
Perspectives on Potential Fatty Acid Modulations of Motility Associated Human Sperm Ion Channels



Abstract
:1. Introduction
1.1. Human Spermatozoa
1.2. Sperm Capacitation
2. Motility-Associated Major Ion Channels
2.1. Calcium Channels
2.2. Potassium Channels
2.3. Proton Channels
2.4. Sodium Channels
2.5. Chloride Channels
3. Lipids and Spermatozoa
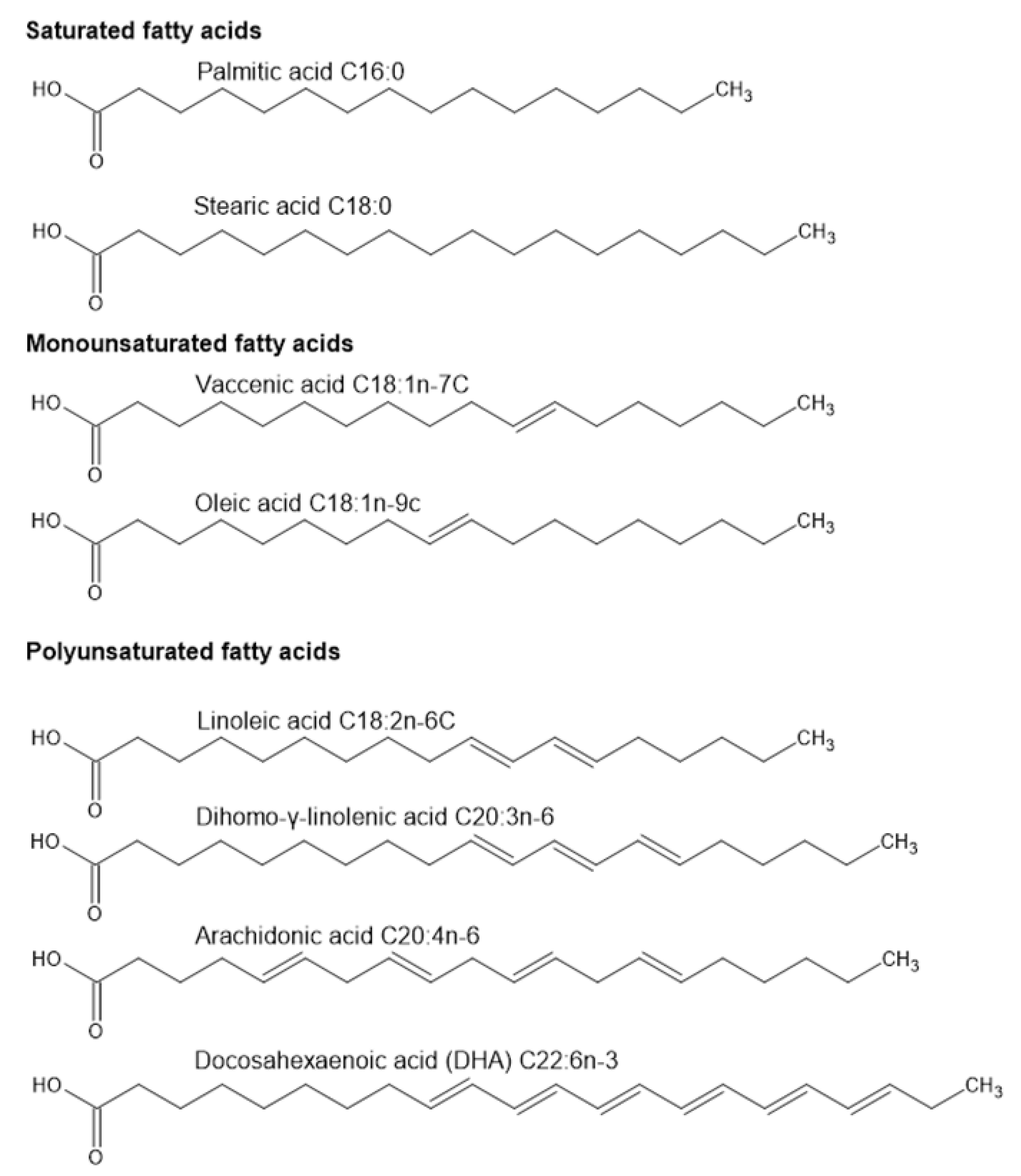
3.1. Fatty Acid Composition of Spermatozoa and Seminal Fluid
3.2. Fatty Acids Associated Sperm Abnormalities
3.3. Normozoospermic and Non-Normozoospermic Spermatozoa FA Profiles
3.4. Fertile and Infertile Subjects’ Spermatozoa FA Profiles
3.5. Sperm FA Metabolism and Modifications
4. Fatty Acids and Sperm Motility
4.1. Effect of Lipids/FAs on Ion Channels
4.1.1. Calcium Channels and FAs
4.1.2. K+ Channels and FAs
4.1.3. Hv1 and FAs
4.1.4. Other Ion Channels and FAs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| ART | Assisted reproductive technology |
| CaCC | Ca2+-activated chloride channel |
| CatSper | Cation channel of sperm |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| ClC3 | Chloride channel 3 |
| CNGK | Cyclic nucleotide-gated K+ channel |
| DHA | Docosahexaenoic acid |
| ENaC | Epithelial sodium channel |
| EPA | Eicosapentaenoic acid |
| FA | Fatty acid |
| FFA | Free fatty acid |
| Hv1 | Voltage-gated proton channel |
| Kir | Inwardly rectifying K+ channel |
| KV | Voltage-gated K+ channel |
| MUFA | Monounsaturated fatty acid |
| PLA2 | Phospholipase A2 |
| PUFA | Polyunsaturated fatty acid |
| ROS | Reactive oxygen species |
| SFA | Saturated fatty acid |
| SOCC | Store-operated Ca2+ channel |
| SSC | Spermatogonial stem cell |
| TRPV | Transient receptor potential vanilloid |
| VGCC | Voltage-gated calcium channel |
| VGNC/Nav | Voltage-gated sodium channel |
| VSD | Voltage sensing domains |
References
- Abou-Haïla, A.; Tulsiani, D.R. Signal transduction pathways that regulate sperm capacitation and the acrosome reaction. Arch. Biochem. Biophys. 2009, 485, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Gadêlha, H.; Hernández-Herrera, P.; Montoya, F.; Darszon, A.; Corkidi, G. Human sperm uses asymmetric and anisotropic flagellar controls to regulate swimming symmetry and cell steering. Sci. Adv. 2020, 6, eaba5168. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Castillo, J.; Estanyol, J.M.; Ballescà, J.L.; Ramalho-Santos, J.; Oliva, R. Human Sperm Tail Proteome Suggests New Endogenous Metabolic Pathways. Mol. Cell. Proteom. 2013, 12, 330–342. [Google Scholar] [CrossRef] [Green Version]
- El-Ghobashy, A.A.; West, C.R. The human sperm head: A key for successful fertilization. J. Androl. 2003, 24, 232–238. [Google Scholar] [CrossRef]
- Ugajin, T.; Terada, Y.; Hasegawa, H.; Nabeshima, H.; Suzuki, K.; Yaegashi, N. The shape of the sperm midpiece in intracytoplasmic morphologically selected sperm injection relates sperm centrosomal function. J. Assist. Reprod. Genet. 2009, 27, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Mundy, A.; Ryder, T.; Edmonds, D. Asthenozoospermia and the human sperm mid-piece. Hum. Reprod. 1995, 10, 116–119. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Szymczak-Cendlak, M. Structure and Function of Ion Channels Regulating Sperm Motility—An Overview. Int. J. Mol. Sci. 2021, 22, 3259. [Google Scholar] [CrossRef] [PubMed]
- Storey, B.T. Sperm Capacitation and the Acrosome Reaction. Ann. N. Y. Acad. Sci. 1991, 637, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Kirichok, Y.; Ren, D.; Navarro, B.; Chung, J.-J.; Clapham, D.E. The Control of Male Fertility by Spermatozoan Ion Channels. Annu. Rev. Physiol. 2012, 74, 453–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourmoghadam, Z.; Aghebati-Maleki, L.; Motallebnezhad, M.; Yousefi, B.; Yousefi, M. Current approaches for the treatment of male infertility with stem cell therapy. J. Cell. Physiol. 2018, 233, 6455–6469. [Google Scholar] [CrossRef]
- Kumar, R.; Gautam, G.; Gupta, N.P. Drug Therapy for Idiopathic Male Infertility: Rationale Versus Evidence. J. Urol. 2006, 176, 1307–1312. [Google Scholar] [CrossRef]
- Finkelstein, M.; Etkovitz, N.; Breitbart, H. Ca2+ signaling in mammalian spermatozoa. Mol. Cell. Endocrinol. 2020, 516, 110953. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; Sá, R.; Barros, A.; Sousa, M. Major regulatory mechanisms involved in sperm motility. Asian J. Androl. 2017. [Google Scholar] [CrossRef] [PubMed]
- Achikanu, C.; Pendekanti, V.; Teague, R.; Publicover, S. Effects of pH manipulation, CatSper stimulation and Ca2+-store mobilization on [Ca2+]i and behaviour of human sperm. Hum. Reprod. 2018, 33, 1802–1811. [Google Scholar] [CrossRef] [PubMed]
- Rahban, R.; Nef, S. CatSper: The complex main gate of calcium entry in mammalian spermatozoa. Mol. Cell. Endocrinol. 2020, 518, 110951. [Google Scholar] [CrossRef] [PubMed]
