
Crosstalk between Melanin Concentrating Hormone and Endocrine Factors: Implications for Obesity

 , , and
, , and {kind=link}
Abstract
:1. Introduction
1.1. MCH: Precursor, Products, and Metabolic Actions
1.2. MCH Regulation
2. MCH Receptors
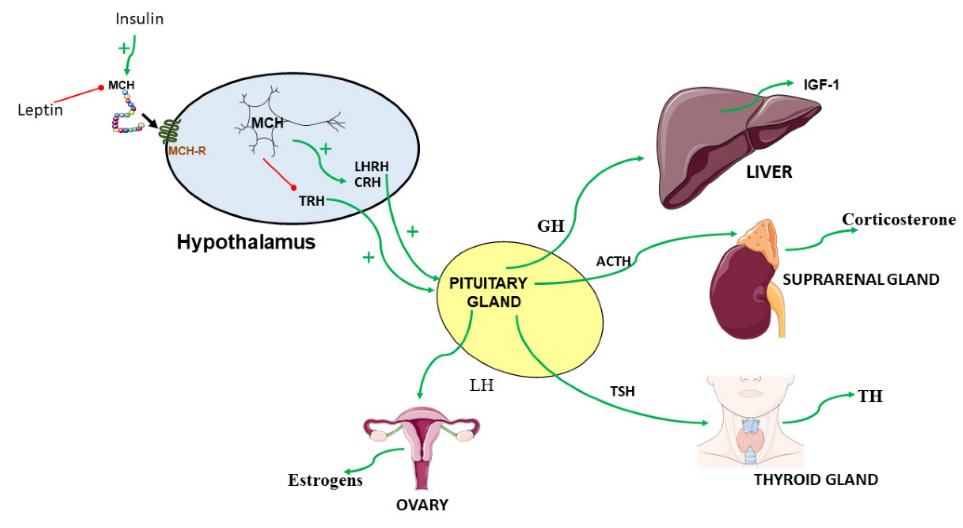
3. Crosstalk of MCH and Endocrine Factors
3.1. MCH and Insulin
3.2. MCH and Leptin
3.3. MCH and Somatotropic Function
3.4. MCH and Hypothalamic-Pituitary Gonadal Axis
Role of Estrogens
3.5. MCH Hypothalamo-Pituitary Thyroid Axis
3.6. MCH and Hypothalamic-Pituitary Adrenal Axis
4. Conclusions and Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kawauchi, H.; Kawazoe, I.; Tsubokawa, M.; Kishida, M.; Baker, B.I. Characterization of melanin-concentrating hormone in chum salmon pituitaries. Nature 1983, 305, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Pissios, P.; Bradley, R.L.; Maratos-Flier, E. Expanding the scales: The multiple roles of MCH in regulating energy balance and other biological functions. Endocr. Rev. 2006, 27, 606–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Massadi, O.; Dieguez, C.; Schneeberger, M.; Lopez, M.; Schwaninger, M.; Prevot, V.; Nogueiras, R. Multifaceted actions of melanin-concentrating hormone on mammalian energy homeostasis. Nat. Rev. Endocrinol. 2021, 17, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Nahon, J.L.; Presse, F.; Bittencourt, J.C.; Sawchenko, P.E.; Vale, W. The rat melanin-concentrating hormone messenger ribonucleic acid encodes multiple putative neuropeptides coexpressed in the dorsolateral hypothalamus. Endocrinology 1989, 125, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, J.C.; Presse, F.; Arias, C.; Peto, C.; Vaughan, J.; Nahon, J.L.; Vale, W.; Sawchenko, P.E. The melanin-concentrating hormone system of the rat brain: An immuno- and hybridization histochemical characterization. J. Comp. Neurol. 1992, 319, 218–245. [Google Scholar] [CrossRef]
- Bittencourt, J.C. Anatomical organization of the melanin-concentrating hormone peptide family in the mammalianbrain. Gen. Comp. Endocrinol. 2011, 172, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.M.; Fischer, W.H.; Hoeger, C.; Rivier, J.; Vale, W. Characterization of melanin-concentrating hormone from rat hypothalamus. Endocrinology 1989, 125, 1660–1665. [Google Scholar] [CrossRef]
- Matsunaga, T.O.; Hruby, V.J.; Lebl, M.; Castrucci, A.M.; Hadley, M.E. Synthesis and bioactivity studies of two isosteric acyclic analogues of melanin concentrating hormone. Life Sci. 1992, 51, 679–685. [Google Scholar] [CrossRef]
- Parkes, D.; Vale, W. Secretion of melanin-concentrating hormone and neuropeptide-EI from cultured rat hypothalamic cells. Endocrinology 1992, 131, 1826–1831. [Google Scholar] [CrossRef]
- Mul, J.D.; La Fleur, S.E.; Toonen, P.W.; Afrasiab-Middelman, A.; Binnekade, R.; Schetters, D.; Verheij, M.M.M.; Sears, R.M.; Homberg, J.R.; Schoffelmeer, A.N.M.; et al. Chronic loss of melanin-concentrating hormone affects motivational aspects of feeding in the rat. PLoS ONE 2011, 6, e19600. [Google Scholar] [CrossRef]
