
Stereoselective Covalent Adduct Formation of Acyl Glucuronide Metabolite of Nonsteroidal Anti-Inflammatory Drugs with UDP-Glucuronosyltransferase

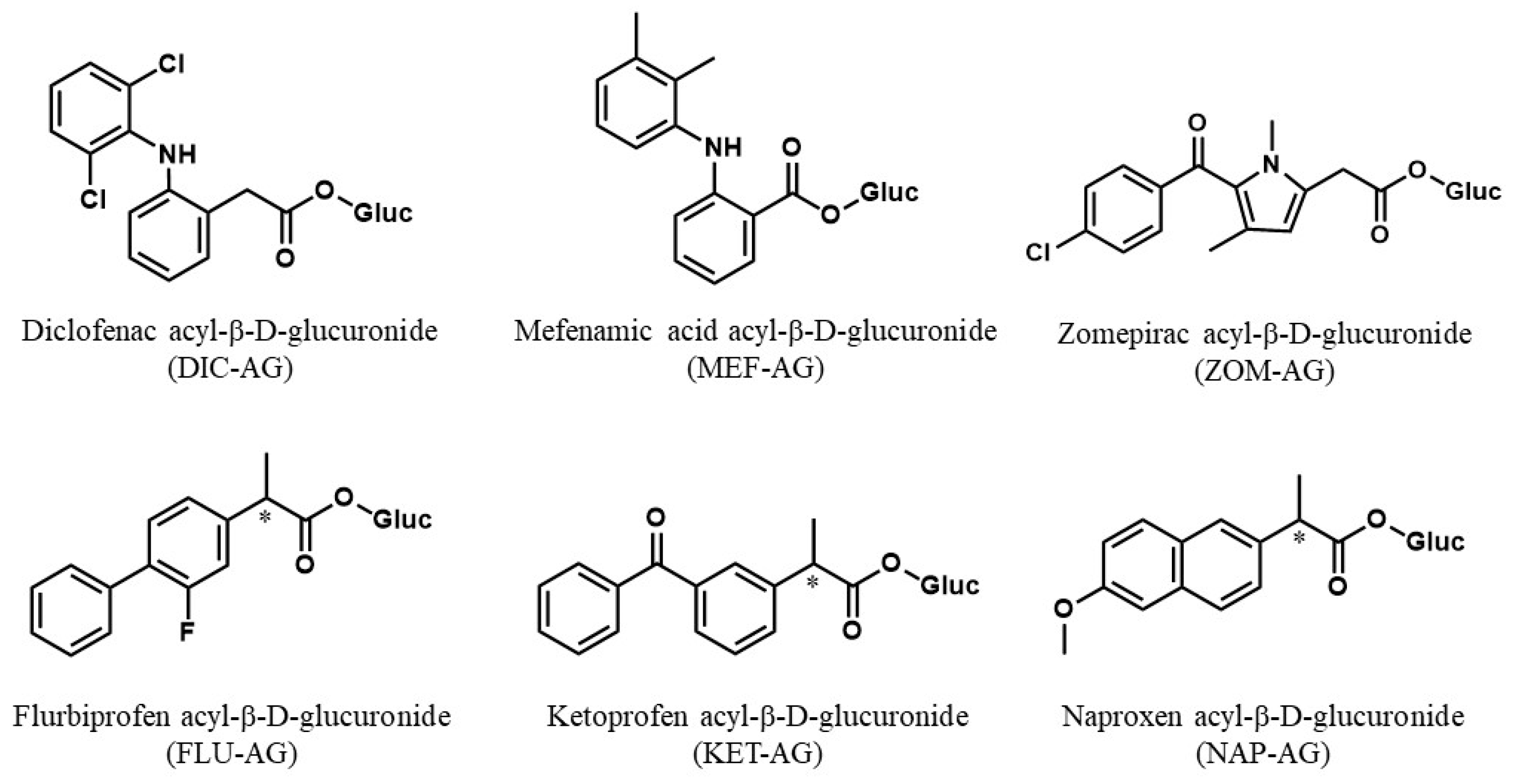{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
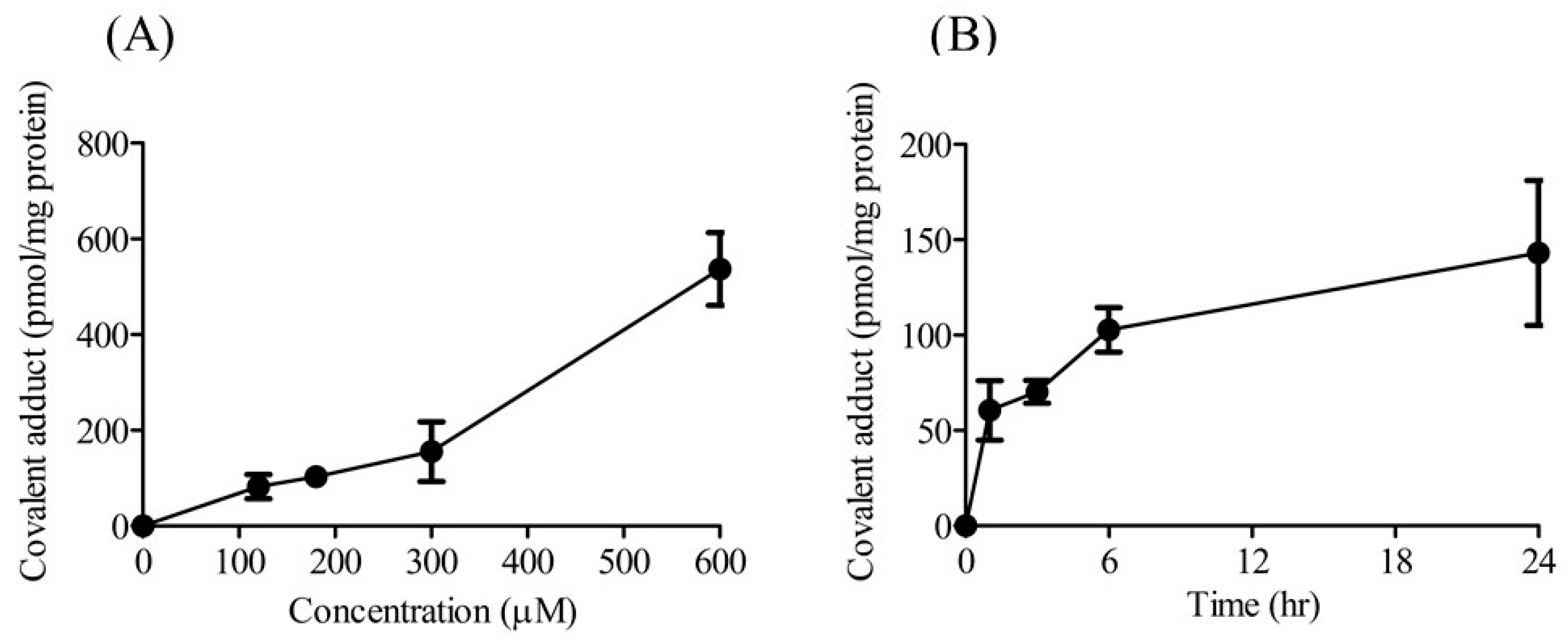
2.1. Concentration- and Time-Dependent Formation of Covalent Adducts
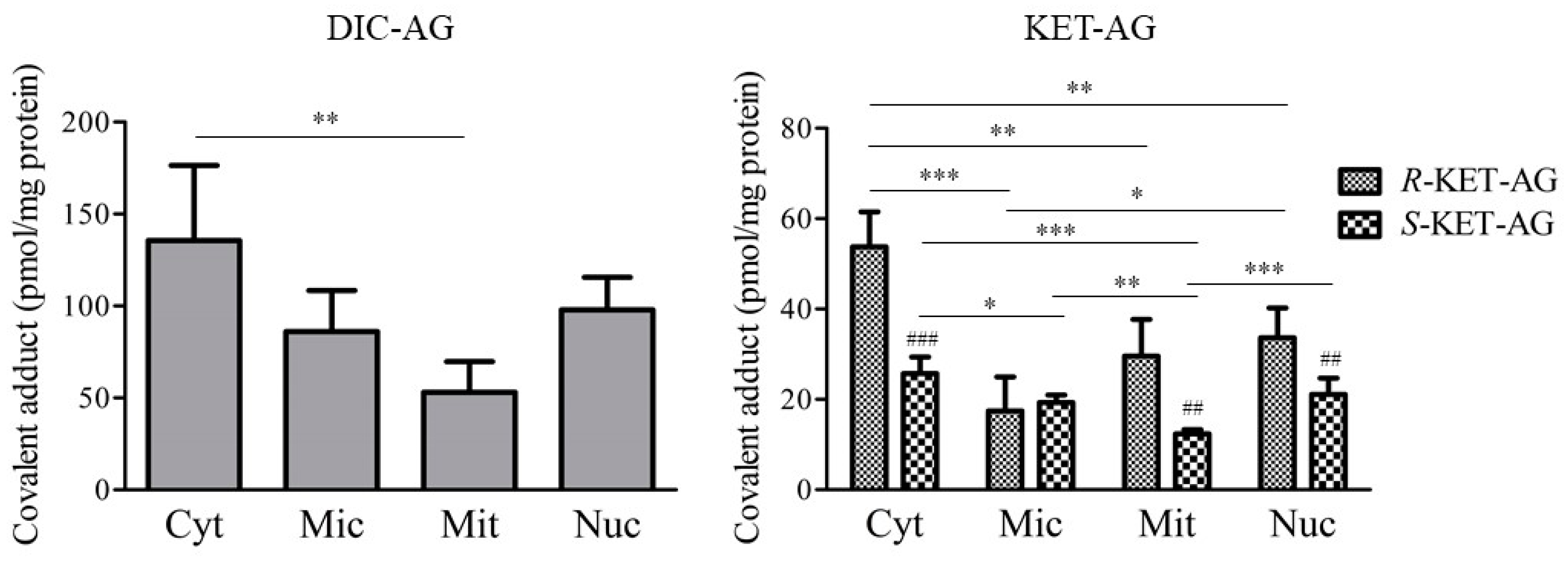
2.2. Covalent Adduct Formation with Organelle Proteins
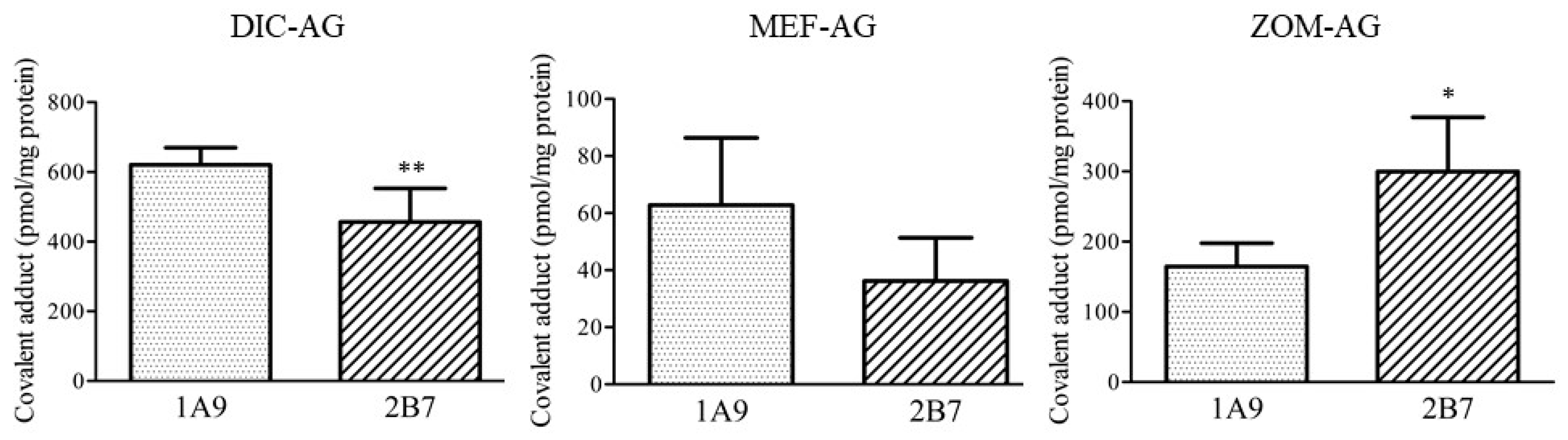
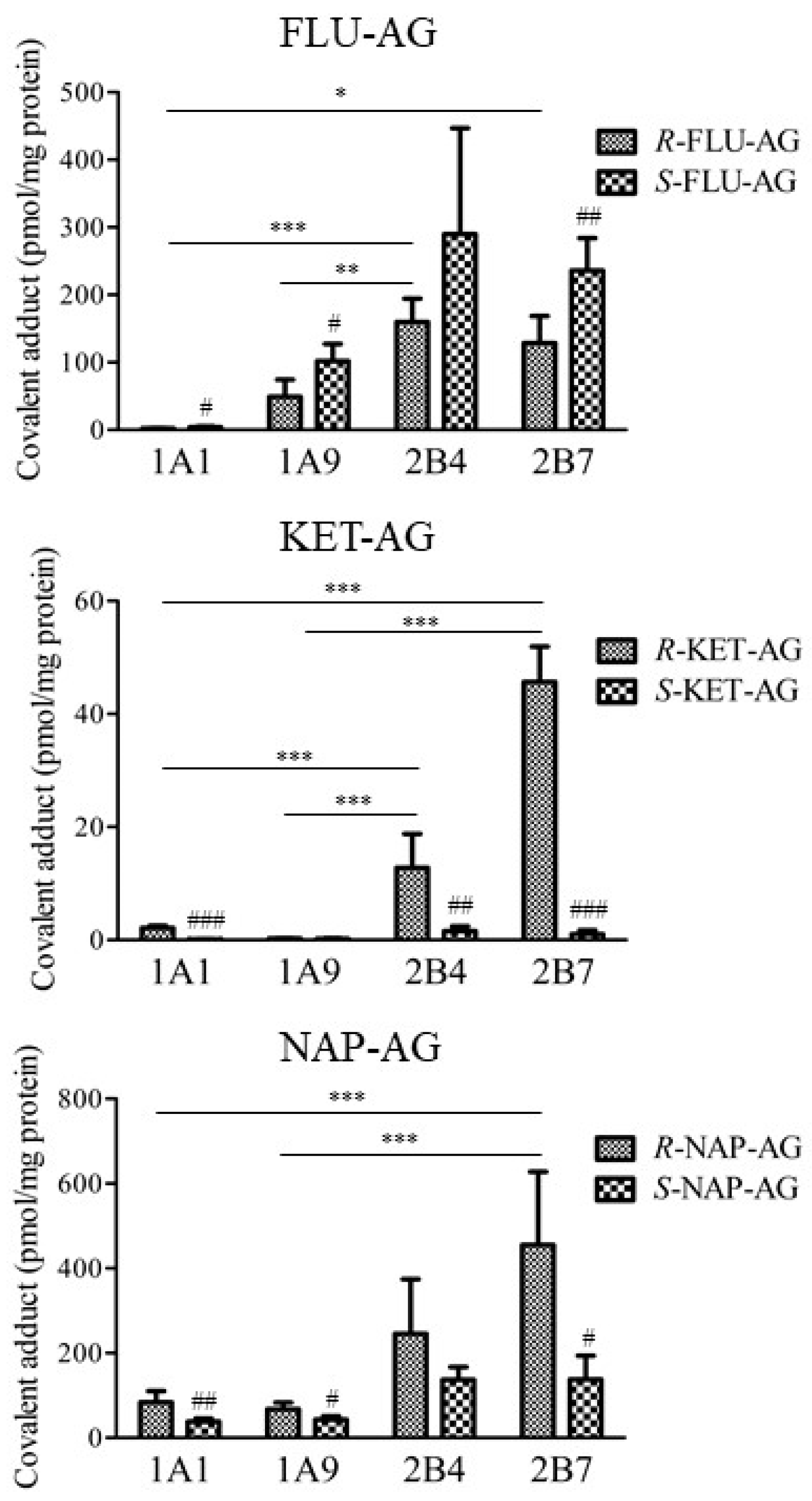
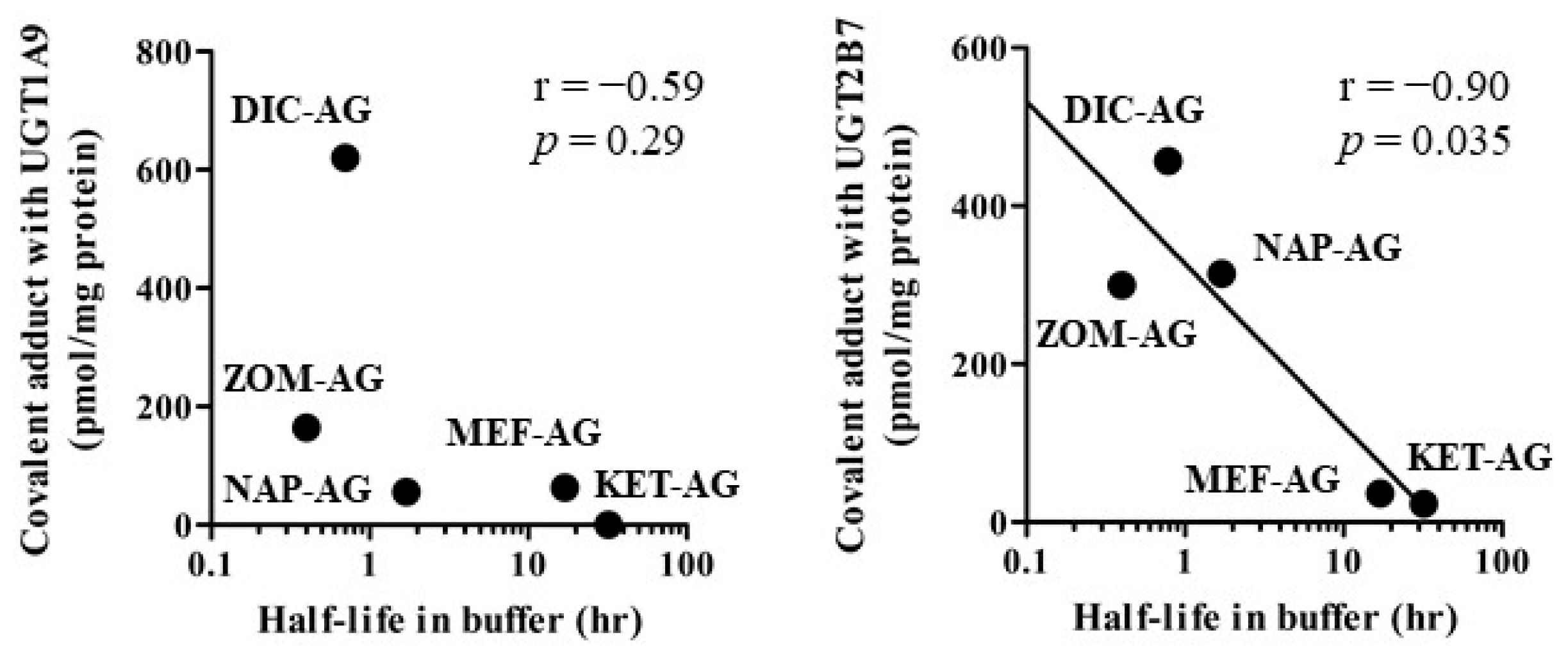
2.3. Covalent Adduct Formation with UGTs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Subcellular Fractionation of Liver Homogenates
4.4. Preparation of NSAIDs-AG
4.5. Covalent Adduct Formation of NSAIDs-AG with Organelle Proteins, UGT1A, and UGT2B Isoforms
4.6. Determination of NSAIDs Concentrations by LC-MS/MS Method
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AG | acyl-β-D-glucuronide |
| Cyt | cytosolic fraction |
| DIC | diclofenac |
| DILI | drug-induced liver injury |
| ER | endoplasmic reticulum |
| FLU | flurbiprofen |
| HPLC | high-performance liquid chromatography |
| HSA | human serum albumin |
| KET | ketoprofen |
| LC-MS/MS | liquid chromatographic-tandem mass spectrometry |
| MEF | mefenamic acid |
| Mic | microsomal fraction |
| Mit | mitochondrial fraction |
| m/z | mass to charge ratio |
| NAP | naproxen |
| NSAIDs | nonsteroidal anti-inflammatory drugs |
| Nuc | nuclear fraction |
