
Translational Bioinformatics for Human Reproductive Biology Research: Examples, Opportunities and Challenges for a Future Reproductive Medicine

Abstract
:1. Introduction
2. Identification and Molecular Regulation of Biomarkers of Infertility and Reproductive Disease
2.1. Development of Male Reproductive and Infertility Biomarkers
2.2. Biomarks for Prostate Cancer
2.3. Biomarkers for Male Infertility
2.4. Development of Female Reproductive and Infertility Biomarkers
2.5. Hereditary Factors
2.6. Inflammatory Factors
2.7. Single-Cell Omics and Multi-Omics
3. Application and Development of AI in Reproductive Medicine
Limitations and Challenges of AI in Reproductive Medicine Applications
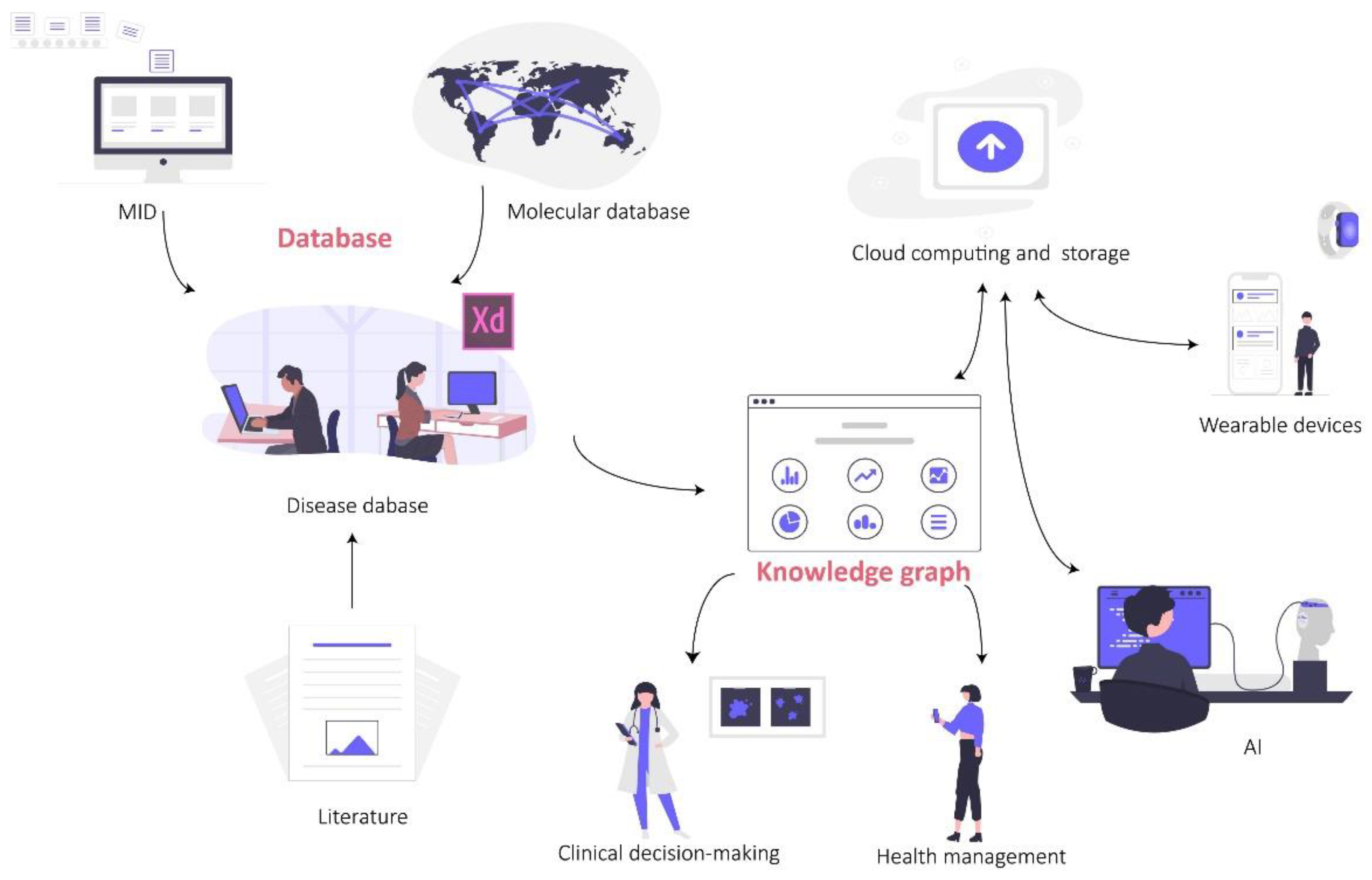
4. The Way from Reproduction Medical Database to Knowledge Graph
4.1. Status of Reproductive Medicine Database
4.2. Constructing Clinical Reproductive Explainable Knowledge Graph Based on Ontology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AI | Artificial intelligence | mRNA | Messenger RNA |
| ANN | Artificial neural network | NF | Nuclear factor |
| ART | Assisted reproductive technology | OA | Obstructive azoospermia |
| AZF | Azoospermia factor | PCA | Prostate cancer |
| BiTEs | Bispecific T cell engagers | PCOS | Polycystic ovary syndrome |
| CAR-T | Chimeric antigen receptor T | PCT | Procalcitonin |
| CAVD | Congenital absence of the vas deferens | PGT | Preimplantation genetic testing |
| CBAVD | Congenital Bilateral Absence of the Vas Deferens | POI | Premature ovarian insufficiency |
| CF | Cystic fibrosis | Prx4 | Peroxiredoxin 4 |
| cfDNA | Circulating free DNA | PSA | Prostate-specific antigen |
| CFTR | Cystic fibrosis transmembrane conductance regulator | STI | Sexually transmitted infection |
| chr | Chromosome | TBI | Translational bioinformatics |
| CNN | Convolutional neural networks | TCGA | The Cancer Genome Atlas |
| CNV | Copy number variation | TFI | Tubal factor infertility |
| CRP | C-reactive protein | TMAO | Trimethylamine-N-Oxide |
| DIE | Deep-infiltrating endometriosis | ToxRefDB | Toxicity Reference Database |
| DNN | Deep neural network | WGS | Whole genome sequencing |
| EMR | Electronic medical record | ML | Machine learning |
| ESTs | Expressed sequence tags | LR | Logistic regression |
| GWAS | Genome-wide association studies | DT | Decision trees |
| HOXB13 | Homeobox B13 | NB | Naive Bayes |
| HSSC | Human spermatogonial stem cells | RF | Random forests |
| IFN | Interferons | SVM | Support vector machines |
| IMIGC | International Male Infertility Genomics Consortium | Nnet | Neural networks |
| IVF | In vitro fertilization | BNN | Back propagation neural networks |
| lnRNA | Long non-coding RNA | GBDT | Gradient boosting decision trees |
| MID | Medical information database | XGBoost | Extreme gradient boosting |
| mpMRI | Multiparametric magnetic resonance imaging | SL | Super learners |
References
- Tenenbaum, J.D. Translational Bioinformatics: Past, Present, and Future. Genom. Proteom. Bioinform. 2016, 14, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oulas, A.; Minadakis, G.; Zachariou, M.; Sokratous, K.; Bourdakou, M.M.; Spyrou, G.M. Systems Bioinformatics: Increasing precision of computational diagnostics and therapeutics through network-based approaches. Brief. Bioinform. 2019, 20, 806–824. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.L.; Altman, R.B. Biomarkers: Delivering on the expectation of molecularly driven, quantitative health. Exp. Biol. Med. 2018, 243, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Zhang, W.; Yu, X.; Liu, X.; Li, M.; Liu, R.; Chen, L. Edge biomarkers for classification and prediction of phenotypes. Sci. China. Life Sci. 2014, 57, 1103–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Angulo, A.M.; Hennessy, B.T.; Mills, G.B. Future of personalized medicine in oncology: A systems biology approach. J. Clin. Oncol. 2010, 28, 2777–2783. [Google Scholar] [CrossRef]
- Toppari, J.; Larsen, J.C.; Christiansen, P.; Giwercman, A.; Grandjean, P.; Guillette, L.J., Jr.; Jegou, B.; Jensen, T.K.; Jouannet, P.; Keiding, N.; et al. Male reproductive health and environmental xenoestrogens. Environ. Health Perspect. 1996, 104 (Suppl. 4), 741–803. [Google Scholar]
- Das, L.; Parbin, S.; Pradhan, N.; Kausar, C.; Patra, S.K. Epigenetics of reproductive infertility. Front. Biosci. 2017, 9, 509–535. [Google Scholar]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; Korrovits, P.; Laan, M. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 2017, 32, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Guzick, D.S.; Overstreet, J.W.; Factor-Litvak, P.; Brazil, C.K.; Nakajima, S.T.; Coutifaris, C.; Carson, S.A.; Cisneros, P.; Steinkampf, M.P.; Hill, J.A.; et al. Sperm morphology, motility, and concentration in fertile and infertile men. N. Engl. J. Med. 2001, 345, 1388–1393. [Google Scholar] [CrossRef]
- Viale, P.H. The American Cancer Society’s Facts & Figures: 2020 Edition. J. Adv. Pr. Oncol. 2020, 11, 135–136. [Google Scholar]
- Conti, D.V.; Darst, B.F.; Moss, L.C.; Saunders, E.J.; Sheng, X.; Chou, A.; Schumacher, F.R.; Olama, A.A.A.; Benlloch, S.; Dadaev, T.; et al. Trans-ancestry genome-wide association meta-analysis of prostate cancer identifies new susceptibility loci and informs genetic risk prediction. Nat. Genet. 2021, 53, 65–75. [Google Scholar] [CrossRef]
- Ye, C.; Wang, H.; Li, Z.; Xia, C.; Yuan, S.; Yan, R.; Yang, X.; Ma, T.; Wen, X.; Yang, D. Comprehensive data analysis of genomics, epigenomics, and transcriptomics to identify specific biomolecular markers for prostate adenocarcinoma. Transl. Androl. Urol. 2021, 10, 3030–3045. [Google Scholar] [CrossRef]
- Bochum, S.; Paiss, T.; Vogel, W.; Herkommer, K.; Hautmann, R.; Haeussler, J. Confirmation of the prostate cancer susceptibility locus HPCX in a set of 104 German prostate cancer families. Prostate 2002, 52, 12–19. [Google Scholar] [CrossRef]
- Xu, J.; Meyers, D.; Freije, D.; Isaacs, S.; Wiley, K.; Nusskern, D.; Ewing, C.; Wilkens, E.; Bujnovszky, P.; Bova, G.S.; et al. Evidence for a prostate cancer susceptibility locus on the X chromosome. Nat. Genet. 1998, 20, 175–179. [Google Scholar] [CrossRef]
- Valeri, A.; Briollais, L.; Azzouzi, R.; Fournier, G.; Mangin, P.; Berthon, P.; Cussenot, O.; Demenais, F. Segregation analysis of prostate cancer in France: Evidence for autosomal dominant inheritance and residual brother-brother dependence. Ann. Hum. Genet. 2003, 67, 125–137. [Google Scholar] [CrossRef]
- Kibel, A.S.; Faith, D.A.; Bova, G.S.; Isaacs, W.B. Xq27-28 deletions in prostate carcinoma. Genes Chromosom. Cancer 2003, 37, 381–388. [Google Scholar] [CrossRef]
- Ren, S.; Wei, G.H.; Liu, D.; Wang, L.; Hou, Y.; Zhu, S.; Peng, L.; Zhang, Q.; Cheng, Y.; Su, H.; et al. Whole-genome and Transcriptome Sequencing of Prostate Cancer Identify New Genetic Alterations Driving Disease Progression. Eur. Urol. 2018, 73, 322–339. [Google Scholar] [CrossRef]
