
Apolipoprotein E, a Crucial Cellular Protein in the Lifecycle of Hepatitis Viruses


{kind=link}
Abstract
:1. Apolipoprotein E
2. Hepatitis C Virus
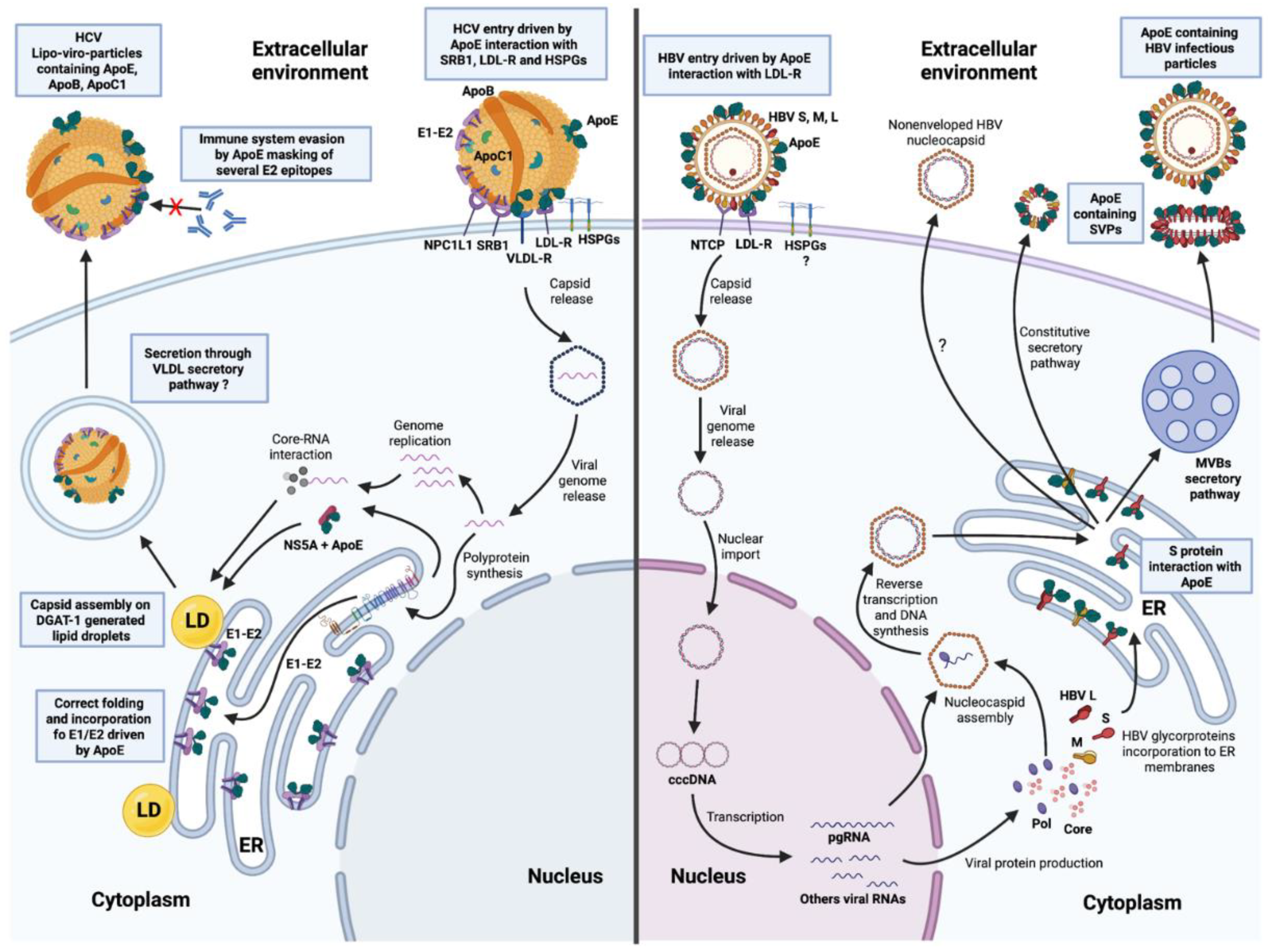
2.1. ApoE on HCV Particles
2.2. ApoE and the Intracellular Lifecycle of HCV
3. Hepatitis B Virus
3.1. Disorders of Global Lipid Metabolism during HBV Infection
3.2. Presence of ApoE on HBV Particles and Its Role in the Viral Entry Process
3.3. ApoE and the Intracellular Lifecycle of HBV
4. Other Hepatitis Viruses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Liu, J.-Q.; Li, W.-X.; Zheng, J.-J.; Tian, Q.-N.; Huang, J.-F.; Dai, S.-X. Gain and loss events in the evolution of the apolipoprotein family in vertebrata. BMC Evol. Biol. 2019, 19, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Mägi, R.; Milani, L.; Lauschke, V.M. Global genetic diversity of human apolipoproteins and effects on cardiovascular disease risk. J. Lipid Res. 2018, 59, 1987–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basavaraju, P.; Balasubramani, R.; Kathiresan, D.S.; Devaraj, I.; Babu, K.; Alagarsamy, V.; Puthamohan, V.M. Genetic Regulatory Networks of Apolipoproteins and Associated Medical Risks. Front. Cardiovasc. Med. 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Halim, A.; Rüetschi, U.; Larson, G.; Nilsson, J. LC-MS/MS Characterization of O-Glycosylation Sites and Glycan Structures of Human Cerebrospinal Fluid Glycoproteins. J. Proteome Res. 2013, 12, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kockx, M.; Raftery, M.J.; Jessup, W.; Griffith, R.; Kritharides, L. Glycosylation and Sialylation of Macrophage-derived Human Apolipoprotein E Analyzed by SDS-PAGE and Mass Spectrometry. Mol. Cell. Proteom. 2010, 9, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Rüetschi, U.; Halim, A.; Hesse, C.; Carlsohn, E.; Brinkmalm, G.; Larson, G. Enrichment of glycopeptides for glycan structure and attachment site identification. Nat. Methods 2009, 6, 809–811. [Google Scholar] [CrossRef]
- Rall, S.C.; Weisgraber, K.H.; Mahley, R.W. Human Apolipoprotein, E. The Complete Amino Acid Sequence. J. Biol. Chem. 1982, 257, 4171–4178. [Google Scholar] [CrossRef]
- Wernette-Hammond, M.E.; Lauer, S.J.; Corsini, A.; Walker, D.; Taylor, J.M.; Rall, S.C. Glycosylation of human apolipoprotein E. The carbohydrate attachment site is threonine 194. J. Biol. Chem. 1989, 264, 9094–9101. [Google Scholar] [CrossRef]
- Huang, Y.; Mahley, R.W. Apolipoprotein E: Structure and function in lipid metabolism, neurobiology, and Alzheimer’s diseases. Neurobiol. Dis. 2014, 72, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Rall, S.C. Apolipoprotein E: Far More Than a Lipid Transport Protein. Annu. Rev. Genom. Hum. Genet. 2000, 1, 507–537. [Google Scholar] [CrossRef]
