
Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
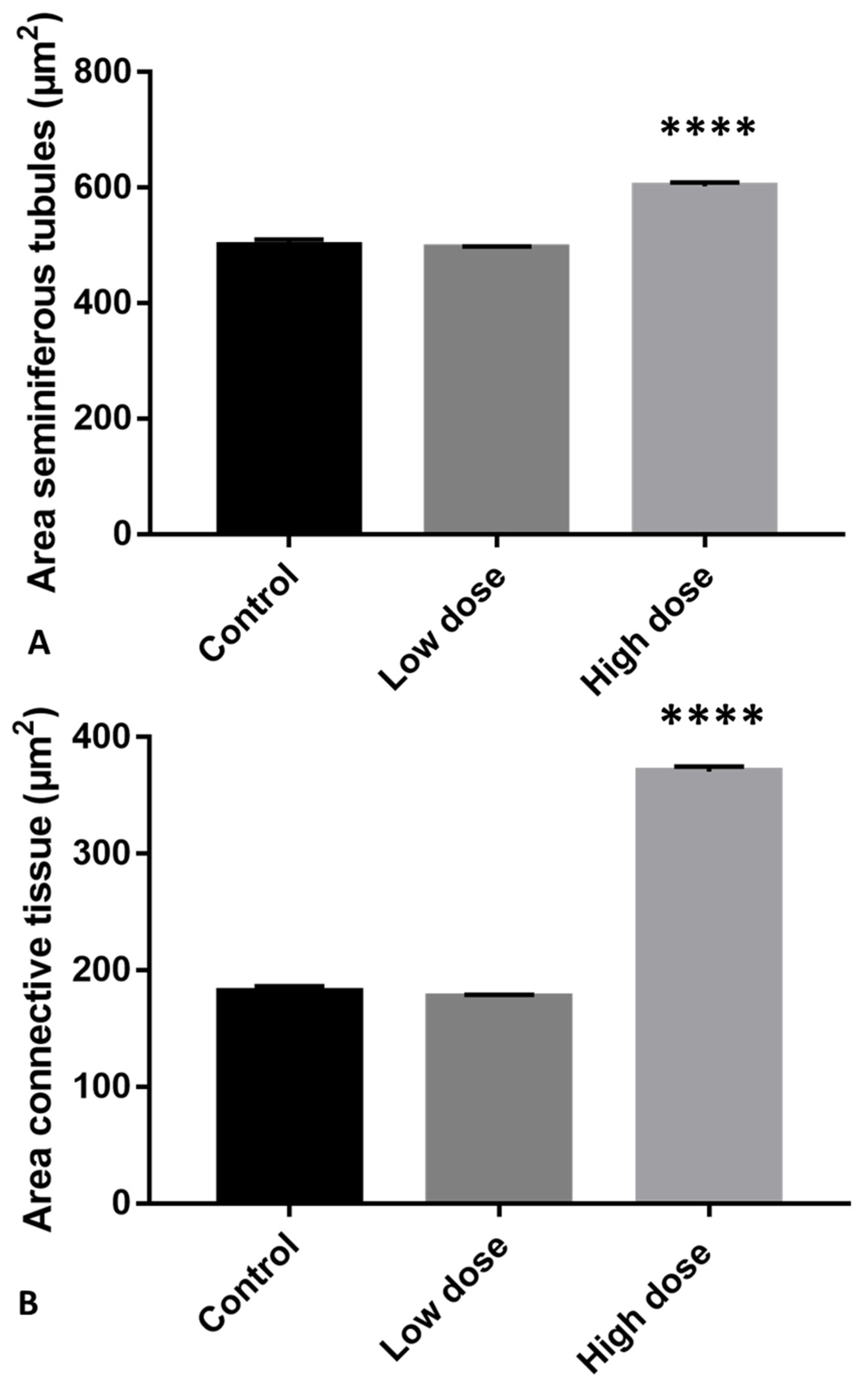
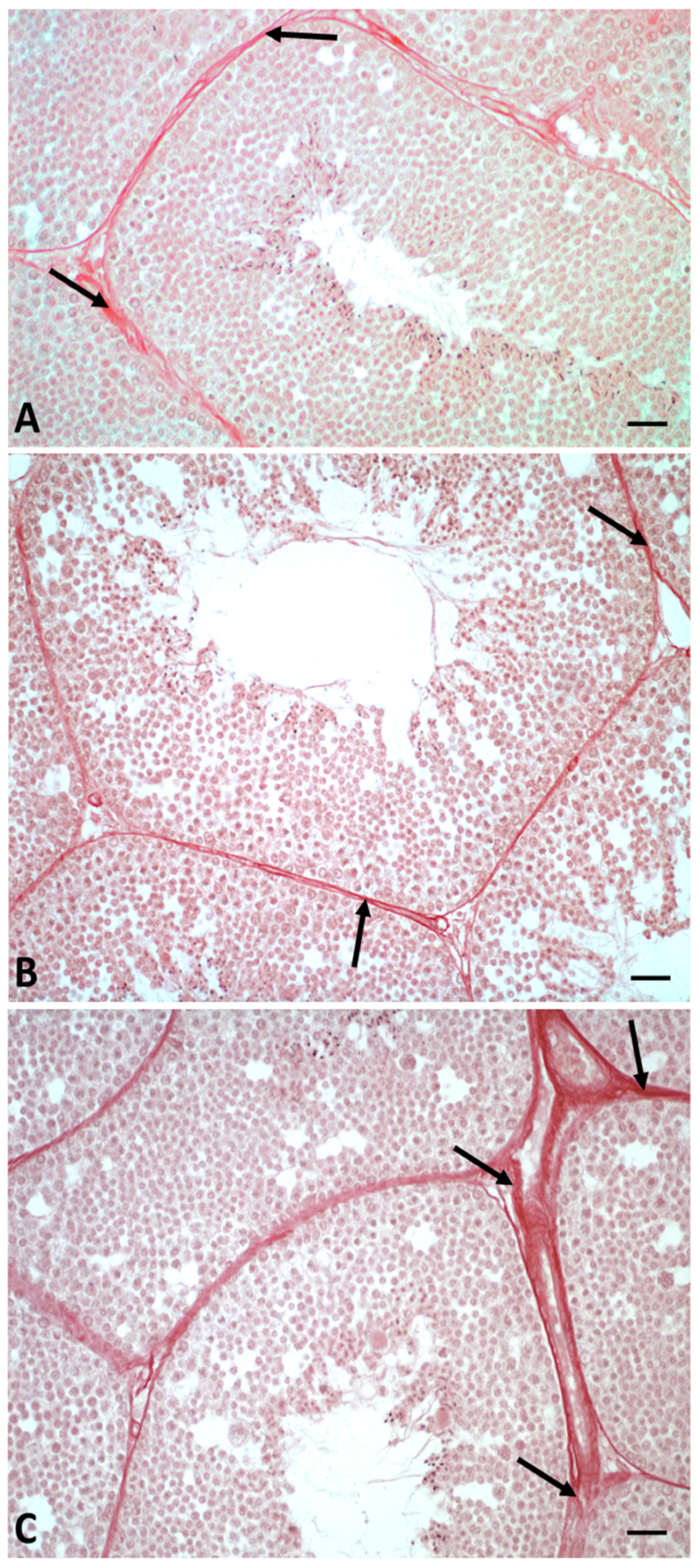
2.1. Morphological Analysis of Testis
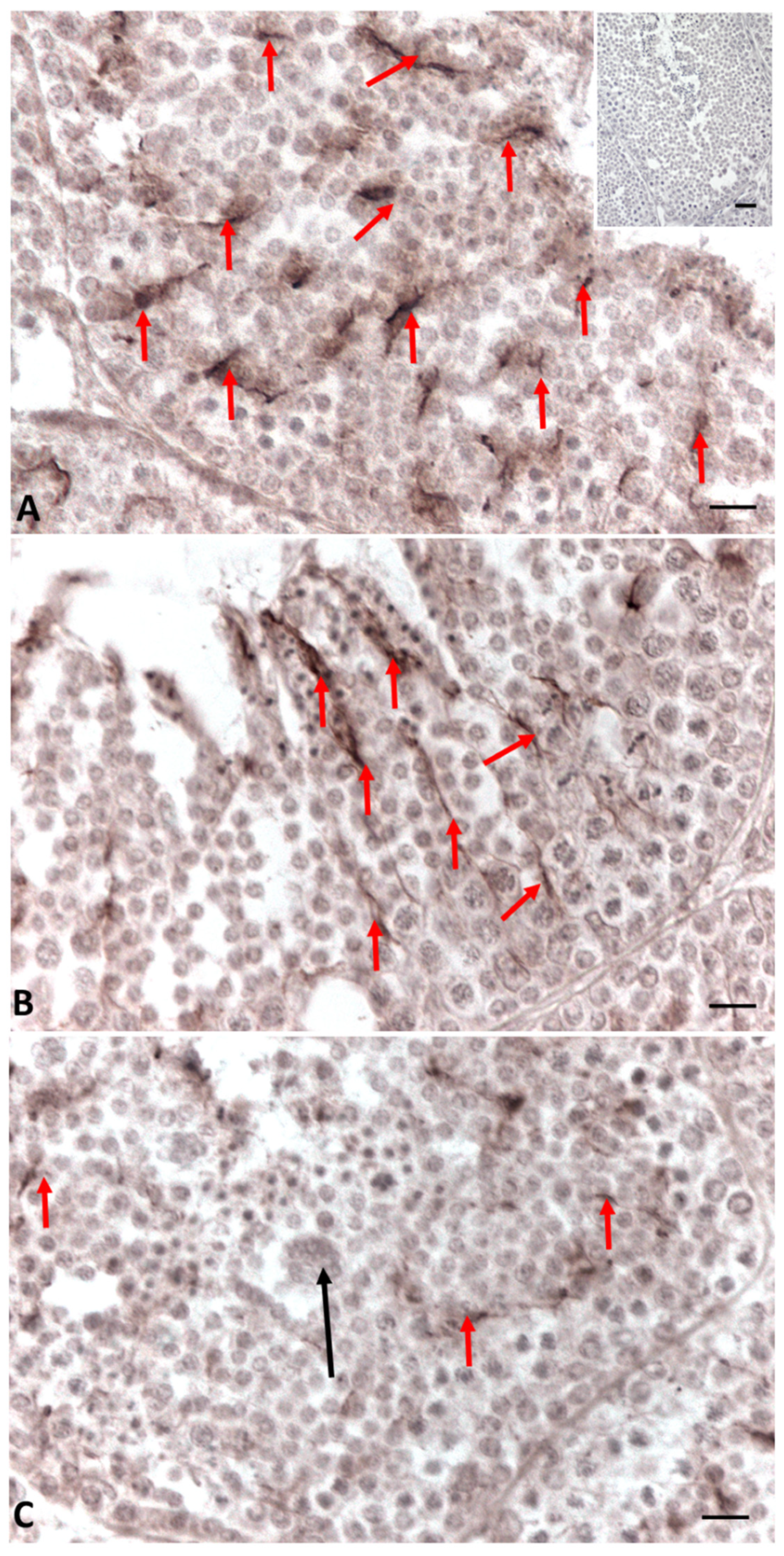
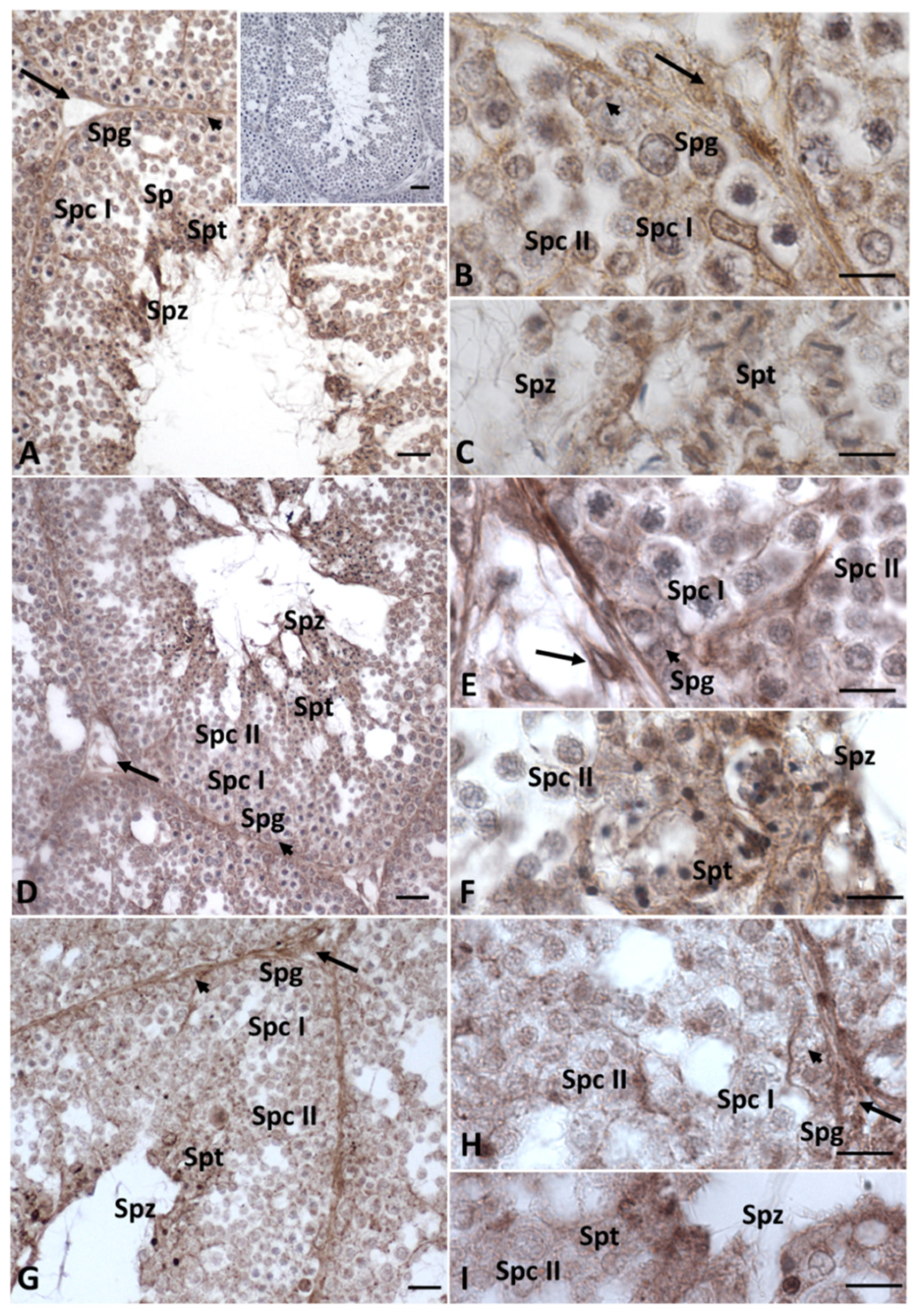
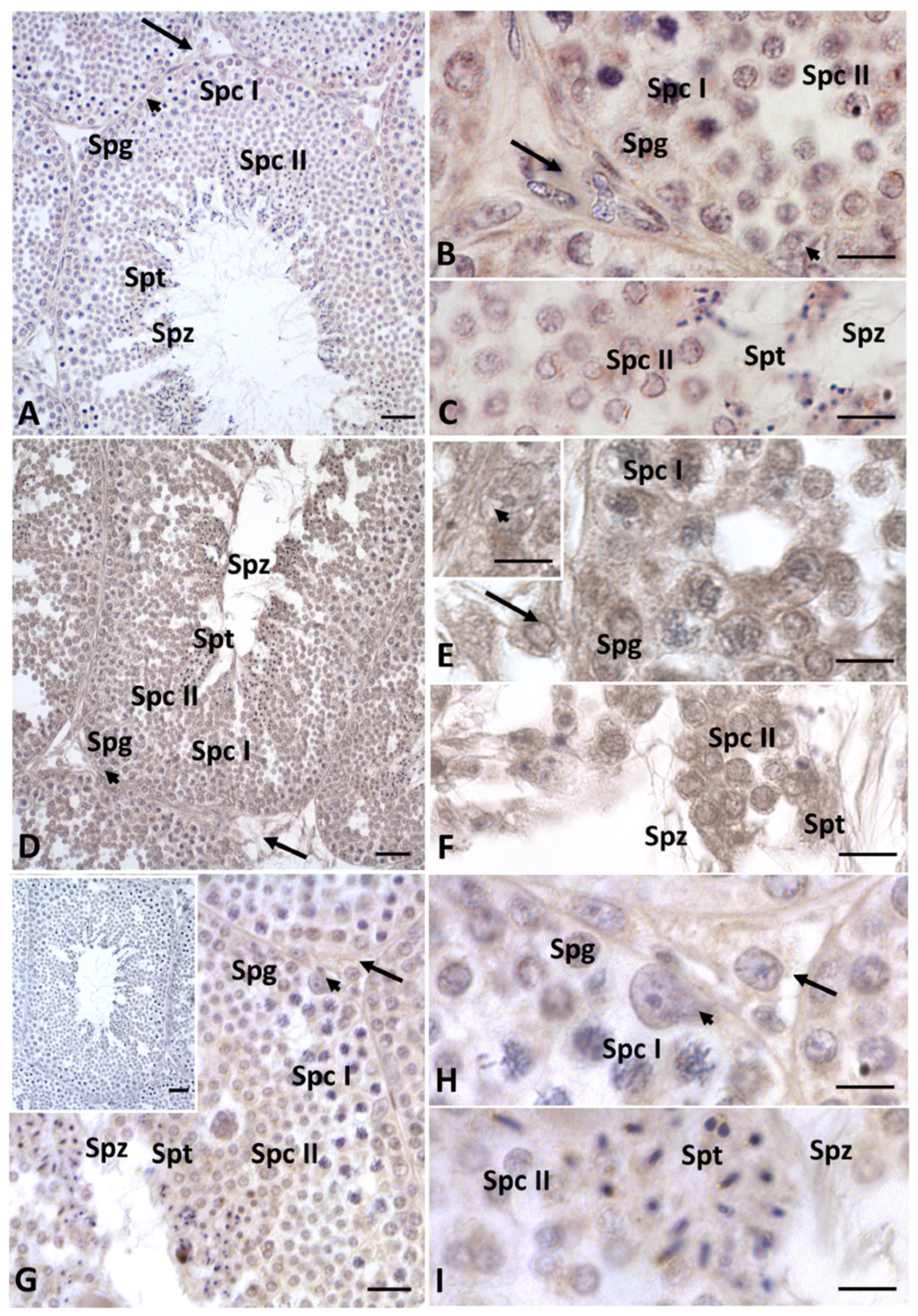
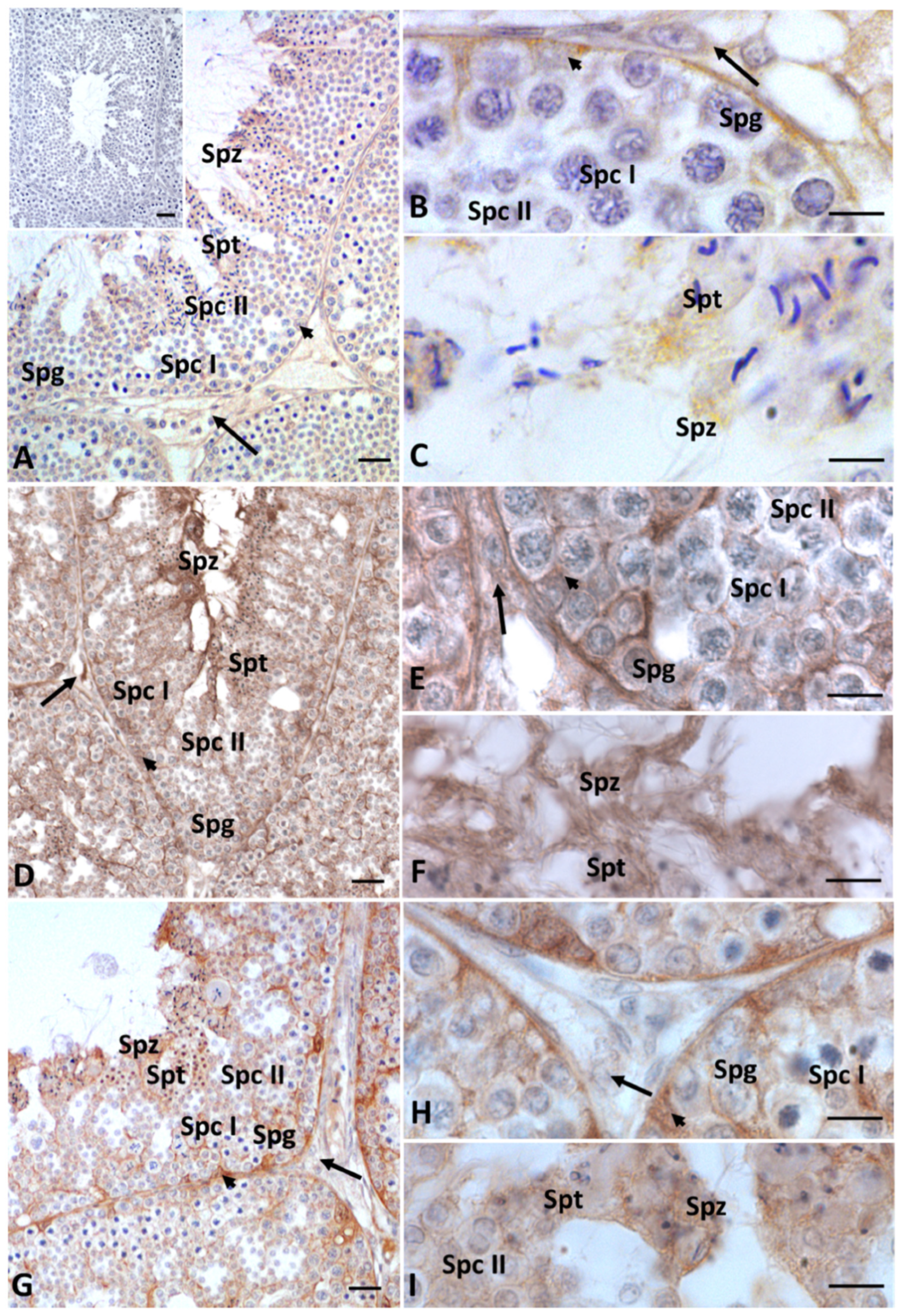
2.2. Immunohistochemistry for Cx43, Steroidogenic Enzymes, Erα and Erβ in Testis Sections
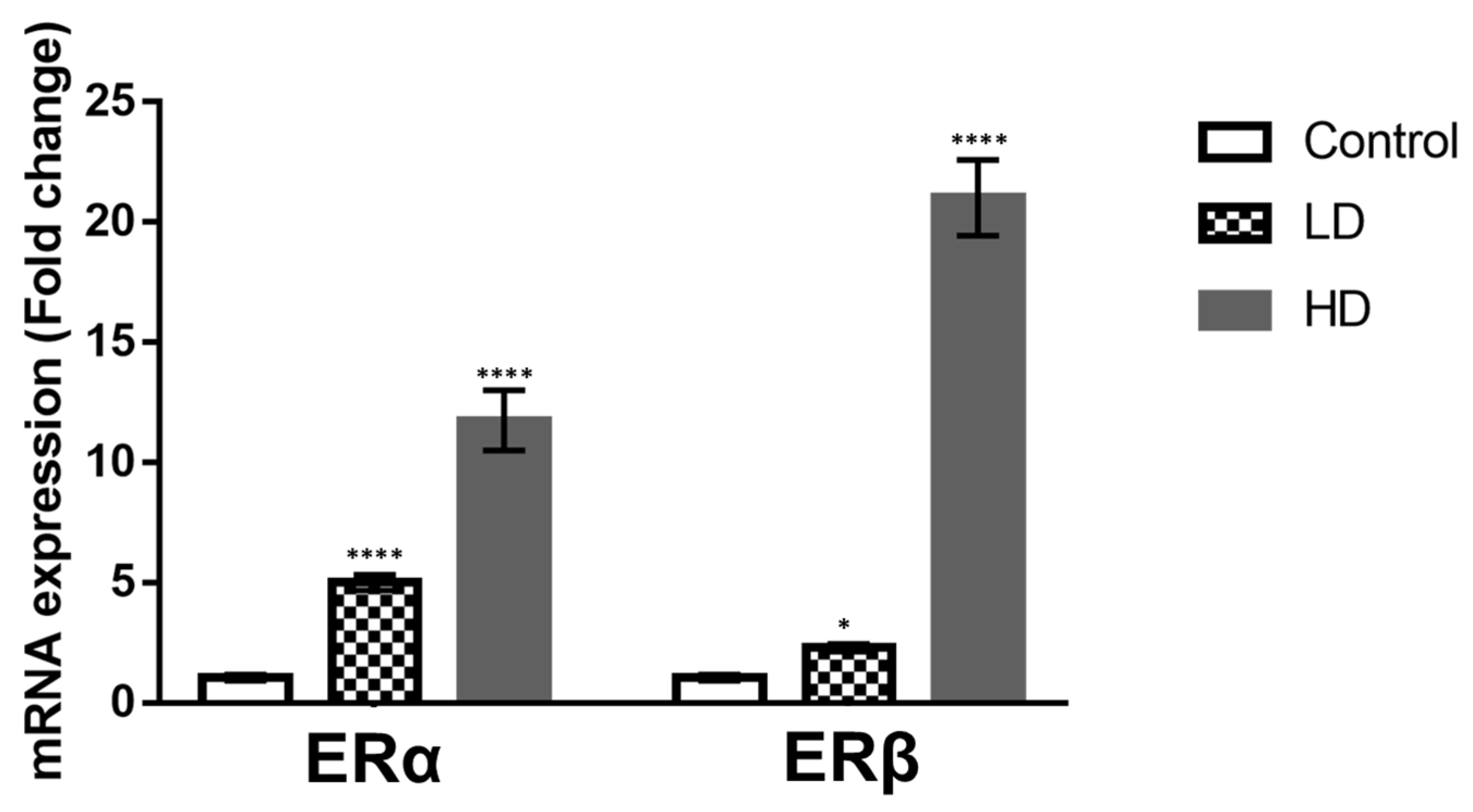
2.3. Gene Expression of ERα and ERβ in Testis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Experimental Design
5.2. Histological and Morphometric Analysis
5.3. Immunoistochemistry
5.4. RNA Isolation and Real-Time Polymerase Chain Reactions
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Griffin, J.; Boudreaux, J.; Miller, D. Herbicides As Harvest Aids. Weed Sci. 2010, 58, 355–358. [Google Scholar] [CrossRef]
- Bellaloui, N.; Bruns, H.A.; Abbas, H.K.; Fisher, D.K.; Mengistu, A. Effects of Harvest-Aids on Seed Nutrition in Soybean under Midsouth USA Conditions. Plants 2020, 9, 1007. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. Glyphosate: Uses Other Than in Glyphosate-Resistant Crops, Mode of Action, Degradation in Plants, and Effects on Non-target Plants and Agricultural Microbes. Rev. Environ. Contam. Toxicol. 2021, 255, 1–65. [Google Scholar] [CrossRef]
- Shumilina, E.; Andreasen, C.; Bitarafan, Z.; Dikiy, A. Determination of Glyphosate in Dried Wheat by 1H-NMR Spectroscopy. Molecules 2020, 25, 1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Kumar, V.; Gill, J.P.K.; Datta, S.; Singh, S.; Dhaka, V.; Kapoor, D.; Wani, A.B.; Dhanjal, D.S.; Kumar, M.; et al. Herbicide Glyphosate: Toxicity and Microbial Degradation. Int. J. Environ. Res. Public Health 2020, 17, 7519. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ 2018, 616–617, 255–268. [Google Scholar] [CrossRef]
- Carles, L.; Gardon, H.; Joseph, L.; Sanchís, J.; Farré, M.; Artigas, J. Meta-analysis of glyphosate contamination in surface waters and dissipation by biofilms. Environ. Int. 2019, 124, 284–293. [Google Scholar] [CrossRef]
