
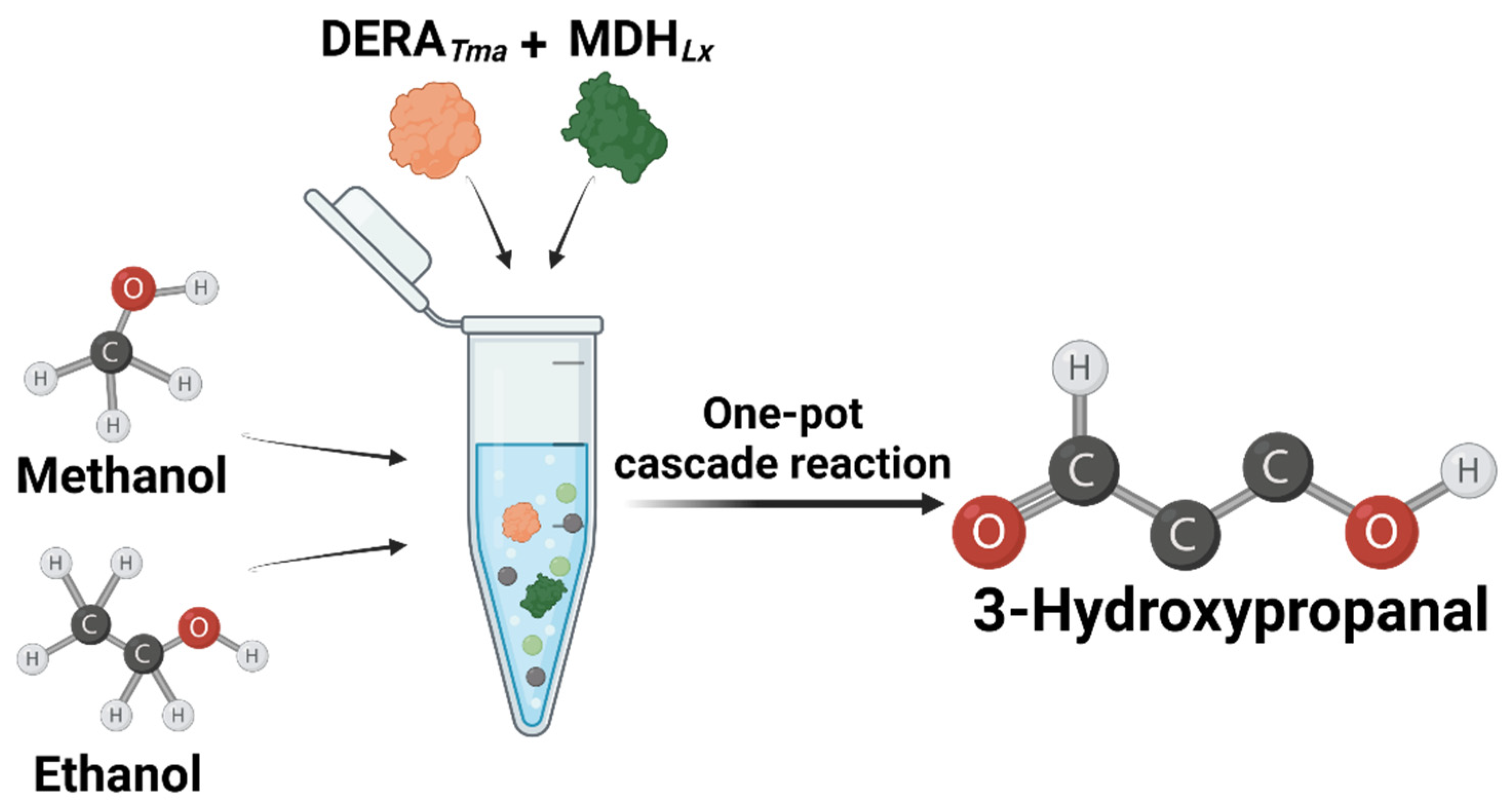
In Vitro One-Pot 3-Hydroxypropanal Production from Cheap C1 and C2 Compounds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
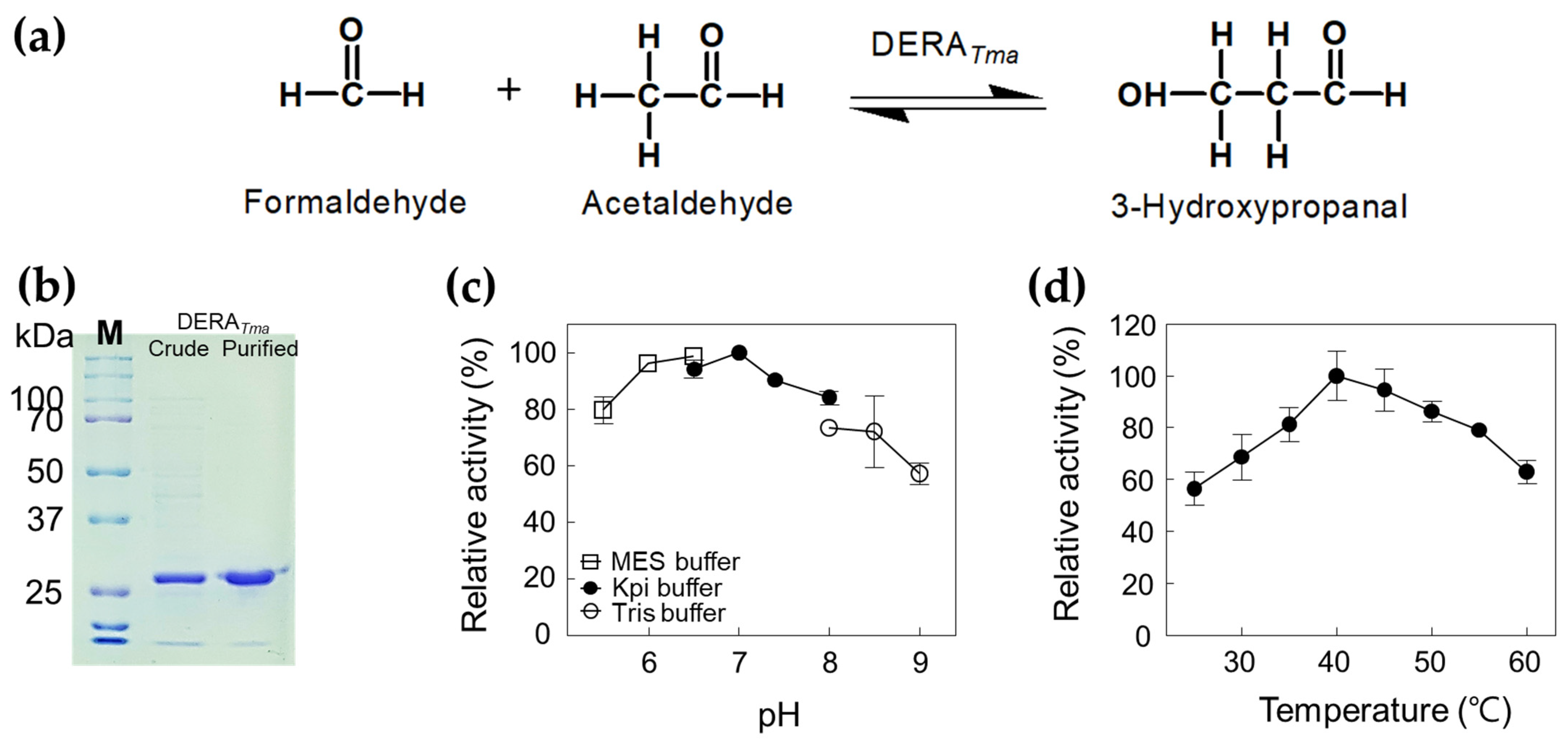
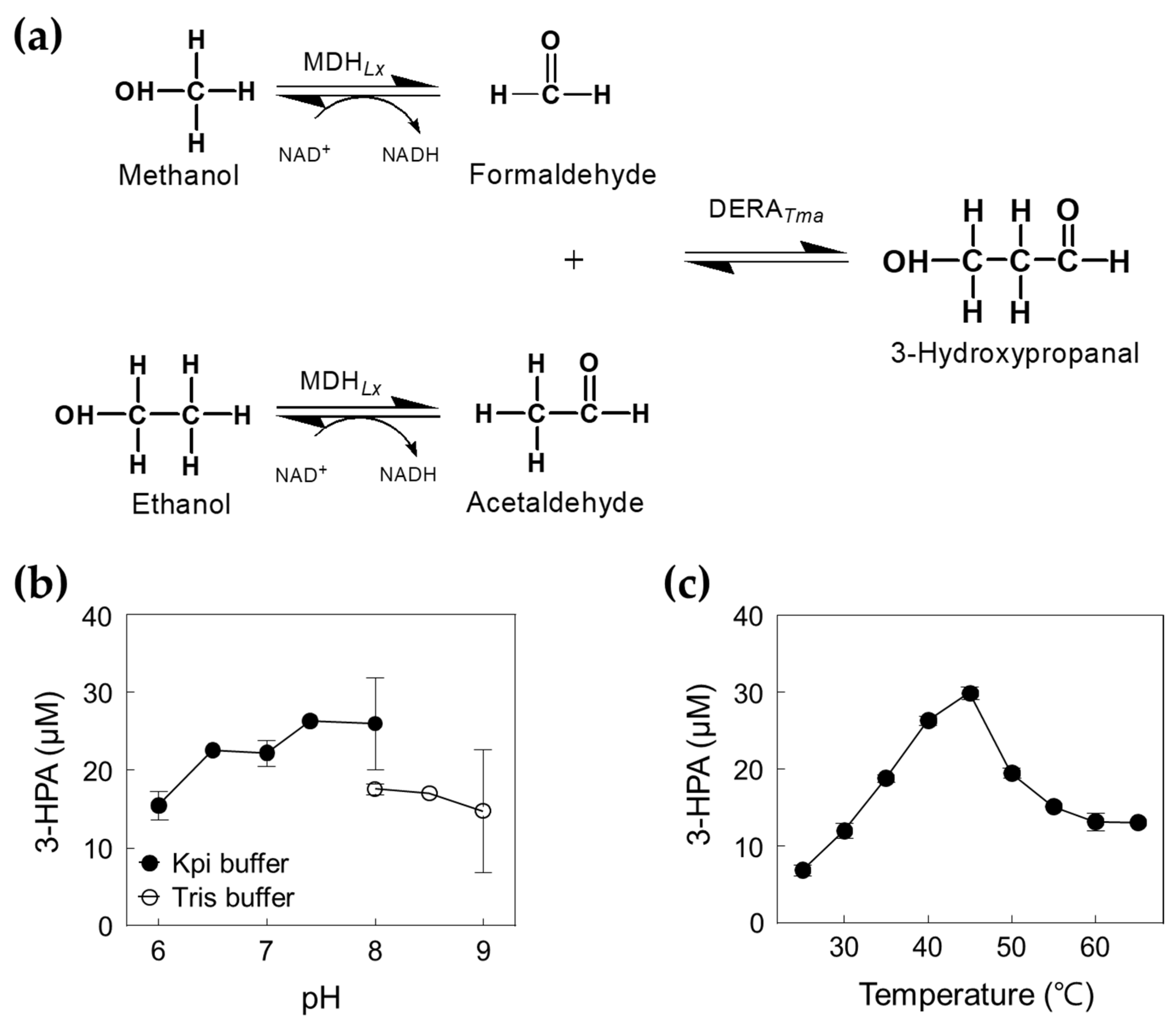
2.1. Characterization of DERATma
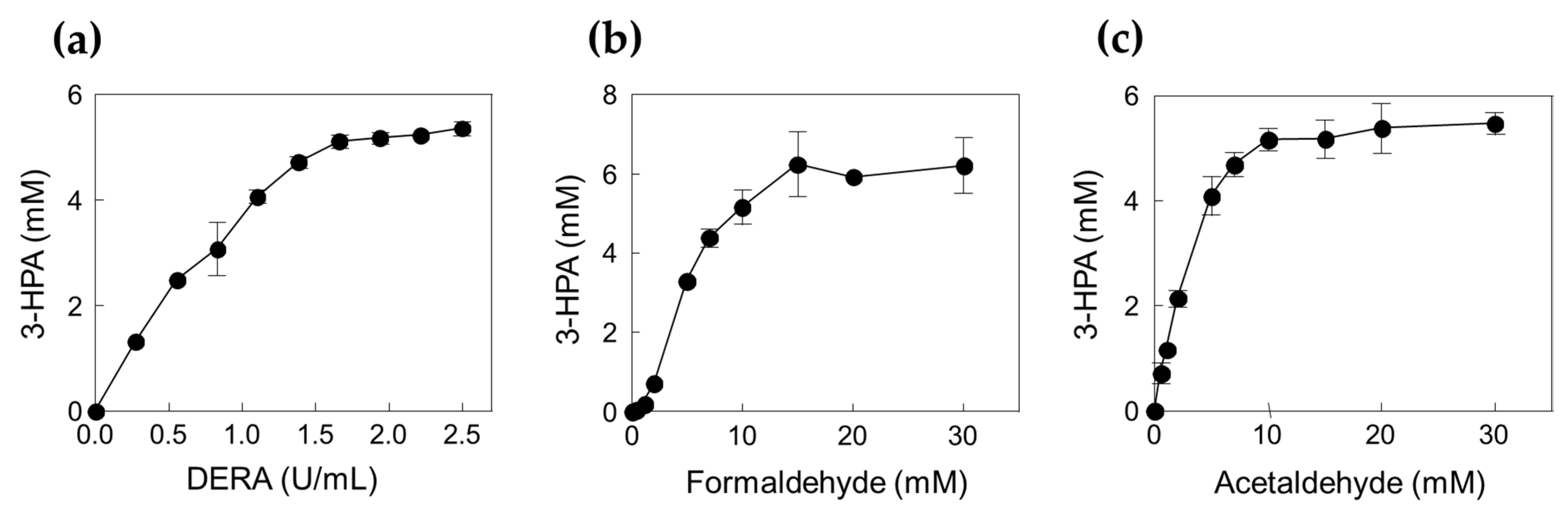
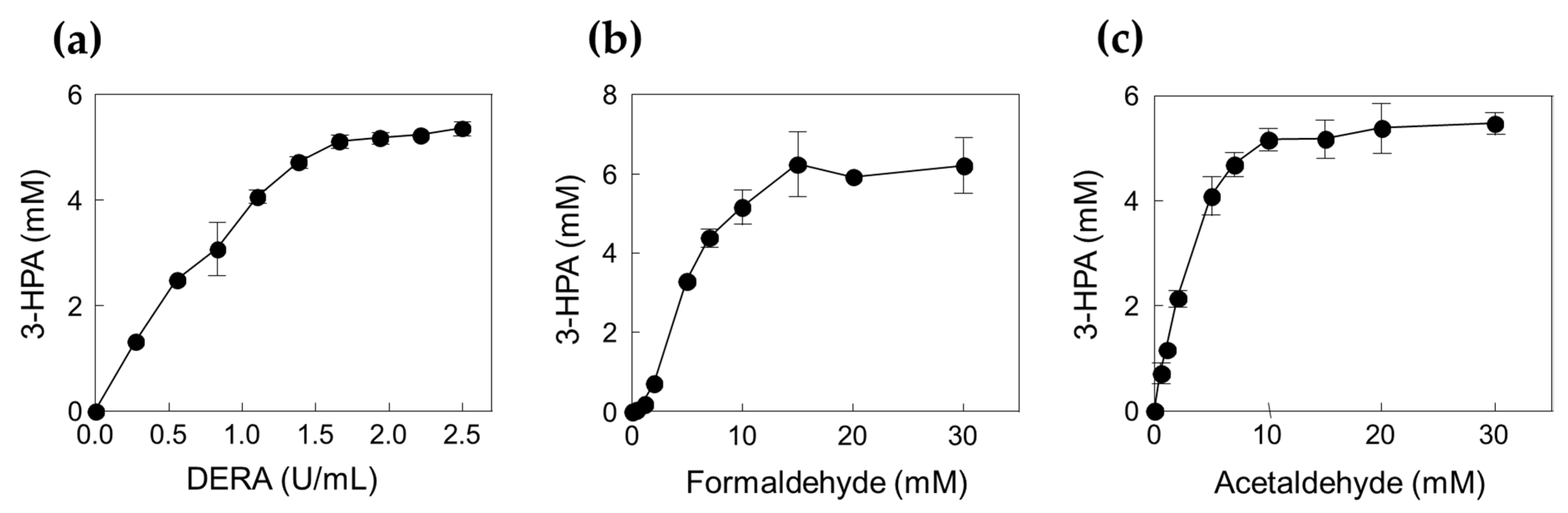
2.2. Production of 3-HPA from Formaldehyde and Acetaldehyde by DERATma
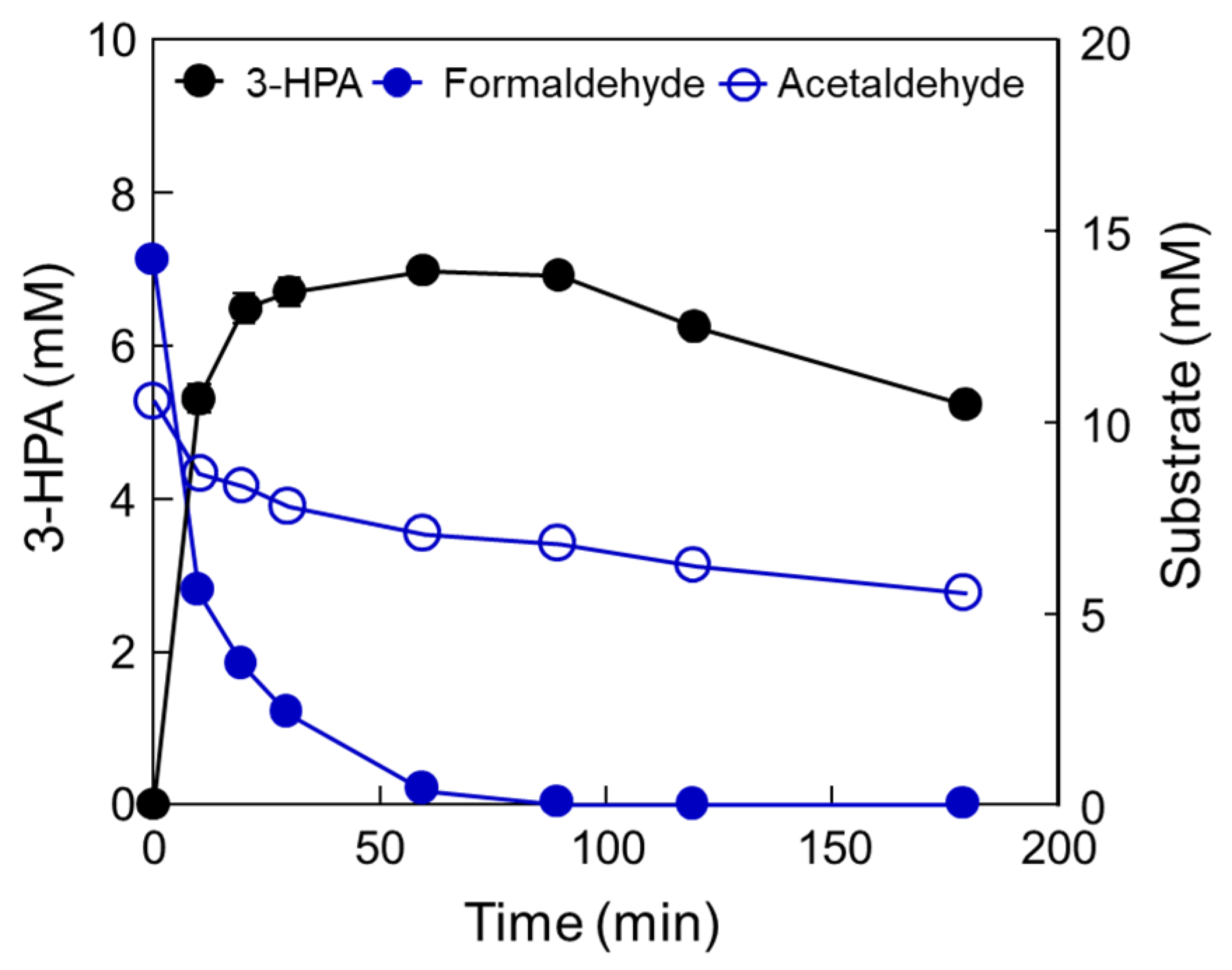
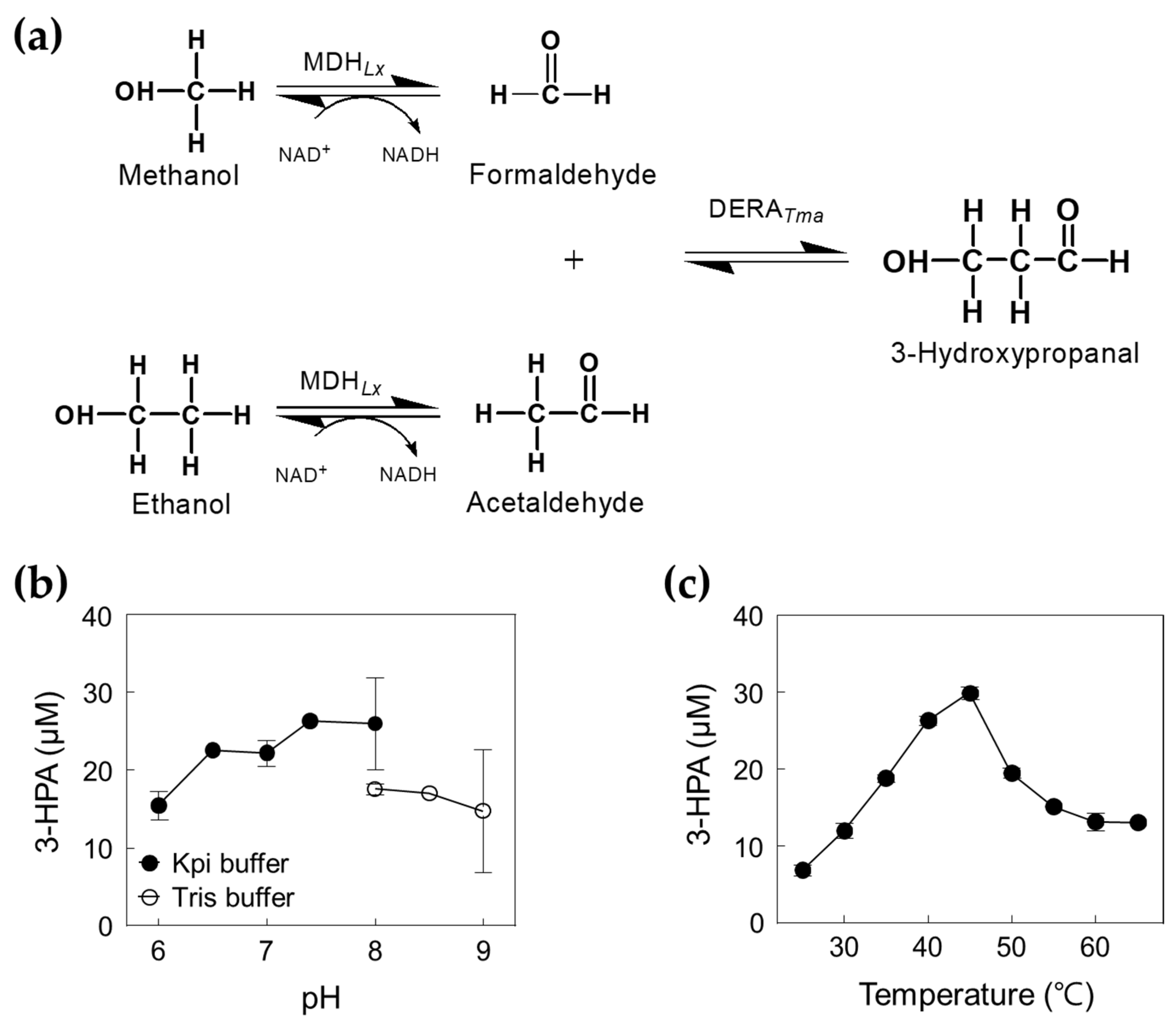
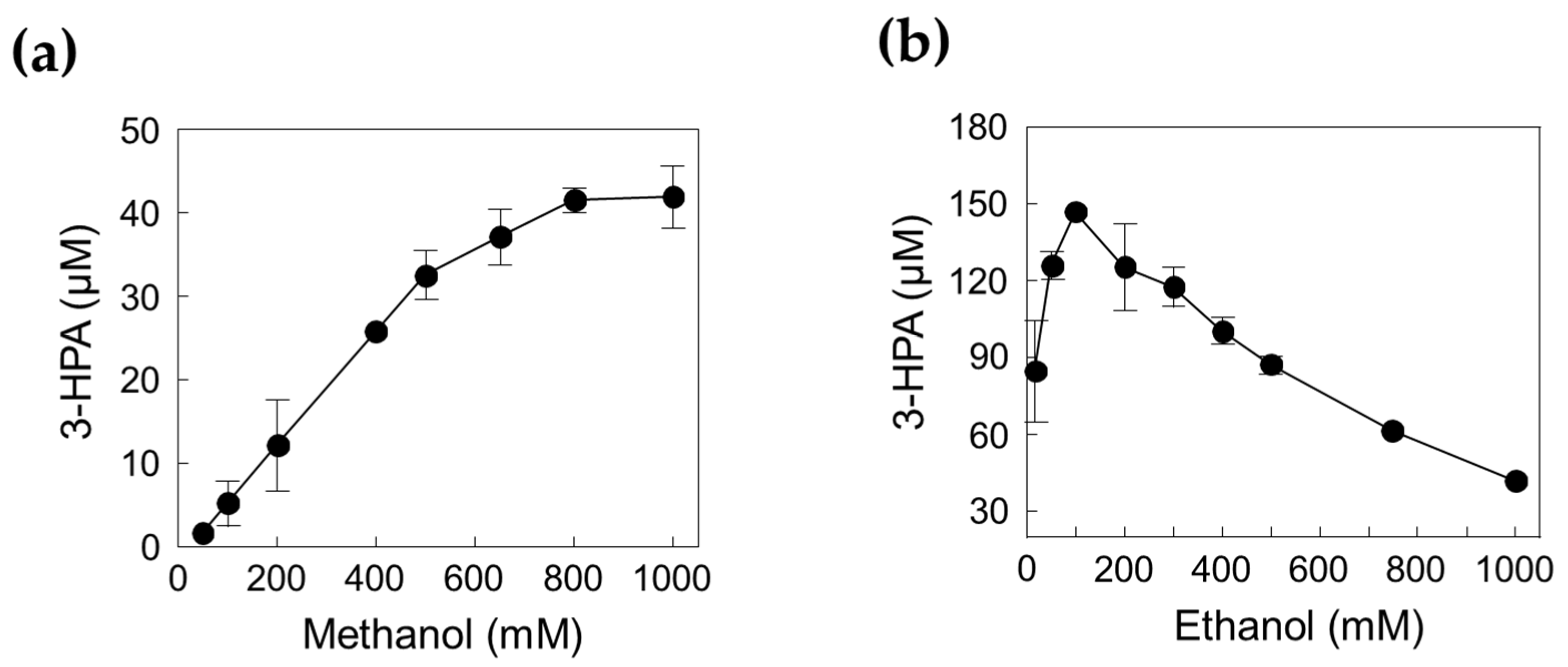
2.3. Optimization of the Cascade Reaction with DERATma and MDHLx
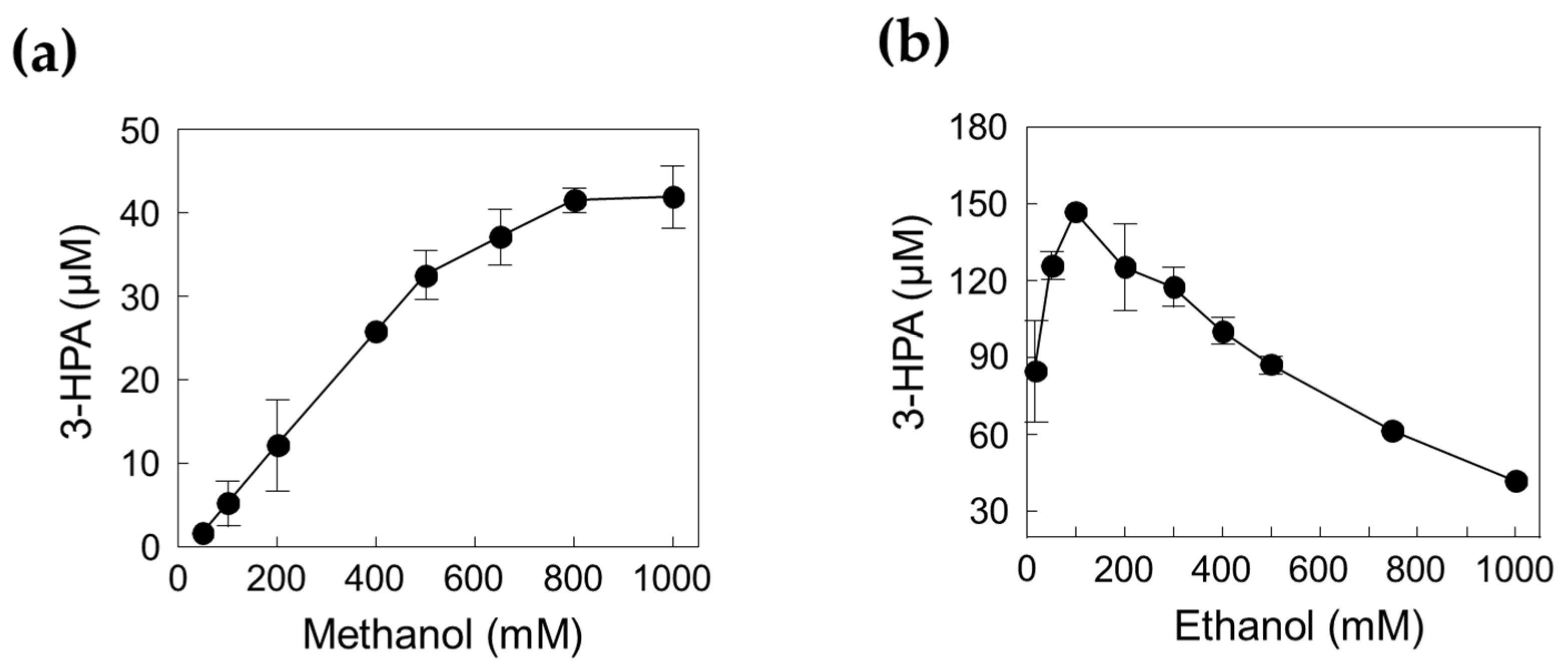
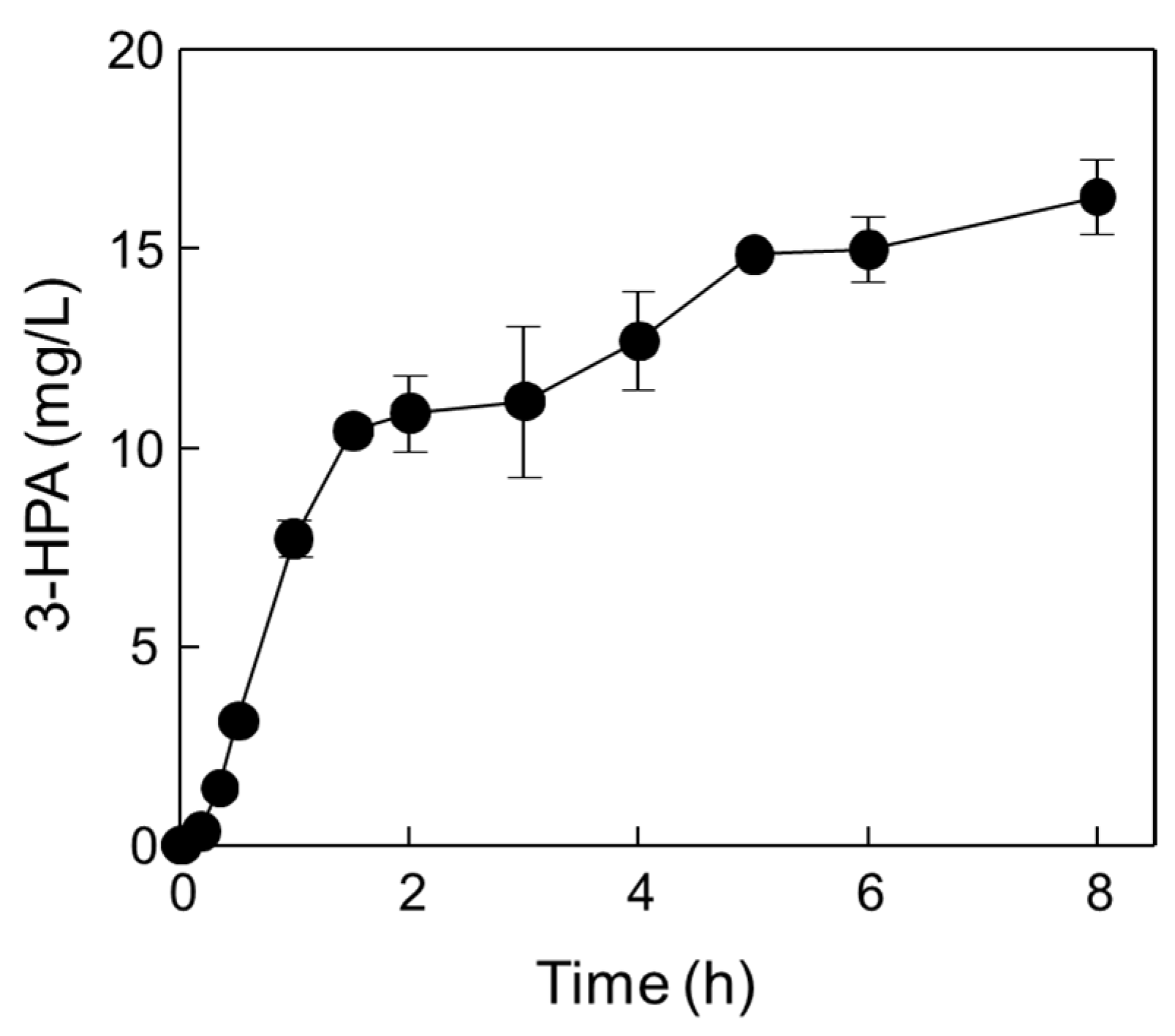
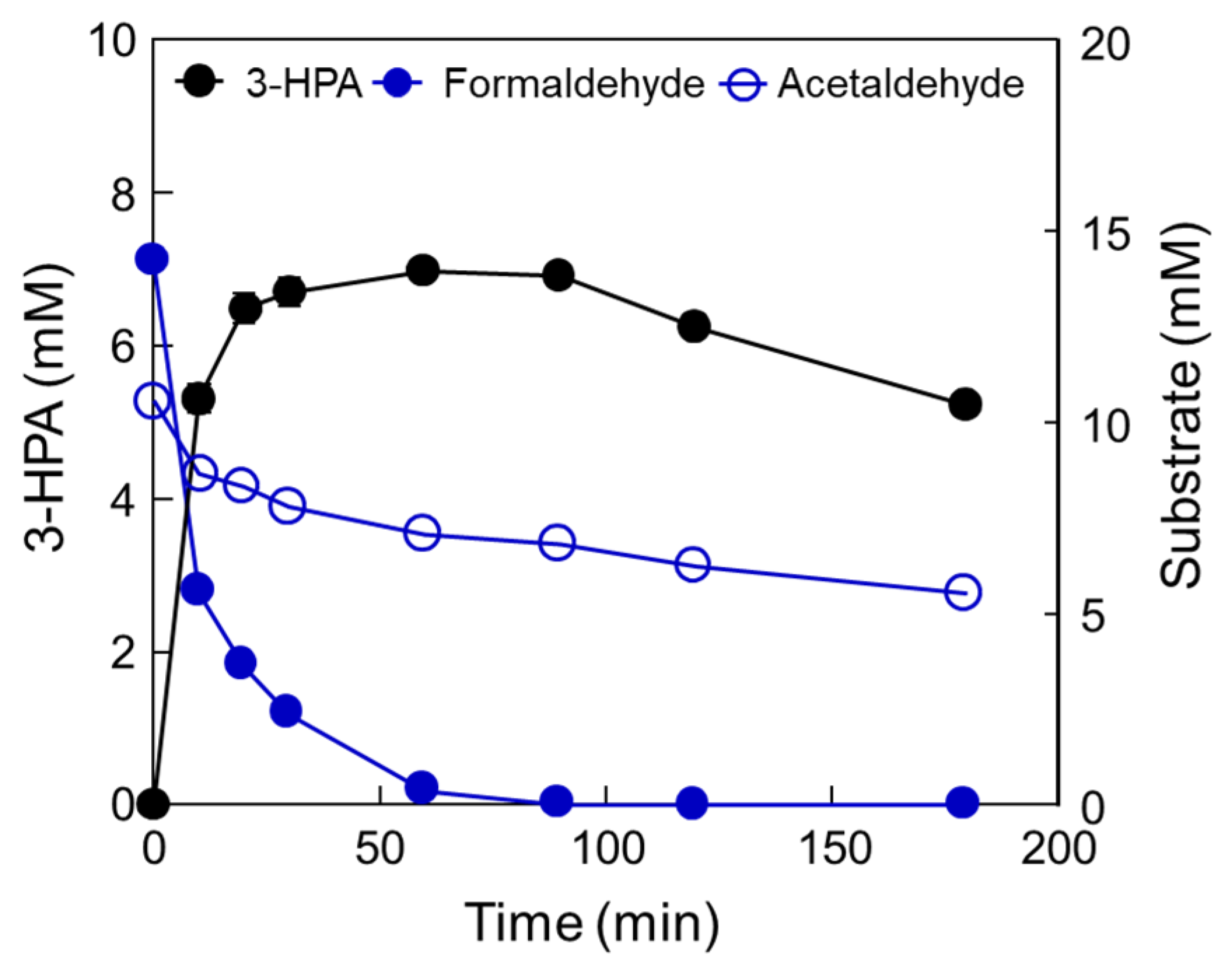
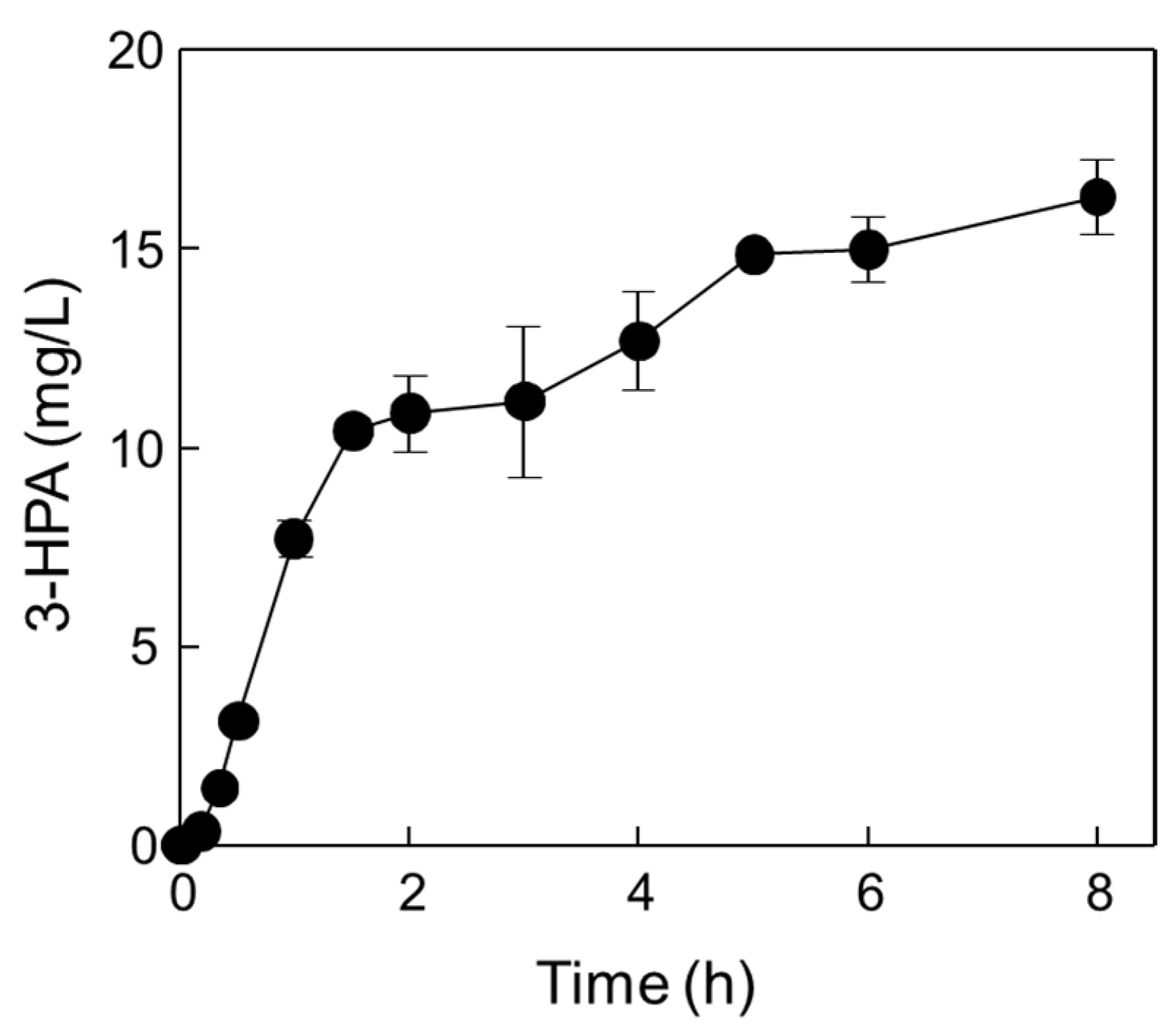
2.4. One-Pot 3-HPA Biosynthesis from Methanol and Ethanol
3. Materials and Methods
3.1. Chemicals and Materials
3.2. Gene Cloning
3.3. Site-Directed Mutagenesis of MDHLx
3.4. Culture Conditions
3.5. DERATma and MDHLx Purification
3.6. Enzyme Assay and Kinetic Parameters
3.7. Effects of pH and Temperature on DERATma Activity
3.8. Effects of Enzyme and Substrate Concentrations on 3-HPA Production
3.9. Optimization of Reaction Conditions for One-Pot 3-HPA Production from C1 and C2 Compounds
3.10. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, W.; Song, M.; Yang, Q.; Dai, Z.; Zhang, S.; Xin, F.; Dong, W.; Ma, J.; Jiang, M. Current advance in bioconversion of methanol to chemicals. Biotechnol. Biofuels 2018, 11, 260. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liang, H.; Panda, S.; Fung, V.K.Y.; Zhou, J.F.J.; Zhou, K. C2 feedstock-based biomanufacturing of value-added chemicals. Curr. Opin. Biotechnol. 2022, 73, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Schwach, P.; Pan, X.; Bao, X. Direct conversion of methane to value-added chemicals over heterogeneous catalysts: Challenges and prospects. Chem. Rev. 2017, 117, 8497–8520. [Google Scholar] [CrossRef]