- Khalil, Y.A.; Rabès, J.-P.; Boileau, C.; Varret, M. APOE gene variants in primary dyslipidemia. Atherosclerosis 2021, 328, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ghebranious, N. Detection of ApoE E2, E3 and E4 alleles using MALDI-TOF mass spectrometry and the homogeneous mass-extend technology. Nucleic Acids Res. 2005, 33, e149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatters, D.M.; Peters-Libeu, C.A.; Weisgraber, K.H. Apolipoprotein E structure: Insights into function. Trends Biochem. Sci. 2006, 31, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Blue, M.L.; Williams, D.L.; Zucker, S.; Khan, S.A.; Blum, C.B. Apolipoprotein E synthesis in human kidney, adrenal gland, and liver. Proc. Natl. Acad. Sci. USA 1983, 80, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Elshourbagy, N.A.; Liao, W.S.; Mahley, R.W.; Taylor, J.M. Apolipoprotein E mRNA is abundant in the brain and adrenals, as well as in the liver, and is present in other peripheral tissues of rats and marmosets. Proc. Natl. Acad. Sci. USA 1985, 82, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Weisgraber, K.H.; Huang, Y. Apolipoprotein E: Structure determines function, from atherosclerosis to Alzheimer’s disease to AIDS. J. Lipid Res. 2009, 50, S183–S188. [Google Scholar] [CrossRef] [Green Version]
- Getz, G.S.; Reardon, C.A. Apoprotein E as a lipid transport and signaling protein in the blood, liver, and artery wall. J. Lipid Res. 2009, 50, S156–S161. [Google Scholar] [CrossRef] [Green Version]
- Dose, J.; Huebbe, P.; Nebel, A.; Rimbach, G. APOE genotype and stress response - a mini review. Lipids Health Dis. 2016, 15, 121. [Google Scholar] [CrossRef] [Green Version]
- van Niel, G.; Bergam, P.; Di Cicco, A.; Hurbain, I.; Cicero, A.L.; Dingli, F.; Palmulli, R.; Fort, C.; Potier, M.-C.; Schurgers, L.; et al. Apolipoprotein E Regulates Amyloid Formation within Endosomes of Pigment Cells. Cell Rep. 2015, 13, 43–51. [Google Scholar] [CrossRef]
- Feinstone, S.M.; Kapikian, A.Z.; Purcell, R.H.; Alter, H.J.; Holland, P.V. Transfusion-Associated Hepatitis Not Due to Viral Hepatitis Type A or B. N. Engl. J. Med. 1975, 292, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Houghton, M. Discovery of the hepatitis C virus. Liver Int. 2009, 29, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Choo, Q.-L.; Kuo, G.; Weiner, A.J.; Overby, L.R.; Bradley, D.W.; Houghton, M. Isolation of a cDNA cLone Derived from a Blood-Borne Non-A, Non-B Viral Hepatitis Genome. Science 1989, 244, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pol, S.; Lagaye, S. The remarkable history of the hepatitis C virus. Genes Immun. 2019, 20, 436–446. [Google Scholar] [CrossRef] [PubMed]
- M.D., S.J.; Gordon, S.C. Epidemiology of Hepatitis, C. Clin. Liver Dis. 2018, 12, 140–142. [Google Scholar] [CrossRef] [Green Version]
- Zoulim, F.; Liang, T.J.; Gerbes, A.L.; Aghemo, A.; Deuffic-Burban, S.; Dusheiko, G.M.; Fried, M.W.; Pol, S.; Rockstroh, J.K.; Terrault, N.A.; et al. Hepatitis C virus treatment in the real world: Optimising treatment and access to therapies: Table 1. Gut 2015, 64, 1824–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geddawy, A.; Ibrahim, Y.F.; Elbahie, N.M.; Ibrahim, M.A. Direct acting anti-hepatitis C virus drugs: Clinical pharmacology and future direction. J. Transl. Intern. Med. 2017, 5, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Roingeard, P.; Beaumont, E. Hepatitis C Vaccine: 10 Good Reasons for Continuing. Hepatology 2020, 71, 1845–1850. [Google Scholar] [CrossRef]
