
Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio rerio)

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
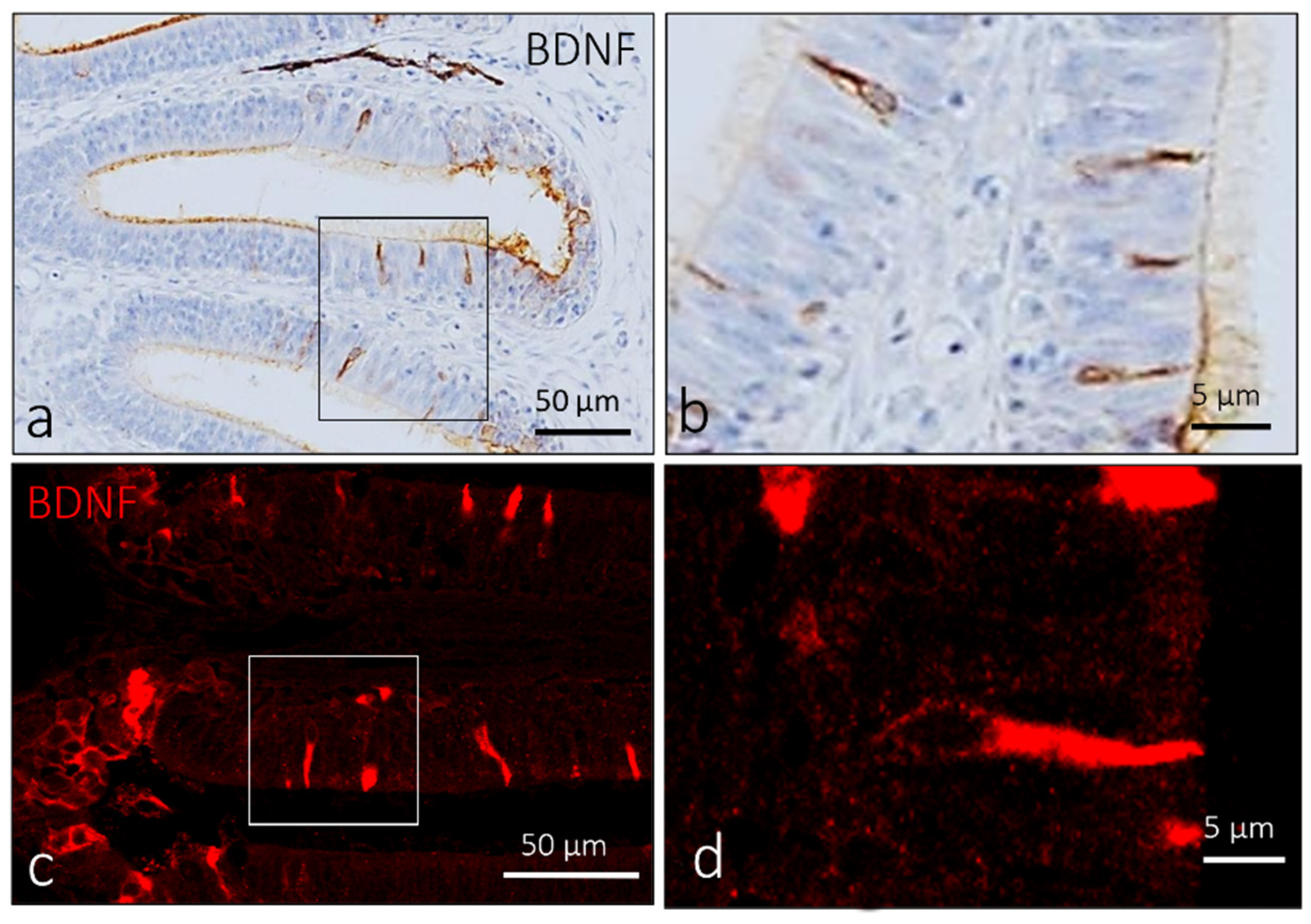
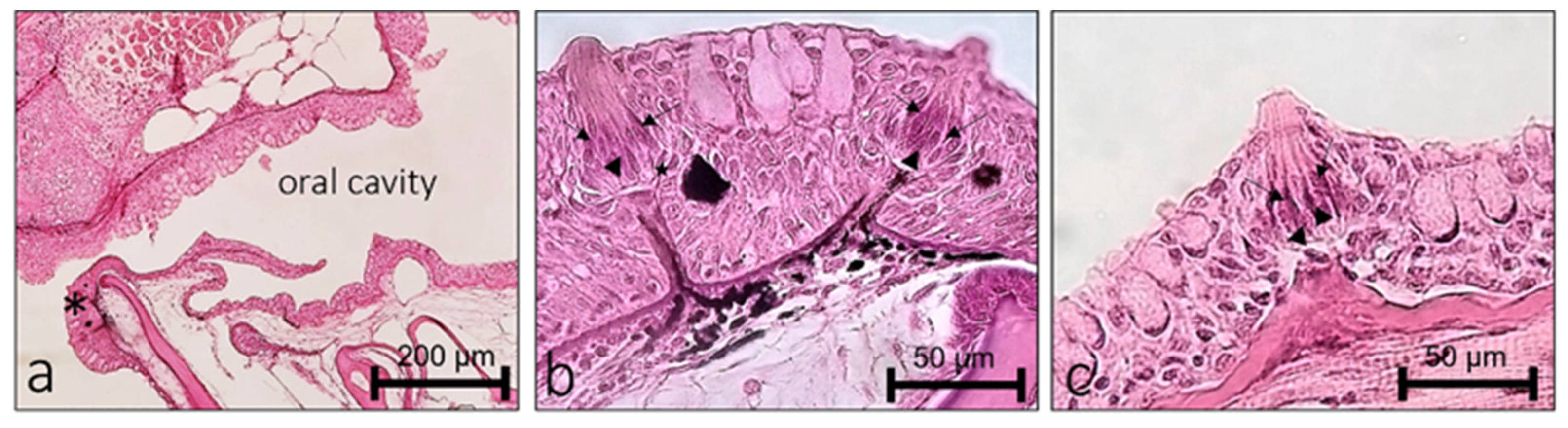
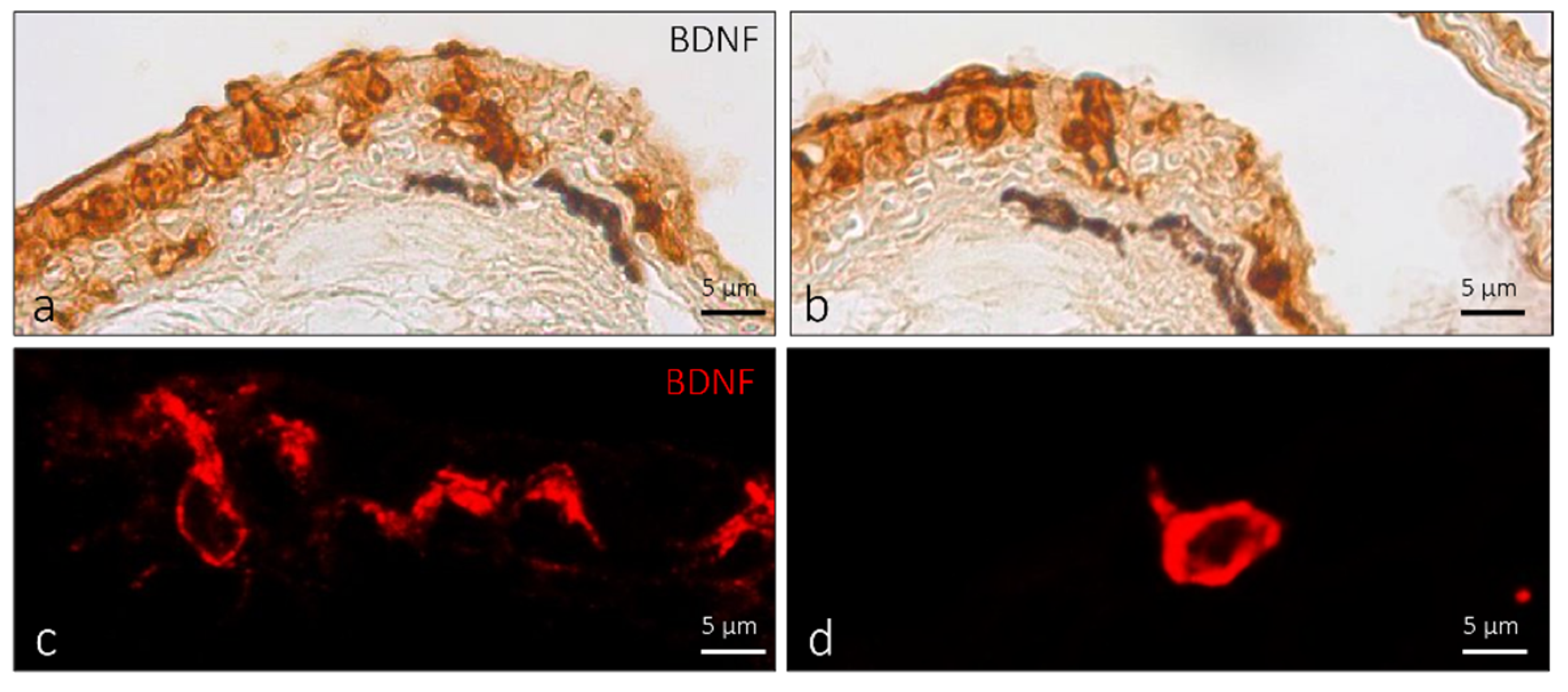
2.1. Localization of BDNF in the Olfactory Epithelium of Zebrafish during Development
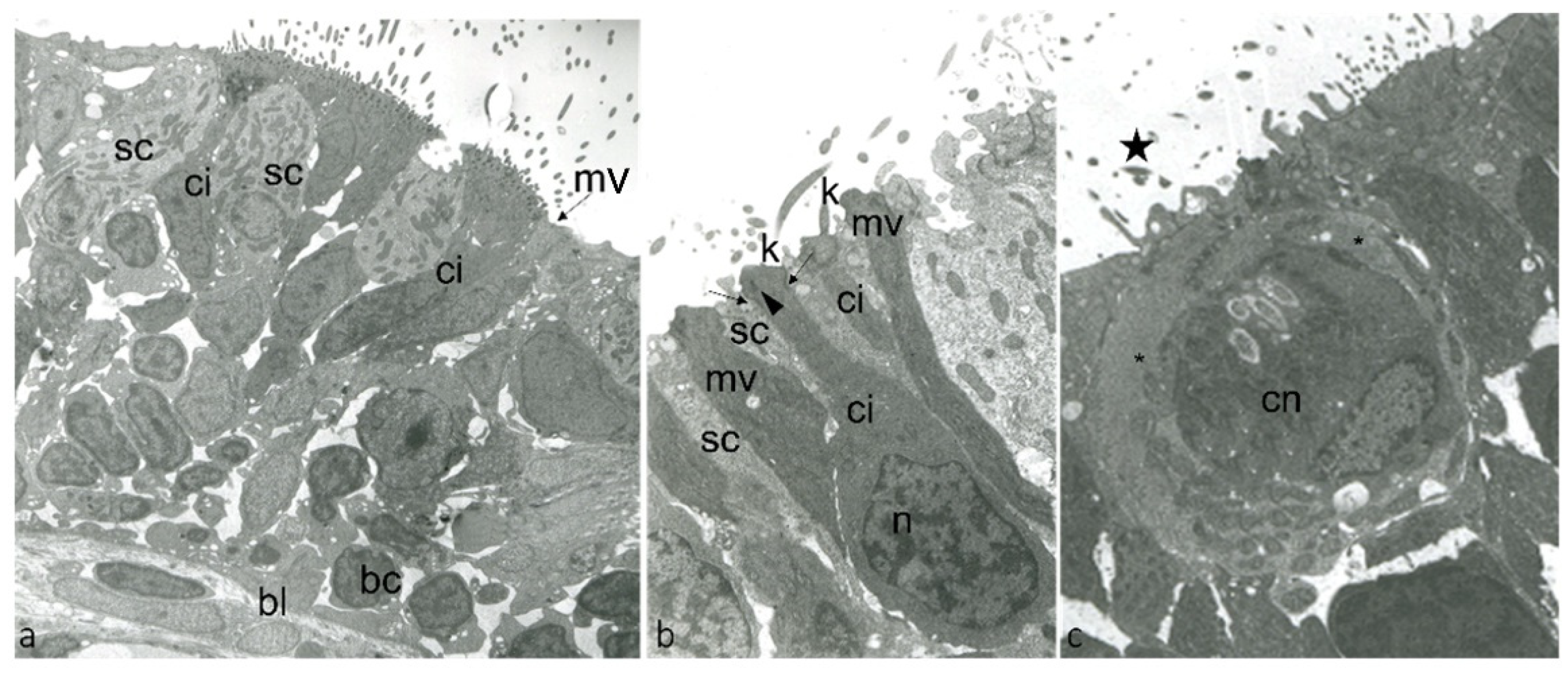
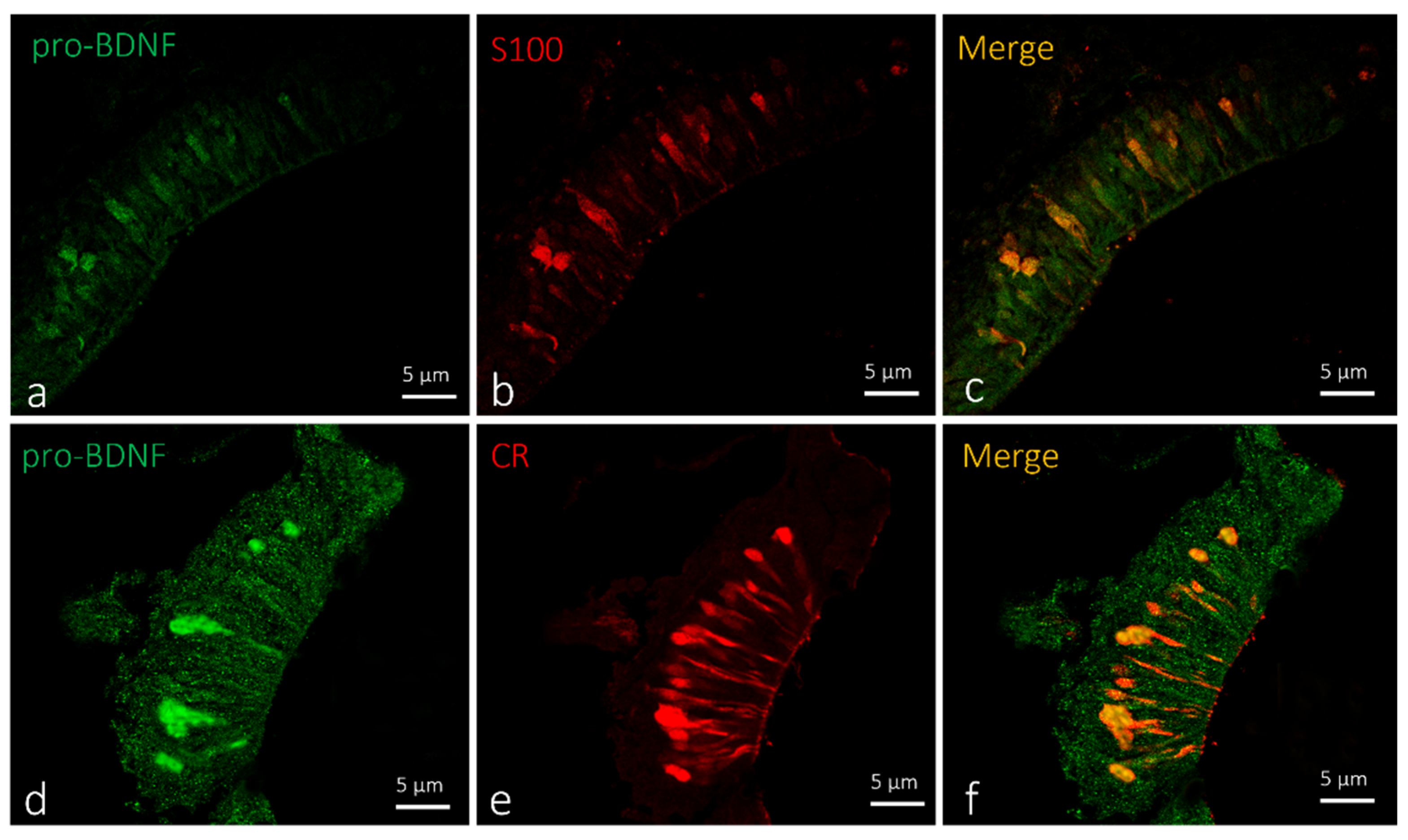
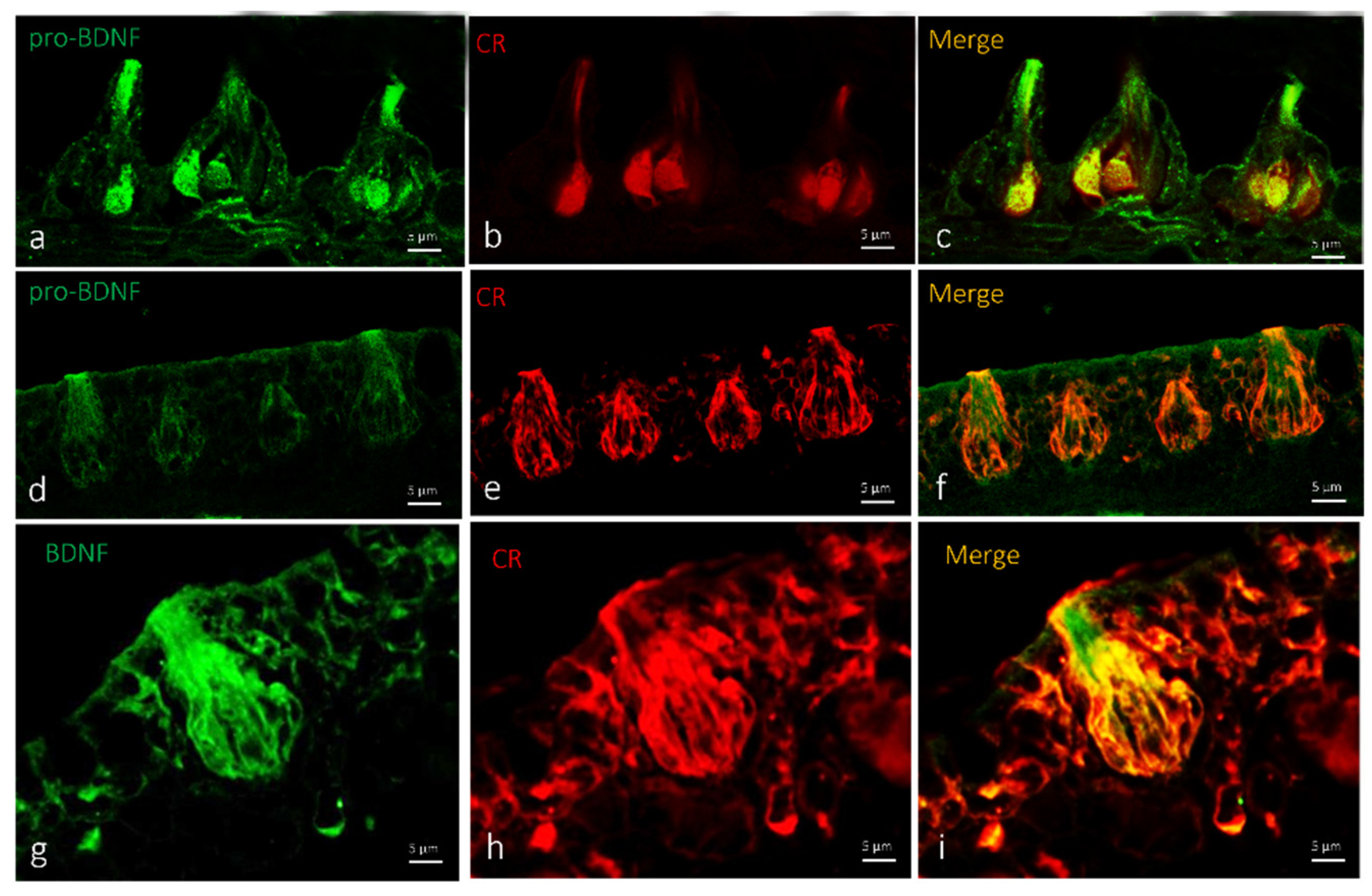
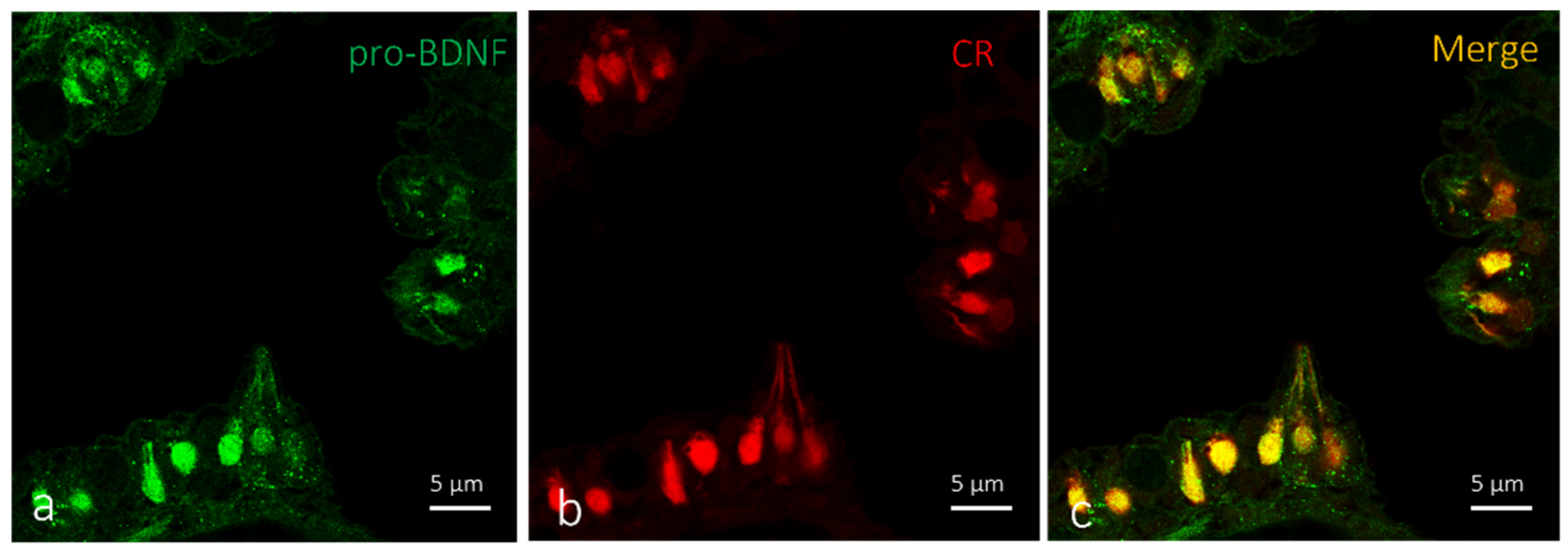
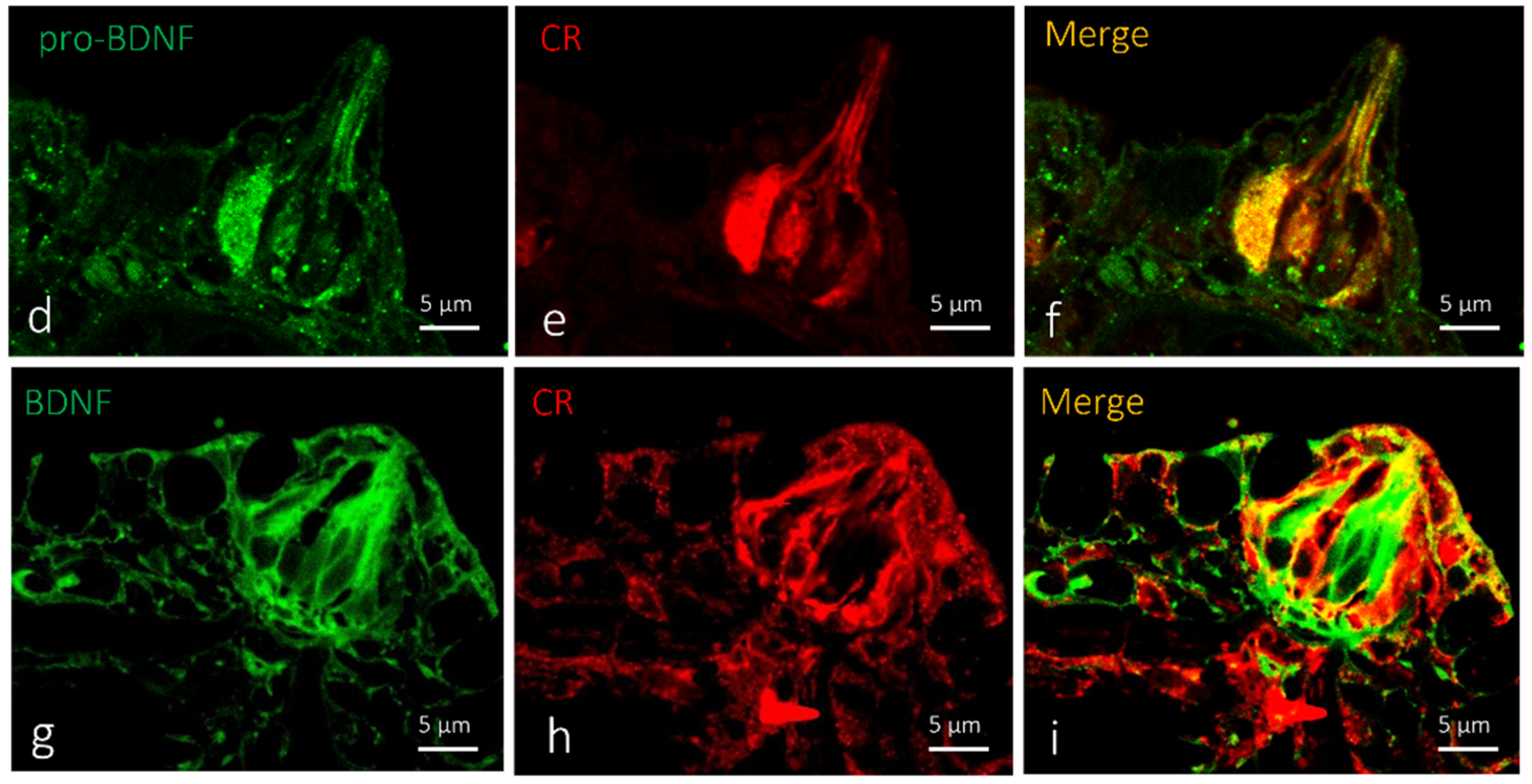
2.2. Localization of BDNF in Taste Buds and ICCC of Zebrafish during Development
3. Discussions
4. Materials and Methods
4.1. Trasmission Electron Microscopy
4.2. Immunohistochemical Detection of BDNF, Pro-BDNF, Calretinin and S100 Protein in the Olfactory Epithelium, Taste Buds and ICCC
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cacialli, P.; Gatta, C.; D’Angelo, L.; Leggieri, A.; Palladino, A.; de Girolamo, P.; Pellegrini, E.; Lucini, C. Nerve growth factor is expressed and stored in central neurons of adult zebrafish. J. Anat. 2019, 235, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Pérez-Pérez, M.; Germanà, A.; Esteban, I.; Germanà, G. Involvement of growth factors in thymic involution. Microsc. Res. Tech. 2003, 62, 514–523. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Germanà, A.; Hannestad, J.; Pérez-Pérez, M.; Esteban, I.; Naves, J.F.; Vega, J.A. Changes in the expression of the nerve growth factor receptors TrkA and p75LNGR in the rat thymus with ageing and increased nerve growth factor plasma levels. Cell Tissue Res. 2000, 301, 225–234. [Google Scholar] [CrossRef]
- Vega, J.A.; García-Suárez, O.; Hannestad, J.; Pérez-Pérez, M.; Germanà, A. Neurotrophins and the immune system. J. Anat. 2003, 203, 1–19. [Google Scholar] [CrossRef]
- Lanave, C.; Colangelo, A.M.; Saccone, C.; Alberghina, L. Molecular evolution of the neurotrophin family members and their Trk receptors. Gene 2007, 394, 1–12. [Google Scholar] [CrossRef]
- Tettamanti, G.; Cattaneo, A.G.; Gornati, R.; de Eguileor, M.; Bernardini, G.; Binelli, G. Phylogenesis of brain-derived neurotrophic factor (BDNF) in vertebrates. Gene 2010, 450, 85–93. [Google Scholar] [CrossRef]
- Hannestad, J.; Marino, F.; Germanà, A.; Catania, S.; Abbate, F.; Ciriaco, E.; Vega, J. Trk neurotrophin receptor-like proteins in the teleost Dicentrarchus labrax. Cell Tissue Res. 2000, 300, 1–9. [Google Scholar] [CrossRef]
- Heinrich, G.; Lum, T. Fish neurotrophins and Trk receptors. Int. J. Dev. Neurosci. 2000, 18, 1–27. [Google Scholar] [CrossRef]