- Harlemon, M.; Ajayi, O.; Kachambwa, P.; Kim, M.S.; Simonti, C.N.; Quiver, M.H.; Petersen, D.C.; Mittal, A.; Fernandez, P.W.; Hsing, A.W.; et al. A Custom Genotyping Array Reveals Population-Level Heterogeneity for the Genetic Risks of Prostate Cancer and Other Cancers in Africa. Cancer Res. 2020, 80, 2956–2966. [Google Scholar] [CrossRef]
- Li, J.; Xu, C.; Lee, H.J.; Ren, S.; Zi, X.; Zhang, Z.; Wang, H.; Yu, Y.; Yang, C.; Gao, X.; et al. A genomic and epigenomic atlas of prostate cancer in Asian populations. Nature 2020, 580, 93–99. [Google Scholar] [CrossRef]
- Gerhauser, C.; Favero, F.; Risch, T.; Simon, R.; Feuerbach, L.; Assenov, Y.; Heckmann, D.; Sidiropoulos, N.; Waszak, S.M.; Hubschmann, D.; et al. Molecular Evolution of Early-Onset Prostate Cancer Identifies Molecular Risk Markers and Clinical Trajectories. Cancer Cell 2018, 34, 996–1011.e8. [Google Scholar] [CrossRef] [Green Version]
- Rubin, M.A.; Girelli, G.; Demichelis, F. Genomic Correlates to the Newly Proposed Grading Prognostic Groups for Prostate Cancer. Eur. Urol. 2016, 69, 557–560. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Zhao, S.G.; Chang, S.L.; Tomlins, S.A.; Erho, N.; Sboner, A.; Schiewer, M.J.; Spratt, D.E.; Kothari, V.; Klein, E.A.; et al. Patient-Level DNA Damage and Repair Pathway Profiles and Prognosis After Prostatectomy for High-Risk Prostate Cancer. JAMA Oncol. 2016, 2, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Cooperberg, M.R.; Erho, N.; Chan, J.M.; Feng, F.Y.; Fishbane, N.; Zhao, S.G.; Simko, J.P.; Cowan, J.E.; Lehrer, J.; Alshalalfa, M.; et al. The Diverse Genomic Landscape of Clinically Low-risk Prostate Cancer. Eur. Urol. 2018, 74, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research, N. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar]
- Kamoun, A.; Cancel-Tassin, G.; Fromont, G.; Elarouci, N.; Armenoult, L.; Ayadi, M.; Irani, J.; Leroy, X.; Villers, A.; Fournier, G.; et al. Comprehensive molecular classification of localized prostate adenocarcinoma reveals a tumour subtype predictive of non-aggressive disease. Ann. Oncol. 2018, 29, 1814–1821. [Google Scholar] [CrossRef]
- Burns, D.; Anokian, E.; Saunders, E.J.; Bristow, R.G.; Fraser, M.; Reimand, J.; Schlomm, T.; Sauter, G.; Brors, B.; Korbel, J.; et al. Rare Germline Variants Are Associated with Rapid Biochemical Recurrence After Radical Prostate Cancer Treatment: A Pan Prostate Cancer Group Study. Eur. Urol. 2022, 82, 201–211. [Google Scholar] [CrossRef]
- Castro, E.; Goh, C.; Olmos, D.; Saunders, E.; Leongamornlert, D.; Tymrakiewicz, M.; Mahmud, N.; Dadaev, T.; Govindasami, K.; Guy, M.; et al. Germline BRCA mutations are associated with higher risk of nodal involvement, distant metastasis, and poor survival outcomes in prostate cancer. J. Clin. Oncol. 2013, 31, 1748–1757. [Google Scholar] [CrossRef] [Green Version]
- Segal, N.; Ber, Y.; Benjaminov, O.; Tamir, S.; Yakimov, M.; Kedar, I.; Rosenbaum, E.; Sela, S.; Ozalvo, R.; Shavit-Grievink, L.; et al. Imaging-based prostate cancer screening among BRCA mutation carriers-results from the first round of screening. Ann. Oncol. 2020, 31, 1545–1552. [Google Scholar] [CrossRef]
- Calvo Chozas, A.; Mahjani, B.; Ronnegard, L. Family history of breast cancer is associated with elevated risk of prostate cancer: Evidence for shared genetic risks. Hum. Hered. 2021, 87, 12–20. [Google Scholar] [CrossRef]
- Mottet, N.; van den Bergh, R.C.N.; Briers, E.; Van den Broeck, T.; Cumberbatch, M.G.; De Santis, M.; Fanti, S.; Fossati, N.; Gandaglia, G.; Gillessen, S.; et al. EAU-EANM-ESTRO-ESUR-SIOG Guidelines on Prostate Cancer-2020 Update. Part 1: Screening, Diagnosis, and Local Treatment with Curative Intent. Eur. Urol. 2021, 79, 243–262. [Google Scholar] [CrossRef]
- Dombernowsky, S.L.; Weischer, M.; Allin, K.H.; Bojesen, S.E.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. Risk of cancer by ATM missense mutations in the general population. J. Clin. Oncol. 2008, 26, 3057–3062. [Google Scholar] [CrossRef]
- Kumar, A.; Coleman, I.; Morrissey, C.; Zhang, X.; True, L.D.; Gulati, R.; Etzioni, R.; Bolouri, H.; Montgomery, B.; White, T.; et al. Substantial interindividual and limited intraindividual genomic diversity among tumors from men with metastatic prostate cancer. Nat. Med. 2016, 22, 369–378. [Google Scholar] [CrossRef]
- Giri, V.N.; Knudsen, K.E.; Kelly, W.K.; Abida, W.; Andriole, G.L.; Bangma, C.H.; Bekelman, J.E.; Benson, M.C.; Blanco, A.; Burnett, A.; et al. Role of Genetic Testing for Inherited Prostate Cancer Risk: Philadelphia Prostate Cancer Consensus Conference 2017. J. Clin. Oncol. 2018, 36, 414–424. [Google Scholar] [CrossRef]
- Zapala, P.; Dybowski, B.; Poletajew, S.; Radziszewski, P. What Can Be Expected from Prostate Cancer Biomarkers A Clinical Perspective. Urol. Int. 2018, 100, 1–12. [Google Scholar] [CrossRef]
- Strom, P.; Kartasalo, K.; Olsson, H.; Solorzano, L.; Delahunt, B.; Berney, D.M.; Bostwick, D.G.; Evans, A.J.; Grignon, D.J.; Humphrey, P.A.; et al. Artificial intelligence for diagnosis and grading of prostate cancer in biopsies: A population-based, diagnostic study. Lancet Oncol. 2020, 21, 222–232. [Google Scholar] [CrossRef]
- Bulten, W.; Kartasalo, K.; Chen, P.C.; Strom, P.; Pinckaers, H.; Nagpal, K.; Cai, Y.; Steiner, D.F.; van Boven, H.; Vink, R.; et al. Artificial intelligence for diagnosis and Gleason grading of prostate cancer: The PANDA challenge. Nat. Med. 2022, 28, 154–163. [Google Scholar] [CrossRef]
- Pantanowitz, L.; Quiroga-Garza, G.M.; Bien, L.; Heled, R.; Laifenfeld, D.; Linhart, C.; Sandbank, J.; Albrecht Shach, A.; Shalev, V.; Vecsler, M.; et al. An artificial intelligence algorithm for prostate cancer diagnosis in whole slide images of core needle biopsies: A blinded clinical validation and deployment study. Lancet Digit. Health 2020, 2, e407–e416. [Google Scholar] [CrossRef]
- Chen, P.C.; Gadepalli, K.; MacDonald, R.; Liu, Y.; Kadowaki, S.; Nagpal, K.; Kohlberger, T.; Dean, J.; Corrado, G.S.; Hipp, J.D.; et al. An augmented reality microscope with real-time artificial intelligence integration for cancer diagnosis. Nat. Med. 2019, 25, 1453–1457. [Google Scholar] [CrossRef]
- Chin, J.; Bauman, G.; Power, N.; Ward, A. The Singularity is Near(ish): Emerging Applications of Artificial Intelligence in Prostate Cancer Management. Eur. Urol. 2020, 77, 293–295. [Google Scholar] [CrossRef]
- Hopstaken, J.S.; Bomers, J.G.R.; Sedelaar, M.J.P.; Valerio, M.; Futterer, J.J.; Rovers, M.M. An Updated Systematic Review on Focal Therapy in Localized Prostate Cancer: What Has Changed over the Past 5 Years? Eur. Urol. 2022, 81, 5–33. [Google Scholar] [CrossRef]
- Deluce, J.E.; Cardenas, L.; Lalani, A.K.; Maleki Vareki, S.; Fernandes, R. Emerging Biomarker-Guided Therapies in Prostate Cancer. Curr. Oncol. 2022, 29, 5054–5076. [Google Scholar] [CrossRef] [PubMed]
