
TSLP as a Potential Therapy in the Treatment of CRLF2 B Cell Acute Lymphoblastic Leukemia

 , ,
, ,
Abstract
:1. Introduction
2. Results
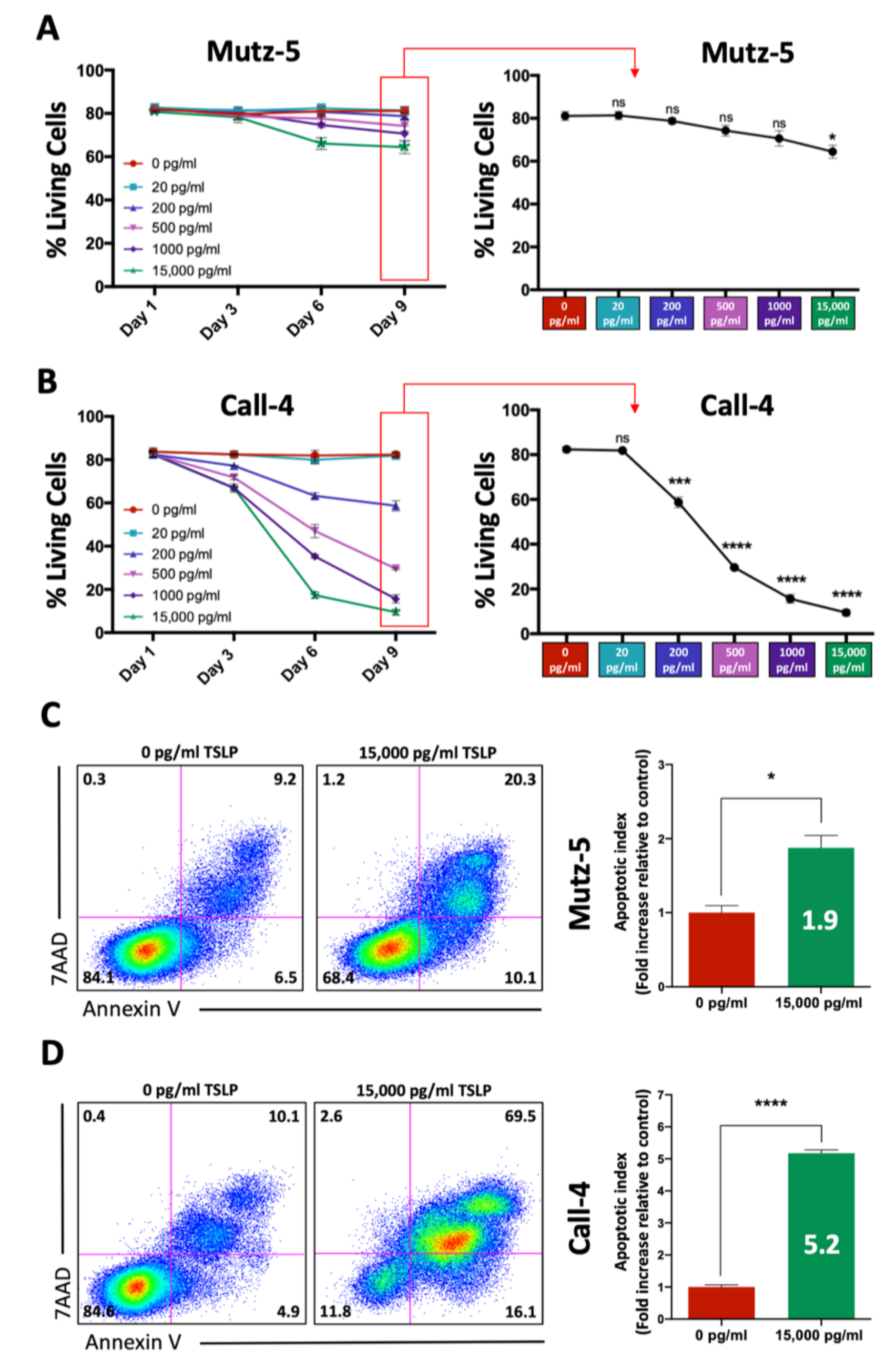
2.1. High TSLP Concentrations Induce Cell Death in CRLF2 B-ALL Cells In Vitro
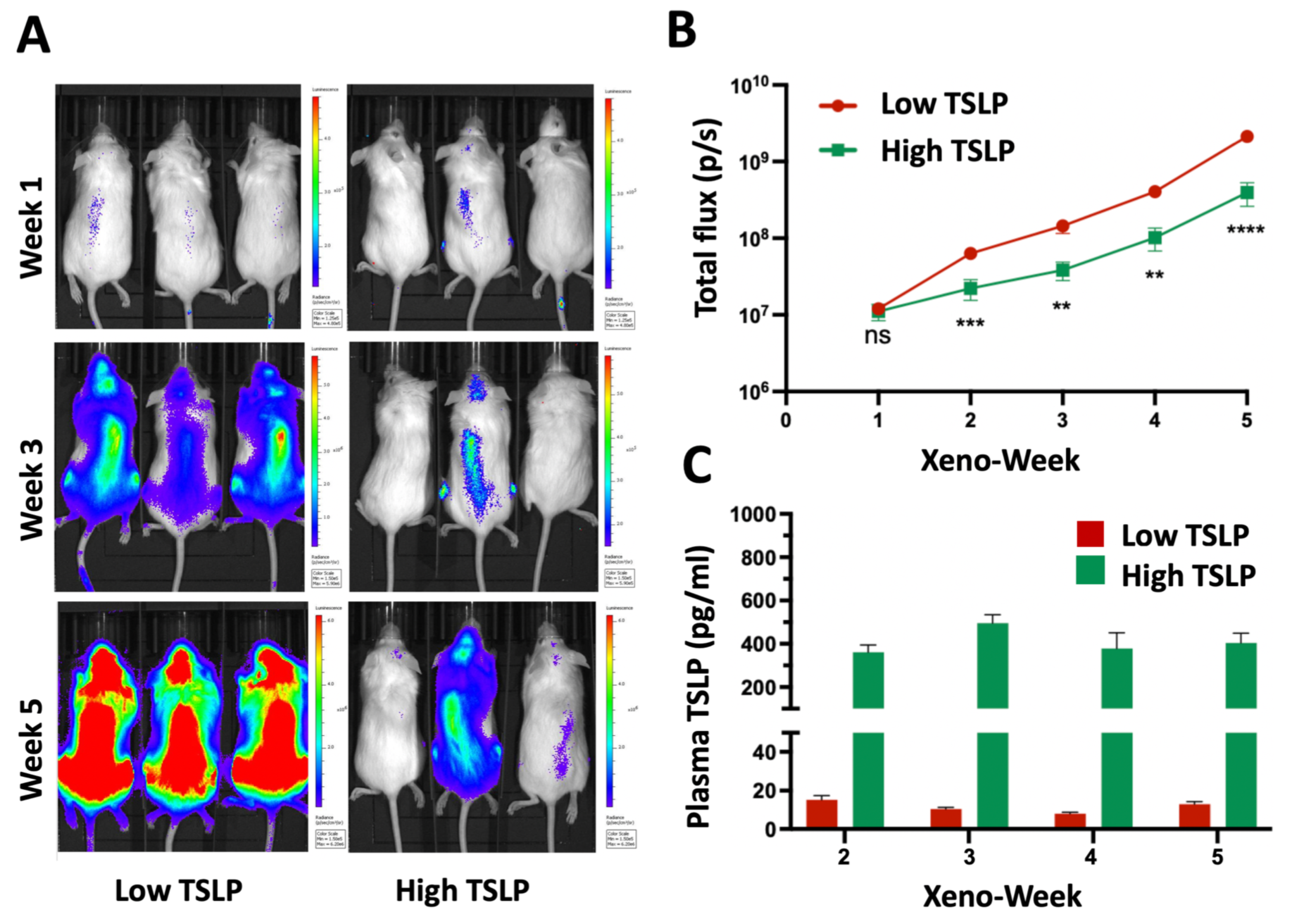