- Bai, H.; Ogbourne, S.M. Glyphosate: Environmental contamination, toxicity and potential risks to human health via food contamination. Environ. Sci. Pollut. Res. Int. 2016, 23, 18988–19001. [Google Scholar] [CrossRef]
- Niemann, L.; Sieke, C.; Pfeil, R.; Solecki, R. A critical review of glyphosate findings in human urine samples and comparison with the exposure of operators and consumers. J. Verbr. Lebensm. 2015, 10, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Gholami-Seyedkolaei, S.J.; Mirvaghefi, A.; Farahmand, H.; Kosari, A. Effect of a glyphosate-based herbicide in Cyprinus carpio: Assessment of acetylcholinesterase activity, hematological responses and serum biochemical parameters. Ecotoxicol. Environ. Saf. 2013, 98, 135–141. [Google Scholar] [CrossRef]
- Webster, T.M.U.; Santos, E.M. Global transcriptomic profiling demonstrates induction of oxidative stress and of compensatory cellular stress responses in brown trout exposed to glyphosate and Roundup. BMC Genom. 2015, 16, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, N.; Reichenbecher, W.; Teichmann, H.; Tappeser, B.; Lötters, S. Questions concerning the potential impact of glyphosate-based herbicides on amphibians. Environ. Toxicol. Chem. 2013, 32, 1688–1700. [Google Scholar] [CrossRef] [PubMed]
- Herek, J.S.; Vargas, L.; Trindade, S.A.R.; Rutkoski, C.F.; Macagnan, N.; Hartmann, P.A.; Hartmann, M.T. Genotoxic effects of glyphosate on Physalaemus tadpoles. Environ. Toxicol. Pharmacol. 2021, 81, 103516. [Google Scholar] [CrossRef] [PubMed]
- Flach, H.; Lenz, A.; Pfeffer, S.; Kühl, M.; Kühl, S.J. Impact of glyphosate-based herbicide on early embryonic development of the amphibian Xenopus laevis. Aquat. Toxicol. 2022, 244, 106081. [Google Scholar] [CrossRef]
- Latorre, M.A.; López González, E.C.; Larriera, A.; Poletta, G.L.; Siroski, P.A. Effects of in vivo exposure to Roundup® on immune system of Caiman latirostris. J. Immunotoxicol. 2013, 10, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Siroski, P.A.; Poletta, G.L.; Latorre, M.A.; Merchant, M.E.; Ortega, H.; Mudry, M.D. Immunotoxicity of commercial-mixed glyphosate in broad snouted caiman (Caiman latirostris). Chem. Biol. Interact. 2016, 244, 64–70. [Google Scholar] [CrossRef]