- Pagliuca, M.; Buonerba, C.; Fizazi, K.; Di Lorenzo, G. The Evolving Systemic Treatment Landscape for Patients with Advanced Prostate Cancer. Drugs 2019, 79, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.Y.; Gleave, M.E.; Beltran, H. Towards precision oncology in advanced prostate cancer. Nat. Rev. Urol. 2019, 16, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Myrskyla, M.; Kohler, H.P.; Billari, F.C. Advances in development reverse fertility declines. Nature 2009, 460, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Caddy, D.; Short, R.V. Use of dizygotic to monozygotic twinning ratio as a measure of fertility. Lancet 1997, 349, 843–845. [Google Scholar] [CrossRef]
- Vollset, S.E.; Goren, E.; Yuan, C.W.; Cao, J.; Smith, A.E.; Hsiao, T.; Bisignano, C.; Azhar, G.S.; Castro, E.; Chalek, J.; et al. Fertility, mortality, migration, and population scenarios for 195 countries and territories from 2017 to 2100: A forecasting analysis for the Global Burden of Disease Study. Lancet 2020, 396, 1285–1306. [Google Scholar] [CrossRef]
- Ventimiglia, E.; Capogrosso, P.; Boeri, L.; Serino, A.; Colicchia, M.; Ippolito, S.; Scano, R.; Papaleo, E.; Damiano, R.; Montorsi, F.; et al. Infertility as a proxy of general male health: Results of a cross-sectional survey. Fertil. Steril. 2015, 104, 48–55. [Google Scholar] [CrossRef]
- Minhas, S.; Bettocchi, C.; Boeri, L.; Capogrosso, P.; Carvalho, J.; Cilesiz, N.C.; Cocci, A.; Corona, G.; Dimitropoulos, K.; Gul, M.; et al. European Association of Urology Guidelines on Male Sexual and Reproductive Health: 2021 Update on Male Infertility. Eur. Urol. 2021, 80, 603–620. [Google Scholar] [CrossRef]
- Hotaling, J.M.; Walsh, T.J. Male infertility: A risk factor for testicular cancer. Nat. Rev. Urol. 2009, 6, 550–556. [Google Scholar] [CrossRef]
- Choy, J.T.; Eisenberg, M.L. Male infertility as a window to health. Fertil. Steril. 2018, 110, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Del Giudice, F.; Kasman, A.M.; Chen, T.; De Berardinis, E.; Busetto, G.M.; Sciarra, A.; Ferro, M.; Lucarelli, G.; Belladelli, F.; Salonia, A.; et al. The Association between Mortality and Male Infertility: Systematic Review and Meta-analysis. Urology 2021, 154, 148–157. [Google Scholar] [CrossRef]
- Egozcue, S.; Blanco, J.; Vendrell, J.M.; García, F.; Veiga, A.; Aran, B.; Barri, P.N.; Vidal, F.; Egozcue, J. Human male infertility: Chromosome anomalies, meiotic disorders, abnormal spermatozoa and recurrent abortion. Hum. Reprod. Update 2000, 6, 93–105. [Google Scholar] [CrossRef] [Green Version]
- du Fosse, N.A.; van der Hoorn, M.P.; van Lith, J.M.M.; le Cessie, S.; Lashley, E. Advanced paternal age is associated with an increased risk of spontaneous miscarriage: A systematic review and meta-analysis. Hum. Reprod. Update 2020, 26, 650–669. [Google Scholar] [CrossRef]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Oud, M.S.; Smits, R.M.; Smith, H.E.; Mastrorosa, F.K.; Holt, G.S.; Houston, B.J.; de Vries, P.F.; Alobaidi, B.K.S.; Batty, L.E.; Ismail, H.; et al. A de novo paradigm for male infertility. Nat. Commun. 2022, 13, 154. [Google Scholar] [CrossRef]
- Tiepolo, L.; Zuffardi, O. Localization of factors controlling spermatogenesis in the nonfluorescent portion of the human Y chromosome long arm. Hum. Genet. 1976, 34, 119–124. [Google Scholar] [CrossRef]
- Ma, K.; Sharkey, A.; Kirsch, S.; Vogt, P.; Keil, R.; Hargreave, T.B.; McBeath, S.; Chandley, A.C. Towards the molecular localisation of the AZF locus: Mapping of microdeletions in azoospermic men within 14 subintervals of interval 6 of the human Y chromosome. Hum. Mol. Genet. 1992, 1, 29–33. [Google Scholar] [CrossRef]
- Vogt, P.H.; Edelmann, A.; Kirsch, S.; Henegariu, O.; Hirschmann, P.; Kiesewetter, F.; Kohn, F.M.; Schill, W.B.; Farah, S.; Ramos, C.; et al. Human Y chromosome azoospermia factors (AZF) mapped to different subregions in Yq11. Hum. Mol. Genet. 1996, 5, 933–943. [Google Scholar] [CrossRef]
- Pryor, J.L.; Kent-First, M.; Muallem, A.; Van Bergen, A.H.; Nolten, W.E.; Meisner, L.; Roberts, K.P. Microdeletions in the Y chromosome of infertile men. N. Engl. J. Med. 1997, 336, 534–539. [Google Scholar] [CrossRef]
- Foresta, C.; Moro, E.; Ferlin, A. Y chromosome microdeletions and alterations of spermatogenesis. Endocr. Rev. 2001, 22, 226–239. [Google Scholar]
- Riordan, J.R.; Rommens, J.M.; Kerem, B.; Alon, N.; Rozmahel, R.; Grzelczak, Z.; Zielenski, J.; Lok, S.; Plavsic, N.; Chou, J.L.; et al. Identification of the cystic fibrosis gene: Cloning and characterization of complementary DNA. Science 1989, 245, 1066–1073. [Google Scholar] [CrossRef] [PubMed]
- Bieth, E.; Hamdi, S.M.; Mieusset, R. Genetics of the congenital absence of the vas deferens. Hum. Genet. 2021, 140, 59–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chillon, M.; Casals, T.; Mercier, B.; Bassas, L.; Lissens, W.; Silber, S.; Romey, M.C.; Ruiz-Romero, J.; Verlingue, C.; Claustres, M.; et al. Mutations in the cystic fibrosis gene in patients with congenital absence of the vas deferens. N. Engl. J. Med. 1995, 332, 1475–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, S.; Mahale, S.D. Male Infertility Knowledgebase: Decoding the genetic and disease landscape. Database J. Biol. Databases Curation 2021, 2021, baab049. [Google Scholar]
- Houston, B.J.; Riera-Escamilla, A.; Wyrwoll, M.J.; Salas-Huetos, A.; Xavier, M.J.; Nagirnaja, L.; Friedrich, C.; Conrad, D.F.; Aston, K.I.; Krausz, C.; et al. A systematic review of the validated monogenic causes of human male infertility: 2020 update and a discussion of emerging gene-disease relationships. Hum. Reprod. Update 2021, 28, 15–29. [Google Scholar] [CrossRef]
- Oud, M.S.; Volozonoka, L.; Smits, R.M.; Vissers, L.; Ramos, L.; Veltman, J.A. A systematic review and standardized clinical validity assessment of male infertility genes. Hum. Reprod. 2019, 34, 932–941. [Google Scholar] [CrossRef]
- Spiliopoulou, A.; Nagy, R.; Bermingham, M.L.; Huffman, J.E.; Hayward, C.; Vitart, V.; Rudan, I.; Campbell, H.; Wright, A.F.; Wilson, J.F.; et al. Genomic prediction of complex human traits: Relatedness, trait architecture and predictive meta-models. Hum. Mol. Genet. 2015, 24, 4167–4182. [Google Scholar] [CrossRef] [Green Version]
- Belhechmi, S.; Bin, R.; Rotolo, F.; Michiels, S. Accounting for grouped predictor variables or pathways in high-dimensional penalized Cox regression models. BMC Bioinform. 2020, 21, 277. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Ren, J.; Wang, Y.; Wang, Z.; Zhang, J. Statistical Approaches for the Construction and Interpretation of Human Protein-Protein Interaction Network. BioMed Res. Int. 2016, 2016, 5313050. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, F.; Fu, J.; Zhang, P.; Wang, Y.; Zeng, X. Systematic identification and characterization of long non-coding RNAs in mouse mature sperm. PLoS ONE 2017, 12, e0173402. [Google Scholar] [CrossRef] [Green Version]
- Casas, E.; Vavouri, T. Sperm epigenomics: Challenges and opportunities. Front. Genet. 2014, 5, 330. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, J.; Rassoulzadegan, M.; Tuorto, F.; Chen, Q. Sperm RNA code programmes the metabolic health of offspring. Nat. Rev. Endocrinol. 2019, 15, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.; Silva, J.V.; Howl, J.; Santos, M.A.S.; Fardilha, M. All you need to know about sperm RNAs. Hum. Reprod. Update 2021, 28, 67–91. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Dix, D.J.; Krawetz, S.A. A bioinformatic strategy to rapidly characterize cDNA libraries. Bioinformatics 2002, 18, 949–952. [Google Scholar] [CrossRef] [Green Version]