- Bedu-Addo, K.; Costello, S.; Harper, C.; Machado-Oliveira, G.; Lefièvre, L.; Ford, C.; Barratt, C.; Publicover, S. Mobilisation of stored calcium in the neck region of human sperm a mechanism for regulation of flagellar activity. Int. J. Dev. Biol. 2008, 52, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Keshtgar, S.; Ghanbari, H.; Ghani, E.; Moosavi, S.M.S. Effect of CatSper and Hv1 Channel Inhibition on Progesterone Stimulated Human Sperm. J. Reprod. Infertil. 2018, 19, 133–139. [Google Scholar]
- Catterall, W.A. Voltage-Gated Calcium Channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- José, O.; Hernández-Hernández, O.; Chirinos, M.; González-González, M.E.; Larrea, F.; Almanza, A.; Felix, R.; Darszon, A.; Treviño, C.L. Recombinant human ZP3-induced sperm acrosome reaction: Evidence for the involvement of T- and L-type voltage-gated calcium channels. Biochem. Biophys. Res. Commun. 2010, 395, 530–534. [Google Scholar] [CrossRef]
- Clapham, D.E.; Julius, D.; Montell, C.; Schultz, G. International Union of Pharmacology. XLIX. Nomenclature and Structure-Function Relationships of Transient Receptor Potential Channels. Pharmacol. Rev. 2005, 57, 427–450. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Yang, F.; Takanishi, C.L.; Zheng, J. Thermosensitive TRPV Channel Subunits Coassemble into Heteromeric Channels with Intermediate Conductance and Gating Properties. J. Gen. Physiol. 2007, 129, 191–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Toni, L.; Garolla, A.; Menegazzo, M.; Magagna, S.; di Nisio, A.; Šabović, I.; Rocca, M.S.; Scattolini, V.; Filippi, A.; Foresta, C. Heat Sensing Receptor TRPV1 Is a Mediator of Thermotaxis in Human Spermatozoa. PLoS ONE 2016, 11, e0167622. [Google Scholar] [CrossRef] [Green Version]
- Majhi, R.K.; Kumar, A.; Yadav, M.; Swain, N.; Kumari, S.; Saha, A.; Pradhan, A.; Goswami, L.; Saha, S.; Samanta, L.; et al. Thermosensitive ion channel TRPV1 is endogenously expressed in the sperm of a fresh water teleost fish (Labeo rohita) and regulates sperm motility. Channels 2013, 7, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, H.; Wang, F.; Chen, C.; Zhang, P.; Song, D.; Luo, T.; Xu, H.; Zeng, X. Sperm motility modulated by Trpv1 regulates zebrafish fertilization. Theriogenology 2020, 151, 41–51. [Google Scholar] [CrossRef]
- Kumar, A.; Mishra, A.K.; Singh, V.; Yadav, S.; Saxena, A.; Garg, S.K.; Swain, D.K. Molecular and functional insights into Transient Receptor Potential Vanilloid 1 (TRPV1) in bull spermatozoa. Theriogenology 2019, 128, 207–217. [Google Scholar] [CrossRef]
- Kumar, A.; Majhi, R.K.; Swain, N.; Giri, S.; Kar, S.; Samanta, L.; Goswami, C. TRPV4 is endogenously expressed in vertebrate spermatozoa and regulates intracellular calcium in human sperm. Biochem. Biophys. Res. Commun. 2016, 473, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Mundt, N.; Spehr, M.; Lishko, P.V. TRPV4 is the temperature-sensitive ion channel of human sperm. eLife 2018, 7, e35853. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Ishikawa, M.; Izumi, H.; de Santis, R.; Morisawa, M. Store-operated calcium channel regulates the chemotactic behavior of ascidian sperm. Proc. Natl. Acad. Sci. USA 2003, 100, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Krasznai, Z.; Krasznai, Z.T.; Morisawa, M.; Bazsáné, Z.K.; Hernádi, Z.; Fazekas, Z.; Trón, L.; Goda, K.; Márián, T. Role of the Na+/Ca2+ exchanger in calcium homeostasis and human sperm motility regulation. Cell Motil. Cytoskelet. 2006, 63, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Arias, R.D.; Vívenes, C.Y.; I Camejo, M.; Piñero, S.; Proverbio, T.; Martínez, E.; Marín, R.; Proverbio, F. ATPases, ion exchangers and human sperm motility. Reproduction 2015, 149, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannowetz, N.; Naidoo, N.M.; Choo, S.-A.S.; Smith, J.F.; Lishko, P.V. Slo1 is the principal potassium channel of human spermatozoa. eLife 2013, 2, e01009. [Google Scholar] [CrossRef]
- Muñoz-Garay, C.; de la Vega-Beltrán, J.L.; Delgado, R.; Labarca, P.; Felix, R.; Darszon, A. Inwardly Rectifying K+ Channels in Spermatogenic Cells: Functional Expression and Implication in Sperm Capacitation. Dev. Biol. 2001, 234, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Swain, D.K.; Singh, V.; Anand, M.; Choudhury, S.; Yadav, S.; Saxena, A.; Garg, S.K. Molecular characterization of voltage-gated potassium channel (Kv) and its importance in functional dynamics in bull spermatozoa. Theriogenology 2018, 114, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Fechner, S.; Alvarez, L.; Bönigk, W.; Müller, A.; Berger, T.K.; Pascal, R.; Troetschel, C.; Poetsch, A.; Stölting, G.; Siegfried, K.; et al. A K+-selective CNG channel orchestrates Ca2+ signalling in zebrafish sperm. eLife 2015, 4, e07624. [Google Scholar] [CrossRef] [PubMed]