- Ye, R.P.; Ding, J.; Gong, W.; Argyle, M.D.; Zhong, Q.; Wang, Y.; Russell, C.K.; Xu, Z.; Russell, A.G.; Li, Q.; et al. CO2 hydrogenation to high-value products via heterogeneous catalysis. Nat. Commun. 2019, 10, 5698. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, K.K.; Goswami, G.; Das, D. Biotransformation of methane and carbon dioxide into high-value products by methanotrophs: Current state of art and future prospects. Front. Microbiol. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Bornscheuer, U.T.; Huisman, G.W.; Kazlauskas, R.J.; Lutz, S.; Moore, J.C.; Robins, K. Engineering the third wave of biocatalysis. Nature 2012, 485, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; Brady, D. Broadening the scope of biocatalysis in sustainable organic synthesis. ChemSusChem 2019, 12, 2859–2881. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.K.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic synthesis for industrial applications. Angew. Chem. Int. Ed. 2021, 60, 88–119. [Google Scholar] [CrossRef]
- Zhang, W.M.; Zhang, T.; Wu, S.H.; Wu, M.K.; Xin, F.X.; Dong, W.L.; Ma, J.F.; Zhang, M.; Jiang, M. Guidance for engineering of synthetic methylotrophy based on methanol metabolism in methylotrophy. Rsc. Adv. 2017, 7, 4083–4091. [Google Scholar] [CrossRef] [Green Version]
- Le, T.K.; Lee, Y.J.; Han, G.H.; Yeom, S.J. Methanol dehydrogenases as a key biocatalysts for synthetic methylotrophy. Front. Bioeng. Biotechnol. 2021, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Le, T.K.; Ju, S.B.; Lee, H.W.; Lee, J.Y.; Oh, S.H.; Kwon, K.K.; Sung, B.H.; Lee, S.G.; Yeom, S.J. Biosensor-Based Directed Evolution of Methanol Dehydrogenase from Lysinibacillus xylanilyticus. Int. J. Mol. Sci. 2021, 22, 1471. