Res 2015, 4, F1000 Faculty Rev-1465. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, C.M.; Kavelaars, A.; Eijkemans, M.J.C.; Amarouchi, K.; Teklenburg, G.; Gutknecht, D.; Fauser, B.J.C.M.; Heijnen, C.J.; Macklon, N.S. Cytokine Profiling in Endometrial Secretions: A Non-Invasive Window on Endometrial Receptivity. Reprod. Biomed. Online 2009, 18, 85–94. [Google Scholar] [CrossRef]
- Huang, H.-Y.; Chan, S.-H.; Yu, H.-T.; Wang, H.-S.; Lai, C.-H.; Soong, Y.-K. Interleukin-18 System Messenger RNA and Protein Expression in Human Endometrium during the Menstrual Cycle. Fertil. Steril. 2006, 86, 905–913. [Google Scholar] [CrossRef]
- Lédée, N.; Petitbarat, M.; Rahmati, M.; Dubanchet, S.; Chaouat, G.; Sandra, O.; Perrier-d’Hauterive, S.; Munaut, C.; Foidart, J.M. New Pre-Conception Immune Biomarkers for Clinical Practice: Interleukin-18, Interleukin-15 and TWEAK on the Endometrial Side, G-CSF on the Follicular Side. J. Reprod. Immunol. 2011, 88, 118–123. [Google Scholar] [CrossRef]
- Lédée, N.; Chaouat, G.; Serazin, V.; Lombroso, R.; Dubanchet, S.; Oger, P.; Louafi, N.; Ville, Y. Endometrial Vascularity by Three-Dimensional Power Doppler Ultrasound and Cytokines: A Complementary Approach to Assess Uterine Receptivity. J. Reprod. Immunol. 2008, 77, 57–62. [Google Scholar] [CrossRef]
- Lédée-Bataille, N.; Bonnet-Chea, K.; Hosny, G.; Dubanchet, S.; Frydman, R.; Chaouat, G. Role of the Endometrial Tripod Interleukin-18, -15, and -12 in Inadequate Uterine Receptivity in Patients with a History of Repeated in Vitro Fertilization-Embryo Transfer Failure. Fertil. Steril. 2005, 83, 598–605. [Google Scholar] [CrossRef]
- Nardo, L.G. Vascular Endothelial Growth Factor Expression in the Endometrium during the Menstrual Cycle, Implantation Window and Early Pregnancy. Curr. Opin. Obstet. Gynecol. 2005, 17, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Petitbarat, M.; Rahmati, M.; Sérazin, V.; Dubanchet, S.; Morvan, C.; Wainer, R.; de Mazancourt, P.; Chaouat, G.; Foidart, J.-M.; Munaut, C.; et al. TWEAK Appears as a Modulator of Endometrial IL-18 Related Cytotoxic Activity of Uterine Natural Killers. PLoS ONE 2011, 6, e14497. [Google Scholar] [CrossRef] [PubMed]