- Germana, A.; Catania, S.; Cavallaro, M.; González-Martínez, T.; Ciriaco, E.; Hannestad, J.; Vega, J. Immunohistochemical localization of BDNF-, TrkB-and TrkA-like proteins in the teleost lateral line system. J. Anat. 2002, 200, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Levanti, M.; Abbate, F.; Laurà, R.; Germanà, A. Localization of Neurotrophin Specific Trk Receptors in Mechanosensory Systems of Killifish (Nothobranchius guentheri). Int. J. Mol. Sci. 2021, 22, 10411. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Heinrich, G. Brain-derived neurotrophic factor gene expression in the developing zebrafish. Int. J. Dev. Neurosci. 1997, 15, 983–997. [Google Scholar] [CrossRef]
- Cacialli, P.; D’Angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.-M.; Pellegrini, E.; Lucini, C. Neuronal expression of brain derived neurotrophic factor in the injured telencephalon of adult zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef]
- Cacialli, P.; Gueguen, M.-M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF expression in larval and adult zebrafish brain: Distribution and cell identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, L.; de Girolamo, P.; Cellerino, A.; Tozzini, E.T.; Castaldo, L.; Lucini, C. Neurotrophin Trk receptors in the brain of a teleost fish, Nothobranchius furzeri. Microsc. Res. Tech. 2012, 75, 81–88. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Levanti, M.B.; Germanà, G.P.; Navarra, M.; Laurà, R.; Vega, J.A.; Ciriaco, E.; Germanà, A. Expression and anatomical distribution of TrkB in the encephalon of the adult zebrafish (Danio rerio). Neurosci. Lett. 2014, 563, 66–69. [Google Scholar] [CrossRef]
- D’Angelo, L.; De Girolamo, P.; Lucini, C.; Terzibasi, E.T.; Baumgart, M.; Castaldo, L.; Cellerino, A. Brain-derived neurotrophic factor: mRNA expression and protein distribution in the brain of the teleost Nothobranchius furzeri. J. Comp. Neurol. 2014, 522, 1004–1030. [Google Scholar] [CrossRef]
- Montalbano, G.; Mania, M.; Guerrera, M.C.; Abbate, F.; Laurà, R.; Navarra, M.; Vega, J.A.; Ciriaco, E.; Germanà, A. Morphological differences in adipose tissue and changes in BDNF/Trkb expression in brain and gut of a diet induced obese zebrafish model. Ann. Anat.-Anat. Anz. 2016, 204, 36–44. [Google Scholar] [CrossRef]
- Nittoli, V.; Sepe, R.M.; Coppola, U.; D’Agostino, Y.; De Felice, E.; Palladino, A.; Vassalli, Q.A.; Locascio, A.; Ristoratore, F.; Spagnuolo, A.; et al. A comprehensive analysis of neurotrophins and neurotrophin tyrosine kinase receptors expression during development of zebrafish. J. Comp. Neurol. 2018, 526, 1057–1072. [Google Scholar] [CrossRef]
- Catania, S.; Germana, A.; Cabo, R.; Ochoa-Erena, F.; Guerrera, M.; Hannestad, J.; Represa, J.; Vega, J. Neurotrophin and Trk neurotrophin receptors in the inner ear of Salmo salar and Salmo trutta. J. Anat. 2007, 210, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Guerrera, M.C.; Laurà, R.; Levanti, M.; Aragona, M.; Mhalhel, K.; Germanà, G.; Montalbano, G.; Abbate, F. Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear. Int. J. Mol. Sci. 2020, 21, 5787. [Google Scholar] [CrossRef] [PubMed]