- Brenker, C.; Zhou, Y.; Müller, A.; Echeverry, F.A.; Troetschel, C.; Poetsch, A.; Xia, X.-M.; Bönigk, W.; Lingle, C.J.; Kaupp, U.B.; et al. The Ca2+-activated K+ current of human sperm is mediated by Slo3. eLife 2014, 3, e01438. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.-H.; Yang, C.; Kim, S.T.; Lingle, C.J.; Xia, X.-M. Deletion of the Slo3 gene abolishes alkalization-activated K + current in mouse spermatozoa. Proc. Natl. Acad. Sci. USA 2011, 108, 5879–5884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lishko, P.V.; Kirichok, Y. The role of Hv1 and CatSper channels in sperm activation. J. Physiol. 2010, 588, 4667–4672. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Botchkina, I.L.; Fedorenko, A.; Kirichok, Y. Acid Extrusion from Human Spermatozoa Is Mediated by Flagellar Voltage-Gated Proton Channel. Cell 2010, 140, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros-Volante, A.; Treviño, C.L. Capacitation-associated alkalization in human sperm is differentially controlled at the subcellular level. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef] [PubMed]
- Candenas, L.; Pinto, F.; Cejudo-Román, A.; González-Ravina, C.; Fernández-Sánchez, M.; Pérez-Hernández, N.; Irazusta, J.; Subirán, N. Veratridine-sensitive Na + channels regulate human sperm fertilization capacity. Life Sci. 2018, 196, 48–55. [Google Scholar] [CrossRef]
- Pinto, F.M.; Ravina, C.G.; Fernández-Sánchez, M.; Gallardo-Castro, M.; Cejudo-Román, A.; Candenas, L. Molecular and functional characterization of voltage-gated sodium channels in human sperm. Reprod. Biol. Endocrinol. 2009, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cejudo-Roman, A.; Pinto, F.M.; Subirán, N.; Ravina, C.G.; Fernández-Sánchez, M.; Pérez-Hernández, N.; Pérez, R.; Pacheco, A.; Irazusta, J.; Candenas, L. The Voltage-Gated Sodium Channel Nav1.8 Is Expressed in Human Sperm. PLoS ONE 2013, 8, e76084. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.-B.; Ma, H.-G.; Li, H.-G.; Xiong, C.-L. Blockade of epithelial sodium channels improves sperm motility in asthenospermia patients. Int. J. Androl. 2009, 32, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Puga Molina, L.C.; Luque, G.M.; Balestrini, P.A.; Marín-Briggiler, C.I.; Romarowski, A.; Buffone, M.G. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-W.; Chen, L.-X.; Liu, S.-W.; Li, Y.; Zou, L.-L.; Guan, Y.-T.; Peng, S.; Zheng, L.-X.; Deng, S.-M.; Zhu, L.-Y. Chloride channels are involved in sperm motility and are downregulated in spermatozoa from patients with asthenozoospermia. Asian J. Androl. 2016, 18, 418–424. [Google Scholar] [CrossRef]
- Yeung, C.; Barfield, J.; Cooper, T. Chloride Channels in Physiological Volume Regulation of Human Spermatozoa. Biol. Reprod. 2005, 73, 1057–1063. [Google Scholar] [CrossRef]
- Smith, G.D.; Wolf, D.P.; Trautman, K.C.; Silva, E.F.D.C.E.; Greengard, P.; Vijayaraghavan, S. Primate Sperm Contain Protein Phosphatase 1, a Biochemical Mediator of Motility. Biol. Reprod. 1996, 54, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Martínez, J.; Reyes-Miguel, T.; Rodríguez-Páez, L.; Garduño, L.; García, D.M.; Roa-Espitia, A.L.; Hernández-González, E.O. TMEM16A inhibition impedes capacitation and acquisition of hyperactivated motility in guinea pig sperm. J. Cell. Biochem. 2018, 119, 5944–5959. [Google Scholar] [CrossRef] [PubMed]
- Chávez, J.C.; Hernández-González, E.O.; Wertheimer, E.; Visconti, P.E.; Darszon, A.; Treviño, C.L. Participation of the Cl−/HCO3− Exchangers SLC26A3 and SLC26A6, the Cl− Channel CFTR, and the Regulatory Factor SLC9A3R1 in Mouse Sperm Capacitation. Biol. Reprod. 2012, 86, 1–14. [Google Scholar] [CrossRef]
- Shan, S.; Xu, F.; Hirschfeld, M.; Brenig, B. Sperm Lipid Markers of Male Fertility in Mammals. Int. J. Mol. Sci. 2021, 22, 8767. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Mukai, C.; Travis, A.J. Lipid Regulation of Acrosome Exocytosis. Adv. Anat. Embryol. Cell Biol. 2016, 220, 107–127. [Google Scholar] [CrossRef]
- Jones, R.; James, P.S.; Howes, L.; Bruckbauer, A.; Klenerman, D. Supramolecular organization of the sperm plasma membrane during maturation and capacitation. Asian J. Androl. 2007, 9, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Flesch, F.; Gadella, B.M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim. Biophys. Acta 2000, 1469, 197–235. [Google Scholar] [CrossRef]
- de Carvalho, M.J.C.R.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, B. Dietary fatty acids. Am. Fam. Physician 2009, 80, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Zalata, A.; Christophe, A.B.; Depuydt, C.E.; Schoonjans, F.; Comhaire, F.H. The fatty acid composition of phospholipids of spermatozoa from infertile patients. Mol. Hum. Reprod. 1998, 4, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collodel, G.; Castellini, C.; Lee, J.C.-Y.; Signorini, C. Relevance of Fatty Acids to Sperm Maturation and Quality. Oxidative Med. Cell. Longev. 2020, 2020, 7038124-14. [Google Scholar] [CrossRef]