- Petitbarat, M.; Serazin, V.; Dubanchet, S.; Wayner, R.; de Mazancourt, P.; Chaouat, G.; Lédée, N. Tumor Necrosis Factor-like Weak Inducer of Apoptosis (TWEAK)/Fibroblast Growth Factor Inducible-14 Might Regulate the Effects of Interleukin 18 and 15 in the Human Endometrium. Fertil. Steril. 2010, 94, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Coulam, C.B.; Roussev, R.G. Increasing Circulating T-Cell Activation Markers Are Linked to Subsequent Implantation Failure after Transfer of in Vitro Fertilized Embryos. Am. J. Reprod. Immunol. 2003, 50, 340–345. [Google Scholar] [CrossRef]
- Fukui, A.; Ntrivalas, E.; Fukuhara, R.; Fujii, S.; Mizunuma, H.; Gilman-Sachs, A.; Beaman, K.; Kwak-Kim, J. Correlation between Natural Cytotoxicity Receptors and Intracellular Cytokine Expression of Peripheral Blood NK Cells in Women with Recurrent Pregnancy Losses and Implantation Failures. Am. J. Reprod. Immunol. 2009, 62, 371–380. [Google Scholar] [CrossRef]
- Fukui, A.; Kwak-Kim, J.; Ntrivalas, E.; Gilman-Sachs, A.; Lee, S.-K.; Beaman, K. Intracellular Cytokine Expression of Peripheral Blood Natural Killer Cell Subsets in Women with Recurrent Spontaneous Abortions and Implantation Failures. Fertil. Steril. 2008, 89, 157–165. [Google Scholar] [CrossRef]
- Koushaeian, L.; Ghorbani, F.; Ahmadi, M.; Eghbal-Fard, S.; Zamani, M.; Danaii, S.; Yousefi, B.; Jadidi-Niaragh, F.; Hamdi, K.; Yousefi, M. The Role of IL-10-Producing B Cells in Repeated Implantation Failure Patients with Cellular Immune Abnormalities. Immunol. Lett. 2019, 214, 16–22. [Google Scholar] [CrossRef]
- Argilés, J.M.; Carbó, N.; López-Soriano, F.J. TNF and Pregnancy: The Paradigm of a Complex Interaction. Cytokine Growth Factor Rev. 1997, 8, 181–188. [Google Scholar] [CrossRef]