- Mestre, A.P.; Amavet, P.S.; Vanzetti, A.I.; Moleón, M.S.; Marcó, M.V.P.; Poletta, G.L.; Siroski, P.A. Effects of cypermethrin (pyrethroid), glyphosate and chlorpyrifos (organophosphorus) on the endocrine and immune system of Salvator merianae (Argentine tegu). Ecotoxicol. Environ. Saf. 2019, 169, 61–67. [Google Scholar] [CrossRef]
- Ruuskanen, S.; Rainio, M.J.; Gómez-Gallego, C.; Selenius, O.; Salminen, S.; Collado, M.C.; Saikkonen, K.; Saloniemi, I.; Helander, M. Glyphosate-based herbicides influence antioxidants, reproductive hormones and gut microbiome but not reproduction: A long-term experiment in an avian model. Environ. Pollut. 2020, 266 Pt 1, 115108. [Google Scholar] [CrossRef]
- Carlsen, E.; Giwercman, A.; Keiding, N.; Skakkebaek, N.E. Evidence for decreasing quality of semen during past 50 years. BMJ 1992, 305, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Merzenich, H.; Zeeb, H.; Blettner, M. Decreasing sperm quality: A global problem? BMC Public Health 2010, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T. Diet and sperm quality: Nutrients, foods and dietary patterns. Reprod. Biol. 2019, 19, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Marra, F.; Moriello, C.; Prisco, M.; Notari, T.; Trifuoggi, M.; Giarra, A.; Bosco, L.; Montano, L.; Piscopo, M. Molecular Alterations in Spermatozoa of a Family Case Living in the Land of Fires—A First Look at Possible Transgenerational Effects of Pollutants. Int. J. Mol. Sci. 2020, 21, 6710. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Akiri, O.F.; Durojaiye, M.A.; Adenike, A. Combined effects of repeated administration of Bretmont Wipeout (glyphosate) and Ultrazin (atrazine) on testosterone, oxidative stress and sperm quality of Wistar rats. Toxicol. Mech. Methods 2015, 25, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Ji, Y.; Song, X.; Guo, H.; Han, L.; Zhang, F.; Liu, X.; Zhang, H.; Zhu, B.; Xu, M. Effects of glyphosate exposure on sperm concentration in rodents: A systematic review and meta-analysis. Environ. Toxicol. Pharmacol. 