2.2. High TSLP Doses Reduce Leukemia Burden in Mutz-5-luc Xenografts
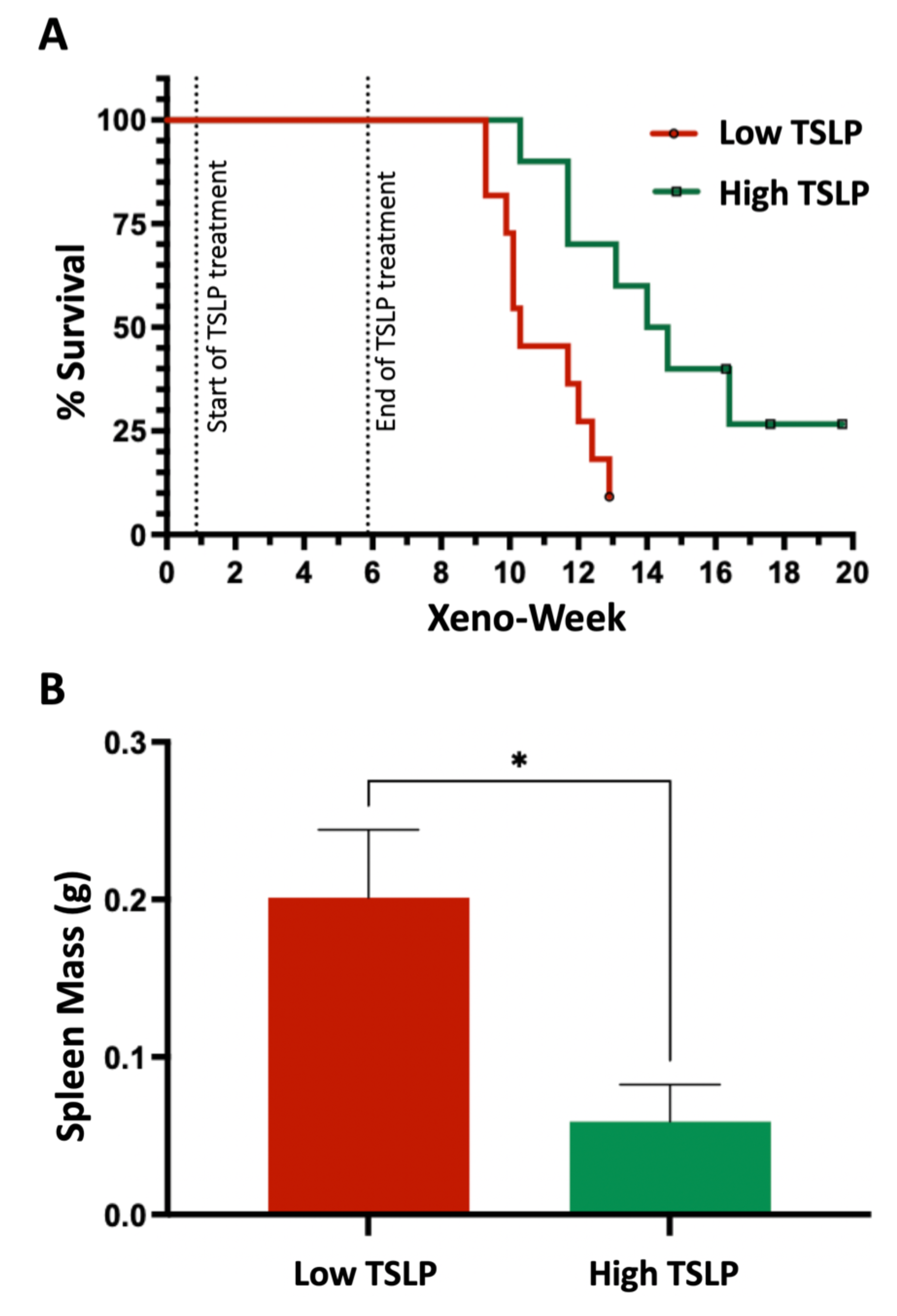
2.3. High TSLP Doses Prolong the Survival of CRLF2-B-ALL