- Zerbinati, C.; Caponecchia, L.; Rago, R.; Leoncini, E.; Bottaccioli, A.G.; Ciacciarelli, M.; Pacelli, A.; Salacone, P.; Sebastianelli, A.; Pastore, A.; et al. Fatty acids profiling reveals potential candidate markers of semen quality. Andrology 2016, 4, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; Bulló, M.; Salas-Salvadó, J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: A systematic review of observational studies. Hum. Reprod. Update 2017, 23, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Karayiannis, D.; Kontogianni, M.; Mendorou, C.; Douka, L.; Mastrominas, M.; Yiannakouris, N. Association between adherence to the Mediterranean diet and semen quality parameters in male partners of couples attempting fertility. Hum. Reprod. 2017, 32, 215–222. [Google Scholar] [CrossRef]
- Afeiche, M.C.; Bridges, N.D.; Williams, P.L.; Gaskins, A.J.; Tanrikut, C.; Petrozza, J.C.; Hauser, R.; Chavarro, J. Dairy intake and semen quality among men attending a fertility clinic. Fertil. Steril. 2014, 101, 1280–1287.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afeiche, M.; Williams, P.L.; Mendiola, J.; Gaskins, A.J.; Jørgensen, N.; Swan, S.H.; Chavarro, J. Dairy food intake in relation to semen quality and reproductive hormone levels among physically active young men. Hum. Reprod. 2013, 28, 2265–2275. [Google Scholar] [CrossRef] [PubMed]
- Attaman, J.A.; Toth, T.L.; Furtado, J.; Campos, H.; Hauser, R.; Chavarro, J.E. Dietary fat and semen quality among men attending a fertility clinic. Hum. Reprod. 2012, 27, 1466–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, T.K.; Heitmann, B.L.; Jensen, M.B.; Halldorsson, T.I.; Andersson, A.-M.; Skakkebaek, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Lassen, T.H.; et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.-R.; Baghestani, A.-R.; Hekmatdoost, A. Dietary fatty acid intakes and asthenozoospermia: A case-control study. Fertil. Steril. 2015, 103, 190–198. [Google Scholar] [CrossRef]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.R.; Hekmatdoost, A. Nutrient patterns and asthenozoospermia: A case-control study. Andrologia 2017, 49, e12624. [Google Scholar] [CrossRef] [PubMed]
- Falsig, A.-M.L.; Gleerup, C.S.; Knudsen, U.B. The influence of omega-3 fatty acids on semen quality markers: A systematic PRISMA review. Andrology 2019, 7, 794–803. [Google Scholar] [CrossRef] [Green Version]
- Chavarro, J.; Minguez-Alarcon, L.; Mendiola, J.; Cutillas-Tolin, A.; Lopez-Espin, J.J.; Torres-Cantero, A.M. Trans fatty acid intake is inversely related to total sperm count in young healthy men. Hum. Reprod. 2014, 29, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Tayebinia, H.; Doosti, M.; Nourmohammadi, I.; Mahjub, H.; Vaisiraygani, A.; Salimi, S.; Hosseinipanah, S.M. Lipid composition of spermatozoa in normozoospermic and asthenozoospermic males. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 45–50. [Google Scholar] [CrossRef]
- Aksoy, Y.; Aksoy, H.; Altınkaynak, K.; Aydın, H.R.; Özkan, A. Sperm fatty acid composition in subfertile men. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 75–79. [Google Scholar] [CrossRef]
- Gulaya, N.M.; Margitich, V.M.; Govseeva, N.M.; Klimashevsky, V.M.; Gorpynchenko, I.I.; Boyko, M.I. Sperm and Seminal Plasma in Relation to sperm fertility. Arch. Androl. 2001, 46, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Soto, J.C.; Landeras, J.; Gadea, J. Spermatozoa and seminal plasma fatty acids as predictors of cryopreservation success. Andrology 2013, 1, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Safarinejad, M.R.; Hosseini, Y.; Dadkhah, F.; Asgari, M.A. Relationship of omega-3 and omega-6 fatty acids with semen characteristics, and anti-oxidant status of seminal plasma: A comparison between fertile and infertile men. Clin. Nutr. 2010, 29, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.; Costa, C.; Bassaizteguy, V.; Santos, M.; Cardozo, R.; Montes, J.; Settineri, R.; Nicolson, G.L. Incubation of human sperm with micelles made from glycerophospholipid mixtures increases sperm motility and resistance to oxidative stress. PLoS ONE 2018, 13, e0197897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collodel, G.; Castellini, C.; Iacoponi, F.; Noto, D.; Signorini, C. Cytosolic phospholipase A 2 and F 2 isoprostanes are involved in semen quality and human infertility—A study on leucocytospermia, varicocele and idiopathic infertility. Andrologia 2020, 52, e13465. [Google Scholar] [CrossRef]
- Collodel, G.; Moretti, E.; Noto, D.; Iacoponi, F.; Signorini, C. Fatty Acid Profile and Metabolism Are Related to Human Sperm Parameters and Are Relevant in Idiopathic Infertility and Varicocele. Mediat. Inflamm. 2020, 2020, 3640450. [Google Scholar] [CrossRef]
- Collodel, G.; Moretti, E.; Longini, M.; Pascarelli, N.A.; Signorini, C. Increased F2-Isoprostane Levels in Semen and Immunolocalization of the 8-Iso Prostaglandin F2α in Spermatozoa from Infertile Patients with Varicocele. Oxidative Med. Cell. Longev. 2018, 2018, 7508014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, E.; Collodel, G.; Salvatici, M.C.; Belmonte, G.; Signorini, C. New insights into sperm with total globozoospermia: Increased fatty acid oxidation and centrin1 alteration. Syst. Biol. Reprod. Med. 2019, 65, 390–399. [Google Scholar] [CrossRef]