- Lum, T.; Huynh, G.; Heinrich, G. Brain-derived neurotrophic factor and TrkB tyrosine kinase receptor gene expression in zebrafish embryo and larva. Int. J. Dev. Neurosci. 2001, 19, 569–587. [Google Scholar] [CrossRef]
- Germanà, A.; Laurà, R.; Montalbano, G.; Guerrera, M.C.; Amato, V.; Zichichi, R.; Campo, S.; Ciriaco, E.; Vega, J.A. Expression of Brain-Derived Neurotrophic Factor and TrkB in the Lateral Line System of Zebrafish During Development. Cell. Mol. Neurobiol. 2010, 30, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Sánchez-Ramos, C.; Guerrera, M.C.; Calavia, M.; Navarro, M.; Zichichi, R.; García-Suárez, O.; Pérez-Piñera, P.; Vega, J.A. Expression and cell localization of brain-derived neurotrophic factor and TrkB during zebrafish retinal development. J. Anat. 2010, 217, 214–222. [Google Scholar] [CrossRef]
- Sánchez-Ramos, C.; Bonnin-Arias, C.; Guerrera, M.C.; Calavia, M.; Chamorro, E.; Montalbano, G.; López-Velasco, S.; López-Muñiz, A.; Germanà, A.; Vega, J.A. Light regulates the expression of the BDNF/TrkB system in the adult zebrafish retina. Microsc. Res. Tech. 2013, 76, 42–49. [Google Scholar] [CrossRef]
- Germana, A.; González-Martínez, T.; Catania, S.; Laura, R.; Cobo, J.; Ciriaco, E.; Vega, J. Neurotrophin receptors in taste buds of adult zebrafish (Danio rerio). Neurosci. Lett. 2004, 354, 189–192. [Google Scholar] [CrossRef]
- Hansen, A.; Reutter, K.; Zeiske, E. Taste bud development in the zebrafish, Danio rerio. Dev. Dyn. 2002, 223, 483–496. [Google Scholar] [CrossRef]
- Hansen, A.; Eckart, Z. The peripheral olfactory organ of the zebrafish, Danio rerio: An ultrastructural study. Chem. Senses 1998, 23, 39–48. [Google Scholar] [CrossRef]
- Bang, P.I.; Sewell, W.F.; Malicki, J.J. Morphology and cell type heterogeneities of the inner ear epithelia in adult and juvenile zebrafish (Danio rerio). J. Compart. Neurol. 2001, 438, 173–190. [Google Scholar] [CrossRef]
- Korsching, S.I.; Argo, S.; Campenhausen, H.; Friedrich, R.W.; Rummrich, A.; Weth, F. Olfaction in zebrafish: What does a tiny teleost tell us? Semin. Cell Dev. Biol. 1997, 8, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, T.T. Zebrafish as a model for hearing and deafness. J. Neurobiol. 2002, 53, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Paruta, S.; Germanà, G.P.; Ochoa-Erena, F.J.; Montalbano, G.; Cobo, J.; Vega, J.A. Differential distribution of S100 protein and calretinin in mechanosensory and chemosensory cells of adult zebrafish (Danio rerio). Brain Res. 2007, 1162, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Amato, V.; Vina, E.; Calavia, M.; Guerrera, M.C.; Laurà, R.; Navarro, M.; De Carlos, F.; Cobo, J.; Germanà, A.; Vega, J. TRPV4 in the sensory organs of adult zebrafish. Microsc. Res. Tech. 2012, 75, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Levanti, M.; Randazzo, B.; Viña, E.; Montalbano, G.; Garcia-Suarez, O.; Germanà, A.; Vega, J.; Abbate, F. Acid-sensing ion channels and transient-receptor potential ion channels in zebrafish taste buds. Ann. Anat.