- Ostermeier, G.C.; Dix, D.J.; Miller, D.; Khatri, P.; Krawetz, S.A. Spermatozoal RNA profiles of normal fertile men. Lancet 2002, 360, 772–777. [Google Scholar] [CrossRef]
- Luk, A.C.; Gao, H.; Xiao, S.; Liao, J.; Wang, D.; Tu, J.; Rennert, O.M.; Chan, W.Y.; Lee, T.L. GermlncRNA: A unique catalogue of long non-coding RNAs and associated regulations in male germ cell development. Database J. Biol. Databases Curation 2015, 2015, bav044. [Google Scholar] [CrossRef]
- Lee, T.L.; Cheung, H.H.; Claus, J.; Sastry, C.; Singh, S.; Vu, L.; Rennert, O.; Chan, W.Y. GermSAGE: A comprehensive SAGE database for transcript discovery on male germ cell development. Nucleic Acids Res. 2009, 37, D891–D897. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Grow, E.J.; Yi, C.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Murphy, P.J.; Wike, C.L.; Carrell, D.T.; Goriely, A.; et al. Chromatin and Single-Cell RNA-Seq Profiling Reveal Dynamic Signaling and Metabolic Transitions during Human Spermatogonial Stem Cell Development. Cell Stem Cell 2017, 21, 533–546.e6. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Murray, E.; Sinha, A.; Laumas, A.; Li, J.; Lesman, D.; Nie, X.; Hotaling, J.; Guo, J.; Cairns, B.R.; et al. Dissecting mammalian spermatogenesis using spatial transcriptomics. Cell Rep. 2021, 37, 109915. [Google Scholar] [CrossRef]
- Chen, H.; Xie, Y.; Li, Y.; Zhang, C.; Lv, L.; Yao, J.; Deng, C.; Sun, X.; Zou, X.; Liu, G. Outcome prediction of microdissection testicular sperm extraction based on extracellular vesicles piRNAs. J. Assist. Reprod. Genet. 2021, 38, 1429–1439. [Google Scholar] [CrossRef]
- Martinez-Heredia, J.; Estanyol, J.M.; Ballesca, J.L.; Oliva, R. Proteomic identification of human sperm proteins. Proteomics 2006, 6, 4356–4369. [Google Scholar] [CrossRef]
- Baker, M.A.; Naumovski, N.; Hetherington, L.; Weinberg, A.; Velkov, T.; Aitken, R.J. Head and flagella subcompartmental proteomic analysis of human spermatozoa. Proteomics 2013, 13, 61–74. [Google Scholar] [CrossRef]
- Nixon, B.; Mitchell, L.A.; Anderson, A.L.; McLaughlin, E.A.; O’Bryan, M.K.; Aitken, R.J. Proteomic and functional analysis of human sperm detergent resistant membranes. J. Cell. Physiol. 2011, 226, 2651–2665. [Google Scholar] [CrossRef]
- Thomas, S.; Kratzsch, D.; Schaab, M.; Scholz, M.; Grunewald, S.; Thiery, J.; Paasch, U.; Kratzsch, J. Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil. Steril. 2013, 99, 1256–1263.e3. [Google Scholar] [CrossRef]
- Towns, J.M.; Leslie, D.E.; Denham, I.; Wigan, R.; Azzato, F.; Williamson, D.A.; Lee, D.; Chow, E.P.F.; Fairley, C.K.; Graves, S.R.; et al. Treponema pallidum detection in lesion and non-lesion sites in men who have sex with men with early syphilis: A prospective, cross-sectional study. Lancet. Infect. Dis. 2021, 21, 1324–1331. [Google Scholar] [CrossRef]
- Imaz, A.; Niubo, J.; Cottrell, M.L.; Perez, E.; Kashuba, A.D.M.; Tiraboschi, J.M.; Morenilla, S.; Garcia, B.; Podzamczer, D. Seminal Tenofovir Concentrations, Viral Suppression, and Semen Quality With Tenofovir Alafenamide, Compared with Tenofovir Disoproxil Fumarate (Spanish HIV/AIDS Research Network, PreEC/RIS 40). Clin. Infect. Dis. 2019, 69, 1403–1409. [Google Scholar] [CrossRef]
- Fauque, P.; De Mouzon, J.; Devaux, A.; Epelboin, S.; Gervoise-Boyer, M.J.; Levy, R.; Valentin, M.; Viot, G.; Bergere, A.; De Vienne, C.; et al. Reproductive technologies, female infertility, and the risk of imprinting-related disorders. Clin. Epigenetics 2020, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Zembutsu, H.; Hirasawa, A.; Takahashi, A.; Kubo, M.; Akahane, T.; Aoki, D.; Kamatani, N.; Hirata, K.; Nakamura, Y. A genome-wide association study identifies genetic variants in the CDKN2BAS locus associated with endometriosis in Japanese. Nat. Genet. 2010, 42, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Nyholt, D.R.; Low, S.K.; Anderson, C.A.; Painter, J.N.; Uno, S.; Morris, A.P.; MacGregor, S.; Gordon, S.D.; Henders, A.K.; Martin, N.G.; et al. Genome-wide association meta-analysis identifies new endometriosis risk loci. Nat. Genet. 2012, 44, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Painter, J.N.; Anderson, C.A.; Nyholt, D.R.; Macgregor, S.; Lin, J.; Lee, S.H.; Lambert, A.; Zhao, Z.Z.; Roseman, F.; Guo, Q.; et al. Genome-wide association study identifies a locus at 7p15.2 associated with endometriosis. Nat. Genet. 2011, 43, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, Y.; Steinthorsdottir, V.; Morris, A.P.; Fassbender, A.; Rahmioglu, N.; De Vivo, I.; Buring, J.E.; Zhang, F.; Edwards, T.L.; Jones, S.; et al. Meta-analysis identifies five novel loci associated with endometriosis highlighting key genes involved in hormone metabolism. Nat. Commun. 2017, 8, 15539. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.J.; Zhao, H.; He, L.; Shi, Y.; Qin, Y.; Shi, Y.; Li, Z.; You, L.; Zhao, J.; Liu, J.; et al. Genome-wide association study identifies susceptibility loci for polycystic ovary syndrome on chromosome 2p16.3, 2p21 and 9q33.3. Nat. Genet. 2011, 43, 55–59. [Google Scholar] [CrossRef]
- Lerchbaum, E.; Trummer, O.; Giuliani, A.; Gruber, H.J.; Pieber, T.R.; Obermayer-Pietsch, B. Susceptibility loci for polycystic ovary syndrome on chromosome 2p16.3, 2p21, and 9q33.3 in a cohort of Caucasian women. Horm. Metab. Res. 2011, 43, 743–747. [Google Scholar] [CrossRef]
- Day, F.; Karaderi, T.; Jones, M.R.; Meun, C.; He, C.; Drong, A.; Kraft, P.; Lin, N.; Huang, H.; Broer, L.; et al. Large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for different diagnosis criteria. PLoS Genet. 2018, 14, e1007813. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhao, H.; Shi, Y.; Cao, Y.; Yang, D.; Li, Z.; Zhang, B.; Liang, X.; Li, T.; Chen, J.; et al. Genome-wide association study identifies eight new risk loci for polycystic ovary syndrome. Nat. Genet. 2012, 44, 1020–1025. [Google Scholar] [CrossRef]
- Day, F.R.; Hinds, D.A.; Tung, J.Y.; Stolk, L.; Styrkarsdottir, U.; Saxena, R.; Bjonnes, A.; Broer, L.; Dunger, D.B.; Halldorsson, B.V.; et al. Causal mechanisms and balancing selection inferred from genetic associations with polycystic ovary syndrome. Nat. Commun. 2015, 6, 8464. [Google Scholar] [CrossRef]
- McAllister, J.M.; Legro, R.S.; Modi, B.P.; Strauss, J.F. 3rd, Functional genomics of PCOS: From GWAS to molecular mechanisms. Trends Endocrinol. Metab. TEM 2015, 26, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Hayes, M.G.; Urbanek, M.; Ehrmann, D.A.; Armstrong, L.L.; Lee, J.Y.; Sisk, R.; Karaderi, T.; Barber, T.M.; McCarthy, M.I.; Franks, S.; et al. Genome-wide association of polycystic ovary syndrome implicates alterations in gonadotropin secretion in European ancestry populations. Nat. Commun. 2015, 6, 7502. [Google Scholar] [CrossRef] [Green Version]