- Aitken, R.J.; Wingate, J.K.; de Iuliis, G.; Koppers, A.J.; McLaughlin, E. Cis-Unsaturated Fatty Acids Stimulate Reactive Oxygen Species Generation and Lipid Peroxidation in Human Spermatozoa. J. Clin. Endocrinol. Metab. 2006, 91, 4154–4163. [Google Scholar] [CrossRef] [Green Version]
- Nissen, H.; Kreysel, H. Polyunsaturated Fatty Acids in Relation to Sperm Motility. Andrologia 1983, 15, 264–269. [Google Scholar] [CrossRef]
- Esmaeili, V.; Shahverdi, A.; Moghadasian, M.H.; Alizadeh, A.R. Dietary fatty acids affect semen quality: A review. Andrology 2015, 3, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Martin, J.B.; Tummon, I.; Watson, L.; Tekpetey, F. Effect of DHA supplementation on DHA status and sperm motility in asthenozoospermic males. Lipids 2000, 35, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Siegel, I.; Dudkiewicz, A.B.; Friberg, J.; Suarez, M.; Gleicher, N. Inhibition of sperm motility and agglutination of sperm cells by free fatty acids in whole semen**Supported by the Foundation for Reproductive Medicine, Inc., Chicago, Illinois. Fertil. Steril. 1986, 45, 273–279. [Google Scholar] [CrossRef]
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nature 2001, 413, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-H.; Zhu, Y.-Y.; Wang, L.; Liu, H.-L.; Ling, Y.; Li, Z.-L.; Sun, L.-B. The Catsper channel and its roles in male fertility: A systematic review. Reprod. Biol. Endocrinol. 2017, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.-F.; Gomez, A.M.; Morgan, J.P.; Lederer, W.J.; Leaf, A. Suppression of voltage-gated L-type Ca 2+ currents by polyunsaturated fatty acids in adult and neonatal rat ventricular myocytes. Proc. Natl. Acad. Sci. USA 1997, 94, 4182–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.J. Inhibition of L-type Ca2+channel current and negative inotropy induced by arachidonic acid in adult rat ventricular myocytes. Am. J. Physiol. Physiol. 2007, 293, C1594–C1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, L.E.; Treviño, C.L.; Rodríguez, D.; Serrano, C.J.; Pacheco, J.; Tsutsumi, V.; Felix, R.; Darszon, A. Transient receptor potential (TRPC) channels in human sperm: Expression, cellular localization and involvement in the regulation of flagellar motility. FEBS Lett. 2003, 541, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Redmond, W.J.; Gu, L.; Camo, M.; McIntyre, P.; Connor, M. Ligand determinants of fatty acid activation of the pronociceptive ion channel TRPA1. PeerJ 2014, 2, e248. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T.; Nilius, B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 2003, 424, 434–438. [Google Scholar] [CrossRef]
- Costello, S.; Michelangeli, F.; Nash, K.; Lefievre, L.; Morris, J.; Machado-Oliveira, G.; Barratt, C.; Kirkman-Brown, J.; Publicover, S. Ca2+-stores in sperm: Their identities and functions. Reproduction 2009, 138, 425–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakriya, M.; Lewis, R.S. Store-operated calcium channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, P.; Moncada, M.; Enrique, N.; Asuaje, A.; Capuccino, J.M.V.; Gonzalez, C.; Milesi, V. Arachidonic acid activation of BKCa (Slo1) channels associated to the β1-subunit in human vascular smooth muscle cells. Pflugers Arch. Eur. J. Physiol. 2014, 466, 1779–1792. [Google Scholar] [CrossRef] [PubMed]
- Denson, D.D.; Wang, X.; Worrell, R.T.; Eaton, D. Effects of fatty acids on BK channels in GH3cells. Am. J. Physiol. Physiol. 2000, 279, C1211–C1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, K.K.; Mahdi, A.A.; Rajender, S. Ion Channels in Sperm Physiology and Male Fertility and Infertility. J. Androl. 2012, 33, 777–788. [Google Scholar] [CrossRef]
- Danthi, S.; Enyeart, J.A. Modulation of Native TREK-1 and Kv1.4 K + Channels by Polyunsaturated Fatty Acids and Lysophospholipids. J. Membr. Biol. 2003, 195, 147–164. [Google Scholar] [CrossRef]
- Honoré, E.; Barhanin, J.; Attali, B.; Lesage, F.; Lazdunski, M. External blockade of the major cardiacdelayed-rectifier K+ channel (Kv1.5) by polyunsaturated fattyacids. Proc. Natl. Acad. Sci. USA 1994, 91, 1937–1941. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.-Y.; Ding, W.-G.; Kojima, A.; Seto, T.; Matsuura, H. Putative binding sites for arachidonic acid on the human cardiac Kv1.5 channel. Br. J. Pharmacol. 2015, 172, 5281–5292. [Google Scholar] [CrossRef] [Green Version]
- Seebungkert, B.; Lynch, J.W. Effects of polyunsaturated fatty acids on voltage-gated K+and Na+channels in rat olfactory receptor neurons. Eur. J. Neurosci. 2002, 16, 2085–2094. [Google Scholar] [CrossRef]
- Guizy, M.; David, M.; Arias, C.; Zhang, L.; Cofán, M.; Ruiz-Gutiérrez, V.; Ros, E.; Lillo, M.P.; Martens, J.R.; Valenzuela, C. Modulation of the atrial specific Kv1.5 channel by the n-3 polyunsaturated fatty acid, α-linolenic acid. J. Mol. Cell. Cardiol. 2008, 44, 323–335. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Morgan, J.P.; Leaf, A. Effects of polyunsaturated fatty acids on cardiac voltage-activated K(+) currents in adult ferret cardiomyocytes. Acta Physiol. Sin. 2002, 43, 271–281. [Google Scholar]