-Anat. Anz. 2016, 207, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Germana, A.; Montalbano, G.; Laura, R.; Ciriaco, E.; Del Valle, M.; Vega, J.A. S100 protein-like immunoreactivity in the crypt olfactory neurons of the adult zebrafish. Neurosci. Lett. 2004, 371, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Gayoso, J.Á.; Castro, A.; Anadón, R.; Manso, M.J. Differential bulbar and extrabulbar projections of diverse olfactory receptor neuron populations in the adult zebrafish (Danio rerio). J. Comp. Neurol. 2011, 519, 247–276. [Google Scholar] [CrossRef]
- Hansen, A.; Zielinski, B.S. Diversity in the olfactory epithelium of bony fishes: Development, lamellar arrangement, sensory neuron cell types and transduction components. J. Neurocytol. 2005, 34, 183–208. [Google Scholar] [CrossRef]
- Cacialli, P.; Lucini, C. Adult neurogenesis and regeneration in zebrafish brain: Are the neurotrophins involved in? Neural Regen. Res. 2019, 14, 2067–2068. [Google Scholar] [CrossRef]
- Cacialli, P. Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human. Int. J. Mol. Sci. 2021, 22, 1585. [Google Scholar] [CrossRef]
- Doty Richard, L.; Shaman, P.; Applebaum Steven, L.; Giberson, R.; Siksorski, L.; Rosenberg, L. Smell Identification Ability: Changes with Age. Science 1984, 226, 1441–1443. [Google Scholar] [CrossRef] [PubMed]
- Methven, L.; Allen, V.J.; Withers, C.A.; Gosney, M.A. Ageing and taste. Proc. Nutr. Soc. 2012, 71, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doty, R.; Kamath, V. The influences of age on olfaction: A review. Front. Psychol. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deems, D.A.; Doty, R.L.; Settle, R.G.; Moore-Gillon, V.; Shaman, P.; Mester, A.F.; Kimmelman, C.P.; Brightman, V.J.; Snow, J.B., Jr. Smell and Taste Disorders, A Study of 750 Patients From the University of Pennsylvania Smell and Taste Center. Arch. Otolaryngol.–Head Neck Surg. 1991, 117, 519–528. [Google Scholar] [CrossRef]
- Doty, R.L. Age-related deficits in taste and smell. Otolaryngol. Clin. N. Am. 2018, 51, 815–825. [Google Scholar] [CrossRef]
- DeVere, R. Disorders of taste and smell. Contin. Lifelong Learn. Neurol. 2017, 23, 421–446. [Google Scholar] [CrossRef]
- Landis, B.N.; Konnerth, C.G.; Hummel, T. A Study on the Frequency of Olfactory Dysfunction. Laryngoscope 2004, 114, 1764–1769. [Google Scholar] [CrossRef]