- Beim, P.Y.; Hu-Seliger, T.; Elashoff, M.; Chodroff, R.; Lee, J.A.; Copperman, A.B. Whole genome sequencing for female infertility biomarker discovery. Fertil. Steril. 2013, 100, s324. [Google Scholar] [CrossRef]
- Kozhabek, Z.; Pang, M.; Zhao, Q.; Yi, J.; Huang, W. Copy number variation in female infertility and candidate gene screening for common infertility-related diseases. Bull. Karaganda Univ. Biol. Med. Geogr. Ser. 2021, 100, 73–79. [Google Scholar] [CrossRef]
- Schubert, M.; Perez Lanuza, L.; Woste, M.; Dugas, M.; Carmona, F.D.; Palomino-Morales, R.J.; Rassam, Y.; Heilmann-Heimbach, S.; Tuttelmann, F.; Kliesch, S.; et al. A GWAS in Idiopathic/Unexplained Infertile Men Detects a Genomic Region Determining Follicle-Stimulating Hormone Levels. J. Clin. Endocrinol. Metab. 2022, 107, 2350–2361. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ma, Y.; Zhang, J.; Jiang, S.; Yuan, G.; Cheng, J.; Lan, T.; Hao, J. Alteration in autophagy gene expression profile correlates with low sperm quality. Reprod. Biol. 2021, 21, 100546. [Google Scholar] [CrossRef] [PubMed]
- Maddirevula, S.; Coskun, S.; Alhassan, S.; Elnour, A.; Alsaif, H.S.; Ibrahim, N.; Abdulwahab, F.; Arold, S.T.; Alkuraya, F.S. Female Infertility Caused by Mutations in the Oocyte-Specific Translational Repressor PATL2. Am. J. Hum. Genet. 2017, 101, 603–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Ni, C.; Wu, L.; Chen, B.; Xu, Y.; Zhang, Z.; Mu, J.; Li, B.; Yan, Z.; Fu, J.; et al. Novel mutations in ZP1, ZP2, and ZP3 cause female infertility due to abnormal zona pellucida formation. Hum. Genet. 2019, 138, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, M.; Gao, J. Alterations in synaptonemal complex coding genes and human infertility. Int. J. Biol. Sci. 2022, 18, 1933–1943. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhao, J.; Liu, F.; Li, Y. Lipid metabolism and endometrial receptivity. Hum. Reprod. Update 2022, 28, 858–889. [Google Scholar] [CrossRef]
- Saei Ghare Naz, M.; Ramezani Tehrani, F.; Behroozi-Lak, T.; Mohammadzadeh, F.; Kholosi Badr, F.; Ozgoli, G. Polycystic Ovary Syndrome and Pelvic Floor Dysfunction: A Narrative Review. Res. Rep. Urol. 2020, 12, 179–185. [Google Scholar] [CrossRef]
- Yu, T.; Wu, D.; Cao, Y.; Zhai, J. Association Between Menstrual Patterns and Adverse Pregnancy Outcomes in Patients with Polycystic Ovary Syndrome. Front. Endocrinol. 2021, 12, 740377. [Google Scholar] [CrossRef]
- Sominsky, L.; Hodgson, D.M.; McLaughlin, E.A.; Smith, R.; Wall, H.M.; Spencer, S.J. Linking Stress and Infertility: A Novel Role for Ghrelin. Endocr. Rev. 2017, 38, 432–467. [Google Scholar] [CrossRef]
- Thong, E.P.; Codner, E.; Laven, J.S.E.; Teede, H. Diabetes: A metabolic and reproductive disorder in women. Lancet Diabetes Endocrinol. 2020, 8, 134–149. [Google Scholar] [CrossRef]
- Turan, V.; Lambertini, M.; Lee, D.Y.; Wang, E.; Clatot, F.; Karlan, B.Y.; Demeestere, I.; Bang, H.; Oktay, K. Association of Germline BRCA Pathogenic Variants with Diminished Ovarian Reserve: A Meta-Analysis of Individual Patient-Level Data. J. Clin. Oncol. 2021, 39, 2016–2024. [Google Scholar] [CrossRef]
- Liu, Q.; Zhu, Z.; Kraft, P.; Deng, Q.; Stener-Victorin, E.; Jiang, X. Genomic correlation, shared loci, and causal relationship between obesity and polycystic ovary syndrome: A large-scale genome-wide cross-trait analysis. BMC Med. 2022, 20, 66. [Google Scholar] [CrossRef]
- Lau, E.S.; Wang, D.; Roberts, M.; Taylor, C.N.; Murugappan, G.; Shadyab, A.H.; Schnatz, P.F.; Farland, L.V.; Wood, M.J.; Scott, N.S.; et al. Infertility and Risk of Heart Failure in the Women’s Health Initiative. J. Am. Coll. Cardiol. 2022, 79, 1594–1603. [Google Scholar] [CrossRef]
- Madjunkova, S.; Sundaravadanam, Y.; Antes, R.; Abramov, R.; Chen, S.; Yin, Y.; Zuzarte, P.C.; Moskovtsev, S.I.; Jorgensen, L.G.T.; Baratz, A.; et al. Detection of Structural Rearrangements in Embryos. N. Engl. J. Med. 2020, 382, 2472–2474. [Google Scholar] [CrossRef]
- Vermeesch, J.R.; Voet, T.; Devriendt, K. Prenatal and pre-implantation genetic diagnosis. Nat. Rev. Genet 2016, 17, 643–656. [Google Scholar] [CrossRef]
- Yan, L.; Huang, L.; Xu, L.; Huang, J.; Ma, F.; Zhu, X.; Tang, Y.; Liu, M.; Lian, Y.; Liu, P.; et al. Live births after simultaneous avoidance of monogenic diseases and chromosome abnormality by next-generation sequencing with linkage analyses. Proc. Natl. Acad. Sci. USA 2015, 112, 15964–15969. [Google Scholar] [CrossRef]
- Kumar, A.; Im, K.; Banjevic, M.; Ng, P.C.; Tunstall, T.; Garcia, G.; Galhardo, L.; Sun, J.; Schaedel, O.N.; Levy, B.; et al. Whole-genome risk prediction of common diseases in human preimplantation embryos. Nat. Med. 2022, 28, 513–516. [Google Scholar] [CrossRef]
- The alarming rise of complex genetic testing in human embryo selection. Nature 2022, 603, 549–550. [CrossRef]
- Capalbo, A.; Poli, M.; Riera-Escamilla, A.; Shukla, V.; Kudo Hoffding, M.; Krausz, C.; Hoffmann, E.R.; Simon, C. Preconception genome medicine: Current state and future perspectives to improve infertility diagnosis and reproductive and health outcomes based on individual genomic data. Hum. Reprod. Update 2021, 27, 254–279. [Google Scholar] [CrossRef]
- Rantsi, T.; Joki-Korpela, P.; Hokynar, K.; Kalliala, I.; Ohman, H.; Surcel, H.M.; Paavonen, J.; Tiitinen, A.; Puolakkainen, M. Serum antibody response to Chlamydia trachomatis TroA and HtrA in women with tubal factor infertility. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1499–1502. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; O’Connell, C.M.; Zhong, W.; Poston, T.B.; Wiesenfeld, H.C.; Hillier, S.L.; Trent, M.; Gaydos, C.; Tseng, G.; Taylor, B.D.; et al. Gene Expression Signatures Can Aid Diagnosis of Sexually Transmitted Infection-Induced Endometritis in Women. Front. Cell Infect. Microbiol. 2018, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, S.F.; De Medeiros, M.A.S.; Barbosa, B.B.; Yamamoto, A.; Yamamoto, M.M.W. The connection of alpha-1 acid glycoprotein inflammatory marker with anthropometric, hormonal, and metabolic characteristic of women with polycystic ovary syndrome. J. Obstet. Gynaecol. Res. 2021, 47, 3571–3582. [Google Scholar] [CrossRef] [PubMed]
- Mazloomi, S.; Sheikh, N.; Sanoee Farimani, M.; Pilehvari, S. Association of Prx4, Total Oxidant Status, and Inflammatory Factors with Insulin Resistance in Polycystic Ovary Syndrome. Int. J. Endocrinol. 2021, 2021, 9949753. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, L.; Chen, C.; Gao, Y. PCOS without hyperandrogenism is associated with higher plasma Trimethylamine N-oxide levels. BMC Endocr. Disord. 2020, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.L.; Chen, Z.; Feng, W.J.; Long, S.L.; Mo, Z.C. Sex hormone-binding globulin and polycystic ovary syndrome. Clin. Chim. Acta 2019, 499, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Meng, X.; Wang, J.; Wei, Y.; Tang, J.; Lei, X.; He, W.; Li, S. Serum amyloid A in polycystic ovary syndrome. Clin. Chim. Acta 2021, 518, 151–155. [Google Scholar] [CrossRef]