| SRM | selected reaction monitoring |
| UGT | UDP-glucuronosyltransferase |
| ZOM | zomepirac |
References
- Kretz-Rommel, A.; Boelsterli, U.A. Selective Protein Adducts to Membrane Proteins in Cultured Rat Hepatocytes Exposed to Diclofenac: Radiochemical and Immunochemical Analysis. Mol. Pharmacol. 1994, 45, 237–244. [Google Scholar]
- Nagao, T.; Tanino, T.; Iwaki, M. Stereoselective Pharmacokinetics of Flurbiprofen and Formation of Covalent Adducts with Plasma Protein in Adjuvant-Induced Arthritic Rats. Chirality 2003, 15, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.; Kretz-Rommel, A.; Oude Elferink, R.P.; Boelsterli, U.A. Selective Protein Adduct Formation of Diclofenac Glucuronide Is Critically Dependent on the Rat Canalicular Conjugate Export Pump (Mrp2). Chem. Res. Toxicol. 1998, 11, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Kretz-Rommel, A.; Boelsterli, U.A. Mechanism of Covalent Adduct Formation of Diclofenac to Rat Hepatic Microsomal Proteins. Retention of the Glucuronic Acid Moiety in the Adduct. Drug Metab. Dispos. 1994, 22, 956–961. [Google Scholar] [PubMed]
- Wang, M.; Gorrell, M.D.; McGaughan, G.W.; Dickinson, R.G. Dipeptidyl Peptidase IV Is a Target for Covalent Adduct Formation with the Acyl Glucuronide Metabolite of the Anti-Inflammatory Drug Zomepirac. Life Sci. 2001, 68, 785–797. [Google Scholar] [CrossRef]