- Kawanabe, A.; Okamura, Y. Effects of unsaturated fatty acids on the kinetics of voltage-gated proton channels heterologously expressed in cultured cells. J. Physiol. 2016, 594, 595–610. [Google Scholar] [CrossRef] [PubMed]
- DeCoursey, T.; Cherny, V. Potential, pH, and arachidonate gate hydrogen ion currents in human neutrophils. Biophys. J. 1993, 65, 1590–1598. [Google Scholar] [CrossRef] [Green Version]
- Kapus, A.; Romanek, R.; Grinstein, S. Arachidonic acid stimulates the plasma membrane H+ conductance of macrophages. J. Biol. Chem. 1994, 269, 4736–4745. [Google Scholar] [CrossRef]
- Gordienko, D.V.; Tare, M.; Parveen, S.; Fenech, C.J.; Robinson, C.; Bolton, T.B. Voltage-activated proton current in eosinophils from human blood. J. Physiol. 1996, 496, 299–316. [Google Scholar] [CrossRef] [Green Version]
- Cherny, V.V.; Henderson, L.M.; Xu, W.; Thomas, L.L.; DeCoursey, T. Activation of NADPH oxidase-related proton and electron currents in human eosinophils by arachidonic acid. J. Physiol. 2001, 535, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.M.; Chappell, J.B. The NADPH-oxidase-associated H+ channel is opened by arachidonate. Biochem. J. 1992, 283, 171–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, M.-P.; Kim, H.I.; Shin, Y.K.; Lee, C.S.; Park, M.; Song, J.-H. Effects of free fatty acids on sodium currents in rat dorsal root ganglion neurons. Brain Res. 2004, 1008, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-R.; Sun, H.-Y.; Zhang, X.-H.; Cheng, L.-C.; Chiu, S.-W.; Tse, H.-F.; Lau, C.-P. Omega-3 polyunsaturated fatty acids inhibit transient outward and ultra-rapid delayed rectifier K+currents and Na+current in human atrial myocytes. Cardiovasc. Res. 2009, 81, 286–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.F.; Kang, J.X.; Morgan, J.P.; Leaf, A. Blocking effects of polyunsaturated fatty acids on Na+ channels of neonatal rat ventricular myocytes. Proc. Natl. Acad. Sci. USA 1995, 92, 11000–11004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.-F.; Sigg, D.; Leaf, A. The Antiarrhythmic Effect of n-3 Polyunsaturated Fatty Acids: Modulation of Cardiac Ion Channels as a Potential Mechanism. J. Membr. Biol. 2005, 206, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Fang, Y.-J.; He, Y.-L.; Sun, J.; Zhu, J.; Mei, Y.-A. Modulation of muscle rNav1.4 Na+channel isoform by arachidonic acid and its non-metabolized analog. J. Cell. Physiol. 2009, 219, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lin, D.-H.; Kemp, R.; Yaddanapudi, G.S.; Nasjletti, A.; Falck, J.R.; Wang, W.-H. Arachidonic Acid Inhibits Epithelial Na Channel Via Cytochrome P450 (CYP) Epoxygenase-dependent Metabolic Pathways. J. Gen. Physiol. 2004, 124, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Orta, G.; Ferreira, G.; José, O.; Treviño, C.L.; Beltrán, C.; Darszon, A. Human spermatozoa possess a calcium-dependent chloride channel that may participate in the acrosomal reaction. J. Physiol. 2012, 590, 2659–2675. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.M.; Shi, Q.X.; Chen, W.Y.; Zhou, C.X.; Ni, Y.; Rowlands, D.K.; Liu, G.Y.; Zhu, H.; Ma, Z.G.; Wang, X.F.; et al. Cystic fibrosis transmembrane conductance regulator is vital to sperm fertilizing capacity and male fertility. Proc. Natl. Acad. Sci. USA 2007, 104, 9816–9821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.-J.; Linsdell, P. Molecular mechanism of arachidonic acid inhibition of the CFTR chloride channel. Eur. J. Pharmacol. 2007, 563, 88–91. [Google Scholar] [CrossRef]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Tamburrino, L.; Marchiani, S.; Minetti, F.; Forti, G.; Muratori, M.; Baldi, E. The CatSper calcium channel in human sperm: Relation with motility and involvement in progesterone-induced acrosome reaction. Hum. Reprod. 2014, 29, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Mannowetz, N.; Miller, M.R.; Lishko, P.V. Regulation of the sperm calcium channel CatSper by endogenous steroids and plant triterpenoids. Proc. Natl. Acad. Sci. USA 2017, 114, 5743–5748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeld, A. Revisiting the action of steroids and triterpenoids on the human sperm Ca2+ channel CatSper. Mol. Hum. Reprod. 2020, 26, 816–824. [Google Scholar] [CrossRef]