- Rossi, M.; Choi, S.J.; O’Shea, D.; Miyoshi, T.; Ghatei, M.A.; Bloom, S.R. Melanin-concentrating hormone acutely stimulates feeding, but chronic administration has no effect on body weight. Endocrinology 1997, 138, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.Y.; Bradley, R.L.; Kokkotou, E.G.; Marino, F.E.; Wang, X.; Pissios, P.; Maratos-Flier, E. MCH-/- mice are resistant to aging-associated increases in body weight and insulin resistance. Diabetes 2006, 55, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chee, M.J.; Pissios, P.; Maratos-Flier, E. Neurochemical characterization of neurons expressing melanin-concentrating hormone receptor 1 in the mouse hypothalamus. J. Comp. Neurol. 2013, 521, 2208–2234. [Google Scholar] [CrossRef] [Green Version]
- Izawa, S.; Chowdhury, S.; Miyazaki, T.; Mukai, Y.; Ono, D.; Inoue, R.; Ohmura, Y.; Mizoguchi, H.; Kimura, K.; Yoshioka, M.; et al. REM sleep-active MCH neurons are involved in forgetting hippocampus-dependent memories. Science 2019, 365, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, B.; Durkin, M.M.; Ogozalek, K.L.; Marzabadi, M.R.; Deleon, J.E.; Heurich, R.O.; Lichtblau, H.; Shaposhnik, Z.; Daniewska, I.; Blackburn, T.P.; et al. Antidepressant, anxiolytic and anorectic effects of a melanin-concentrating hormone-1 receptor antagonist. Nat. Med. 2002, 8, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Santollo, J.; Eckel, L.A. The orexigenic effect of melanin-concentrating hormone (MCH) is influenced by sex and stage of the estrous cycle. Physiol. Behav. 2008, 93, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, D.; Ludwig, D.; Gammeltoft, S.; Piper, M.E.; Pelleymounter, M.A.; Cullen, M.J.; Mathes, W.F.; Przypek, J.; Kanarek, R.B.; Maratos-Flier, E. A role for melanin-concentrating hormone in the central regulation of feeding behaviour. Nature 1996, 380, 243–247. [Google Scholar] [CrossRef]
- Gomori, A.; Ishihara, A.; Ito, M.; Mashiko, S.; Matsushita, H.; Yumoto, M.; Ito, M.; Tanaka, T.; Tokita, S.; Moriya, M.; et al. Chronic intracerebroventricular infusion of MCH causes obesity in mice. Melanin-concentrating hormone. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E583–E588. [Google Scholar] [CrossRef] [Green Version]
- Imbernon, M.; Beiroa, D.; Vázquez, M.J.; Morgan, D.A.; Veyrat–Durebex, C.; Porteiro, B.; Díaz–Arteaga, A.; Senra, A.; Busquets, S.; Velásquez, D.A.; et al. Central melanin-concentrating hormone influences liver and adipose metabolism via specific hypothalamic nuclei and efferent autonomic/JNK1 pathways. Gastroenterology 2013, 144, 636–649.e6. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, D.S.; Tritos, N.A.; Mastaitis, J.W.; Kulkarni, R.; Kokkotou, E.; Elmquist, J.; Lowell, B.; Flier, J.S.; Maratos-Flier, E. Melanin-concentrating hormone overexpression in transgenic mice leads to obesity and insulin resistance. J. Clin. Investig. 2001, 107, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Ishihara, A.; Gomori, A.; Egashira, S.; Matsushita, H.; Mashiko, S.; Ito, J.; Ito, M.; Nakase, K.; Haga, Y.; et al. Melanin-concentrating hormone 1-receptor antagonist suppresses body weight gain correlated with high receptor occupancy levels in diet-induced obesity mice. Eur. J. Pharm. 2009, 624, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Mashiko, S.; Ishihara, A.; Gomori, A.; Moriya, R.; Ito, M.; Iwaasa, H.; Matsuda, M.; Feng, Y.; Shen, Z.; Marsh, D.J.; et al. Antiobesity effect of a melanin-concentrating hormone 1 receptor antagonist in diet-induced obese mice. Endocrinology 2005, 146, 3080–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearman, L.P.; Camacho, R.E.; Stribling, D.S.; Zhou, D.; Bednarek, M.A.; Hreniuk, D.L.; Feighner, S.D.; Tan, C.P.; Howard, A.D.; Van der Ploeg, L.H.; et al. Chronic MCH-1 receptor modulation alters appetite, body weight and adiposity in rats. Eur. J. Pharm. 2003, 475, 37–47. [Google Scholar] [CrossRef]