- Bettini, S.; Lazzari, M.; Ferrando, S.; Gallus, L.; Franceschini, V. Histopathological analysis of the olfactory epithelium of zebrafish (Danio rerio) exposed to sublethal doses of urea. J. Anat. 2016, 228, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-Y.; Lee, S.J.; Yun, J.H.; Bae, J.H. Taste Dysfunction after Tonsillectomy: A Meta-analysis. Ann. Otol. Rhinol. Laryngol. 2020, 130, 205–210. [Google Scholar] [CrossRef]
- Agyeman, A.A.; Chin, K.L.; Landersdorfer, C.B.; Liew, D.; Ofori-Asenso, R. Smell and Taste Dysfunction in Patients With COVID-19: A Systematic Review and Meta-analysis. Mayo Clin. Proc. 2020, 95, 1621–1631. [Google Scholar] [CrossRef]
- Evans, D.H.; Claiborne, J.B.; Currie, S. The Physiology of Fishes, 4th ed.; Taylor & Francis: Oxfordshire, UK, 2013. [Google Scholar]
- Lazzari, M.; Bettini, S.; Milani, L.; Maurizii, M.G.; Franceschini, V. Differential response of olfactory sensory neuron populations to copper ion exposure in zebrafish. Aquat. Toxicol. 2017, 183, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Parisi, V.; Guerrera, M.C.; Abbate, F.; Garcia-Suarez, O.; Viña, E.; Vega, J.A.; Germanà, A. Immunohistochemical characterization of the crypt neurons in the olfactory epithelium of adult zebrafish. Ann. Anat. Anat. Anz. 2014, 196, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, P.; Caprio, J. Chemoreception. In The Physiology of Fishes, 2nd ed.; CRC Marine Science Series 9; CRC Press: Boca Raton, FL, USA, 1998; pp. 375–405. ISBN 0-8493-8427-3. [Google Scholar]
- Hamdani, E.H.; Døving, K.B. Specific Projection of the Sensory Crypt Cells in the Olfactory System in Crucian Carp, Carassius carassius. Chem. Senses 2006, 31, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisthen, H.; Polese, G. Evolution of vertebrate olfactory subsystems. Brain Behav. Evol. 2007, 50, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.Y.; Rubel, E.W.; Raible, D.W. Notch Signaling Regulates the Extent of Hair Cell Regeneration in the Zebrafish Lateral Line. J. Neurosci. 2008, 28, 2261. [Google Scholar] [CrossRef] [Green Version]
- Uranagase, A.; Katsunuma, S.; Doi, K.; Nibu, K.-I. BDNF expression in olfactory bulb and epithelium during regeneration of olfactory epithelium. Neurosci. Lett. 2012, 516, 45–49. [Google Scholar] [CrossRef]
- Ernfors, P.; Lee, K.-F.; Jaenisch, R. Mice lacking brain-derived neurotrophic factor develop with sensory deficits. Nature 1994, 368, 147–150. [Google Scholar] [CrossRef]
- Lindsay, R.M. Neurotrophic growth factors and neurodegenerative diseases: Therapeutic potential of the neurotrophins and ciliary neurotrophic factor. Neurobiol. Aging 1994, 15, 249–251. [Google Scholar] [CrossRef]
- Cohen-Cory, S.; Fraser, S.E. Effects of brain-derived neurotrophic factor on optic axon branching and remodelling in vivo. Nature 1995, 378, 192–196. [Google Scholar] [CrossRef]
- De Felice, E.; Porreca, I.; Alleva, E.; De Girolamo, P.; Ambrosino, C.; Ciriaco, E.; Germanà, A.; Sordino, P. Localization of BDNF expression in the developing brain of zebrafish. J. Anat. 2014, 224, 564–574. [Google Scholar] [CrossRef]