- Kalyan, S.; Goshtesabi, A.; Sarray, S.; Joannou, A.; Almawi, W.Y. Assessing C reactive protein/albumin ratio as a new biomarker for polycystic ovary syndrome: A case-control study of women from Bahraini medical clinics. BMJ Open 2018, 8, e021860. [Google Scholar] [CrossRef]
- Deng, H.; Li, Z.; Liu, G.; Li, X.; Chen, Y.; Zhang, Y.; Sun, Y.; Fu, J. Elevated serum interferon gamma-inducible protein-10 in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2017, 33, 363–367. [Google Scholar] [CrossRef]
- Gao, H.; Meng, J.; Xing, H.; Nie, S.; Xu, M.; Zhang, S.; Jin, Y.; Sun, T.; Huang, H.; Zhang, H.; et al. Association of heme oxygenase-1 with the risk of polycystic ovary syndrome in non-obese women. Hum. Reprod. 2014, 29, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Rashad, N.M.; El-Shal, A.S.; Abdelaziz, A.M. Association between inflammatory biomarker serum procalcitonin and obesity in women with polycystic ovary syndrome. J. Reprod. Immunol. 2013, 97, 232–239. [Google Scholar] [CrossRef]
- Pepene, C.E. Soluble platelet/endothelial cell adhesion molecule (sPECAM)-1 is increased in polycystic ovary syndrome and related to endothelial dysfunction. Gynecol. Endocrinol. 2012, 28, 370–374. [Google Scholar] [CrossRef]
- Sari, U.; Kaygusuz, I.; Kafali, H. Is pentraxin 3 a new cardiovascular risk marker in polycystic ovary syndrome? Gynecol. Obstet. Investig. 2014, 78, 173–178. [Google Scholar] [CrossRef]
- Daan, N.M.; Koster, M.P.; de Wilde, M.A.; Dalmeijer, G.W.; Evelein, A.M.; Fauser, B.C.; de Jager, W. Biomarker Profiles in Women with PCOS and PCOS Offspring; A Pilot Study. PLoS ONE 2016, 11, e0165033. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Nicholes, K.; Shih, I.M. The Origin and Pathogenesis of Endometriosis. Annu. Rev. Pathol. 2020, 15, 71–95. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, X.; Zhou, C.; Huang, X.; Lin, J.; Xu, H. Elevated immunoreactivity of RANTES and CCR1 correlate with the severity of stages and dysmenorrhea in women with deep infiltrating endometriosis. Acta Histochem. 2013, 115, 434–439. [Google Scholar] [CrossRef]
- Chen, L.; Ding, B.; Wu, L.; Qiu, J.; Li, Q.; Ye, Z.; Yang, J. Transcriptome Analysis Reveals the Mechanism of Natural Ovarian Ageing. Front. Endocrinol. 2022, 13, 918212. [Google Scholar] [CrossRef]
- Yasen, A.; Aini, A.; Wang, H.; Li, W.; Zhang, C.; Ran, B.; Tuxun, T.; Maimaitinijiati, Y.; Shao, Y.; Aji, T.; et al. Progress and applications of single-cell sequencing techniques. Infect. Genet. Evol. 2020, 80, 104198. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A.; et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Fan, W.; Yan, L.; Li, R.; Lian, Y.; Huang, J.; Li, J.; Xu, L.; Tang, F.; Xie, X.S.; et al. Genome analyses of single human oocytes. Cell 2013, 155, 1492–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Yang, M.; Guo, H.; Yang, L.; Wu, J.; Li, R.; Liu, P.; Lian, Y.; Zheng, X.; Yan, J.; et al. Single-cell RNA-Seq profiling of human preimplantation embryos and embryonic stem cells. Nat. Struct. Mol. Biol. 2013, 20, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Guo, H.; Ren, Y.; Hou, Y.; Dong, J.; Li, R.; Lian, Y.; Fan, X.; Hu, B.; Gao, Y.; et al. Single-cell DNA methylome sequencing of human preimplantation embryos. Nat. Genet. 2018, 50, 12–19. [Google Scholar] [CrossRef]
- Li, L.; Li, L.; Li, Q.; Liu, X.; Ma, X.; Yong, J.; Gao, S.; Wu, X.; Wei, Y.; Wang, X.; et al. Dissecting the epigenomic dynamics of human fetal germ cell development at single-cell resolution. Cell Res 2021, 31, 463–477. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Li, J.; Yu, Y.; Zhang, W.; Song, M.; Liu, Z.; Min, Z.; Hu, H.; Jing, Y.; et al. Single-Cell Transcriptomic Atlas of Primate Ovarian Aging. Cell 2020, 180, 585–600.e19. [Google Scholar] [CrossRef]
- Stevant, I.; Nef, S. Single cell transcriptome sequencing: A new approach for the study of mammalian sex determination. Mol. Cell. Endocrinol. 2018, 468, 11–18. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, L.; Zhan, H.; Mo, Y.; Ren, Z.; Shao, A.; Lin, J. Single-cell transcriptomic analysis of endometriosis provides insights into fibroblast fates and immune cell heterogeneity. Cell Biosci. 2021, 11, 125. [Google Scholar] [CrossRef]
- Lv, H.; Zhao, G.; Jiang, P.; Wang, H.; Wang, Z.; Yao, S.; Zhou, Z.; Wang, L.; Liu, D.; Deng, W.; et al. Deciphering the endometrial niche of human thin endometrium at single-cell resolution. Proc. Natl. Acad. Sci. USA 2022, 119, e2115912119. [Google Scholar] [CrossRef]
- Chung, R.H.; Kang, C.Y. A multi-omics data simulator for complex disease studies and its application to evaluate multi-omics data analysis methods for disease classification. GigaScience 2019, 8, giz045. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [Green Version]
- Pouryahya, M.; Oh, J.H.; Javanmard, P.; Mathews, J.C.; Belkhatir, Z.; Deasy, J.O.; Tannenbaum, A.R. aWCluster: A Novel Integrative Network-Based Clustering of Multiomics for Subtype Analysis of Cancer Data. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 1472–1483. [Google Scholar] [CrossRef]
- Singh, A.; Shannon, C.P.; Gautier, B.; Rohart, F.; Vacher, M.; Tebbutt, S.J.; Le Cao, K.A. DIABLO: An integrative approach for identifying key molecular drivers from multi-omics assays. Bioinformatics 2019, 35, 3055–3062. [Google Scholar] [CrossRef]
- Argelaguet, R.; Velten, B.; Arnol, D.; Dietrich, S.; Zenz, T.; Marioni, J.C.; Buettner, F.; Huber, W.; Stegle, O. Multi-Omics Factor Analysis—A framework for unsupervised integration of multi-omics data sets. Mol. Syst. Biol. 2018, 14, e8124. [Google Scholar] [CrossRef] [PubMed]
- Rappoport, N.; Shamir, R. Multi-omic and multi-view clustering algorithms: Review and cancer benchmark. Nucleic Acids Res. 2018, 46, 10546–10562. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Probst, P.; Hornung, R.; Jurinovic, V.; Boulesteix, A.L. Large-scale benchmark study of survival prediction methods using multi-omics data. Brief. Bioinform. 2021, 22, bbaa167. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.Y.; Li, S.; Narasimhan, B.; Tibshirani, R. Cooperative learning for multiview analysis. Proc. Natl. Acad. Sci. USA 2022, 119, e2202113119. [Google Scholar] [CrossRef] [PubMed]
- Jia, P.; Pei, G.; Zhao, Z. CNet: A multi-omics approach to detecting clinically associated, combinatory genomic signatures. Bioinformatics 2019, 35, 5207–5215. [Google Scholar] [CrossRef]
- Fatima, N.; Rueda, L. iSOM-GSN: An integrative approach for transforming multi-omic data into gene similarity networks via self-organizing maps. Bioinformatics 2020, 36, 4248–4254. [Google Scholar] [CrossRef]
- Zhao, R.; Jiang, Y.; Zhao, S.; Zhao, H. Multiomics Analysis Reveals Molecular Abnormalities in Granulosa Cells of Women with Polycystic Ovary Syndrome. Front. Genet. 2021, 12, 648701. [Google Scholar] [CrossRef]
- Xia, Y. Correlation and association analyses in microbiome study integrating multiomics in health and disease. Prog. Mol. Biol. Transl. Sci. 2020, 171, 309–491. [Google Scholar]
- Prasnikar, E.; Kunej, T.; Knez, J.; Repnik, K.; Potocnik, U.; Kovacic, B. Determining the Molecular Background of Endometrial Receptivity in Adenomyosis. Biomolecules 2020, 10, 1311. [Google Scholar] [CrossRef]