- Roger, S.; Ducancelle, A.; Le Guillou-Guillemette, H.; Gaudy, C.; Lunel, F. HCV virology and diagnosis. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101626. [Google Scholar] [CrossRef]
- André, P.; Komurian-Pradel, F.; Deforges, S.; Perret, M.; Berland, J.L.; Sodoyer, M.; Pol, S.; Bréchot, C.; Paranhos-Baccalà, G.; Lotteau, V. Characterization of Low- and Very-Low-Density Hepatitis C Virus RNA-Containing Particles. J. Virol. 2002, 76, 7040–7048. [Google Scholar] [CrossRef] [Green Version]
- Boyer, A.; Dumans, A.; Beaumont, E.; Etienne, L.; Roingeard, P.; Meunier, J.-C. The Association of Hepatitis C Virus Glycoproteins with Apolipoproteins E and B Early in Assembly Is Conserved in Lipoviral Particles. J. Biol. Chem. 2014, 289, 18904–18913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, T.; Ono, C.; Puig-Basagoiti, F.; Matsuura, Y. Roles of Lipoproteins and Apolipoproteins in Particle Formation of Hepatitis C Virus. Trends Microbiol. 2015, 23, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J.-C.; Russell, R.S.; Engle, R.E.; Faulk, K.N.; Purcell, R.H.; Emerson, S.U. Apolipoprotein C1 Association with Hepatitis C Virus. J. Virol. 2008, 82, 9647–9656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piver, E.; Boyer, A.; Gaillard, J.; Bull, A.; Beaumont, E.; Roingeard, P.; Meunier, J.-C. Ultrastructural organisation of HCV from the bloodstream of infected patients revealed by electron microscopy after specific immunocapture. Gut 2016, 66, 1487–1495. [Google Scholar] [CrossRef]
- Butt, A.A.; Yan, P.; Simon, T.G.; Chung, R.T.; Abou-Samra, A.-B. ERCHIVES study team Changes in circulating lipids level over time after acquiring HCV infection: Results from ERCHIVES. BMC Infect. Dis. 2015, 15, 510. [Google Scholar] [CrossRef] [Green Version]
- Felmlee, D.J.; Sheridan, D.A.; Bridge, S.H.; Nielsen, S.U.; Milne, R.W.; Packard, C.J.; Caslake, M.J.; McLauchlan, J.; Toms, G.L.; Neely, R.D.G.; et al. Intravascular Transfer Contributes to Postprandial Increase in Numbers of Very-Low-Density Hepatitis C Virus Particles. Gastroenterology 2010, 139, 1774–1783. [Google Scholar] [CrossRef]
- Fauvelle, C.; Felmlee, D.J.; Crouchet, E.; Lee, J.; Heydmann, L.; Lefèvre, M.; Magri, A.; Hiet, M.-S.; Fofana, I.; Habersetzer, F.; et al. Apolipoprotein E Mediates Evasion From Hepatitis C Virus Neutralizing Antibodies. Gastroenterology 2016, 150, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Escobar, E.; Burlaud-Gaillard, J.; Visdeloup, C.; Silva, A.R.E.; Coutant, P.; Roingeard, P.; Beaumont, E. Incorporation of apolipoprotein E into HBV–HCV subviral envelope particles to improve the hepatitis vaccine strategy. Sci. Rep. 2021, 11, 21856. [Google Scholar] [CrossRef]
- Harris, H.J.; Davis, C.; Mullins, J.; Hu, K.; Goodall, M.; Farquhar, M.J.; Mee, C.; McCaffrey, K.; Young, S.; Drummer, H.; et al. Claudin Association with CD81 Defines Hepatitis C Virus Entry. J. Biol. Chem. 2010, 285, 21092–21102. [Google Scholar] [CrossRef] [Green Version]