- Anand, S.K.; Mondal, A.C. Neuroanatomical distribution and functions of brain-derived neurotrophic factor in zebrafish (Danio rerio) brain. J. Neurosci. Res. 2019, 98, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Lucini, C.; D’Angelo, L.; Cacialli, P.; Palladino, A.; De Girolamo, P. BDNF, Brain, and Regeneration: Insights from Zebrafish. Int. J. Mol. Sci. 2018, 19, 3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Cometa, M.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; et al. The BDNF/TrkB Neurotrophin System in the Sensory Organs of Zebrafish. Int. J. Mol. Sci. 2022, 23, 2621. [Google Scholar] [CrossRef]
- Díaz-Regueira, S.M.; Lamas, I.; Anadón, R. Calretinin immunoreactivity in taste buds and afferent fibers of the grey mullet Chelon Labrosus. Brain Res. 2005, 1031, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Porteros, A.; Arévalo, R.; Weruaga, E.; Crespo, C.; Briñón, J.G.; Alonso, J.R.; Aijón, J. Calretinin immunoreactivity in the developing olfactory system of the rainbow trout. Dev. Brain Res. 1997, 100, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Kress, S.; Biechl, D.; Wullimann, M.F. Combinatorial analysis of calcium-binding proteins in larval and adult zebrafish primary olfactory system identifies differential olfactory bulb glomerular projection fields. Brain Struct. Funct. 2015, 220, 1951–1970. [Google Scholar] [CrossRef]
- Hansen, A.; Rolen, S.H.; Anderson, K.; Morita, Y.; Caprio, J.; Finger, T.E. Correlation between Olfactory Receptor Cell Type and Function in the Channel Catfish. J. Neurosci. 2003, 23, 9328. [Google Scholar] [CrossRef]
- Kolmakov, N.N.; Hubbard, P.C.; Lopes, O.; Canario, A.V.M. Effect of Acute Copper Sulfate Exposure on Olfactory Responses to Amino Acids and Pheromones in Goldfish (Carassius auratus). Environ. Sci. Technol. 2009, 43, 8393–8399. [Google Scholar] [CrossRef] [Green Version]
- Jakubowski, M.; Whitear, M. Comparative morphology and cytology of taste buds in teleosts. Jahrb. Für Morphol. Und Mikrosk. Anatomie. 2. Abt. Z. Für Mikrosk. Anat. Forsch. 1990, 104, 529–560. [Google Scholar]
- Reutter, K. Taste Organ in the Bullhead (Teleostei); Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Mistretta, C.M.; Goosens, K.A.; Farinas, I.; Reichardt, L.F. Alterations in size, number, and morphology of gustatory papillae and taste buds in BDNF null mutant mice demonstrate neural dependence of developing taste organs. J. Comp. Neurol. 1999, 409, 13–24. [Google Scholar] [CrossRef]