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, S.H.; Oh, S.H.; Lee, J.J.; Kwon, K.K.; Kim, S.J.; Choi, M.; Rha, E.; Lee, H.; Lee, D.H.; et al. Discovery and biochemical characterization of a methanol dehydrogenase From Lysinibacillus xylanilyticus. Front. Bioeng. Biotechnol. 2020, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.Q.; Huang, B.; Wu, H.; Li, Z.M.; Ye, Q. Efficient 3-hydroxypropionic acid production from glycerol by metabolically engineered Klebsiella pneumoniae. Bioresour. Bioprocess. 2018, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Vollenweider, S.; Lacroix, C. 3-Hydroxypropionaldehyde: Applications and perspectives of biotechnological production. Appl. Microbiol. Biotechnol. 2004, 64, 16–27. [Google Scholar] [CrossRef]
- Rathnasingh, C.; Raj, S.M.; Jo, J.E.; Park, S. Development and evaluation of efficient recombinant Escherichia coli strains for the production of 3-hydroxypropionic acid from glycerol. Biotechnol. Bioeng. 2009, 104, 729–739. [Google Scholar] [CrossRef]
- Celinska, E. Debottlenecking the 1,3-propanediol pathway by metabolic engineering. Biotechnol. Adv. 2010, 28, 519–530. [Google Scholar] [CrossRef]
- Ko, Y.; Ashok, S.; Zhou, S.; Kumar, V.; Park, S. Aldehyde dehydrogenase activity is important to the production of 3-hydroxypropionic acid from glycerol by recombinant Klebsiella pneumoniae. Process. Biochem. 2012, 47, 1135–1143. [Google Scholar] [CrossRef]
- Kumar, V.; Ashok, S.; Park, S. Recent advances in biological production of 3-hydroxypropionic acid. Biotechnol. Adv. 2013, 31, 945–961. [Google Scholar] [CrossRef]
- Zhou, S.F.; Catherine, C.; Rathnasingh, C.; Somasundar, A.; Park, S. Production of 3-hydroxypropionic acid from glycerol by recombinant Pseudomonas denitrificans. Biotechnol. Bioeng. 2013, 110, 3177–3187. [Google Scholar] [CrossRef]