2017, 55, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Mele, E.; Viggiano, A.; Nori, S.L.; Meccariello, R.; Santoro, A. Pleiotropic Outcomes of Glyphosate Exposure: From Organ Damage to Effects on Inflammation, Cancer, Reproduction and Development. Int. J. Mol. Sci. 2021, 22, 12606. [Google Scholar] [CrossRef] [PubMed]
- Gorga, A.; Rindone, G.M.; Centola, C.L.; Sobarzo, C.M.; Pellizzari, E.H.; Camberos, M.D.C.; Marín-Briggiler, C.I.; Cohen, D.J.; Riera, M.F.; Galardo, M.N.; et al. Low Doses of Glyphosate/Roundup Alter Blood-Testis Barrier Integrity in Juvenile Rats. Front. Endocrinol. 2021, 12, 615678. [Google Scholar] [CrossRef]
- Liu, J.-B.; Li, Z.-F.; Lu, L.; Wang, Z.-Y.; Wang, L. Glyphosate damages blood-testis barrier via NOX1-triggered oxidative stress in rats: Long-term exposure as a potential risk for male reproductive health. Environ. Int. 2022, 159, 107038. [Google Scholar] [CrossRef]
- Avdatek, F.; Birdane, Y.O.; Türkmen, R.; Demirel, H.H. Ameliorative effect of resveratrol on testicular oxidative stress, spermatological parameters and DNA damage in glyphosate-based herbicide-exposed rats. Andrologia 2018, 50, 13036. [Google Scholar] [CrossRef]
- Walsh, L.P.; McCormick, C.; Martin, C.; Stocco, D.M. Roundup inhibits steroidogenesis by disrupting steroidogenic acute regulatory (StAR) protein expression. Environ. Health Perspect. 2000, 108, 769–776. [Google Scholar] [CrossRef]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef]
- Cassault-Meyer, E.; Gress, S.; Séralini, G.É.; Galeraud-Denis, I. An acute exposure to glyphosate-based herbicide alters aromatase levels in testis and sperm nuclear quality. Environ. Toxicol. Pharmacol. 2014, 38, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Carreau, S.; Bouraima-Lelong, H.; Delalande, C. Estrogens: New players in spermatogenesis. Reprod. Biol. Endocrinol. 2011, 1, 90–94. [Google Scholar] [CrossRef]