- Tailor, A.; Waddington, J.C.; Meng, X.; Park, B.K. Mass Spectrometric and Functional Aspects of Drug-Protein Conjugation. Chem. Res. Toxicol. 2016, 29, 1912–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrier, N.; Benoit, E.; Senay, C.; Lapicque, F.; Radominska-Pandya, A.; Magdalou, J.; Fournel-Gigleux, S. Human and Rat Liver UDP-Glucuronosyltransferases Are Targets of Ketoprofen Acylglucuronide. Mol. Pharmacol. 1999, 56, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Bischer, A.; Zia-Amirhosseini, P.; Iwaki, M.; McDonagh, A.F.; Benet, L.Z. Stereoselective Binding Properties of Naproxen Glucuronide Diastereomers to Proteins. J. Pharmacokinet. Biopharm. 1995, 23, 379–395. [Google Scholar] [CrossRef]
- Presle, N.; Lapicque, F.; Fournel-Gigleux, S.; Magdalou, J.; Netter, P. Stereoselective Irreversible Binding of Ketoprofen Glucuronides to Albumin. Characterization of the Site and the Mechanism. Drug Metab. Dispos. 1996, 24, 1050–1057. [Google Scholar]
- Shimada, H.; Kobayashi, Y.; Tanahashi, S.; Kawase, A.; Ogiso, T.; Iwaki, M. Correlation between Glucuronidation and Covalent Adducts Formation with Proteins of Nonsteroidal Anti-Inflammatory Drugs. Eur. J. Pharm. Sci. 2018, 112, 132–138. [Google Scholar] [CrossRef]
- Meech, R.; Hu, D.G.; McKinnon, R.A.; Mubarokah, S.N.; Haines, A.Z.; Nair, P.C.; Rowland, A.; Mackenzie, P.I. The UDP-Glycosyltransferase (UGT) Superfamily: New Members, New Functions, and Novel Paradigms. Physiol. Rev. 2019, 99, 1153–1222. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Miners, J.O.; Lillywhite, K.J.; Mackenzie, P.I. Complementary Deoxyribonucleic Acid Cloning and Expression of a Human Liver Uridine Diphosphate-Glucuronosyltransferase Glucuronidating Carboxylic Acid-Containing Drugs. J. Pharmacol. Exp. Ther. 1993, 264, 475–479. [Google Scholar] [PubMed]