- Shimada, M.; Tritos, N.A.; Lowell, B.B.; Flier, J.S.; Maratos-Flier, E. Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature 1998, 396, 670–674. [Google Scholar] [CrossRef]
- Alon, T.; Friedman, J.M. Late-onset leanness in mice with targeted ablation of melanin concentrating hormone neurons. J. Neurosci. 2006, 26, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ziogas, D.C.; Biddinger, S.; Kokkotou, E. You deserve what you eat: Lessons learned from the study of the melanin-concentrating hormone (MCH)-deficient mice. Gut 2010, 59, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiddon, B.B.; Palmiter, R.D. Ablation of neurons expressing melanin-concentrating hormone (MCH) in adult mice improves glucose tolerance independent of MCH signaling. J. Neurosci. 2013, 33, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D.J.; Weingarth, D.T.; Novi, D.E.; Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Guan, X.-M.; Jiang, M.M.; Feng, Y.; Camacho, R.E.; et al. Melanin-concentrating hormone 1 receptor-deficient mice are lean, hyperactive, and hyperphagic and have altered metabolism. Proc. Natl. Acad. Sci. USA 2002, 99, 3240–3245. [Google Scholar] [CrossRef] [Green Version]
- Szalai, K.K.; Beke, G.; Eles, J.; Kitka, T.; Kovacs, P.; Nagy, J.; Farkas, S.; Boros, A. Recent patents on novel MCH1 receptor antagonists as potential anti-obesity drugs. Recent. Pat. CNS Drug Discov. 2014, 9, 122–140. [Google Scholar] [CrossRef] [PubMed]
- Elias, C.F.; Lee, C.E.; Kelly, J.F.; Ahima, R.S.; Kuhar, M.; Saper, C.B.; Elmquist, J.K. Characterization of CART neurons in the rat and human hypothalamus. J. Comp. Neurol. 2001, 432, 442–459. [Google Scholar] [CrossRef]
- Fort, P.; Salvert, D.; Hanriot, L.; Jego, S.; Shimizu, H.; Hashimoto, K.; Mori, M.; Luppi, P.H. The satiety molecule nesfatin-1 is co-expressed with melanin concentrating hormone in tuberal hypothalamic neurons of the rat. Neuroscience 2008, 155, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Tan, K.; Nectow, A.R.; Parolari, L.; Caglar, C.; Azevedo, E.; Li, Z.; Domingos, A.; Friedman, J.M. Functional analysis reveals differential effects of glutamate and MCH neuropeptide in MCH neurons. Mol. Metab. 2018, 13, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Nieh, E.H.; Vander Weele, C.M.; Matthews, G.A.; Presbrey, K.N.; Wichmann, R.; Leppla, C.A.; Izadmehr, E.M.; Tye, K.M. Inhibitory Input from the Lateral Hypothalamus to the Ventral Tegmental Area Disinhibits Dopamine Neurons and Promotes Behavioral Activation. Neuron 2016, 90, 1286–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naufahu, J.; Alzaid, F.; Fiuza Brito, M.; Doslikova, B.; Valencia, T.; Cunliffe, A.; Murray, J.F. Melanin-concentrating hormone in peripheral circulation in the human. J. Endocrinol. 2017, 232, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Carnier, J.; de Piano, A.; Sanches, P.D.L.; Tock, L.; Nascimento, C.M.O.D.; Oyama, L.M.; Corrêa, F.A.; Ernandes, R.H.; Lederman, H.; de Mello, M.T.; et al. The role of orexigenic and anorexigenic factors in an interdisciplinary weight loss therapy for obese adolescents with symptoms of eating disorders. Int. J. Clin. Pract. 2010, 64, 784–790. [Google Scholar] [CrossRef]