- Blois, S.M.; Freitag, N.; Tirado-González, I.; Cheng, S.-B.; Heimesaat, M.M.; Bereswill, S.; Rose, M.; Conrad, M.L.; Barrientos, G.; Sharma, S. NK Cell-Derived IL-10 Is Critical for DC-NK Cell Dialogue at the Maternal-Fetal Interface. Sci. Rep. 2017, 7, 2189. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Xu, H.; Zhang, D.; Si, C.; Zhou, X.; Zhao, H.; Liu, Q.; Xu, B.; Zhang, A. Decreased PIBF1/IL6/p-STAT3 during the Mid-Secretory Phase Inhibits Human Endometrial Stromal Cell Proliferation and Decidualization. J. Adv. Res. 2021, 30, 15–25. [Google Scholar] [CrossRef]
- Tu, B.; Du, L.; Fan, Q.-M.; Tang, Z.; Tang, T.-T. STAT3 Activation by IL-6 from Mesenchymal Stem Cells Promotes the Proliferation and Metastasis of Osteosarcoma. Cancer Lett. 2012, 325, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Halasz, M.; Polgar, B.; Berta, G.; Czimbalek, L.; Szekeres-Bartho, J. Progesterone-Induced Blocking Factor Differentially Regulates Trophoblast and Tumor Invasion by Altering Matrix Metalloproteinase Activity. Cell. Mol. Life Sci. 2013, 70, 4617–4630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskarin, G.; Tokmadzić, V.S.; Strbo, N.; Bogović, T.; Szekeres-Bartho, J.; Randić, L.; Podack, E.R.; Rukavina, D. Progesterone Induced Blocking Factor (PIBF) Mediates Progesterone Induced Suppression of Decidual Lymphocyte Cytotoxicity. Am. J. Reprod. Immunol. 2002, 48, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Turienzo, A.; Lledó, B.; Ortiz, J.A.; Morales, R.; Sanz, J.; Llácer, J.; Bernabeu, R. Prevalence of Candidate Single Nucleotide Polymorphisms on P53, IL-11, IL-10, VEGF and APOE in Patients with Repeated Implantation Failure (RIF) and Pregnancy Loss (RPL). Hum. Fertil. 2020, 23, 117–122. [Google Scholar] [CrossRef]
- Jung, Y.W.; Ahn, E.H.; Kim, J.O.; An, H.J.; Cho, S.H.; Kim, Y.R.; Lee, W.S.; Kim, N.K. Association of Genetic Polymorphisms in VEGF -460, -7 and -583 and Hematocrit Level with the Development of Idiopathic Recurrent Pregnancy Loss and a Meta-Analysis. J. Gene Med. 2018, 20, e3048. [Google Scholar] [CrossRef]