- Tsai, V.F.; Zhuang, B.; Pong, Y.H.; Hsieh, J.T.; Chang, H.C. Web- and Artificial Intelligence-Based Image Recognition For Sperm Motility Analysis: Verification Study. JMIR Med. Inform. 2020, 8, e20031. [Google Scholar] [CrossRef]
- Liu, L.; Jiao, Y.; Li, X.; Ouyang, Y.; Shi, D. Machine learning algorithms to predict early pregnancy loss after in vitro fertilization-embryo transfer with fetal heart rate as a strong predictor. Comput. Methods Programs Biomed. 2020, 196, 105624. [Google Scholar] [CrossRef]
- Chavez-Badiola, A.; Flores-Saiffe Farias, A.; Mendizabal-Ruiz, G.; Garcia-Sanchez, R.; Drakeley, A.J.; Garcia-Sandoval, J.P. Predicting pregnancy test results after embryo transfer by image feature extraction and analysis using machine learning. Sci. Rep. 2020, 10, 4394. [Google Scholar] [CrossRef] [Green Version]
- Rotshenker-Olshinka, K.; Srebnik Moshe, N.; Weiss, O.; Shaviv, S.; Freireich, O.; Segel, R.; Zeligson, S.; Eldar-Geva, T.; Altarescu, G. Preimplantation genetic testing (PGT) for copy number variants of uncertain significance (CNV- VUS) in the genomic era: To do or not to do? J. Assist. Reprod. Genet. 2021, 38, 719–725. [Google Scholar] [CrossRef]
- Cornelisse, S.; Zagers, M.; Kostova, E.; Fleischer, K.; van Wely, M.; Mastenbroek, S. Preimplantation genetic testing for aneuploidies (abnormal number of chromosomes) in in vitro fertilisation. Cochrane Database Syst. Rev. 2020, 9, CD005291. [Google Scholar]
- Brezina, P.R.; Anchan, R.; Kearns, W.G. Preimplantation genetic testing for aneuploidy: What technology should you use and what are the differences? J. Assist. Reprod. Genet. 2016, 33, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.C.; Jiang, V.S.; Kanakasabapathy, M.K.; Thirumalaraju, P.; Kandula, H.; Dimitriadis, I.; Souter, I.; Bormann, C.L.; Shafiee, H. Using artificial intelligence to avoid human error in identifying embryos: A retrospective cohort study. J. Assist. Reprod. Genet. 2022, 39, 2343–2348. [Google Scholar] [CrossRef]
- Fordham, D.E.; Rosentraub, D.; Polsky, A.L.; Aviram, T.; Wolf, Y.; Perl, O.; Devir, A.; Rosentraub, S.; Silver, D.H.; Gold Zamir, Y.; et al. Embryologist agreement when assessing blastocyst implantation probability: Is data-driven prediction the solution to embryo assessment subjectivity? Hum. Reprod. 2022, 37, 2275–2290. [Google Scholar] [CrossRef]
- Cheredath, A.; Uppangala, S.; CS, A.; Jijo, A.; Kumar, P.; Joseph, D.; GA, N.G.; Kalthur, G.; Adiga, S.K. Combining Machine Learning with Metabolomic and Embryologic Data Improves Embryo Implantation Prediction. Reprod. Sci. 2022. [Google Scholar] [CrossRef]
- Liu, R.; Bai, S.; Jiang, X.; Luo, L.; Tong, X.; Zheng, S.; Wang, Y.; Xu, B. Multifactor Prediction of Embryo Transfer Outcomes Based on a Machine Learning Algorithm. Front. Endocrinol. 2021, 12, 745039. [Google Scholar] [CrossRef]
- Yigit, P.; Bener, A.; Karabulut, S. Comparison of machine learning classification techniques to predict implantation success in an IVF treatment cycle. Reprod. Biomed. Online 2022, 45, 923–934. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, L.; Li, S.; Liu, L.; Yang, T.; Yang, P.; Zhao, J.; Arkin, I.T.; Liu, H. Predicting the reproductive toxicity of chemicals using ensemble learning methods and molecular fingerprints. Toxicol. Lett. 2021, 340, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Diakiw, S.M.; Hall, J.M.M.; VerMilyea, M.D.; Amin, J.; Aizpurua, J.; Giardini, L.; Briones, Y.G.; Lim, A.Y.X.; Dakka, M.A.; Nguyen, T.V.; et al. Development of an artificial intelligence model for predicting the likelihood of human embryo euploidy based on blastocyst images from multiple imaging systems during IVF. Hum. Reprod. 2022, 37, 1746–1759. [Google Scholar] [CrossRef]
- Huang, C.; Xiang, Z.; Zhang, Y.; Tan, D.S.; Yip, C.K.; Liu, Z.; Li, Y.; Yu, S.; Diao, L.; Wong, L.Y.; et al. Using Deep Learning in a Monocentric Study to Characterize Maternal Immune Environment for Predicting Pregnancy Outcomes in the Recurrent Reproductive Failure Patients. Front. Immunol. 2021, 12, 642167. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Duan, H.; Wang, S.; Wu, J.; Li, Y. Establishment of an Artificial Neural Network Model Using Immune-Infiltration Related Factors for Endometrial Receptivity Assessment. Vaccines 2022, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Kangasniemi, M.H.; Komsi, E.K.; Rossi, H.R.; Liakka, A.; Khatun, M.; Chen, J.C.; Paulson, M.; Hirschberg, A.L.; Arffman, R.K.; Piltonen, T.T. Artificial intelligence deep learning model assessment of leukocyte counts and proliferation in endometrium from women with and without polycystic ovary syndrome. F S Sci. 2022, 3, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.G.; Mello, M.M. HIPAA and Protecting Health Information in the 21st Century. Jama 2018, 320, 231–232. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Sun, H.; Cao, X.; Wen, N.; Wang, J.; Wang, C. Learning representations for gene ontology terms by jointly encoding graph structure and textual node descriptors. Brief. Bioinform. 2022, 23, bbac318. [Google Scholar] [CrossRef]
- Rosenwaks, Z. Artificial intelligence in reproductive medicine: A fleeting concept or the wave of the future? Fertil. Steril. 2020, 114, 905–907. [Google Scholar] [CrossRef]
- Telenti, A.; Jiang, X. Treating medical data as a durable asset. Nat. Genet. 2020, 52, 1005–1010. [Google Scholar] [CrossRef]
- Carter, H.; Hofree, M.; Ideker, T. Genotype to phenotype via network analysis. Curr. Opin. Genet. Dev. 2013, 23, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Houtchens, M.K.; Edwards, N.C.; Hayward, B.; Mahony, M.C.; Phillips, A.L. Live birth rates, infertility diagnosis, and infertility treatment in women with and without multiple sclerosis: Data from an administrative claims database. Mult. Scler. Relat. Disord. 2020, 46, 102541. [Google Scholar] [CrossRef]
- Watford, S.; Ly Pham, L.; Wignall, J.; Shin, R.; Martin, M.T.; Friedman, K.P. ToxRefDB version 2.0: Improved utility for predictive and retrospective toxicology analyses. Reprod. Toxicol. 2019, 89, 145–158. [Google Scholar] [CrossRef]
- Luke, B.; Stern, J.E.; Kotelchuck, M.; Declercq, E.R.; Cohen, B.; Diop, H. Birth Outcomes by Infertility Diagnosis Analyses of the Massachusetts Outcomes Study of Assisted Reproductive Technologies (MOSART). J. Reprod. Med. 2015, 60, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Ignatieva, E.V.; Osadchuk, A.V.; Kleshchev, M.A.; Bogomolov, A.G.; Osadchuk, L.V. A Catalog of Human Genes Associated with Pathozoospermia and Functional Characteristics of These Genes. Front. Genet. 2021, 12, 662770. [Google Scholar] [CrossRef]
- Jiang, X.; Zhao, D.; Ali, A.; Xu, B.; Liu, W.; Wen, J.; Zhang, H.; Shi, Q.; Zhang, Y. MeiosisOnline: A Manually Curated Database for Tracking and Predicting Genes Associated with Meiosis. Front. Cell Dev. Biol. 2021, 9, 673073. [Google Scholar] [CrossRef]