Patient-Derived Xenografts
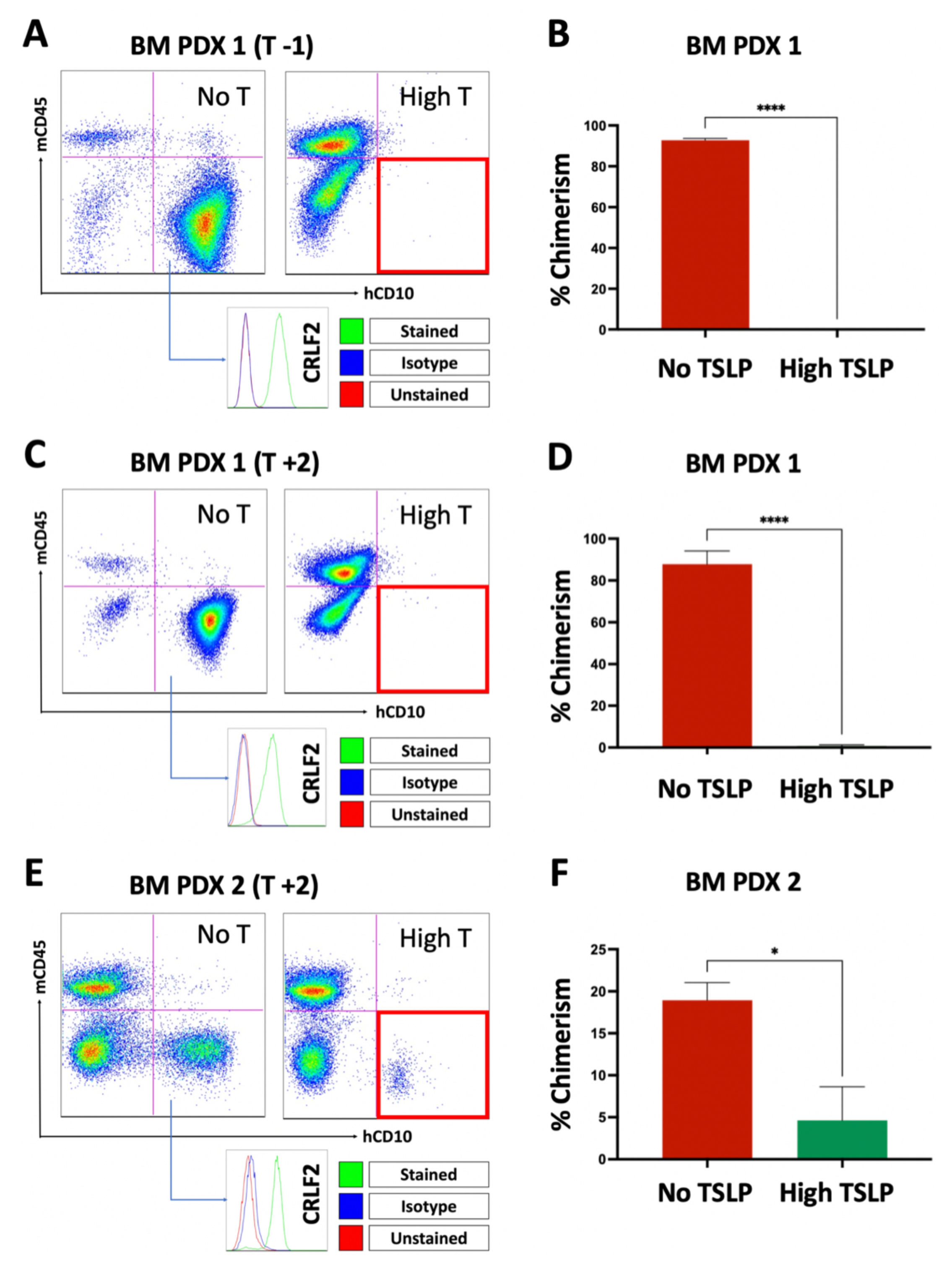
2.4. High TSLP Doses Prevent Engraftment of CRLF2 B-ALL Cells in PDXs
