
Regulatory Processes of the Canonical Wnt/β-Catenin Pathway and Photobiomodulation in Diabetic Wound Repair


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
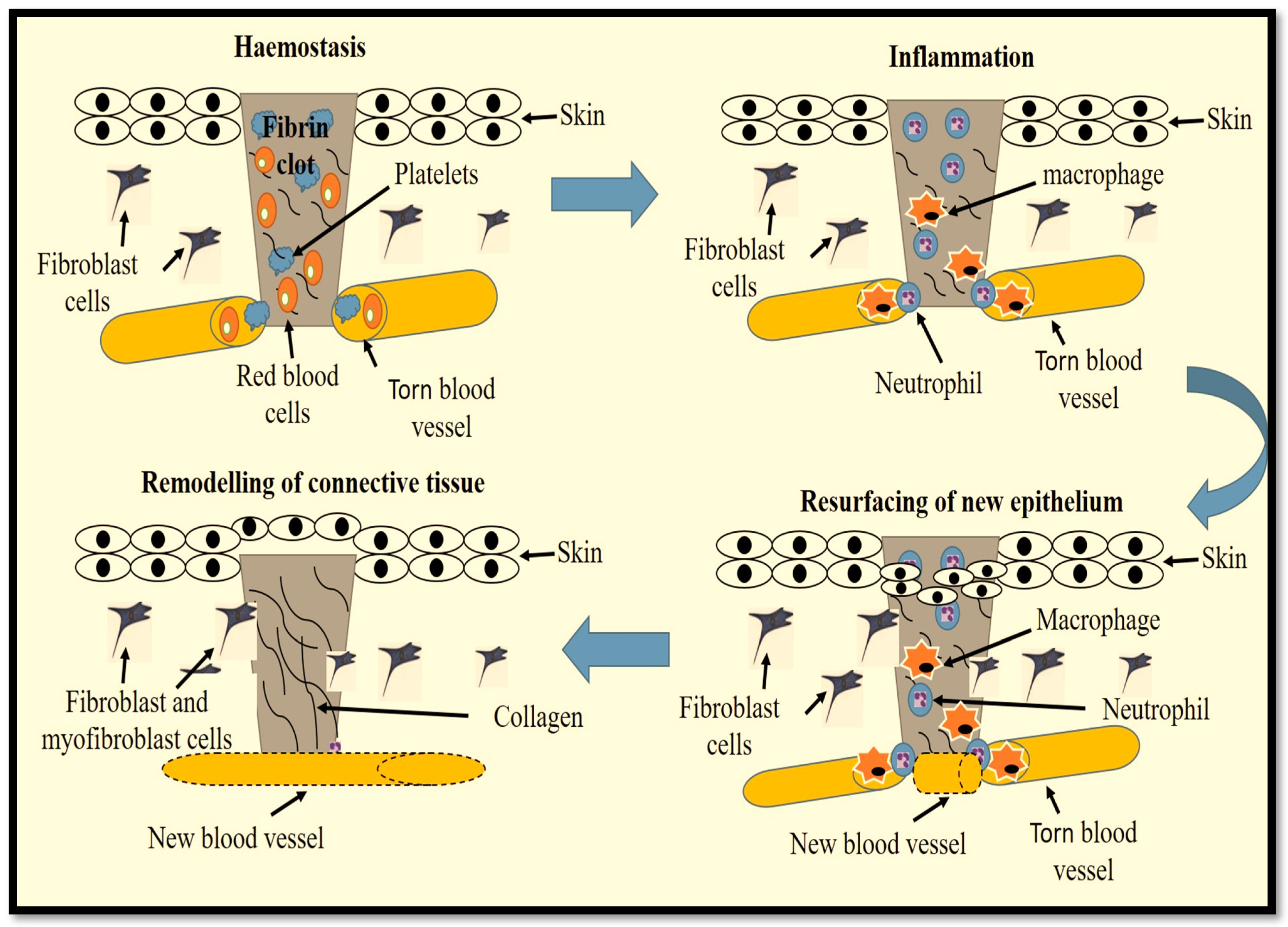
2. Intracellular Signalling in Wound Healing
2.1. Wnt/β-Catenin Pathway in Wound Healing
2.2. Regulation of the Wnt/β-Catenin Pathway in Diabetic Wound Healing
3. Photobiomodulation (PBM) and the Activation of Signalling Pathways in Diabetic Wound Healing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bielefeld, K.A.; Amini-Nik, S.; Alman, B.A. Cutaneous wound healing: Recruiting developmental pathways for regeneration. Cell. Mol. Life Sci. 2013, 70, 2059–2081. [Google Scholar]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265–266. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223. [Google Scholar] [CrossRef] [PubMed]
- Rosińczuk, J.; Taradaj, J.; Dymarek, R.; Sopel, M. Mechanoregulation of Wound Healing and Skin Homeostasis. BioMed Res. Int. 2016, 2016, 3943481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Shu, B.; Yang, R.; Xu, Y.; Xing, B.; Liu, J.; Chen, L.; Qi, S.; Liu, X.; Wang, P.; et al. Wnt and Notch signaling pathway involved in wound healing by targeting c-Myc and Hes1 separately. Stem Cell Res Ther. 2015, 6, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Nie, X.; Shi, X.; Zhao, J.; Chen, Y.; Yao, Q.; Sun, C.; Yang, J. Regulatory Mechanisms of the Wnt/β-Catenin Pathway in Diabetic Cutaneous Ulcers. Front. Pharmacol. 2018, 9, 1114. [Google Scholar] [CrossRef]
- Qing, C. The molecular biology in wound healing & nonhealing wound. Chin. J. Traumatol. 2017, 20, 189–193. [Google Scholar]
- Lin, B.S.; Chang, C.C.; Su, C.L.; Li, J.R.; Chen, M.L.; Chen, M.Y.; Huang, Y.K. The assessment of Buerger’s exercise on dorsal foot skin circulation in patients with vasculopathic diabetic foot ulcer by using wireless near-infrared spectroscope: A cohort prospective study. Lasers Med. Sci. 2018, 33, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Rigato, M.; Pizzol, D.; Tiago, A.; Putoto, G.; Avogaro, A.; Fadini, G.P. Characteristics, prevalence, and outcomes of diabetic foot ulcers in Africa. A systemic review and meta-analysis. Diabetes Res. Clin. Pract. 2018, 142, 63–73. [Google Scholar] [PubMed]
- Zhao, Y.; Ming, L.; Wei, Z.; Bin, W.; Yudong, Z.; Haiwen, S.U.; Xiaolan, R.E.N.; Qingzhi, H.A.O. Changes in the expression of Wnt/β-catenin signaling pathway in diabetic ulcers. Chin. J. Pathophysiol. 2015, 17, 2033–2038. [Google Scholar]
- Everett, E.; Mathioudakis, N. Update on management of diabetic foot ulcers. Ann. N. Y. Acad. Sci. 2018, 1, 153–165. [Google Scholar]