- Krenz, H.; Gromoll, J.; Darde, T.; Chalmel, F.; Dugas, M.; Tuttelmann, F. The Male Fertility Gene Atlas: A web tool for collecting and integrating OMICS data in the context of male infertility. Hum. Reprod. 2020, 35, 1983–1990. [Google Scholar] [CrossRef]
- Schuster, A.; Tang, C.; Xie, Y.; Ortogero, N.; Yuan, S.; Yan, W. SpermBase: A Database for Sperm-Borne RNA Contents. Biol. Reprod. 2016, 95, 99. [Google Scholar] [CrossRef] [Green Version]
- Dawe, A.S.; Radovanovic, A.; Kaur, M.; Sagar, S.; Seshadri, S.V.; Schaefer, U.; Kamau, A.A.; Christoffels, A.; Bajic, V.B. DESTAF: A database of text-mined associations for reproductive toxins potentially affecting human fertility. Reprod. Toxicol. 2012, 33, 99–105. [Google Scholar] [CrossRef]
- Matthews, E.J.; Kruhlak, N.L.; Daniel Benz, R.; Contrera, J.F. A comprehensive model for reproductive and developmental toxicity hazard identification: I. Development of a weight of evidence QSAR database. Regul. Toxicol. Pharmacol. RTP 2007, 47, 115–135. [Google Scholar] [CrossRef]
- Joseph, S.; Mahale, S.D. Endometriosis Knowledgebase: A gene-based resource on endometriosis. Database J. Biol. Databases Curation 2019, 2019, baz062. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database Name | Website | Development Agencies | Characteristic |
|---|---|---|---|
| Society for Assisted Reproductive Technology (SART) | https://www.sart.org | US ART Society | One of the largest reproductive medicine societies in the world, with over 90% of fertility centres in the US as members. Annual assisted reproduction statistics and industry standard setting in the US. |
| International Committee Monitoring Assissted Reproductive Technologies (ICMART) | https://www.icmartivf.org | International Conference Services | It takes a leading role in the development, collection and dissemination of worldwide data on ART through its World Report series. |
| Centers for Disease Control and Prevention (CDC) | https://www.cdc.org | US CDC | Annual assisted reproduction statistics and industry standard setting in the US. |
| European Society of Human Reproduction and Embryology (ESHRE) | https://www.eshre.eu/en | ESHRE | Annual assisted reproduction statistics and industry standard setting in Europe. |
| Human Fertilisation and Embryology Authority (HFEA) | https://www.bionews.org.uk | UK Department of Health | It is responsible for the regulation and inspection of all UK clinics offering in vitro fertilization, artificial insemination and human egg, sperm or embryo storage. It is also responsible for human embryo research. |
| Chinese Society of Reproductive Medicine (CSRM) | http://csrm1.meetingchina.org/msite/main/cn | Chinese Medical Association | Annual assisted reproduction statistics and industry standard setting in China. |
| Massachusetts Outcomes Study of Assisted Reproductive Technology (MOSART) | - | MGH Center for Child and Adolescent Health Research and Policy, MassGeneral Hospital for Children, US | It linked the SART Clinical Outcomes Reporting and the Massachusetts Pregnancy to Early Life Longitudinal (PELL) data systems, to provide a strong basis for further longitudinal ART outcomes studies. It also supports the continued development of potentially powerful linked clinical-public health databases [183]. |
| The Catalog of Genes Associated with Different Forms of Lowered Semen Quality Caused by Impaired Spermatogenesis (HGAPat) | https://www.sysbio.ru/hgap/ | Novosibirsk State University | A catalog of human genes associated with lowered semen quality (HGAPat) and analyzed their functional characteristics [184]. |
| MeiosisOnline | https://mcg.ustc.edu.cn/bsc/meiosis/index.html | University of Science and Technology of China | A manually curated database for tracking and predicting genes associated with meiosis [185] |
| Male Fertility Gene Atlas (MFGA) | https://mfga.uni-muenster.de | Germany Centre of Reproductive Medicine and Andrology, University Hospital Münster | It enables a more targeted search and interpretation of OMICS data on male infertility and germ cells in the context of relevant publications [186]. |
| SpermBase | http://www.spermbase.org | Department of Physiology and Cell Biology, University of Nevada School of Medicine, Reno, Nevada | A database for sperm-borne RNA contents [187] |
| GermlncRNA | http://germlncrna.cbiit.cuhk.edu.hk/ | The Chinese University of Hong Kong | A unique catalogue of long non-coding RNAs and associated regulations in male germ cell development [76]. |
| Dragon Exploration System for Toxicants and Fertility (DESTAF) | http://cbrc.kaust.edu.sa/destaf | King Abdullah University of Science and Technology (KAUST), Thuwal, Saudi Arabia | A database of text-mined associations for reproductive toxins potentially affecting human [188]. |
| GermSAGE | http://germsage.nichd.nih.gov | Eunice Kennedy Shriver National Institute of Child Health and Human Development | A comprehensive SAGE database for transcript discovery on male germ cell development [77]. |
| Reproductive and developmental toxicology (REPROTOX) | http://www.fda.gov/cder/Offices/OPS_IO/default.htm | US FDA | The database is suitable for QSAR modeling and human hazard identification of untested chemicals [189]. |
| Male Infertility Knowledgebase (MIK) | http://mik.bicnirrh.res.in/ | ICMR-National Institute for Research in Reproductive Health, India | A platform for review of genetic information on male infertility, identification pleiotropic genes, prediction of novel candidate genes for the different male infertility diseases and for portending future high-risk diseases associated with male infertility [64]. |
| Endometriosis Knowledgebase | http://www.ek.bicnirrh.res.in | ICMR-National Institute for Research in Reproductive Health, India | The database includes genes, pathways, gene ontologies and and protein functions common to endometriosis [190]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Zhang, Y.; Martin, C.; Ma, X.; Shen, B. Translational Bioinformatics for Human Reproductive Biology Research: Examples, Opportunities and Challenges for a Future Reproductive Medicine. Int. J. Mol. Sci. 2023, 24, 4. https://doi.org/10.3390/ijms24010004
Liu K, Zhang Y, Martin C, Ma X, Shen B. Translational Bioinformatics for Human Reproductive Biology Research: Examples, Opportunities and Challenges for a Future Reproductive Medicine. International Journal of Molecular Sciences. 2023; 24(1):4. https://doi.org/10.3390/ijms24010004
Chicago/Turabian StyleLiu, Kun, Yingbo Zhang, César Martin, Xiaoling Ma, and Bairong Shen. 2023. "Translational Bioinformatics for Human Reproductive Biology Research: Examples, Opportunities and Challenges for a Future Reproductive Medicine" International Journal of Molecular Sciences 24, no. 1: 4. https://doi.org/10.3390/ijms24010004
APA StyleLiu, K., Zhang, Y., Martin, C., Ma, X., & Shen, B. (2023). Translational Bioinformatics for Human Reproductive Biology Research: Examples, Opportunities and Challenges for a Future Reproductive Medicine. International Journal of Molecular Sciences, 24(1), 4. https://doi.org/10.3390/ijms24010004