- Sawamura, R.; Okudaira, N.; Watanabe, K.; Murai, T.; Kobayashi, Y.; Tachibana, M.; Ohnuki, T.; Masuda, K.; Honma, H.; Kurihara, A.; et al. Predictability of Idiosyncratic Drug Toxicity Risk for Carboxylic Acid-Containing Drugs Based on the Chemical Stability of Acyl Glucuronide. Drug Metab. Dispos. 2010, 38, 1857–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knadler, M.P.; Hall, S.D. Stereoselective Hydrolysis of Flurbiprofen Conjugates. Drug Metab. Dispos. 1991, 19, 280–282. [Google Scholar]
- Hayball, P.J.; Nation, R.L.; Bochner, F. Stereoselective Interactions of Ketoprofen Glucuronides with Human Plasma Protein and Serum Albumin. Biochem. Pharmacol. 1992, 44, 291–299. [Google Scholar] [CrossRef]
- Dubois-Presle, N.; Lapicque, F.; Maurice, M.H.; Fournel-Gigleux, S.; Magdalou, J.; Abiteboul, M.; Siest, G.; Netter, P. Stereoselective Esterase Activity of Human Serum Albumin toward Ketoprofen Glucuronide. Mol. Pharmacol. 1995, 47, 647–653. [Google Scholar]
- Hasegawa, J.; Smith, P.C.; Benet, L.Z. Apparent Intramolecular Acyl Migration of Zomepirac Glucuronide. Drug Metab. Dispos. 1982, 10, 469–473. [Google Scholar]
- Hyneck, M.L.; Munafo, A.; Benet, L.Z. Effect of PH on Acyl Migration and Hydrolysis of Tolmetin Glucuronide. Drug Metab. Dispos. 1988, 16, 322–324. [Google Scholar]