- Luo, L.H.; Seo, J.W.; Heo, S.Y.; Oh, B.R.; Kim, D.H.; Kim, C.H. Identification and characterization of Klebsiella pneumoniae aldehyde dehydrogenases increasing production of 3-hydroxypropionic acid from glycerol. Bioprocess. Biosyst. Eng. 2013, 36, 1319–1326. [Google Scholar] [CrossRef]
- Kim, K.; Kim, S.K.; Park, Y.C.; Seo, J.H. Enhanced production of 3-hydroxypropionic acid from glycerol by modulation of glycerol metabolism in recombinant Escherichia coli. Bioresour. Technol. 2014, 156, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.S.; Kim, Y.S.; Lee, C.M.; Lee, J.H.; Jung, W.S.; Ahn, J.H.; Song, S.H.; Choi, I.S.; Cho, K.M. Metabolic engineering of 3-hydroxypropionic acid biosynthesis in Escherichia coli. Biotechnol. Bioeng. 2015, 112, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Burge, G.; Flourat, A.L.; Pollet, B.; Spinnler, H.E.; Allais, F. 3-Hydroxypropionaldehyde (3-HPA) quantification by HPLC using a synthetic acrolein-free 3-hydroxypropionaldehyde system as analytical standard. RSC. Adv. 2015, 5, 96870. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.G.; Noh, M.H.; Jeong, J.H.; Park, S.; Jung, G.Y. Optimum rebalancing of the 3-hydroxypropionic acid production pathway from glycerol in Escherichia coli. Acs. Synth. Biol. 2016, 5, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Wang, C.; Yuan, Q.P.; Ren, J.; Zeng, A.P. An aldolase-based new pathway for bioconversion of formaldehyde and ethanol into 1,3-propanediol in Escherichia coli. Acs. Synth. Biol. 2021, 10, 799–809. [Google Scholar] [CrossRef]