- Mequinion, M.; Le Thuc, O.; Zgheib, S.; Alexandre, D.; Chartrel, N.; Rovere, C.; Hardouin, P.; Viltart, O.; Chauveau, C. Long-Term Energy Deficit in Mice Causes Long-Lasting Hypothalamic Alterations after Recovery. Neuroendocrinology 2017, 105, 372–383. [Google Scholar] [CrossRef]
- Hanada, R.; Nakazato, M.; Matsukura, S.; Murakami, N.; Yoshimatsu, H.; Sakata, T. Differential regulation of melanin-concentrating hormone and orexin genes in the agouti-related protein/melanocortin-4 receptor system. Biochem. Biophys. Res. Commun. 2000, 268, 88–91. [Google Scholar] [CrossRef]
- Stricker-Krongrad, A.; Dimitrov, T.; Beck, B. Central and peripheral dysregulation of melanin-concentrating hormone in obese Zucker rats. Brain Res. Mol. Brain Res. 2001, 92, 43–48. [Google Scholar] [CrossRef]
- Mizuno, T.M.; Kleopoulos, S.P.; Bergen, H.T.; Roberts, J.L.; Priest, C.A.; Mobbs, C.V. Hypothalamic pro-opiomelanocortin mRNA is reduced by fasting and [corrected] in ob/ob and db/db mice, but is stimulated by leptin. Diabetes 1998, 47, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Gerics, B.; Szalay, F.; Sotonyi, P.; Jancsik, V. Diurnal variation of the melanin-concentrating hormone level in the hypothalamus. Acta Biol. Hung. 2017, 68, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Plascencia, O.D.; Saderi, N.; Escobar, C.; Salgado-Delgado, R.C. Feeding during the rest phase promotes circadian conflict in nuclei that control energy homeostasis and sleep-wake cycle in rats. Eur. J. Neurosci. 2017, 45, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Dias Abdo Agamme, A.L.; Aguilar Calegare, B.F.; Fernandes, L.; Costa, A.; Lagos, P.; Torterolo, P.; D’Almeida, V. MCH levels in the CSF, brain preproMCH and MCHR1 gene expression during paradoxical sleep deprivation, sleep rebound and chronic sleep restriction. Peptides 2015, 74, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Pelluru, D.; Konadhode, R.; Shiromani, P.J. MCH neurons are the primary sleep-promoting group. Sleep 2013, 36, 1779–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stutz, A.M.; Staszkiewicz, J.; Ptitsyn, A.; Argyropoulos, G. Circadian expression of genes regulating food intake. Obesity 2007, 15, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harthoorn, L.F.; Sane, A.; Nethe, M.; Van Heerikhuize, J.J. Multi-transcriptional profiling of melanin-concentrating hormone and orexin-containing neurons. Cell Mol. Neurobiol. 2005, 25, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nothacker, H.P.; Wang, Z.; Lin, S.H.; Leslie, F.; Civelli, O. Molecular characterization of the melanin-concentrating-hormone receptor. Nature 1999, 400, 265–269. [Google Scholar] [CrossRef]
- Chambers, J.; Ames, R.S.; Bergsma, D.; Muir, A.I.; Fitzgerald, L.R.; Hervieu, G.; Dytko, G.M.; Foley, J.J.; Martin, J.F.; Liu, W.-S.; et al. Melanin-concentrating hormone is the cognate ligand for the orphan G-protein-coupled receptor SLC-1. Nature 1999, 400, 261–265. [Google Scholar] [CrossRef]
- Lembo, P.M.; Grazzini, E.; Cao, J.; Hubatsch, D.A.; Pelletier, M.; Hoffert, C.; St-Onge, S.; Pou, C.; Labrecque, J.; Groblewski, T.; et al. The receptor for the orexigenic peptide melanin-concentrating hormone is a G-protein-coupled receptor. Nat. Cell Biol. 1999, 1, 267–271. [Google Scholar] [CrossRef]