- Akira, K.; Taira, T.; Hasegawa, H.; Sakuma, C.; Shinohara, Y. Studies on the Stereoselective Internal Acyl Migration of Ketoprofen Glucuronides Using 13C Labeling and Nuclear Magnetic Resonance Spectroscopy. Drug Metab. Dispos. 1998, 26, 457–464. [Google Scholar]
- Mizuma, T.; Benet, L.Z.; Lin, E.T. Interaction of Human Serum Albumin with Furosemide Glucuronide: A Role of Albumin in Isomerization, Hydrolysis, Reversible Binding and Irreversible Binding of a 1-O-Acyl Glucuronide Metabolite. Biopharm. Drug Dispos. 1999, 20, 131–136. [Google Scholar] [CrossRef]
- Inoue, K.; Mizuo, H.; Ishida, T.; Komori, T.; Kusano, K. Bioactivation of Diclofenac in Human Hepatocytes and the Proposed Human Hepatic Proteins Modified by Reactive Metabolites. Xenobiotica 2020, 50, 919–928. [Google Scholar] [CrossRef] [PubMed]
- King, C.; Tang, W.; Ngui, J.; Tephly, T.; Braun, M. Characterization of Rat and Human UDP-Glucuronosyltransferases Responsible for the in Vitro Glucuronidation of Diclofenac. Toxicol. Sci. 2001, 61, 49–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuehl, G.E.; Lampe, J.W.; Potter, J.D.; Bigler, J. Glucuronidation of Nonsteroidal Anti-Inflammatory Drugs: Identifying the Enzymes Responsible in Human Liver Microsomes. Drug Metab. Dispos. 2005, 33, 1027–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunduz, M.; Argikar, U.A.; Cirello, A.L.; Dumouchel, J.L. New Perspectives on Acyl Glucuronide Risk Assessment in Drug Discovery: Investigation of In Vitro Stability, In Situ Reactivity, and Bioactivation. Drug Metab. Lett. 2018, 12, 84–92. [Google Scholar] [CrossRef]