- Meola, A.; Sbardellati, A.; Ercole, B.B.; Cerretani, M.; Pezzanera, M.; Ceccacci, A.; Vitelli, A.; Levy, S.; Nicosia, A.; Traboni, C.; et al. Binding of Hepatitis C Virus E2 Glycoprotein to CD81 Does Not Correlate with Species Permissiveness to Infection. J. Virol. 2000, 74, 5933–5938. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Randall, G.; Higginbottom, A.; Monk, P.; Rice, C.M.; McKeating, J.A. CD81 Is Required for Hepatitis C Virus Glycoprotein-Mediated Viral Infection. J. Virol. 2004, 78, 1448–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquhar, M.J.; Hu, K.; Harris, H.J.; Davis, C.; Brimacombe, C.L.; Fletcher, S.J.; Baumert, T.F.; Rappoport, J.Z.; Balfe, P.; McKeating, J.A. Hepatitis C Virus Induces CD81 and Claudin-1 Endocytosis. J. Virol. 2012, 86, 4305–4316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Wu, X.; Tang, H.; Luo, G. Apolipoprotein E Mediates Attachment of Clinical Hepatitis C Virus to Hepatocytes by Binding to Cell Surface Heparan Sulfate Proteoglycan Receptors. PLoS ONE 2013, 8, e67982. [Google Scholar] [CrossRef] [Green Version]
- Libeu, C.P.; Lund-Katz, S.; Phillips, M.C.; Wehrli, S.; Hernaiz, M.; Capila, I.; Linhardt, R.J.; Raffaï, R.L.; Newhouse, Y.M.; Zhou, F.; et al. New Insights into the Heparan Sulfate Proteoglycan-binding Activity of Apolipoprotein E. J. Biol. Chem. 2001, 276, 39138–39144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahley, R.W. Apolipoprotein E: Cholesterol transport protein with expanding role in cell biology. Science 1988, 240, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Mahley, R.W.; Ji, Z.-S. Remnant lipoprotein metabolism: Key pathways involving cell-surface heparan sulfate proteoglycans and apolipoprotein E. J. Lipid Res. 1999, 40, 1–16. [Google Scholar] [CrossRef]
- Agnello, V.; Ábel, G.; Elfahal, M.; Knight, G.B.; Zhang, Q.-X. Hepatitis C virus and other Flaviviridae viruses enter cells via low density lipoprotein receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 12766–12771. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B., Jr.; Barretto, N.; Martin, D.N.; Hiraga, N.; Imamura, M.; Hussain, S.; Marsh, K.A.; Yu, X.; Chayama, K.; Alrefai, W.A.; et al. Identification of the Niemann-Pick C1–like 1 cholesterol absorption receptor as a new hepatitis C virus entry factor. Nat. Med. 2012, 18, 281–285. [Google Scholar] [CrossRef]
- Scarselli, E.; Ansuini, H.; Cerino, R.; Roccasecca, R.M.; Acali, S.; Filocamo, G.; Traboni, C.; Nicosia, A.; Cortese, R.; Vitelli, A. The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus. EMBO J. 2002, 21, 5017–5025. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-S.; Jiang, J.; Cai, Z.; Luo, G. Human Apolipoprotein E Is Required for Infectivity and Production of Hepatitis C Virus in Cell Culture. J. Virol. 2007, 81, 13783–13793. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.-M.; Zhao, Y.-Z.; Chen, M.-T.; Zhang, F.-L.; Liu, X.-L.; Wang, Y.; Liu, C.-Z.; Yu, J.; Zhang, J.