- Lledo, B.; Turienzo, A.; Ortiz, J.A.; Morales, R.; Ten, J.; Llácer, J.; Bernabeu, R. Negative Effect of P72 Polymorphism on P53 Gene in IVF Outcome in Patients with Repeated Implantation Failure and Pregnancy Loss. J. Assist. Reprod. Genet. 2014, 31, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, M.; Mao, B.; Cheng, X.; Zhang, X.; Xu, C. Association between Vascular Endothelial Growth Factor Polymorphism and Recurrent Pregnancy Loss: A Systematic Review and Meta-Analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 211, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Feng, Z.; Teresky, A.K.; Levine, A.J. P53 Regulates Maternal Reproduction through LIF. Nature 2007, 450, 721–724. [Google Scholar] [CrossRef]
- Magdoud, K.; Dendana, M.; Herbepin, V.; Hizem, S.; Ben Jazia, K.; Messaoudi, S.; Almawi, W.Y.; Touraine, R.; Mahjoub, T. Identification of Specific Vascular Endothelial Growth Factor Susceptible and Protective Haplotypes Associated with Recurrent Spontaneous Miscarriages. Hum. Reprod. 2012, 27, 1536–1541. [Google Scholar] [CrossRef] [Green Version]
- Abdolmohammadi-Vahid, S.; Pashazadeh, F.; Pourmoghaddam, Z.; Aghebati-Maleki, L.; Abdollahi-Fard, S.; Yousefi, M. The Effectiveness of IVIG Therapy in Pregnancy and Live Birth Rate of Women with Recurrent Implantation Failure (RIF): A Systematic Review and Meta-Analysis. J. Reprod. Immunol. 2019, 134–135, 28–33. [Google Scholar] [CrossRef]
- Martini, A.E.; Jasulaitis, S.; Fogg, L.F.; Uhler, M.L.; Hirshfeld-Cytron, J.E. Evaluating the Utility of Intralipid Infusion to Improve Live Birth Rates in Patients with Recurrent Pregnancy Loss or Recurrent Implantation Failure. J. Hum. Reprod. Sci. 2018, 11, 261–268. [Google Scholar] [CrossRef] [PubMed]
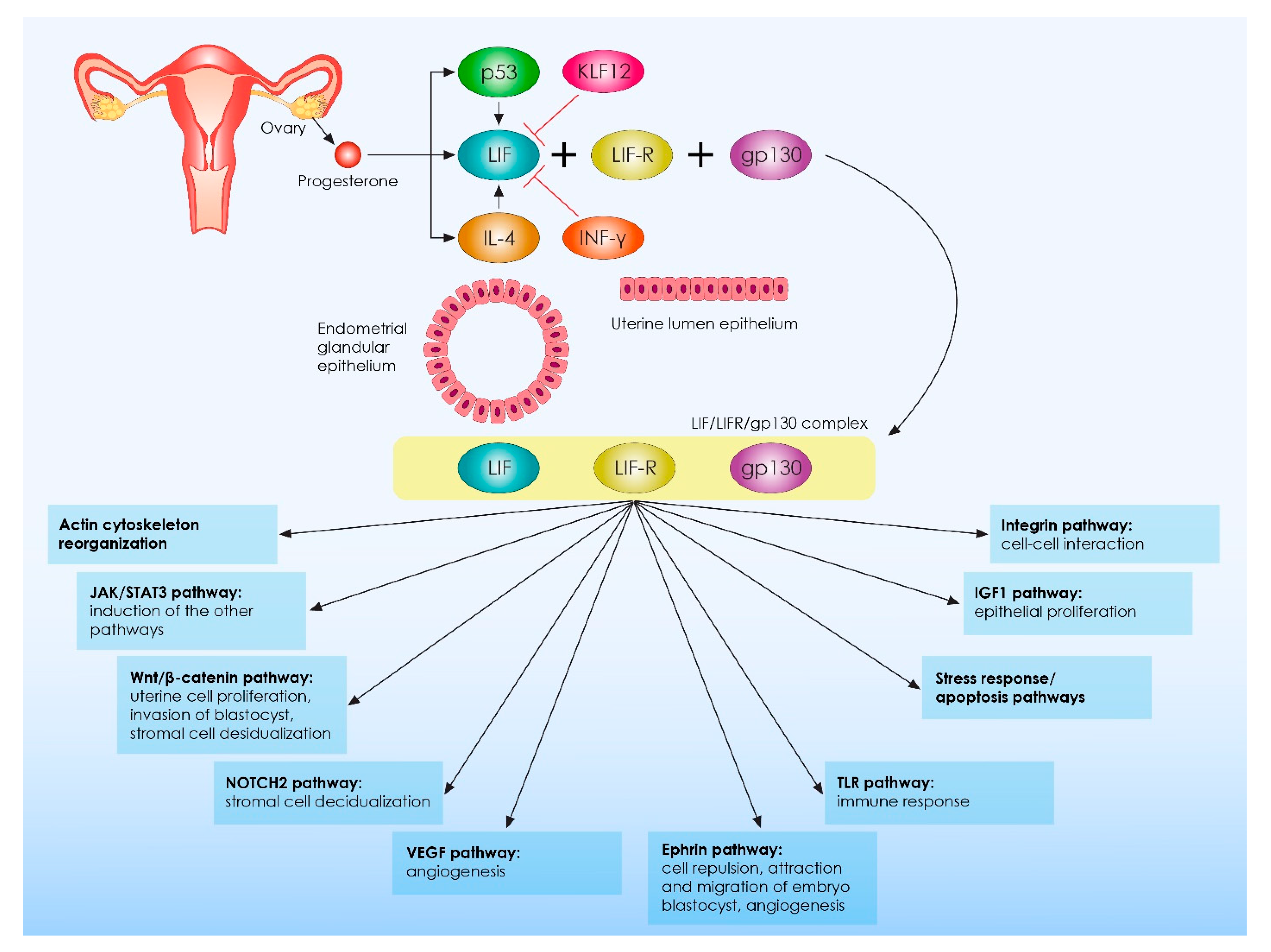



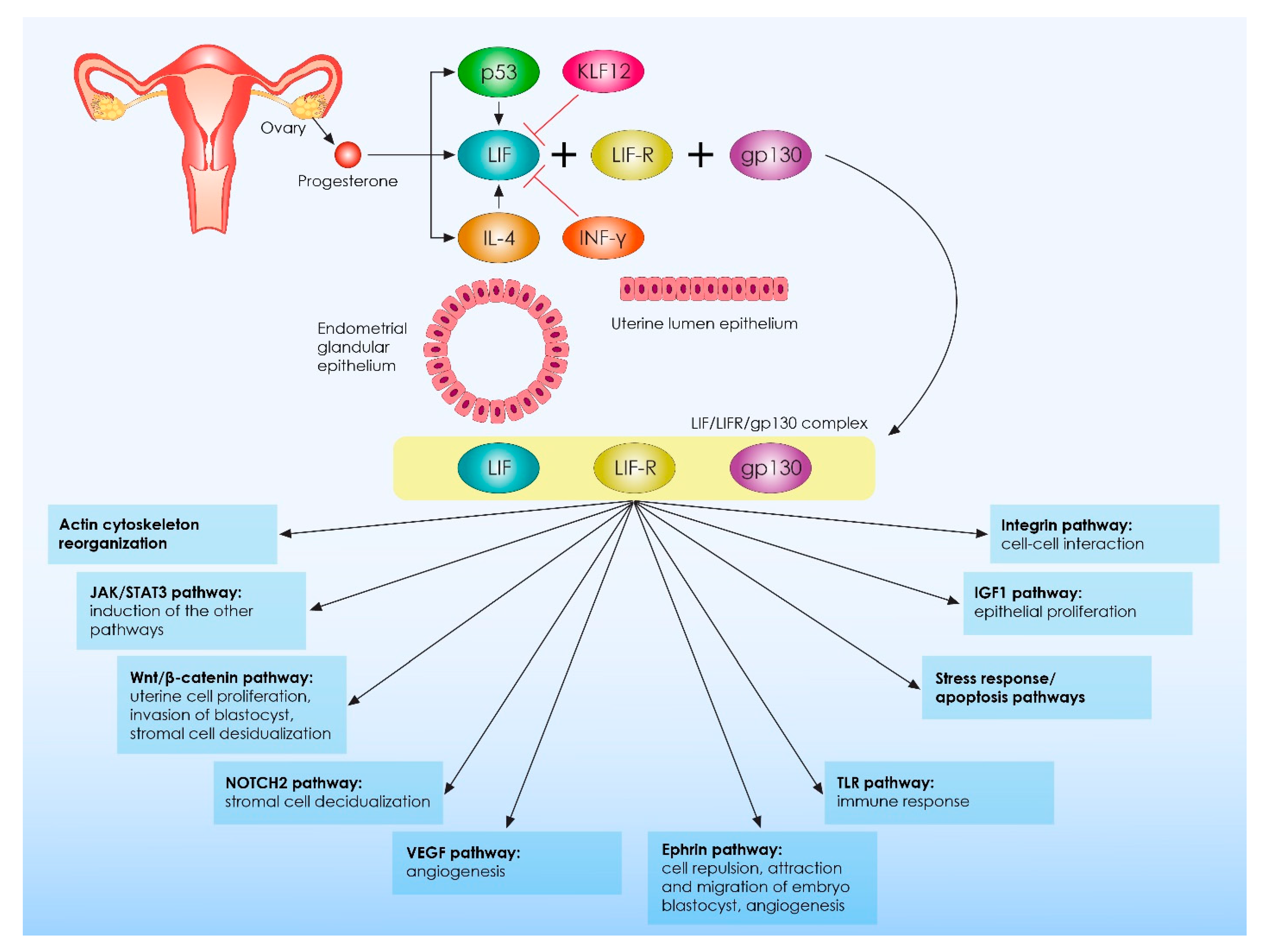{kind=link}
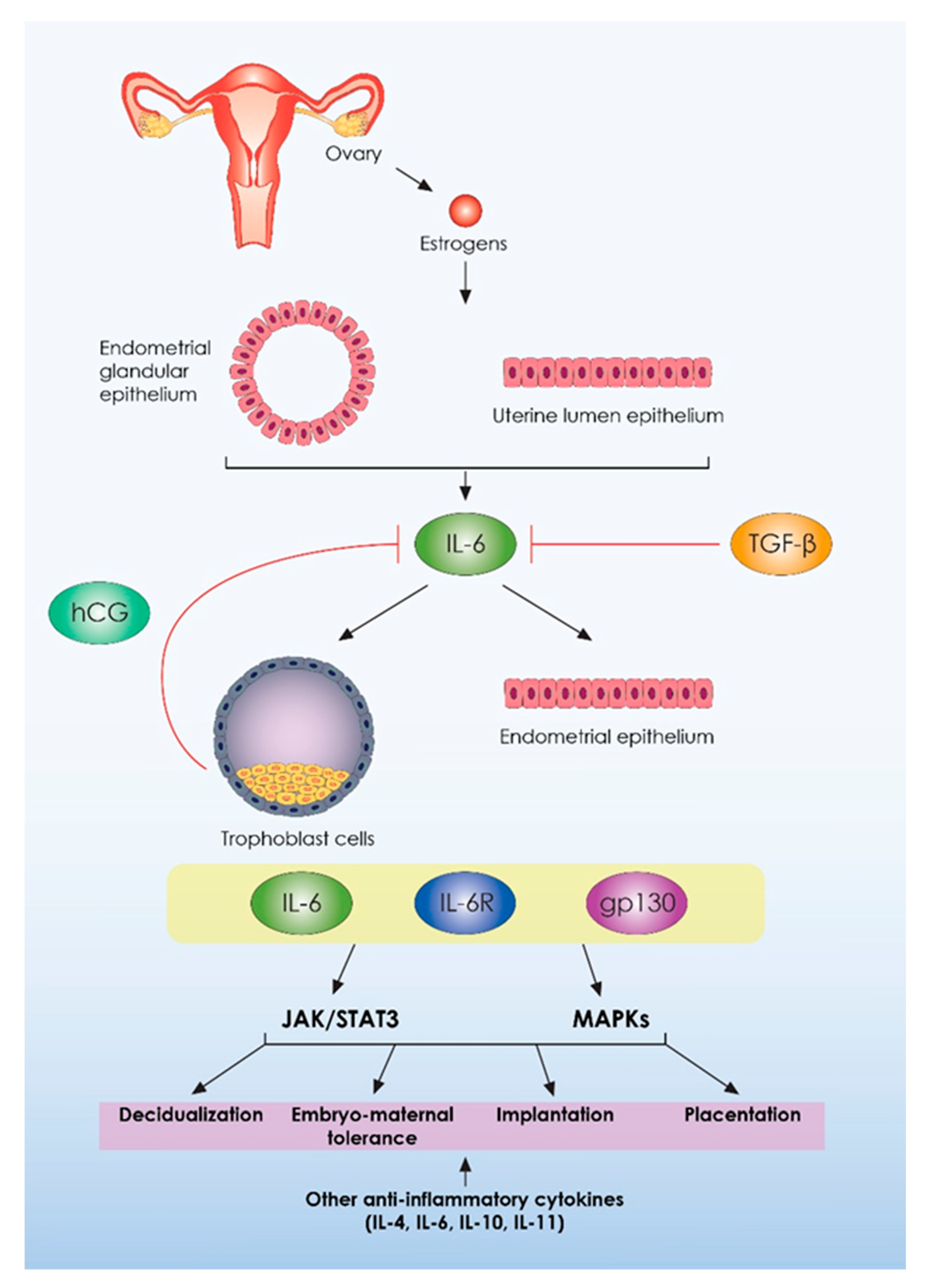
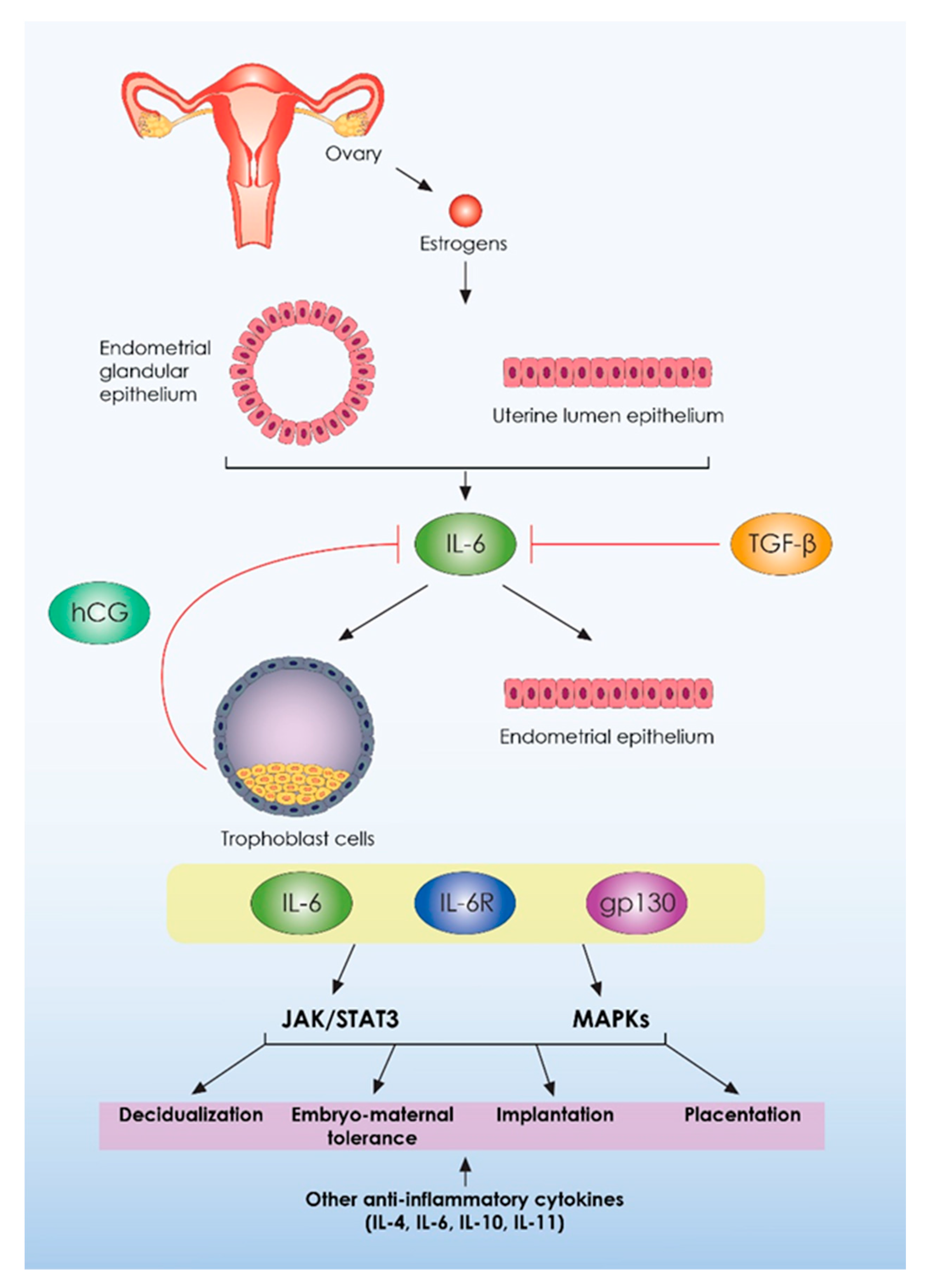{kind=link}
{kind=link}
{kind=link}
| Publication. | Study Design | Study Group(s) | Control Group | Outcome Measures | Conclusion |
|---|---|---|---|---|---|
| [134] | Retrospective case control study | -RIF patients -RPL patients | Oocyte donors with no history of RIF or RPL | Prevalence of polymorphisms in p53, VEGF, IL-10, IL-11 and APOE | Correlation of p53 and VEGF polymorphisms with RIF and APOE polymorphisms with RPL. |