- Rosati, L.; Agnese, M.; Di Fiore, M.M.; Andreuccetti, P.; Prisco, M. P450 aromatase: A key enzyme in the spermatogenesis of the Italian wall lizard Podarcis sicula. J. Exp. Biol. 2016, 219, 2402–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, L.; Prisco, M.; Di Lorenzo, M.; de Falco, M.; Andreuccetti, P. Immunolocalization of aromatase P450 in the epididymis of Podarcis sicula and Rattus rattus. Eur. J. Histochem. 2020, 64, 3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razi, M.; Najafi, G.; Feyzi, S.; Karimi, A.; Shahmohamadloo, S.; Nejati, V. Histological and histochemical effects of Glyphosate on testicular tissue and function. Iran. J. Reprod. Med. 2012, 10, 181–192. [Google Scholar] [PubMed]
- Oliveira, A.G.; Telles, L.F.; Hess, R.A.; Mahecha, G.A.; Oliveira, C.A. Effects of the herbicide Roundup on the epididymal region of drakes Anas platyrhynchos. Reprod. Toxicol. 2007, 23, 182–191. [Google Scholar] [CrossRef]
- Chaki, S.P.; Misro, M.M.; Gautam, D.K.; Kaushik, M.; Ghosh, D.; Chainy, G.B. Estradiol treatment induces testicular oxidative stress and germ cell apoptosis in rats. Apoptosis 2006, 11, 1427–1437. [Google Scholar] [CrossRef]
- de Falco, M.; Forte, M.; Laforgia, V. Estrogenic and anti-androgenic endocrine disrupting chemicals and their impact on the male reproductive system. Front. Environ. Sci. 2015, 3, 3. [Google Scholar] [CrossRef]
- Verderame, M.; Limatola, E.; Scudiero, R. Metallothionein expression and synthesis in the testis of the lizard Podarcis sicula under natural conditions and following estrogenic exposure. Eur. J. Histochem. 2017, 61, 2777. [Google Scholar] [CrossRef] [Green Version]
- González-Rojo, S.; Lombó, M.; Fernández-Díez, C.; Herráez, M.P. Male exposure to bisphenol a impairs spermatogenesis and triggers histone hyperacetylation in zebrafish testes. Environ. Pollut. 2019, 248, 368–379. [Google Scholar] [CrossRef]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.C.; Séralini, G.E. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Ingaramo, P.; Alarcón, R.; Muñoz-de-Toro, M.; Luque, E.H. Are glyphosate and glyphosate-based herbicides endocrine disruptors that alter female fertility? Mol. Cell. Endocrinol. 2020, 518, 110934. [Google Scholar] [CrossRef]