-W. Hypoxia-inducible factor-1 (HIF-1) promotes LDL and VLDL uptake through inducing VLDLR under hypoxia. Biochem. J. 2011, 441, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Kouiavskaia, D.; Migliorini, M.; Robinson, S.; Saenko, E.L.; Gorlatova, N.; Li, D.; Lawrence, D.; Hyman, B.T.; Weisgraber, K.H.; et al. The apoE isoform binding properties of the VLDL receptor reveal marked differences from LRP and the LDL receptor. J. Lipid Res. 2005, 46, 1721–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujino, S.; Nishitsuji, H.; Hishiki, T.; Sugiyama, K.; Takaku, H.; Shimotohno, K. Hepatitis C virus utilizes VLDLR as a novel entry pathway. Proc. Natl. Acad. Sci. USA 2015, 113, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Price, D.A.; Bassendine, M.F.; Norris, S.M.; Golding, C.; Toms, G.L.; Schmid, M.L.; Morris, C.; Burt, A.; Donaldson, P.T. Apolipoprotein 3 allele is associated with persistent hepatitis C virus infection. Gut 2006, 55, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Luo, G. Apolipoprotein E but Not B Is Required for the Formation of Infectious Hepatitis C Virus Particles. J. Virol. 2009, 83, 12680–12691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hueging, K.; Doepke, M.; Vieyres, G.; Bankwitz, D.; Frentzen, A.; Doerrbecker, J.; Gumz, F.; Haid, S.; Wölk, B.; Kaderali, L.; et al. Apolipoprotein E Codetermines Tissue Tropism of Hepatitis C Virus and Is Crucial for Viral Cell-to-Cell Transmission by Contributing to a Postenvelopment Step of Assembly. J. Virol. 2014, 88, 1433–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murayama, A.; Sugiyama, N.; Wakita, T.; Kato, T. Completion of the Entire Hepatitis C Virus Life Cycle in Vero Cells Derived from Monkey Kidney. mBio 2016, 7, e00273-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herker, E.; Harris, C.; Hernandez, C.; Carpentier, A.; Kaehlcke, K.; Rosenberg, A.R.; Farese, R.V., Jr.; Ott, M. Efficient hepatitis C virus particle formation requires diacylglycerol acyltransferase-1. Nat. Med. 2010, 16, 1295–1298. [Google Scholar] [CrossRef] [Green Version]
- Roingeard, P.; Melo, R.C.N. Lipid Droplet Hijacking by Intracellular Pathogens. Cell Microbiol. 2017, 19, e12688. [Google Scholar] [CrossRef]
- Benga, W.J.A.; Krieger, S.E.; Dimitrova, M.; Zeisel, M.B.; Parnot, M.; Lupberger, J.; Hildt, E.; Luo, G.; McLauchlan, J.; Baumert, T.F.; et al. Apolipoprotein E interacts with hepatitis C virus nonstructural protein 5A and determines assembly of infectious particles. Hepatology 2010, 51, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Appel, N.; Zayas, M.; Miller, S.; Krijnse-Locker, J.; Schaller, T.; Friebe, P.; Kallis, S.; Engel, U.; Bartenschlager, R. Essential Role of Domain III of Nonstructural Protein 5A for Hepatitis C Virus Infectious Particle Assembly. PLoS Pathog. 2008, 4, e1000035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camus, G.; Herker, E.; Modi, A.A.; Haas, J.T.; Ramage, H.R.; Farese, R.V., Jr.; Ott, M. Diacylglycerol Acyltransferase-1 Localizes Hepatitis C Virus NS5A Protein to Lipid Droplets and Enhances NS5A Interaction with the Viral Capsid Core. J. Biol. Chem. 2013, 288, 9915–9923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-Y.; Acosta, E.G.; Stoeck, I.K.; Long, G.; Hiet, M.