| [20] | Prospective study | RIF patients | Fertile women | Expression of IL-6, IFNG, IL-17A, IL-23A, IFN-A1, IFN-B1, CD40L, CCR-4, CCR-5, CCR-6, CXR-3, CCL-2, IL-2, TLR-4, IRF-3, STAT3, RAG1, IFNA-R1, IL-1B, IL-8, NFKB, HLA-A, HLA-E, CD80, CD40 | Activation of pNK cells, Th17 signaling pathway and TLR signaling pathway in RIF patients; shift to inflammatory immune responses |
| [130] | Prospective study | RIF patients | Women with tubal obstruction or unexplained infertility who achieved a clinical pregnancy after the first embryo transfer | Evaluation of PIBF1, IL-6 and p-STAT3 in mid-secretory endometrium | Decreased PIBF1/IL6/p-STAT3 in RIF group. Decreased PIBF1/IL6/p-STAT3 during the mid-secretory phase inhibits human endometrial stromal cell proliferation and decidualization |
| [127] | Prospective study | RIF patients with immune system abnormalities | Women with at least one successful pregnancy | Evaluation of expression levels of PD-L1 and IL-10 in peripheral blood; serum levels of several autoantibodies including anti-TPO, anti-TG, ANA, ACA, APA | IL-10 producing B- cells are down-regulated in peripheral blood of patients with RIF |
| [69] | Prospective study in humans and in vivo experiments in mouse model | RIF patients | Age-matched fertile women | Evaluation of KLF12 and LIF expression patterns; evaluation of embryo adhesion in mouse model | KLF12 inhibits embryo adhesion in vivo and in vitro by repressing LIF expression. Increased expression of KLF12 and decreased expression of LIF in RIF patient endometrium |
| [19] | Observational cohort study | RIF patients | Women who underwent an endometrial sampling 3 months before their ETs and who all successfully gave birth at the first subsequent attempt of fresh or frozen-thawed ET | Ratio of IL-15/Fn-14 mRNA; biomarker of uNK cell activation/maturation and IL-18/TWEAK mRNA ratio; biomarker of both angiogenesis and the Th1/Th2 balance. | Endometrial immune profiles were dysregulated in RIF patients; mostly over activated |