- Taniguchi, H.; Okano, K.; Honda, K. Modules for in vitro metabolic engineering: Pathway assembly for bio-based production of value-added chemicals. Synth. Syst. Biotechnol. 2017, 2, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rashid, N.; Imanaka, H.; Fukui, T.; Atomi, H.; Imanaka, T. Presence of a novel phosphopentomutase and a 2-deoxyribose 5-phosphate aldolase reveals a metabolic link between pentoses and central carbon metabolism in the hyperthermophilic archaeon Thermococcus kodakaraensis. J. Bacteriol. 2004, 186, 4185–4191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.Y.; Chen, R.; Du, P.F.; Wu, H.L.; Pei, X.L.; Yang, B.; Yang, L.H.; Huang, L.F.; Liu, J.H.; Xie, T. Cloning and characterization of thermostable-deoxy-D-ribose-5-phosphate aldolase from Hyperthermus butylicus. Afr. J. Biotechnol. 2010, 9, 2898–2905. [Google Scholar]
- Sakuraba, H.; Yoneda, K.; Yoshihara, K.; Satoh, K.; Kawakami, R.; Uto, Y.; Tsuge, H.; Takahashi, K.; Hori, H.; Ohshima, T. Sequential aldol condensation catalyzed by hyperthermophilic 2-deoxy-D-ribose-5-phosphate aldolase. Appl. Environ. Microbiol. 2007, 73, 7427–7434. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Chang, Y.H.; Choi, N.S.; Kim, Y.; Song, J.J.; Kim, J.S. Cloning, expression, and characterization of a new deoxyribose 5-phosphate aldolase from Yersinia sp. EA015. Protein. Expr. Purif. 2009, 68, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Choi, N.S.; Kim, Y.O.; Son, D.H.; Chang, Y.H.; Song, J.J.; Kim, J.S. Expression and characterization of a novel deoxyribose 5-phosphate aldolase from Paenibacillus sp. EA001. J. Microbiol. Biotechnol. 2010, 20, 995–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshida, T.; Hayashi, J.; Satomura, T.; Kawakami, R.; Ohshima, T.; Sakuraba, H. First characterization of extremely halophilic 2-deoxy-D-ribose-5-phosphate aldolase. Protein. Expr. Purif. 2016, 126, 62–68. [Google Scholar] [CrossRef]
- Yin, X.P.; Wang, Q.Y.; Zhao, S.J.; Du, P.F.; Xie, K.L.; Jin, P.; Xie, T. Cloning and characterization of a thermostable 2-deoxy-D-ribose-5-phosphate aldolase from Aciduliprofundum boonei. Afr. J. Biotechnol. 2011, 10, 16260–16266. [Google Scholar] [CrossRef]