- Khan, I.; Arany, P. Biophysical Approaches for Oral Wound Healing: Emphasis on Photobiomodulation. Adv. Wound Care 2015, 12, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, S.Y.; Tam, V.; Ramkumar, S.; Khaw, M.L.; Law, H.; Lee, S. Review on the Cellular Mechanisms of Low-Level Laser Therapy Use in Oncology. Front. Oncol. 2020, 10, 1255. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.; Salman, B.; Razak, N.; Samsudin, A.A. Comparative Clinical Study between Concentrated Growth Factor and Low-Level Laser Therapy in the Management of Dry Socket. Eur. J. Dent. 2020, 14, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J. Cellular signaling in tissue regeneration. Yonsei Med. J. 2000, 6, 692–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghilardi, S.J.; O’Reilly, B.M.; Sgro, A.E. Intracellular signaling dynamics and their role in coordinating tissue repair. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 3, e1479. [Google Scholar]
- Bodnar, R.J. Epidermal Growth Factor and Epidermal Growth Factor Receptor: The Yin and Yang in the Treatment of Cutaneous Wounds and Cancer. Adv. Wound Care 2013, 1, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Klass, B.R.; Grobbelaar, A.O.; Rolfe, K.J. Transforming growth factor β1 signalling, wound healing and repair: A multifunctional cytokine with clinical implications for wound repair, a delicate balance. Postgrad. Med. J. 2009, 85, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, L.; Zheng, Z.; Li, Z.; Shi, J.; Li, Y.; Han, S.; Gao, J.; Tang, C.; Su, L.; et al. Src promotes cutaneous wound healing by regulating MMP-2 through the ERK pathway. Int. J. Mol. Med. 2016, 3, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepe, L.; Ferrari, M.C.; Cantarella, C.; Fioretti, F.; Paolella, G. Ras activated ERK and PI3K pathways differentially affect directional movement of cultured fibroblasts. Cell. Physiol. Biochem. 2013, 1, 123–142. [Google Scholar] [CrossRef]
- Koivisto, L.; Heino, J.; Häkkinen, L.; Larjava, H. Integrins in Wound Healing. Adv. Wound Care 2014, 12, 762–783. [Google Scholar] [CrossRef] [Green Version]
- Chigurupati, S.; Arumugam, T.V.; Son, T.G.; Lathia, J.D.; Jameel, S.; Mughal, M.R.; Tang, S.; Jo, D.G.; Camandola, S.; Giunta, M.; et al. Involvement of notch signaling in wound healing. PLoS ONE 2007, 2, e1167. [Google Scholar] [CrossRef]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Wei, X.; Ma, H.; Fan, L.; Chen, W.D. The complex role of Wnt ligands in type 2 diabetes mellitus and related complications. J. Cell. Mol. Med. 2021, 25, 6479–6495. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 1, 9–26. [Google Scholar]
- Nie, X.; Liu, H.; Liu, L.; Wang, Y.D.; Chen, W.D. Emerging Roles of Wnt Ligands in Human Colorectal Cancer. Front. Oncol. 2020, 10, 1341. [Google Scholar] [PubMed]
- Tamura, M.; Nemoto, E. Role of the Wnt signaling molecules in the tooth. Jpn. Dent. Sci. Rev. 2016, 52, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.J.; Tao, R.; Han, Y.Q.; Xu, G.; Liu, J.; Han, Y.F. Therapeutic potential of umbilical cord mesenchymal stem cells with Wnt/β-catenin signaling pathway pre-activated for the treatment of diabetic wounds. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2460–2464. [Google Scholar] [PubMed]