| [104] | Prospective study | RIF patients | Women under 40 years old, with regular menstrual cycle, who had at least one normal pregnancy and delivery | Endometrial expression of p-STAT3), ERα and PR | Systematic dysregulation of LIF-JAK-STAT3 pathway in RIF patients |
| [109] | Retrospective study | RIF patients | Fertile women | Evaluation of IL-15 and LIF levels in peripheral blood and endometrial samples, association with uNK cell number | Altered expression of LIF and IL-15 in the endometrium of RIF patients. Correlation between the uNK cell number and the stromal cell IL-15. |
| [36] | Prospective study | RIF patients of advanced maternal age (age > 40 years old) | Women with proven fertility and under 40 years old, with at least one live birth and no history of abortion or infertility with regular menstrual cycles | Comparison of cytokine profile (IL-10, TGF-β, IFN-γ, IL-6, IL-8 and IL-17) of whole endometrial cells and endometrial stromal cells between the RIF and the control group | Higher levels of IL-6, IL-8 and TGF-β in the endometrial stromal cells and whole endometrial cells of normal fertile women compared to RIF group. Lower levels of IL-10 in endometrial stromal cells of the control group compared to RIF group. |
| [122] | Ex vivo model | Cell culture from endometrial biopsies from patients with RPL and RIF who had over expression of IL-18 with TWEAK | Cell culture from endometrial biopsies from women with RPL and women with RIF who had over expression of IL-18 without TWEAK | Ex vivo model to study mRNA expressions of NKp46 (uNK cytotoxic receptor) and TGF-β (regulates uNK cytokine production) | TWEAK is a modulator for prevention of endometrial uNK cytotoxicity |
| [123] | Prospective study | Unexplained RIF patients | Fertile women | Expression of TWEAK, IL-18, b2-microglobulin (b2 M), ribosomal protein L13A (RPL13A), and TATA box-binding protein (TBP) | High levels of IL-18/TWEAK ratio might lead to high levels of uNK cells but low levels of IL-15/Fn-14 might indicate uNK depletion |