- Gaganis, P.; Miners, J.O.; Knights, K.M. Glucuronidation of Fenamates: Kinetic Studies Using Human Kidney Cortical Microsomes and Recombinant UDP-Glucuronosyltransferase (UGT) 1A9 and 2B7. Biochem. Pharmacol. 2007, 73, 1683–1691. [Google Scholar] [CrossRef]
- Bolze, S.; Bromet, N.; Gay-Feutry, C.; Massiere, F.; Boulieu, R.; Hulot, T. Development of an in Vitro Screening Model for the Biosynthesis of Acyl Glucuronide Metabolites and the Assessment of Their Reactivity toward Human Serum Albumin. Drug Metab. Dispos. 2002, 30, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.-Z.; Li, L.; Wang, L.; Yuan, L.-M.; Xu, M.-C.; Gu, J.-K.; Jiang, H.; Yu, L.-S.; Zeng, S. The Regioselective Glucuronidation of Morphine by Dimerized Human UGT2B7, 1A1, 1A9 and Their Allelic Variants. Acta Pharmacol. Sin. 2017, 38, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Ishii, Y.; Iwanaga, M.; Nurrochmad, A.; Ito, Y.; Mackenzie, P.I.; Nagata, K.; Yamazoe, Y.; Oguri, K.; Yamada, H. Interaction of Cytochrome P450 3A4 and UDP-Glucuronosyltransferase 2B7: Evidence for Protein-Protein Association and Possible Involvement of CYP3A4 J-Helix in the Interaction. Mol. Pharmacol. 2009, 75, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Baba, A.; Yoshioka, T. Structure-Activity Relationships for Degradation Reaction of 1-Beta-o-Acyl Glucuronides: Kinetic Description and Prediction of Intrinsic Electrophilic Reactivity under Physiological Conditions. Chem. Res. Toxicol. 