- Bastakoty, D.; Young, P.P. Wnt/β-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. FASEB 2016, 10, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, Y.; Sun, C.; Wang, T.; Shen, Y.; Cai, W.; Sun, J.; Chi, L.; Wang, H.; Song, N.; et al. Feedback Activation of Basic Fibroblast Growth Factor Signaling via the Wnt/β-Catenin Pathway in Skin Fibroblasts. Front. Pharmacol. 2017, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; He, Y.; Li, W.; Li, H. Wnt/β-catenin interacts with the FGF pathway to promote proliferation and regenerative cell proliferation in the zebrafish lateral line neuromast. Exp. Mol. Med. 2019, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, S.; Hayashida, K. Advances in surgical applications of growth factors for wound healing. Burn Trauma 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Lauer, G.; Sollberg, S.; Cole, M.; Flamme, I.; Stürzebecher, J.; Mann, K.; Krieg, T.; Eming, S.A. Expression and proteolysis of vascular endothelial growth factor is increased in chronic wounds. J. Investig. Dermatol. 2000, 115, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Haughey, M.; Wang, N.Y.; Blease, K.; Kapoun, A.M.; Couto, S.; Belka, I.; Hoey, T.; Groza, M.; Hartke, J.; et al. Targeting the Wnt signaling pathway through R-spondin 3 identifies an anti-fibrosis treatment strategy for multiple organs. PLoS ONE 2020, 3, e0229445. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.N.; Dash, N.R.; Guru, B.; Mohapatra, P.C. Towards reaching the target: Clinical application of mesenchymal stem cells for diabetic foot ulcers. Rejuvenation Res. 2014, 1, 40–53. [Google Scholar] [CrossRef]
- Hoke, G.D.; Ramos, C.; Hoke, N.N.; Crossland, M.C.; Shawler, L.G.; Boykin, J.V. Atypical diabetic foot ulcer keratinocyte protein signaling correlates with impaired wound healing. J. Diabetes Res. 2016, 2016, 1586927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.F.; Lin, Y.N.; Liu, K.F.; Wang, C.T.; Ramachandran, S.; Wang, C.J.; Kuo, Y. The Acceleration of Diabetic Wound Healing by Low-Intensity Extracorporeal Shockwave Involves in the GSK-3_ Pathway. Biomedicines 2021, 9, 21. [Google Scholar] [CrossRef]
- Lin, C.L.; Wang, J.Y.; Huang, Y.T.; Kuo, Y.H.; Surendran, K.; Wang, F.S. Wnt/beta-catenin signaling modulates survival of high glucose-stressed mesangial cells. J. Am. Soc. Nephrol. 2006, 17, 2812–2820. [Google Scholar] [CrossRef]
- Kuffler, D.P. Photobiomodulation in promoting wound healing: A review. Regen. Med. 2016, 1, 107–122. [Google Scholar] [CrossRef]
- Zhao, M.; Penninger, J.; Isseroff, R.R. Electrical Activation of Wound-Healing Pathways. Adv. Skin Wound Care 2010, 1, 567–573. [Google Scholar]
- Santana-Blank, L.; Rodríguez-Santana, E.; Santana-Rodríguez, K.E.; Reyes, H. Quantum Leap in Photobiomodulation Therapy Ushers in a New Generation of Light-Based Treatments for Cancer and Other Complex Diseases: Perspective and Mini-Review. Photomed. Laser Surg. 2016, 3, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosca, R.C.; Ong, A.A.; Albasha, O.; Bass, K.; Arany, P. Photobiomodulation Therapy for Wound Care: A Potent, Noninvasive, Photoceutical Approach. Adv. Skin Wound Care 2019, 4, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Courtois, E.; Bouleftour, W.; Guy, J.B.; Louati, S.; Bensadoun, R.J.; Rodriguez-Lafrasse, C.; Magné, N. Mechanisms of PhotoBioModulation (PBM) focused on oral mucositis prevention and treatment: A scoping review. BMC Oral Health 2021, 21, 220. [Google Scholar] [CrossRef] [PubMed]