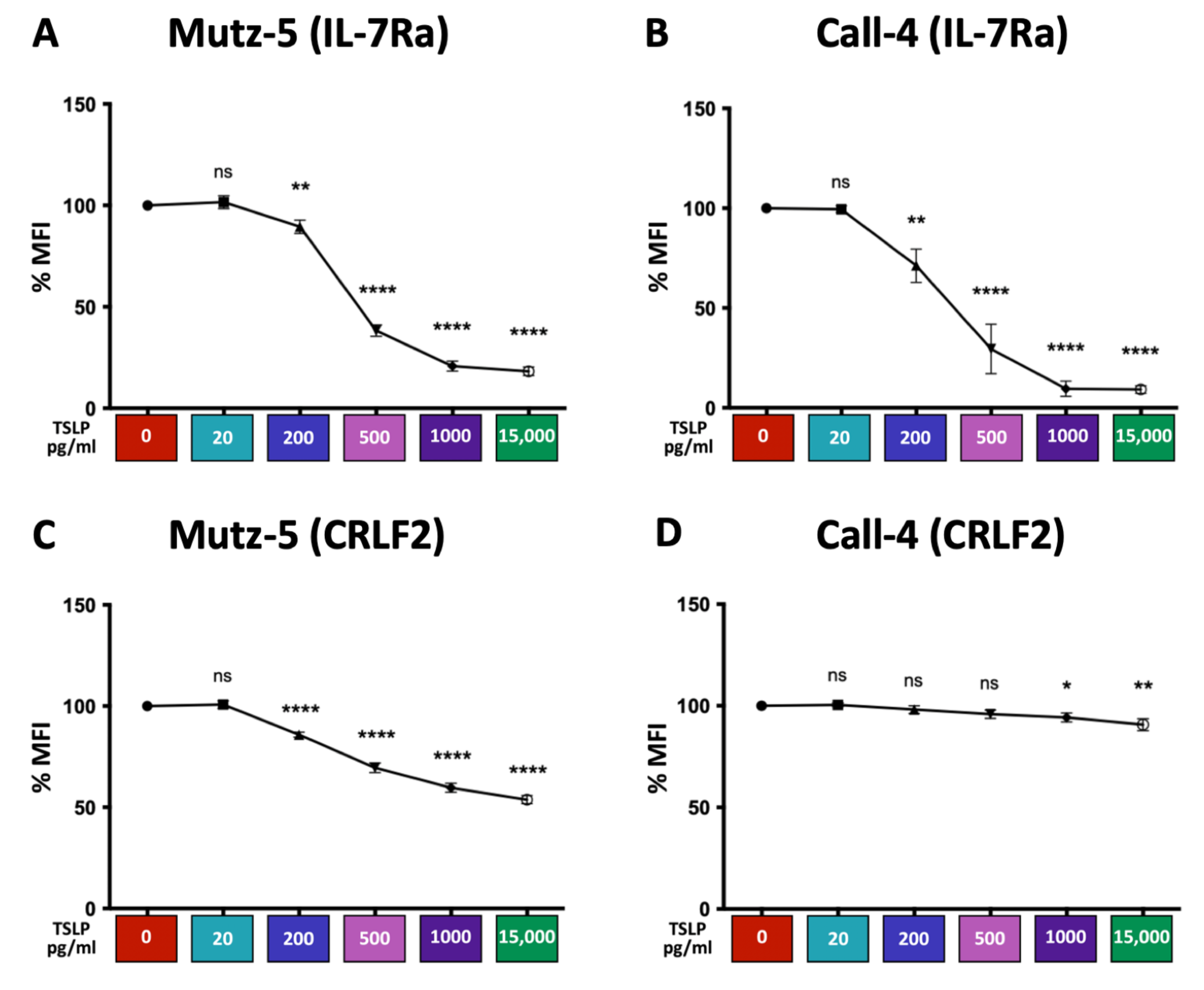
2.5. High TSLP Concentrations Reduce the Expression of IL-7Rα and CRLF2 in CRLF2 B-ALL Cells in a Dose-Dependent Manner In Vitro