- Scudiero, R.; Verderame, M.; Motta, C.M.; Simoniello, P. Unravelling the Role of Metallothionein on Development, Reproduction and Detoxification in the Wall Lizard Podarcis sicula. Int. J. Mol. Sci. 2017, 18, 1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.R.; Campbell, T.S. Lizard contaminant data for ecological risk assessment. Rev. Environ. Contam. Toxicol. 2000, 165, 39–116. [Google Scholar] [CrossRef]
- Verderame, M.; Limatola, E.; Scudiero, R. Estrogenic contamination by manure fertilizer in organic farming: A case study with the lizard Podarcis sicula. Ecotoxicology 2016, 25, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verderame, M.; Scudiero, R. Health status of the lizard Podarcis siculus (Rafinesque-Schmaltz, 1810) subject to different anthropogenic pressures. CR Biol. 2019, 342, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Piovia-Scott, J.; Yang, L.H.; Wright, A.N.; Spiller, D.A.; Schoener, T.W. Pulsed seaweed subsidies drive sequential shifts in the effects of lizard predators on island food webs. Ecol. Lett. 2019, 22, 1850–1859. [Google Scholar] [CrossRef]
- Silva, J.M.; Navoni, J.A.; Freire, E.M.X. Lizards as model organisms to evaluate environmental contamination and biomonitoring. Environ. Monit. Assess. 2020, 192, 454. [Google Scholar] [CrossRef]
- Verderame, M.; Scudiero, R. How Glyphosate Impairs Liver Condition in the Field Lizard Podarcis siculus (Rafinesque-Schmaltz, 1810): Histological and Molecular Evidence. Biomed. Res. Int. 2019, 2019, 4746283. [Google Scholar] [CrossRef] [Green Version]
- Rosati, L.; Agnese, M.; Di Lorenzo, M.; Barra, T.; Valiante, S.; Prisco, M. Spermatogenesis and regulatory factors in the wall lizard Podarcis sicula. Gen. Comp. Endocrinol. 2020, 298, 113579. [Google Scholar] [CrossRef]