- De Castro, J.R.; da Silva Pereira, F.; Chen, L.; Arana-Chavez, V.E.; Ballester, R.Y.; DiPietro, L.A.; Simões, A. Improvement of full-thickness rat skin wounds by photobiomodulation therapy (PBMT): A dosimetric study. J. Photochem. Photobiol. B 2020, 206, 111850. [Google Scholar] [CrossRef]
- Dompe, C.; Moncrieff, L.; Matys, J.; Grzech-Leśniak, K.; Kocherova, I.; Bryja, A.; Bruska, B.; Dominiak, M.; Mozdziak, P.; Hiroshi, T.; et al. Photobiomodulation-Underlying Mechanism and Clinical Applications. J. Clin. Med. 2020, 6, 1724. [Google Scholar] [CrossRef]
- Hamblin, M.; Demidova, T.N. Mechanisms of low level light therapy. Proc. SPIE ISOE 2006, 6140, 1–12. [Google Scholar]
- Oyebode, O.; Houreld, N.N.; Abrahamse, H. Photobiomodulation in diabetic wound healing: A review of red and near-infrared wavelength applications. Cell Biochem. Funct 2021, 39, 596–612. [Google Scholar] [CrossRef]
- Kushibiki, T.; Ishihara, M. Biological Function of Low Reactive Level Laser Therapy (LLLT). Photomed. Adv. Clin. Pract. InTech. 2017. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.C.Y.; Wu, D.F.; Zhu, L.; Peng, P.; Liu, L.; Yang, X.B. Microenvironment Dependent Photobiomodulation on Function-Specific Signal Transduction Pathways. Int. J. Photoenergy 2014, 904304. [Google Scholar] [CrossRef]
- Feng, J.; Li, X.; Zhu, S.; Xie, Y.; Du, J.; Ge, H.; Bai, Y. Photobiomodulation with 808-nm diode laser enhances gingival wound healing by promoting migration of human gingival mesenchymal stem cells via ROS/JNK/NF-?B/MMP-1 pathway. Lasers Med. Sci. 2020, 8, 1831. [Google Scholar] [CrossRef]
- Rajendran, N.K.; Houreld, N.N.; Abrahamse, H. In Vitro Wound Healing Potential of Photobiomodulation Is Possibly Mediated by Its Stimulatory Effect on AKT Expression in Adipose-Derived Stem Cells. Oxid. Med. Cell. Longev. 2021, 9, 6664627. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.H.; Moon, J.H.; Jung, J.Y.; Oh, C.; Ahn, J.C.; Chung, P.S. Effect of photobiomodulation therapy on neuronal injuries by ouabain: The regulation of Na, K-ATPase; Src; and mitogen-activated protein kinase signaling pathway. BMC Neurosci. 2019, 20, 19. [Google Scholar] [CrossRef]
- Ye, X.; Wang, L.; Dang, Y.; Liu, B.; Zhao, D. Investigation of the 1064 nm Q-switched Nd:YAG laser on collagen expression in an animal model. Photomed. Laser Surg. 2012, 10, 6049. [Google Scholar] [CrossRef]
- Neves, L.M.S.; Gonçalves, E.C.D.; Cavalli, J.; Vieira, G.; Laurindo, L.R.; Simões, R.R.; Coelho, R.S.; Santos, A.R.S.; Marcolino, A.M.; Cola, M.; et al. Photobiomodulation Therapy Improves Acute Inflammatory Response in Mice: The Role of Cannabinoid Receptors/ATP-Sensitive K+ Channel/p38-MAPK Signalling Pathway. Mol. Neurobiol. 2018, 7, 5580–5593. [Google Scholar] [CrossRef] [PubMed]