- Kolakowski, L.F., Jr.; Jung, B.P.; Nguyen, T.; Johnson, M.P.; Lynch, K.R.; Cheng, R.; Heng, H.H.; George, S.R.; O’Dowd, B.F. Characterization of a human gene related to genes encoding somatostatin receptors. FEBS Lett. 1996, 398, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Hawes, B.E.; Kil, E.; Green, B.; O’Neill, K.; Fried, S.; Graziano, M.P. The melanin-concentrating hormone receptor couples to multiple G proteins to activate diverse intracellular signaling pathways. Endocrinology 2000, 141, 4524–4532. [Google Scholar] [CrossRef]
- Hervieu, G.J.; Cluderay, J.E.; Harrison, D.; Meakin, J.; Maycox, P.; Nasir, S.; Leslie, R.A. The distribution of the mRNA and protein products of the melanin-concentrating hormone (MCH) receptor gene, slc-1, in the central nervous system of the rat. Eur. J. Neurosci. 2000, 12, 1194–1216. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Cheng, M.; Leslie, F.M.; Civelli, O. Expression of the melanin-concentrating hormone (MCH) receptor mRNA in the rat brain. J. Comp. Neurol. 2001, 435, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.; Duckworth, M.; Murdock, P.; Rennie, G.; Sabido-David, C.; Ames, R.S.; Szekeres, P.; Wilson, S.; Bergsma, D.J.; Gloger, I.S.; et al. Molecular cloning and functional characterization of MCH2, a novel human MCH receptor. J. Biol. Chem. 2001, 276, 20125–20129. [Google Scholar] [CrossRef] [Green Version]
- Sailer, A.W.; Sano, H.; Zeng, Z.; McDonald, T.P.; Pan, J.; Pong, S.-S.; Feighner, S.D.; Tan, C.P.; Fukami, T.; Iwaasa, H.; et al. Identification and characterization of a second melanin-concentrating hormone receptor, MCH-2R. Proc. Natl. Acad. Sci. USA 2001, 98, 7564–7569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlumberger, S.E.; Talke-Messerer, C.; Zumsteg, U.; Eberle, A.N. Expression of receptors for melanin-concentrating hormone (MCH) in different tissues and cell lines. J. Recept. Signal Transduct. Res. 2002, 22, 509–531. [Google Scholar] [CrossRef]
- Fried, S.; O’Neill, K.; Hawes, B.E. Cloning and characterization of rhesus monkey MCH-R1 and MCH-R2. Peptides 2002, 23, 1401–1408. [Google Scholar] [CrossRef]
- Tan, C.P.; Sanob, H.; Iwaasab, H.; Pana, J.; Sailer, A.W.; Hreniuk, D.L.; Feighner, S.D.; Palyha, O.C.; Ponga, S.-S.; Figueroa, D.J.; et al. Melanin-concentrating hormone receptor subtypes 1 and 2: Species-specific gene expression. Genomics 2002, 79, 785–792. [Google Scholar] [CrossRef]
- Al Massadi, O.; Lopez, M.; Tschop, M.; Dieguez, C.; Nogueiras, R. Current Understanding of the Hypothalamic Ghrelin Pathways Inducing Appetite and Adiposity. Trends Neurosci. 2017, 40, 167–180. [Google Scholar] [CrossRef]
- Belgardt, B.F.; Bruning, J.C. CNS leptin and insulin action in the control of energy homeostasis. Ann. N. Y. Acad. Sci. 2010, 1212, 97–113. [Google Scholar] [CrossRef]
- Waterson, M.J.; Horvath, T.L. Neuronal Regulation of Energy Homeostasis: Beyond the Hypothalamus and Feeding. Cell Metab. 2015, 22, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmet, P.; Alberti, G.; Kaufman, F.; Tajima, N.; Silink, M.; Arslanian, S.; Wong, G.; Bennett, P.; Shaw, J.; Caprio, S.; et al. International Diabetes Federation Task Force on Epidemiology and Prevention of Diabetes. The metabolic syndrome in children and adolescents. Lancet 2007, 369, 2059–2061. [Google Scholar] [CrossRef]
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef] [PubMed]
- NCD Risk Factor Collaboration. Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [Green Version]
- Olshansky, S.J.; Passaro, D.J.; Hershow, R.C.; Layden, J.; Carnes, B.A.; Brody, J.; Hayflick, L.; Butler, R.N.; Allison, D.; Ludwig, D. A Potential Decline in Life Expectancy in the United States in the 21st Century. N. Engl. J. Med. 2005, 352, 1138–1145. [Google Scholar] [CrossRef] [Green Version]