| [116] | Prospective cohort study | RIF patients | Women undergoing IVF/ICSI treatment with successful implantation | Endometrial secretion analysis for 17 soluble regulators of implantation prior to embryo transfer | Clinical pregnancy was correlated with higher concentrations of TNF-α and lower levels of IL-1β. It is suggested that the ratio of TNF-α and IL-1β may appear as an indicator of endometrial receptivity. |
| [125] | Sequential study | RIF and RPL patients | Fertile women | Peripheral blood NK cells for NCRs (NKp46, NKp44 and NKp30) and cytokine expression (TNF-α, IFN-c, IL-4, IL-10) | Excessive pro-inflammatory cytokine expression in NK cells in RPL and RIF patients |
| [119] | Control study | RIF patients | Fertile women | Endometrial IL-18, IL-18BP, IL-15 mRNA expressions and number of CD56+ cells | Ultrasonographical indicators appear to be related to insufficient or excessive uNK recruitment and inadequate endothelial vascular remodeling. |
| [126] | Prospective cohort study | Patients with recurrent spontaneous abortions and RIF | Fertile women | Cytokine expression of interferon-γ, TNF-a, IL-4, IL-5, IL-10, IL-13, and granulocyte-macrophage colony-stimulating factor (GM-CSF) in NK cells | Natural killer-1 shift in peripheral blood samples of patients with recurrent spontaneous abortions and RIF |
| [120] | Pilot study | RIF patients | Fertile women | Uterine artery Doppler, count of uterine CD56 bright cells and quantification of IL-12 family, IL-18 system, and IL-15 mRNA | Uterine artery pulsatility index is negatively correlated with the IL-18/actin ratio. IL-18 and IL-15 are involved in the local recruitment and the activation of uNK cells. Correlation of IL-18 with IL-15 and IL-12. |
| [114] | Controlled clinical study | RIF patients | Fertile women | Evaluation of the balance between IL-12 and IL-18; evaluation of the number of uNK cells; evaluation of the endometrial vascular status | Abnormal vascular parameters among RIF group; higher number of uNK cells and altered IL-12 and IL-18 expression patterns. |
| [103] | Cohort study | RIF and unexplained infertility patients | Fertile women | Genetic analysis for LIF gene mutations | No functional mutations in LIF gene in women with RIF |