- Antollini, S.S.; Barrantes, F.J. Fatty Acid Regulation of Voltage- and Ligand-Gated Ion Channel Function. Front. Physiol. 2016, 7, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Börjesson, S.I.; Hammarström, S.; Elinder, F. Lipoelectric Modification of Ion Channel Voltage Gating by Polyunsaturated Fatty Acids. Biophys. J. 2008, 95, 2242–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elinder, F.; Liin, S.I. Actions and Mechanisms of Polyunsaturated Fatty Acids on Voltage-Gated Ion Channels. Front. Physiol. 2017, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Moreno, C.; De La Cruz, A.; Valenzuela, C. In-Depth Study of the Interaction, Sensitivity, and Gating Modulation by PUFAs on K+ Channels; Interaction and New Targets. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Aursnes, M.; Hansen, T.V.; Tungen, J.E.; Galpin, J.D.; Leisle, L.; Ahern, C.A.; Xu, R.; Heinemann, S.H.; Hoshi, T. Atomic determinants of BK channel activation by polyunsaturated fatty acids. Proc. Natl. Acad. Sci. USA 2016, 113, 13905–13910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, T.; Xu, R.; Hou, S.; Heinemann, S.H.; Tian, Y. A point mutation in the human Slo1 channel that impairs its sensitivity to omega-3 docosahexaenoic acid. J. Gen. Physiol. 2013, 142, 507–522. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, T.; Tian, Y.; Xu, R.; Heinemann, S.H.; Hou, S. Mechanism of the modulation of BK potassium channel complexes with different auxiliary subunit compositions by the omega-3 fatty acid DHA. Proc. Natl. Acad. Sci. USA 2013, 110, 4822–4827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-Y.; Letts, J.A.; MacKinnon, R. Dimeric subunit stoichiometry of the human voltage-dependent proton channel Hv1. Proc. Natl. Acad. Sci. USA 2008, 105, 7692–7695. [Google Scholar] [CrossRef] [Green Version]
- Lowenthal, A.; Levy, R. Essential Requirement of Cytosolic Phospholipase A2for Activation of the H+ Channel in Phagocyte-like Cells. J. Biol. Chem. 1999, 274, 21603–21608. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.X.; Leaf, A. Evidence that free polyunsaturated fatty acids modify Na+ channels by directly binding to the channel proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 3542–3546. [Google Scholar] [CrossRef] [Green Version]
- Lishko, P.; Clapham, D.E.; Navarro, B.; Kirichok, Y. Sperm Patch-Clamp. Methods Enzymol. 2013, 525, 59–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, D.A.; Archer, S.L.; Weir, E.K. Accelerated disulfide reduction with polyunsaturated fatty acids: A mechanism of ionic channel modulation? Redox Rep. 1996, 2, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, K.; Eder, P.; Łykowska-Szuber, L.; Dobrowolska, A.; Krela-Kaźmierczak, I. Diet and Nutritional Factors in Male (In)fertility—Underestimated Factors. J. Clin. Med. 2020, 9, 1400. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Ion | Ion Channel | Sub Cellular Localization on Sperm | Function | Fatty Acids/ Lipids on Ion Channels | |
|---|---|---|---|---|---|
| Related Animal Studies | Related Human Studies | ||||
| Calcium | CatSper (cation channel of sperm) | Principal piece of the flagellum [84] | Calcium influx and activation of Ca-dependent hyperactivated motility [85] | N.A. | N.A. |
| Voltage gated Ca2+ channel (VGCC) | Neck and principal piece of the flagellum [18] | Mediating Ca2+ influx in response to action potential and subthreshold signals participating in sperm acrosome function [18] | 0.8 µM and 2.1 µM EPA inhibited the voltage gated L-type Ca2+ current by 50% in neonatal rat heart cell and in adult ventricular myocytes accordingly. Other PUFAs (DHA, AA, linolenic acid, linoleic acid, conjugaed linoleic acid, and eicosatetraynoic acid) had similar effects on calcium currents. MUFAs (Oleic acid) and SFA (Stearic acid) had no effect on L-type calcium current. [86] | N.A. | |
| N.A. | 8.5µM of AA caused 50% inhibition of L-type Ca2+ channel current in adult rat ventricular myocytes. [87] | N.A. | |||
| Transient receptor potential vanilloid (TRPV) | Sperm head, acrosome, neck and the flagellum, mostly in the flagellum of the human sperms [88] | Regulation of voltage gated Ca2+ influx and temperature reception [88] | N.A. | AA activated TRPA1, then ethanolamide or aminoacid/neurotransmitter derivatives in HEK293 expressing hTRPA1 cells. [89] | |
| AA activates the TRPV4 channel via epoxyeicosatrienoic acids in HEK293 cells. [90] | |||||
| Store-operated Ca2+ channel (SOCC) | Sperm head, neck and midpiece [91] | Supplying Ca2+ from extracellular environment in a voltage independent manner [92] | N.A. | N.A. | |
| Potassium | Slo1 | Sperm flagellum [31] | K+ efflux of the sperm [31] | N.A. | 10 µM arachidonic acid increases the open probability of BKca (Slo1) channel of human vascular smooth muscle cells (VSMCs) and HEK cells in the presence of the β1-subunit. [93] |
| cis FFA, palmitoleic, oleic, linoleic, linolenic, EPA increased Slo1 activity in GH3 cells. [94] | |||||
| Slo3 | Principal piece of the flagellum [7] | K+ efflux of the sperm [35] | N.A. | N.A. | |
| Voltage gated potassium channels | Principal piece of the flagellum [95] | K+ efflux [95] | Arachidonic acid (1–20 µM), cis-linoleic acid inhibit the Kv1.4 current in bovine zona fasciculata cells. Trans-PUFA (linoleladiac acid) and SFA (stearic acid) failed to inhibit the Kv1.4 current [96]. | 10 uM AA did not affect the Kv activity of VSMCs cells [93]. | |
| Extracellular application of long chain PUFAs (AA, DHA) inhibited the K+ current channels (Kv 1.5) in mouse and rat cardiomyocytes. [97] | AA acts as an open channel blocker for hKv1.5 channel of human cardiac cells. AA itself can interact with multiple amino acids located in the pore domain of the channel. [98] | ||||