- Krimm, R.F.; Miller, K.K.; Kitzman, P.H.; Davis, B.M.; Albers, K.M. Epithelial Overexpression of BDNF or NT4 Disrupts Targeting of Taste Neurons That Innervate the Anterior Tongue. Dev. Biol. 2001, 232, 508–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzsch, B.; Pauley, S.; Beisel, K.W. Cells, molecules and morphogenesis: The making of the vertebrate ear. Brain Res. 2006, 1091, 151–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farbman, A.I. Neurotrophins and taste buds. J. Comp. Neurol. 2003, 459, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ganchrow, D.; Ganchrow, J.R.; Verdin-Alcazar, M.; Whitehead, M.C. Brain-derived neurotrophic factor-, neurotrophin-3-, and tyrosine kinase receptor-like immunoreactivity in lingual taste bud fields of mature hamster. J. Comp. Neurol. 2003, 455, 11–24. [Google Scholar] [CrossRef]
- Uchida, N.; Kanazawa, M.; Suzuki, Y.; Takeda, M. Expression of BDNF and TrkB in mouse taste buds after denervation and in circumvallate papillae during development. Arch. Histol. Cytol. 2003, 66, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Hallböök, F. Evolution of the vertebrate neurotrophin and Trk receptor gene families. Curr. Opin. Neurobiol. 1999, 9, 616–621. [Google Scholar] [CrossRef]
- Miura, H.; Barlow, L.A. Taste bud regeneration and the search for taste progenitor cells. Arch. Ital. Biol. 2010, 148, 107–118. [Google Scholar]
- Calvo-Ochoa, E.; Byrd-Jacobs, C.A.; Fuss, S.H. Diving into the streams and waves of constitutive and regenerative olfactory neurogenesis: Insights from zebrafish. Cell Tissue Res. 2021, 383, 227–253. [Google Scholar] [CrossRef]
- Calvo-Ochoa, E.; Byrd-Jacobs, C.A. The Olfactory System of Zebrafish as a Model for the Study of Neurotoxicity and Injury: Implications for Neuroplasticity and Disease. Int. J. Mol. Sci. 2019, 20, 1639. [Google Scholar] [CrossRef] [Green Version]
- Pombal, M.A.; de Arriba, M.C.; Sampedro, C.; Alvarez, R.; Megías, M. Immunocytochemical localization of calretinin in the olfactory system of the adult lamprey, Lampetra fluviatilis. Brain Res. Bull. 2002, 57, 281–283. [Google Scholar] [CrossRef]
- Castro, A.; Becerra, M.; Anadón, R.; Manso, M.J. Distribution of calretinin during development of the olfactory system in the brown trout, Salmo trutta fario: Comparison with other immunohistochemical markers. J. Chem. Neuroanat. 2008, 35, 306–316. [Google Scholar] [CrossRef] [PubMed]
- White, E.J.; Kounelis, S.K.; Byrd-Jacobs, C.A. Plasticity of glomeruli and olfactory-mediated behavior in zebrafish following detergent lesioning of the olfactory epithelium. Neuroscience 2015, 284, 622–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, D.; Ghysen, A. Somatotopy of the lateral line projection in larval zebrafish. Proc. Natl. Acad. Sci. USA 1999, 96, 7558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northcutt, R.G.; Holmes, P.H.; Albert, J.S. Distribution and innervation of lateral line organs in the channel catfish. J. Comp. Neurol. 2000, 421, 570–592. [Google Scholar] [CrossRef]