-S.; Mueller, B.; Fackler, O.T.; Kallis, S.; Bartenschlager, R. Apolipoprotein E Likely Contributes to a Maturation Step of Infectious Hepatitis C Virus Particles and Interacts with Viral Envelope Glycoproteins. J. Virol. 2014, 88, 12422–12437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roingeard, P.; Dreneau, J.; Meunier, J.-C. Unravelling the multiple roles of apolipoprotein E in the hepatitis C virus life cycle. Gut 2016, 66, 759–761. [Google Scholar] [CrossRef] [PubMed]
- Alter, M.H.J.; Blumberg, M.B.S. Further Studies on a “New” Human Isoprecipitin System (Australia Antigen). Blood 1966, 27, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Marzio, D.H.-D.; Hann, H.-W. Then and now: The progress in hepatitis B treatment over the past 20 years. World, J. Gastroenterol. 2014, 20, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Maupas, P.; Coursaget, P.; Goudeau, A.; Drucker, J.; Bagros, P. Immunisation against Hepatitis B in Man. Lancet 1976, 307, 1367–1370. [Google Scholar] [CrossRef]
- Mokaya, J.; McNaughton, A.L.; Bester, P.A.; Goedhals, D.; Barnes, E.; Marsden, B.D.; Matthews, P.C. Hepatitis B virus resistance to tenofovir: Fact or fiction? A systematic literature review and structural analysis of drug resistance mechanisms. Wellcome Open Res. 2020, 5, 151. [Google Scholar] [CrossRef]
- Jiang, Y.; Han, Q.; Zhao, H.; Zhang, J. The Mechanisms of HBV-Induced Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2021, 8, 435–450. [Google Scholar] [CrossRef]
- Beck, J. Hepatitis B virus replication. World J. Gastroenterol. 2007, 13, 48–64. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Guo, H. Hepatitis B virus cccDNA: Formation, regulation and therapeutic potential. Antivir. Res. 2020, 180, 104824. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.-J.; Wang, T.-T.; Gao, Y.-F.; Wang, Y.-Q.; Bao, T.; Zou, G.-Z. Serum Lipid Metabolic Derangement is Associated with Disease Progression During Chronic HBV Infection. Clin. Lab. 2019, 65, 65. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Kim, D.K.; Kim, S.S.; Bae, C.B.; Cho, H.J.; Kim, H.G.; Kim, Y.J.; Lee, J.H.; Lee, H.J.; Lee, M.Y.; et al. Association between apolipoprotein E genotype, chronic liver disease, and hepatitis B virus. Clin. Mol. Hepatol. 2012, 18, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, M.; Ye, X.; Bi, Q. Association of apolipoprotein E with the progression of hepatitis B virus-related liver disease. Int. J. Clin. Exp. Pathol. 2015, 8, 14749–14756. [Google Scholar] [PubMed]
- Toniutto, P.; Fattovich, G.; Fabris, C.; Minisini, R.; Burlone, M.; Pravadelli, C.; Peraro, L.; Falleti, E.; Caldera, F.; Bitetto, D.; et al. Genetic polymorphism at the apolipoprotein E locus affects the outcome of chronic hepatitis B. J. Med Virol. 2009, 82, 224–331. [Google Scholar] [CrossRef]
- Yin, Z.; Xiong, C.; Wang, Y.; Zhou, X.; Yan, S.-K. Investigation of the relationship between apolipoprotein E gene polymorphisms and hepatitis B virus infection in northern China. Clin. Chem. Lab. Med. (CCLM) 2010, 48, 1803–1807. [Google Scholar] [CrossRef]