- Kullartz, I.; Pietruszka, J. Cloning and characterisation of a new 2-deoxy-D-ribose-5-phosphate aldolase from Rhodococcus erythropolis. J. Biotechnol. 2012, 161, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.H.; Kim, M.S.; Chung, N.; Kim, J.S. Expression and characterization of a novel 2-deoxyribose-5-phosphate aldolase from Haemophilus influenzae Rd KW20. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 655–660. [Google Scholar] [CrossRef]
- He, F.F.; Xin, Y.Y.; Ma, Y.X.; Yang, S.; Fei, H. Rational design to enhance the catalytic activity of 2-deoxy-D-ribose-5-phosphate aldolase from Pseudomonas syringae pv. syringae B728a. Protein Expr. Purif. 2021, 183, 105863. [Google Scholar] [CrossRef] [PubMed]
- Groth, D.P. Deoxyribose 5-phosphate aldolase. II. Purification and properties of the rat liver enzyme. J. Biol. Chem. 1967, 242, 155–159. [Google Scholar] [CrossRef]
- Osen, O.M.; Hoffee, P.; Horecker, B.L. The Mechanism of action of aldolases. Vii. formation of a 2-methyl-2-deoxypentose catalyzed by deoxyribose 5-phosphate aldolase. J. Biol. Chem. 1965, 240, 1517–1524. [Google Scholar]
- Chen, L.; Dumas, D.P.; Wong, C.H. Deoxyribose 5-phosphate aldolase as a catalyst in asymmetric aldol condensation. J. Am. Chem. Soc. 1992, 114, 741–748. [Google Scholar] [CrossRef]
- Hoffee, P.A. 2-deoxyribose-5-phosphate aldolase of Salmonella typhimurium: Purification and properties. Arch. Biochem. Biophys. 1968, 126, 795–802. [Google Scholar] [CrossRef]
- Sankaranarayanan, M.; Somasundar, A.; Seol, E.; Chauhan, A.S.; Kwon, S.; Jung, G.Y.; Park, S. Production of 3-hydroxypropionic acid by balancing the pathway enzymes using synthetic cassette architecture. J. Biotechnol. 2017, 259, 140–147. [Google Scholar] [CrossRef]