| [105] | Prospective study | RIF patients | -Multiparous women with a history of tubal sterilization who were undergoing surgical tubal anastomosis -Patients having three or more consecutive spontaneous abortions without a live birth | Matrix metalloproteinase (MMP) score and cytokine concentrations in endometrial fluids | In RIF group, the MMP score and IL-1β concentration were significantly higher than those in the control group, whereas concentrations of IFN-γ and IL-10 were significantly lower |
| [124] | Prospective study | Women undergoing in vitro fertilization treatment | Fertile women | Percentage of expression of CD69+, HLA-DR+ and CD11b+ on CD4 and CD8 T cells and association with implantation failure incidence | T-cell activation markers CD 69+ and HLA-DR+ are associated with increased implantation failure |
| [102] | Prospective blinded clinical and biochemical study | Women with primary or secondary infertility of unexplained etiology and RIF | Fertile women | Evaluation of LIF secretion in endometrial cultures | Decreased LIF production in endometrial samples obtained from RIF patients |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pantos, K.; Grigoriadis, S.; Maziotis, E.; Pistola, K.; Xystra, P.; Pantou, A.; Kokkali, G.; Pappas, A.; Lambropoulou, M.; Sfakianoudis, K.; et al. The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2022, 23, 2198. https://doi.org/10.3390/ijms23042198
Pantos K, Grigoriadis S, Maziotis E, Pistola K, Xystra P, Pantou A, Kokkali G, Pappas A, Lambropoulou M, Sfakianoudis K, et al. The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature. International Journal of Molecular Sciences. 2022; 23(4):2198. https://doi.org/10.3390/ijms23042198
Chicago/Turabian StylePantos, Konstantinos, Sokratis Grigoriadis, Evangelos Maziotis, Kalliopi Pistola, Paraskevi Xystra, Agni Pantou, Georgia Kokkali, Athanasios Pappas, Maria Lambropoulou, Konstantinos Sfakianoudis, and et al. 2022. "The Role of Interleukins in Recurrent Implantation Failure: A Comprehensive Review of the Literature" International Journal of Molecular Sciences 23, no. 4: 2198. https://doi.org/10.3390/ijms23042198