- Patient, R.; Hourioux, C.; Roingeard, P. Morphogenesis of hepatitis B virus and its subviral envelope particles. Cell. Microbiol. 2009, 11, 1561–1570. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Himmelsbach, K.; Ren, H.; Boller, K.; Hildt, E. Subviral Hepatitis B Virus Filaments, like Infectious Viral Particles, Are Released via Multivesicular Bodies. J. Virol. 2016, 90, 3330–3341. [Google Scholar] [CrossRef] [Green Version]
- Patient, R.; Hourioux, C.; Sizaret, P.-Y.; Trassard, S.; Sureau, C.; Roingeard, P. Hepatitis B Virus Subviral Envelope Particle Morphogenesis and Intracellular Trafficking. J. Virol. 2007, 81, 3842–3851. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Lu, Y.; Geschwindt, R.; Dwek, R.A.; Block, T.M. Hepatitis B Virus MHBs Antigen Is Selectively Sensitive to Glucosidase-Mediated Processing in the Endoplasmic Reticulum. Dna Cell Biol. 2001, 20, 647–656. [Google Scholar] [CrossRef]
- Inoue, J.; Krueger, E.W.; Chen, J.; Cao, H.; Ninomiya, M.; McNiven, M.A. HBV secretion is regulated through the activation of endocytic and autophagic compartments via Rab7 stimulation. J. Cell Sci. 2015, 128, 1696–1706. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Hildt, E. Intracellular Trafficking of HBV Particles. Cells 2020, 9, 2023. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Luo, G.G. Human apolipoprotein E promotes hepatitis B virus infection and production. PLoS Pathog. 2019, 15, e1007874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Luo, G. Human low-density lipoprotein receptor plays an important role in hepatitis B virus infection. PLoS Pathog. 2021, 17, e1009722. [Google Scholar] [CrossRef] [PubMed]
- Leistner, C.M.; Gruen-Bernhard, S.; Glebe, D. Role of glycosaminoglycans for binding and infection of hepatitis B virus. Cell. Microbiol. 2007, 10, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Schulze, A.; Gripon, P.; Urban, S. Hepatitis B virus infection initiates with a large surface protein-dependent binding to heparan sulfate proteoglycans. Hepatology 2007, 46, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Sureau, C.; Guerra, B.; Lanford, R.E. Role of the large hepatitis B virus envelope protein in infectivity of the hepatitis delta virion. J. Virol. 1993, 67, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Watashi, K.; Wakita, T. Hepatitis B Virus and Hepatitis D Virus Entry, Species Specificity, and Tissue Tropism. Cold Spring Harb. Perspect. Med. 2015, 5, a021378. [Google Scholar] [CrossRef]
- Feng, Z.D.; Hensley, L.; McKnight, K.L.; Hu, F.Y.; Madden, V.; Ping, L.F.; Jeong, S.-H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the ORF3 protein of hepatitis E virus (HEV) can capture HEV particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Jirintai; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; et al. Hepatitis E Virus (HEV) Strains in Serum Samples Can Replicate Efficiently in Cultured Cells Despite the Coexistence of HEV Antibodies: Characterization of HEV Virions in Blood Circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primadharsini, P.P.; Nagashima, S.; Nishiyama, T.; Takahashi, M.; Murata, K.; Okamoto, H. Development of recombinant infectious hepatitis E virus harboring the nanoKAZ gene and its application in drug screening. J. Virol. 2022, 92, 0190621. [Google Scholar] [CrossRef] [PubMed]