- Bahjaoui-Bouhaddi, M.; Fellmann, D.; Griffond, B.; Bugnon, C. Insulin treatment stimulates the rat melanin-concentrating hormone-producing neurons. Neuropeptides 1994, 27, 251–258. [Google Scholar] [CrossRef]
- Hausen, A.C.; Ruud, J.; Jiang, H.; Hess, S.; Varbanov, H.; Kloppenburg, P.; Bruning, J.C. Insulin-Dependent Activation of MCH Neurons Impairs Locomotor Activity and Insulin Sensitivity in Obesity. Cell Rep. 2016, 17, 2512–2521. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Viale, A.; Picard, F.; Nahon, J.; Richard, D. Effects of leptin on melanin-concentrating hormone expression in the brain of lean and obese Lep(ob)/Lep(ob) mice. Neuroendocrinology 1999, 69, 145–153. [Google Scholar] [CrossRef]
- Gavrila, A.; Chan, J.L.; Miller, L.C.; Heist, K.; Yiannakouris, N.; Mantzoros, C.S. Circulating melanin-concentrating hormone, agouti-related protein, and alpha-melanocyte-stimulating hormone levels in relation to body composition: Alterations in response to food deprivation and recombinant human leptin administration. J. Clin. Endocrinol. Metab. 2005, 90, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Bjursell, M.; Gerdin, A.K.; Ploj, K.; Svensson, D.; Svensson, L.; Oscarsson, J.; Snaith, M.; Tornell, J.; Bohlooly, Y.M. Melanin-concentrating hormone receptor 1 deficiency increases insulin sensitivity in obese leptin-deficient mice without affecting body weight. Diabetes 2006, 55, 725–733. [Google Scholar] [CrossRef] [Green Version]
- Segal-Lieberman, G.; Bradley, R.L.; Kokkotou, E.; Carlson, M.; Trombly, D.J.; Wang, X.; Bates, S.; Myers, M.G.; Flier, J.S.; Maratos-Flier, E. Melanin-concentrating hormone is a critical mediator of the leptin-deficient phenotype. Proc. Natl. Acad. Sci. USA 2003, 100, 10085–10090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Whiddon, B.B.; Palmiter, R.D. Ablation of neurons expressing agouti-related protein, but not melanin concentrating hormone, in leptin-deficient mice restores metabolic functions and fertility. Proc. Natl. Acad. Sci. USA 2012, 109, 3155–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egecioglu, E.; Stenstrom, B.; Pinnock, S.B.; Tung, L.Y.; De La Cour, C.D.; Lindqvist, A.; Håkanson, R.; Syversen, U.; Chen, D.; Dickson, S.L. Hypothalamic gene expression following ghrelin therapy to gastrectomized rodents. Regul. Pept. 2008, 146, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Egecioglu, E.; Gerdin, A.K.; Svensson, L.; Oscarsson, J.; Morgan, D.; Snaith, M.; Tornell, J.; Bohlooly, Y.M. Importance of melanin-concentrating hormone receptor for the acute effects of ghrelin. Biochem. Biophys. Res. Commun. 2005, 326, 759–765. [Google Scholar] [CrossRef]
- Segal-Lieberman, G.; Rubinfeld, H.; Glick, M.; Kronfeld-Schor, N.; Shimon, I. Melanin-concentrating hormone stimulates human growth hormone secretion: A novel effect of MCH on the hypothalamic-pituitary axis. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E982–E988. [Google Scholar] [CrossRef]
- Takahashi, K.; Totsune, K.; Murakami, O.; Sone, M.; Satoh, F.; Kitamuro, T.; Noshiro, T.; Hayashi, Y.; Sasano, H.; Shibahara, S. Expression of melanin-concentrating hormone receptor messenger ribonucleic acid in tumor tissues of pheochromocytoma, ganglioneuroblastoma, and neuroblastoma. J. Clin. Endocrinol. Metab. 2001, 86, 369–374. [Google Scholar] [CrossRef]