- Sobolov, M.; Smiley, K.L. Metabolism of glycerol by an acrolein-forming lactobacillus. J. Bacteriol. 1960, 79, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Smiley, K.L.; Sobolov, M. A cobamide-requiring glycerol dehydrase from an acrolein-forming Lactobacillus. Arch. Biochem. Biophys. 1962, 97, 538–543. [Google Scholar] [CrossRef]
- Slininger, P.J.; Bothast, R.J.; Smiley, K.L. Production of 3-hydroxypropionaldehyde from glycerol. Appl. Environ. Microbiol. 1983, 46, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talarico, T.L.; Casas, I.A.; Chung, T.C.; Dobrogosz, W.J. Production and isolation of reuterin, a growth inhibitor produced by Lactobacillus reuteri. Antimicrob. Agents Chemother. 1988, 32, 1854–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthi-Peng, Q.; Dileme, F.B.; Puhan, Z. Effect of glucose on glycerol bioconversion by Lactobacillus reuteri. Appl. Microbiol. Biotechnol. 2002, 59, 289–296. [Google Scholar] [CrossRef]
- Luthi-Peng, Q.; Scharer, S.; Puhan, Z. Production and stability of 3-hydroxypropionaldehyde in Lactobacillus reuteri. Appl. Microbiol. Biotechnol. 2002, 60, 73–80. [Google Scholar] [CrossRef]
- Vollenweider, S.; Grassi, G.; Konig, I.; Puhan, Z. Purification and structural characterization of 3-hydroxypropionaldehyde and its derivatives. J. Agric. Food Chem. 2003, 51, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Slininger, P.J.; Bothast, R.J. Optimizing aerobic conversion of glycerol to 3-hydroxypropionaldehyde. Appl. Environ. Microbiol. 1985, 50, 1444–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancauwenberge, J.E.; Slininger, P.J.; Bothast, R.J. Bacterial conversion of glycerol to beta-hydroxypropionaldehyde. Appl. Environ. Microbiol. 1990, 56, 329–332. [Google Scholar] [CrossRef] [Green Version]