2009, 22, 158–172. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, L.; Zhou, H.; Jiang, H.; Yu, L.; Zeng, S. Stereoselective Binding of Chiral Drugs to Plasma Proteins. Acta Pharmacol. Sin. 2013, 34, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Volland, C.; Sun, H.; Dammeyer, J.; Benet, L.Z. Stereoselective Degradation of the Fenoprofen Acyl Glucuronide Enantiomers and Irreversible Binding to Plasma Protein. Drug Metab. Dispos. 1991, 19, 1080–1086. [Google Scholar]
- Kawase, A.; Yamamoto, T.; Egashira, S.; Iwaki, M. Stereoselective Inhibition of Methotrexate Excretion by Glucuronides of Nonsteroidal Anti-Inflammatory Drugs via Multidrug Resistance Proteins 2 and 4. J. Pharmacol. Exp. Ther. 2016, 356, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, M.; Bischer, A.; Nguyen, A.C.; McDonagh, A.F.; Benet, L.Z. Stereoselective Disposition of Naproxen Glucuronide in the Rat. Drug Metab. Dispos. 1995, 23, 1099–1103. [Google Scholar] [PubMed]
- Nozaki, Y.; Kusuhara, H.; Kondo, T.; Iwaki, M.; Shiroyanagi, Y.; Nakayama, H.; Horita, S.; Nakazawa, H.; Okano, T.; Sugiyama, Y. Species Difference in the Inhibitory Effect of Nonsteroidal Anti-Inflammatory Drugs on the Uptake of Methotrexate by Human Kidney Slices. J. Pharmacol. Exp. 2007, 322, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, M.; Ogiso, T.; Inagawa, S.; Kakehi, K. In Vitro Regioselective Stability of Beta-1-O- and 2-O-Acyl Glucuronides of Naproxen and Their Covalent Binding to Human Serum Albumin. J. Pharm. Sci. 1999, 88, 52–57. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawase, A.; Yamashita, R.; Yoshizato, T.; Yoshikawa, M.; Shimada, H.; Iwaki, M. Stereoselective Covalent Adduct Formation of Acyl Glucuronide Metabolite of Nonsteroidal Anti-Inflammatory Drugs with UDP-Glucuronosyltransferase. Int. J. Mol. Sci. 2022, 23, 4724. https://doi.org/10.3390/ijms23094724
Kawase A, Yamashita R, Yoshizato T, Yoshikawa M, Shimada H, Iwaki M. Stereoselective Covalent Adduct Formation of Acyl Glucuronide Metabolite of Nonsteroidal Anti-Inflammatory Drugs with UDP-Glucuronosyltransferase. International Journal of Molecular Sciences. 2022; 23(9):4724. https://doi.org/10.3390/ijms23094724
Chicago/Turabian StyleKawase, Atsushi, Rio Yamashita, Tsubasa Yoshizato, Mashiro Yoshikawa, Hiroaki Shimada, and Masahiro Iwaki. 2022. "Stereoselective Covalent Adduct Formation of Acyl Glucuronide Metabolite of Nonsteroidal Anti-Inflammatory Drugs with UDP-Glucuronosyltransferase" International Journal of Molecular Sciences 23, no. 9: 4724. https://doi.org/10.3390/ijms23094724
APA StyleKawase, A., Yamashita, R., Yoshizato, T., Yoshikawa, M., Shimada, H., & Iwaki, M. (2022). Stereoselective Covalent Adduct Formation of Acyl Glucuronide Metabolite of Nonsteroidal Anti-Inflammatory Drugs with UDP-Glucuronosyltransferase. International Journal of Molecular Sciences, 23(9), 4724. https://doi.org/10.3390/ijms23094724