| Physiological concentrations of DHA and AA (3–10µM) potently and irreversibly inhibited the Kv of rat olfactory receptor neurons. [99] | α-linolenic acid (ALA) blocked arterial specific Kv1.5 protein with Ic50 ~ 3.7µM. EPA and DHA reduced the steady state levels of the ion channel protein, but ALA did not. [100] | ||||
| Two outward K+ currents of ferret cardiomyocytes were inhibited by EPA and DHA whereas inwardly rectifying K+ current was unaffected. MUFAs and SFAs lacked the effect on K+ channels. [101] | N.A. | ||||
| Proton | Hv1 | Principal piece of the flagellum [38] | Proton extrusion resulting intracellular alkalization and activation of spermatozoa [38] | N.A. | AA increased the proton current through mouse Hv channel (mHv1) in heterologous expressed in HEK293T cells. Application of phospholipase 2 (PLA2) that generates AA from cell membrane phospholipids also stimulated the channel activity. (Similar to approx. 20µM AA) [102] |
| AA increased H+ selective conductance of human neutrophils [103] | |||||
| AA greatly enhanced the slowly activating H+ currents of murine macrophages. Effects of AA were not mediated by lipoxygenases (LOX) or cyclooxygenases (COX). The order of potency to stimulate the conductance was: AA > palmitoleate = palmitelaidate > linoleate > oleate > elaidate. Saturated fatty acids were inactive against Hv1. [104] | |||||
| The proton current of human eosinophils were augmented in the presence of 10µM AA. [105] | |||||
| AA enhanced the voltage gated proton conductance in human eosinophils. Upon activation by AA, activation was four times faster, H+ current amplitude was approximately five times higher, and gated voltage was shifted to more negative voltages. [106] | |||||
| Application of exogenous arachidonate can open the H+ channel of human cytoplasts in the absence of superoxide generation and the used concentrations of AA did not induce H+ permeability in liposomes of the cells. [107] | |||||
| Sodium | Voltage gated Na+ channel (VGNCs,NaV) | Principal piece, connecting piece, head, midpiece [41,42] | Maintenance of progressive motility [41,42] | PUFAs (DHA, EPA, linolenic acid, linoleic acid) inhibited the sodium current in dorsal rat ganglion cells in a dose dependent manner. Higher degree of unsaturation was resulted greater inhibition. MUFAs (Oleic acid, palmitoleic acid) and SFAs (Stearic acid, palmitic acid) inhibited the current to a lesser extent than PUFAs did. [108] | Eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) reduced the voltage gated Na+ current in human atrial myocytes in a concentration dependent manner. IC50 EPA-10.8 µM; DHA-41.2 µM [109] |
| Extracellular application of EPA (5 or 10 µM) caused an inhibition of voltage gated sodium currents of neonatal rat ventricular myocytes. The same concentrations of AA caused lesser inhibition. [110] | Both saturated and unsaturated fatty acids inhibited the current via α subunit of the human cardiac Na+ channel. But only PUFAs (EPA, DHA) inhibited the complete human myocardial Na+ channel. [111] | ||||
| AA had both inhibitory and activating effects on rNav1.4 depending on the depolarization potential. At −30 or −40 mV depolarization potential, AA activated the ion channel, but potential over −10 mv significantly inhibited the ion channel activity. [112] | |||||
| Epithelial Na+ channel (ENaC) | Flagellar midpiece [43] | Regulation of capacitation-associated hyperpolarization [43] | 2µM AA inhibited the ENaC activity by 50% in rat cortical collecting duct (CCD) cells. [113] | ||
| Chloride | Chloride channel-3 (ClC3) | Flagellum, neck, midpiece [46] | Regulation of sperm volume and motility [45] | N.A. | N.A. |
| Calcium activated chloride channels (CaCC) | Head [114] | Assist in Ca2+ dependent Cl- currents of the sperm [114] | N.A. | N.A. | |
| Cystic fibrosis transmembrane conductance regulator (CFTR) | Midpiece of the sperm [115] | Efflux of Cl- upon capacitation and mediating HCO3 entry [44,115] | AA inhibited the activity of CFTR chloride channels of baby hamster kidney (BHK) cells. AA may interact with the positively charged amino acid side chains in the cytoplasmic vestibule to block the channel. [116] | N.A. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooray, A.; Kim, J.H.; Chae, M.R.; Lee, S.; Lee, K.P. Perspectives on Potential Fatty Acid Modulations of Motility Associated Human Sperm Ion Channels. Int. J. Mol. Sci. 2022, 23, 3718. https://doi.org/10.3390/ijms23073718
Cooray A, Kim JH, Chae MR, Lee S, Lee KP. Perspectives on Potential Fatty Acid Modulations of Motility Associated Human Sperm Ion Channels. International Journal of Molecular Sciences. 2022; 23(7):3718. https://doi.org/10.3390/ijms23073718
Chicago/Turabian StyleCooray, Akila, Ji Hyun Kim, Mee Ree Chae, Sungwon Lee, and Kyu Pil Lee. 2022. "Perspectives on Potential Fatty Acid Modulations of Motility Associated Human Sperm Ion Channels" International Journal of Molecular Sciences 23, no. 7: 3718. https://doi.org/10.3390/ijms23073718