- Cvetkovic, V.; Brischoux, F.; Griffond, B.; Bernard, G.; Jacquemard, C.; Fellmann, D.; Risold, P.Y. Evidence of melanin-concentrating hormone-containing neurons supplying both cortical and neuroendocrine projections. Neuroscience 2003, 116, 31–35. [Google Scholar] [CrossRef]
- Glick, M.; Segal-Lieberman, G.; Cohen, R.; Kronfeld-Schor, N. Chronic MCH infusion causes a decrease in energy expenditure and body temperature, and an increase in serum IGF-1 levels in mice. Endocrine 2009, 36, 479–485. [Google Scholar] [CrossRef]
- Wade, G.N.; Jones, J.E. Neuroendocrinology of nutritional infertility. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1277–R1296. [Google Scholar] [CrossRef] [Green Version]
- Williamson-Hughes, P.S.; Grove, K.L.; Smith, M.S. Melanin concentrating hormone (MCH): A novel neural pathway for regulation of GnRH neurons. Brain Res. 2005, 1041, 117–124. [Google Scholar] [CrossRef]
- Chiocchio, S.R.; Gallardo, M.G.; Louzan, P.; Gutnisky, V.; Tramezzani, J.H. Melanin-concentrating hormone stimulates the release of luteinizing hormone-releasing hormone and gonadotropins in the female rat acting at both median eminence and pituitary levels. Biol. Reprod. 2001, 64, 1466–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.I.; Baker, B.I.; Wilson, C.A. Stimulatory effect of melanin-concentrating hormone on luteinising hormone release. Neuroendocrinology 1997, 66, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.F.; Mercer, J.G.; Adan, R.A.; Datta, J.J.; Aldairy, C.; Moar, K.M.; Baker, B.I.; Stock, M.J.; Wilson, C.A. The effect of leptin on luteinizing hormone release is exerted in the zona incerta and mediated by melanin-concentrating hormone. J. Neuroendocr. 2000, 12, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.J.; Romero-Ruiz, A.; Tena-Sempere, M. Roles of leptin in reproduction, pregnancy and polycystic ovary syndrome: Consensus knowledge and recent developments. Metabolism 2015, 64, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Dragano, N.; Milbank, E.; Lopez, M. Estradiol and appetite: To eat or not to eat. Mol. Metab. 2020, 42, 101061. [Google Scholar] [CrossRef]
- Gonzalez-Garcia, I.G.; de Morentin, P.B.M.; Estévez-Salguero, Á.; Contreras, C.; Romero-Picó, A.; Fernø, J.; Nogueiras, R.; Diéguez, C.; Tena-Sempere, M.; Tovar, S.; et al. mTOR signaling in the arcuate nucleus of the hypothalamus mediates the anorectic action of estradiol. J. Endocrinol. 2018, 238, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Messina, M.M.; Boersma, G.; Overton, J.M.; Eckel, L.A. Estradiol decreases the orexigenic effect of melanin-concentrating hormone in ovariectomized rats. Physiol. Behav. 2006, 88, 523–528. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, C.; Hsu, C.-K.; Zhang, Q.; Bi, C.; Asnicar, M.; Hsiung, H.M.; Fox, N.; Slieker, L.J.; Yang, D.D.; et al. Targeted disruption of the melanin-concentrating hormone receptor-1 results in hyperphagia and resistance to diet-induced obesity. Endocrinology 2002, 143, 2469–2477. [Google Scholar] [CrossRef]
- Lopez, M.; Varela, L.; Vázquez, M.J.; Rodríguez-Cuenca, S.; González, C.R.; Velagapudi, V.R.; Morgan, D.A.; Schoenmakers, E.; Agassandian, K.; Lage, R.; et al. Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance. Nat. Med. 2010, 16, 1001–1008. [Google Scholar] [CrossRef]
- Capelli, V.; Dieguez, C.; Mittag, J.; Lopez, M. Thyroid wars: The rise of central actions. Trends Endocrinol. Metab. 2021, 32, 659–671. [Google Scholar] [CrossRef]