- Bamps, M.; Dok, R.; Nuyts, S. Low-Level Laser Therapy Stimulates Proliferation in Head and Neck Squamous Cell Carcinoma. Cells Front. Oncol. 2018, 8, 343. [Google Scholar] [CrossRef]
- Shingyochi, Y.; Kanazawa, S.; Tajima, S.; Tanaka, R.; Mizuno, H.; Tobita, M. A Low-Level Carbon Dioxide Laser Promotes Fibroblast Proliferation and Migration through Activation of Akt, ERK, and JNK. PLoS ONE 2017, 1, e0168937. [Google Scholar] [CrossRef]
- Song, J.M.; Park, B.S.; Shin, S.H.; Kim, I.R. Low-Level Laser Irradiation Stimulates RANKL-Induced Osteoclastogenesis via the MAPK Pathway in RAW264.7 Cells. Appl. Sci. 2021, 11, 5360. [Google Scholar] [CrossRef]
- Rajendran, N.K.; Houreld, N.N.; Abrahamse, H. Photobiomodulation reduces oxidative stress in diabetic wounded fibroblast cells by inhibiting the FOXO1 signaling pathway. J. Cell. Commun. Signal. 2021, 2, 195–206. [Google Scholar] [CrossRef]
- Kim, J.E.; Woo, Y.J.; Sohn, K.M.; Jeong, K.H.; Kang, H. Wnt/β-catenin and ERK pathway activation: A possible mechanism of photobiomodulation therapy with light-emitting diodes that regulate the proliferation of human outer root sheath cells. Lasers Surg. Med. 2017, 10, 940–947. [Google Scholar] [CrossRef]
- Liang, J.; Liu, L.; Xin, D. Photobiomodulation by low-power laser irradiation attenuates Aβ-induced cell apoptosis through the Akt/GSK3β/β-catenin pathway. Free Radic. Biol. Med. 2012, 7, 1459–1467. [Google Scholar]
- Han, L.; Liu, B.; Chen, X.; Chen, H.; Deng, W.; Yang, C.; Ji, B.; Wan, M. Activation of Wnt/β-catenin signaling is involved in hair growth-promoting effect of 655-nm red light and LED in in vitro culture model. Lasers Med. Sci. 2018, 3, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Jere, S.W.; Abrahamse, H.; Houreld, N.N. Photobiomodulation at 660 nm stimulates proliferation and migration of diabetic wounded cells via the expression of epidermal growth factor and the JAK/STAT pathway. J. Photochem. Photobiol. B 2018, 179, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Jere, S.W.; Houreld, N.N.; Abrahamse, H. Role of the PI3K/AKT (mTOR and GSK3β) signalling pathway and photobiomodulation in diabetic wound healing. Cytokine Growth Factor Rev. 2019, 50, 52–59. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jere, S.W.; Houreld, N.N. Regulatory Processes of the Canonical Wnt/β-Catenin Pathway and Photobiomodulation in Diabetic Wound Repair. Int. J. Mol. Sci. 2022, 23, 4210. https://doi.org/10.3390/ijms23084210
Jere SW, Houreld NN. Regulatory Processes of the Canonical Wnt/β-Catenin Pathway and Photobiomodulation in Diabetic Wound Repair. International Journal of Molecular Sciences. 2022; 23(8):4210. https://doi.org/10.3390/ijms23084210
Chicago/Turabian StyleJere, Sandy Winfield, and Nicolette Nadene Houreld. 2022. "Regulatory Processes of the Canonical Wnt/β-Catenin Pathway and Photobiomodulation in Diabetic Wound Repair" International Journal of Molecular Sciences 23, no. 8: 4210. https://doi.org/10.3390/ijms23084210