- Benedetti, A.L.; Vituri, C.D.L.; Trentin, A.; Domingues, M.A.C.; Alvarez-Silva, M. The effects of sub-chronic exposure of Wistar rats to the herbicide Glyphosate-Biocarb. Toxicol. Lett. 2004, 153, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, G.A.; Delconte, M.B.; Gomez, A.L.; Ingaramo, P.I.; Bosquiazzo, V.L.; Luque, E.H.; Muñoz-de-Toro, M.; Kass, L. Postnatal exposure to a glyphosate-based herbicide modifies mammary gland growth and development in Wistar male rats. Food Chem. Toxicol. 2018, 118, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peillex, C.; Pelletier, M. The impact and toxicity of glyphosate and glyphosate-based herbicides on health and immunity. J. Immunotoxicol. 2020, 17, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Brehm, R.; Bergmann, M.; Cooke, P.S. Role of connexin 43 in Sertoli cells of testis. Ann. N. Y. Acad. Sci. 2007, 1120, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr. Rev. 2004, 25, 747–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondet, A.; Martin, G.; Durand, P.; Perrard, M.H. Low concentrations of glyphosate alone affect the pubertal male rat meiotic step: An in vitro study. Toxicol. In Vitro 2022, 79, 105291. [Google Scholar] [CrossRef]
- Fietz, D.; Pilatz, A.; Diemer, T.; Wagenlehner, F.; Bergmann, M.; Schuppe, H.C. Excessive unilateral proliferation of spermatogonia in a patient with non-obstructive azoospermia-adverse effect of clomiphene citrate pre-treatment? Basic Clin. Androl. 2020, 30, 13. [Google Scholar] [CrossRef]
- Moinfar, Z.; Dambach, H.; Schoenebeck, B.; Förster, E.; Prochnow, N.; Faustmann, P.M. Estradiol Receptors Regulate Differential Connexin 43 Expression in F98 and C6 Glioma Cell Lines. PLoS ONE 2016, 11, 0150007. [Google Scholar] [CrossRef] [Green Version]
- Rosati, L.; Falvo, S.; Chieffi Baccari, G.; Santillo, A.; Di Fiore, M.M. The Aromatase-Estrogen System in the Testes of Non-Mammalian Vertebrates. Animals 2021, 11, 1763. [Google Scholar] [CrossRef]