- Hirai-Yuki, A.; Hensley, L.; Whitmire, J.K.; Lemon, S.M. Biliary Secretion of Quasi-Enveloped Human Hepatitis A Virus. mBio 2016, 7, e01998-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Hirai-Yuki, A.; McKnight, K.L.; Lemon, S.M. Naked Viruses That Aren’t Always Naked: Quasi-Enveloped Agents of Acute Hepatitis. Annu. Rev. Virol. 2014, 1, 539–560. [Google Scholar] [CrossRef]
- McKnight, K.L.; Xie, L.; González-López, O.; Rivera-Serrano, E.E.; Chen, X.; Lemon, S.M. Protein composition of the hepatitis A virus quasi-envelope. Proc. Natl. Acad. Sci. USA 2017, 114, 6587–6592. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Tanggis Characterization of the Quasi-Enveloped Hepatitis E Virus Particles Released by the Cellular Exosomal Pathway. J. Virol. 2017, 91, e00822-17. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, S.; Takahashi, M.; Jirintai, S.; Tanaka, T.; Nishizawa, T.; Yasuda, J.; Okamoto, H. Tumour susceptibility gene 101 and the vacuolar protein sorting pathway are required for the release of hepatitis E virions. J. Gen. Virol. 2011, 92, 2838–2848. [Google Scholar] [CrossRef]
- Taneja, S.; Ahmad, I.; Sen, S.; Kumar, S.; Arora, R.; Gupta, V.K.; Aggarwal, R.; Narayanasamy, K.; Reddy, V.S.; Jameel, S. Plasma peptidome profiling of acute hepatitis E patients by MALDI-TOF/TOF. Proteome Sci. 2011, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.; Qiu, J.; Guo, D.; Xu, Y.; Liu, X.; Shen, C.; Dong, C. Evaluation of candidate genes associated with hepatitis A and E virus infection in Chinese Han population. Virol. J. 2018, 15, 47. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yesupriya, A.; Chang, M.-H.; Teshale, E.; Teo, C.-G. Apolipoprotein E and protection against hepatitis E viral infection in American non-Hispanic blacks. Hepatology 2015, 62, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tréguier, Y.; Bull-Maurer, A.; Roingeard, P. Apolipoprotein E, a Crucial Cellular Protein in the Lifecycle of Hepatitis Viruses. Int. J. Mol. Sci. 2022, 23, 3676. https://doi.org/10.3390/ijms23073676
Tréguier Y, Bull-Maurer A, Roingeard P. Apolipoprotein E, a Crucial Cellular Protein in the Lifecycle of Hepatitis Viruses. International Journal of Molecular Sciences. 2022; 23(7):3676. https://doi.org/10.3390/ijms23073676
Chicago/Turabian StyleTréguier, Yannick, Anne Bull-Maurer, and Philippe Roingeard. 2022. "Apolipoprotein E, a Crucial Cellular Protein in the Lifecycle of Hepatitis Viruses" International Journal of Molecular Sciences 23, no. 7: 3676. https://doi.org/10.3390/ijms23073676
APA StyleTréguier, Y., Bull-Maurer, A., & Roingeard, P. (2022). Apolipoprotein E, a Crucial Cellular Protein in the Lifecycle of Hepatitis Viruses. International Journal of Molecular Sciences, 23(7), 3676. https://doi.org/10.3390/ijms23073676