- Ostrander, G.K.; Hopkins, J. The Laboratory Fish; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Gompel, N.; Dambly-Chaudiere, C.; Ghysen, A. Neuronal differences prefigure somatotopy in the zebrafish lateral line. Development 2001, 128, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Collazo, A.; Fraser, S.E.; Mabee, P.M. A dual embryonic origin for vertebrate mechanoreceptors. Science 1994, 264, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Germanà, A.; Montalbano, G.; Guerrera, M.C.; Laura, R.; Levanti, M.; Abbate, F.; de Carlos, F.; Vega, J.A.; Ciriaco, E. Sox-2 in taste bud and lateral line system of zebrafish during development. Neurosci. Lett. 2009, 467, 36–39. [Google Scholar] [CrossRef]
- Germanà, A.; Marino, F.; Guerrera, M.C.; Campo, S.; de Girolamo, P.; Montalbano, G.; Germanà, G.P.; Ochoa-Erena, F.J.; Ciriaco, E.; Vega, J.A. Expression and distribution of S100 protein in the nervous system of the adult zebrafish (Danio rerio). Microsc. Res. Tech. 2008, 71, 248–255. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Supplier | Catalogue Number | Source | Diluition | Antibody ID |
|---|---|---|---|---|---|
| BDNF | Merck Millipore | AB1534SP | rabbit | 1:100 | AB_90748 |
| pro-BDNF | Santa Cruz Biotechnology | sc-65513 | mouse | 1:100 | AB_831028 |
| Calretinin | Santa Cruz Biotechnology | sc-11644 | goat | 1:100 | AB_634545 |
| S100 | Dako | Z0311 | rabbit | 1:100 | AB_10013383 |
| Secondary Antibodies | Supplier | Catalogue Number | Source | Diluition | Antibody ID |
|---|---|---|---|---|---|
| Antirabbit IgG (H+L) Alexa Fluor 594 | Invitrogen | A-21207 | donkey | 1:300 | AB_141607 |
| Antirabbit IgG (H+L) Alexa Fluor 488 | Invitrogen | A-11008 | goat | 1:300 | AB_143165 |
| Antigoat IgG (H+L) Alexa Fluor 594 | Invitrogen | A-11058 | donkey | 1:300 | AB_2534105 |
| Antimouse IgG (H+L) Alexa Fluor 488 | Invitrogen | A-11001 | goat | 1:300 | AB_2534069 |
| Antirabbit IgG-peroxidase conjugate | Amersham | NA934 | donkey | 1:100 | AB_772206 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; Calapai, F.; et al. Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio rerio). Int. J. Mol. Sci. 2022, 23, 4696. https://doi.org/10.3390/ijms23094696
Aragona M, Porcino C, Guerrera MC, Montalbano G, Laurà R, Levanti M, Abbate F, Cobo T, Capitelli G, Calapai F, et al. Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio rerio). International Journal of Molecular Sciences. 2022; 23(9):4696. https://doi.org/10.3390/ijms23094696
Chicago/Turabian StyleAragona, Marialuisa, Caterina Porcino, Maria Cristina Guerrera, Giuseppe Montalbano, Rosaria Laurà, Maria Levanti, Francesco Abbate, Teresa Cobo, Gabriel Capitelli, Fabrizio Calapai, and et al. 2022. "Localization of BDNF and Calretinin in Olfactory Epithelium and Taste Buds of Zebrafish (Danio rerio)" International Journal of Molecular Sciences 23, no. 9: 4696. https://doi.org/10.3390/ijms23094696