- Harris, A.R.; Fang, S.L.; Azizi, F.; Lipworth, L.; Vagenakis, A.G.; Barverman, L.E. Effect of starvation on hypothalamic-pituitary-thyroid function in the rat. Metabolism 1978, 27, 1074–1083. [Google Scholar] [CrossRef]
- Chung, S.; Liao, X.H.; Di Cosmo, C.; Van Sande, J.; Wang, Z.; Refetoff, S.; Civelli, O. Disruption of the melanin-concentrating hormone receptor 1 (MCH1R) affects thyroid function. Endocrinology 2012, 153, 6145–6154. [Google Scholar] [CrossRef]
- Kennedy, A.R.; Todd, J.F.; Stanley, S.A.; Abbott, C.R.; Small, C.J.; Ghatei, M.A.; Bloom, S.R. Melanin-concentrating hormone (MCH) suppresses thyroid stimulating hormone (TSH) release, in vivo and in vitro, via the hypothalamus and the pituitary. Endocrinology 2001, 142, 3265–3268. [Google Scholar] [CrossRef]
- Tsigos, C.; Chrousos, G.P. Hypothalamic-pituitary-adrenal axis, neuroendocrine factors and stress. J. Psychosom. Res. 2002, 53, 865–871. [Google Scholar] [CrossRef] [Green Version]
- Jezova, D.; Bartanusz, V.; Westergren, I.; Johansson, B.B.; Rivier, J.; Vale, W.; Rivier, C. Rat melanin-concentrating hormone stimulates adrenocorticotropin secretion: Evidence for a site of action in brain regions protected by the blood-brain barrier. Endocrinology 1992, 130, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Todd, J.F.; Dhillo, W.S.; Seal, L.J.; Ghatei, M.A.; O’Toole, C.P.; Jones, M.; Witty, D.; Winborne, K.; Riley, G.; et al. Effect of direct injection of melanin-concentrating hormone into the paraventricular nucleus: Further evidence for a stimulatory role in the adrenal axis via SLC-1. J. Neuroendocr. 2003, 15, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.G.; Davis, R.J.; Rorick-Kehn, L.; Morin, M.; Witkin, J.M.; McKinzie, D.L.; Nomikos, G.G.; Gehlert, D.R. Melanin-concentrating hormone-1 receptor modulates neuroendocrine, behavioral, and corticolimbic neurochemical stress responses in mice. Neuropsychopharmacology 2006, 31, 1135–1145. [Google Scholar] [CrossRef]
- Georgescu, D.; Sears, R.M.; Hommel, J.D.; Barrot, M.; Bolaños, C.A.; Marsh, D.J.; Bednarek, M.A.; Bibb, J.A.; Maratos-Flier, E.; Nestler, E.J.; et al. The hypothalamic neuropeptide melanin-concentrating hormone acts in the nucleus accumbens to modulate feeding behavior and forced-swim performance. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 2933–2940. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prida, E.; Fernández-González, S.; Pena-León, V.; Pérez-Lois, R.; Fernø, J.; Seoane, L.M.; Quiñones, M.; Al Massadi, O. Crosstalk between Melanin Concentrating Hormone and Endocrine Factors: Implications for Obesity. Int. J. Mol. Sci. 2022, 23, 2436. https://doi.org/10.3390/ijms23052436
Prida E, Fernández-González S, Pena-León V, Pérez-Lois R, Fernø J, Seoane LM, Quiñones M, Al Massadi O. Crosstalk between Melanin Concentrating Hormone and Endocrine Factors: Implications for Obesity. International Journal of Molecular Sciences. 2022; 23(5):2436. https://doi.org/10.3390/ijms23052436
Chicago/Turabian StylePrida, Eva, Sara Fernández-González, Verónica Pena-León, Raquel Pérez-Lois, Johan Fernø, Luisa María Seoane, Mar Quiñones, and Omar Al Massadi. 2022. "Crosstalk between Melanin Concentrating Hormone and Endocrine Factors: Implications for Obesity" International Journal of Molecular Sciences 23, no. 5: 2436. https://doi.org/10.3390/ijms23052436