- Barbirato, F.; Grivet, J.P.; Soucaille, P.; Bories, A. 3-Hydroxypropionaldehyde, an inhibitory metabolite of glycerol fermentation to 1,3-propanediol by enterobacterial species. Appl. Environ. Microbiol. 1996, 62, 1448–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, Y.; Du, C.; Liu, M.; Cao, Z. Inactivation of aldehyde dehydrogenase: A key factor for engineering 1,3-propanediol production by Klebsiella pneumoniae. Metab. Eng. 2006, 8, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.Y.; Seo, J.W.; Heo, S.Y.; Baek, J.O.; Rairakhwada, D.; Oh, B.R.; Seo, P.S.; Choi, M.H.; Kim, C.H. Elimination of by-product formation during production of 1,3-propanediol in Klebsiella pneumoniae by inactivation of glycerol oxidative pathway. Appl. Microbiol. Biotechnol. 2009, 84, 527–534. [Google Scholar] [CrossRef]
- Ju, J.H.; Jeon, S.G.; Lee, K.M.; Heo, S.Y.; Kim, M.S.; Kim, C.H.; Oh, B.R. The biocatalytic production of 3-hydroxypropionaldehyde and evaluation of its stability. Catalysts 2021, 11, 1139. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, S.-B.; Seo, M.-J.; Yeom, S.-J. In Vitro One-Pot 3-Hydroxypropanal Production from Cheap C1 and C2 Compounds. Int. J. Mol. Sci. 2022, 23, 3990. https://doi.org/10.3390/ijms23073990
Ju S-B, Seo M-J, Yeom S-J. In Vitro One-Pot 3-Hydroxypropanal Production from Cheap C1 and C2 Compounds. International Journal of Molecular Sciences. 2022; 23(7):3990. https://doi.org/10.3390/ijms23073990
Chicago/Turabian StyleJu, Su-Bin, Min-Ju Seo, and Soo-Jin Yeom. 2022. "In Vitro One-Pot 3-Hydroxypropanal Production from Cheap C1 and C2 Compounds" International Journal of Molecular Sciences 23, no. 7: 3990. https://doi.org/10.3390/ijms23073990
APA StyleJu, S.-B., Seo, M.-J., & Yeom, S.-J. (2022). In Vitro One-Pot 3-Hydroxypropanal Production from Cheap C1 and C2 Compounds. International Journal of Molecular Sciences, 23(7), 3990. https://doi.org/10.3390/ijms23073990