- Falvo, S.; Rosati, L.; Di Fiore, M.M.; Di Giacomo Russo, F.; Chieffi Baccari, G.; Santillo, A. Proliferative and Apoptotic Pathways in the Testis of Quail Coturnix coturnix during the Seasonal Reproductive Cycle. Animals 2021, 11, 1729. [Google Scholar] [CrossRef]
- Mesnage, R.; Phedonos, A.; Biserni, M.; Arno, M.; Balu, S.; Corton, J.C.; Ugarte, R.; Antoniou, M.N. Evaluation of estrogen receptor alpha activation by glyphosate-based herbicide constituents. Food Chem. Toxicol. 2017, 108 Pt A, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Thongprakaisang, S.; Thiantanawat, A.; Rangkadilok, N.; Suriyo, T.; Satayavivad, J. Glyphosate induces human breast cancer cells growth via estrogen receptors. Food Chem. Toxicol. 2013, 59, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate. EFSA J. 2015, 13, 4302. [Google Scholar]
- Di Lorenzo, M.; Mileo, A.; Laforgia, V.; de Falco, M.; Rosati, L. Alkyphenol Exposure Alters Steroidogenesis in Male Lizard Podarcis siculus. Animals 2021, 11, 1003. [Google Scholar] [CrossRef] [PubMed]
- Verderame, M.; Limatola, E. Molecular identification of estrogen receptors (ER alpha and ER beta) and their differential expression during VTG synthesis in the liver of lizard Podarcis sicula. Gen. Comp. Endocrinol. 2010, 168, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verderame, M.; Chianese, T.; Rosati, L.; Scudiero, R. Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus. Int. J. Mol. Sci. 2022, 23, 4850. https://doi.org/10.3390/ijms23094850
Verderame M, Chianese T, Rosati L, Scudiero R. Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus. International Journal of Molecular Sciences. 2022; 23(9):4850. https://doi.org/10.3390/ijms23094850
Chicago/Turabian StyleVerderame, Mariailaria, Teresa Chianese, Luigi Rosati, and Rosaria Scudiero. 2022. "Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus" International Journal of Molecular Sciences 23, no. 9: 4850. https://doi.org/10.3390/ijms23094850
APA StyleVerderame, M., Chianese, T., Rosati, L., & Scudiero, R. (2022). Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus. International Journal of Molecular Sciences, 23(9), 4850. https://doi.org/10.3390/ijms23094850