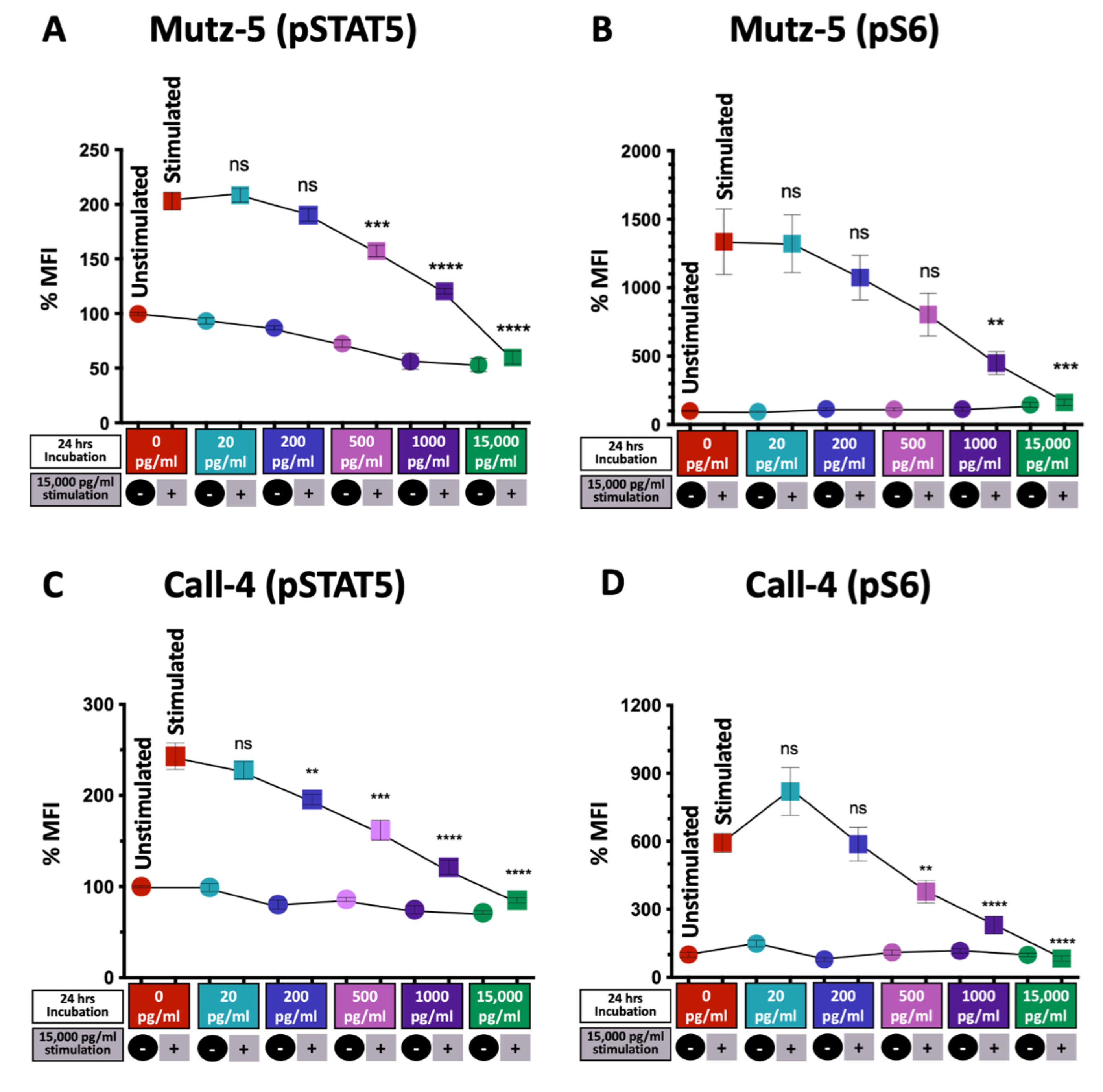
2.6. High TSLP Concentrations Reduce CRLF2 Signaling via the JAK/STAT and PI3/AKT/mTOR Pathways in CRLF2 B-ALL Cells
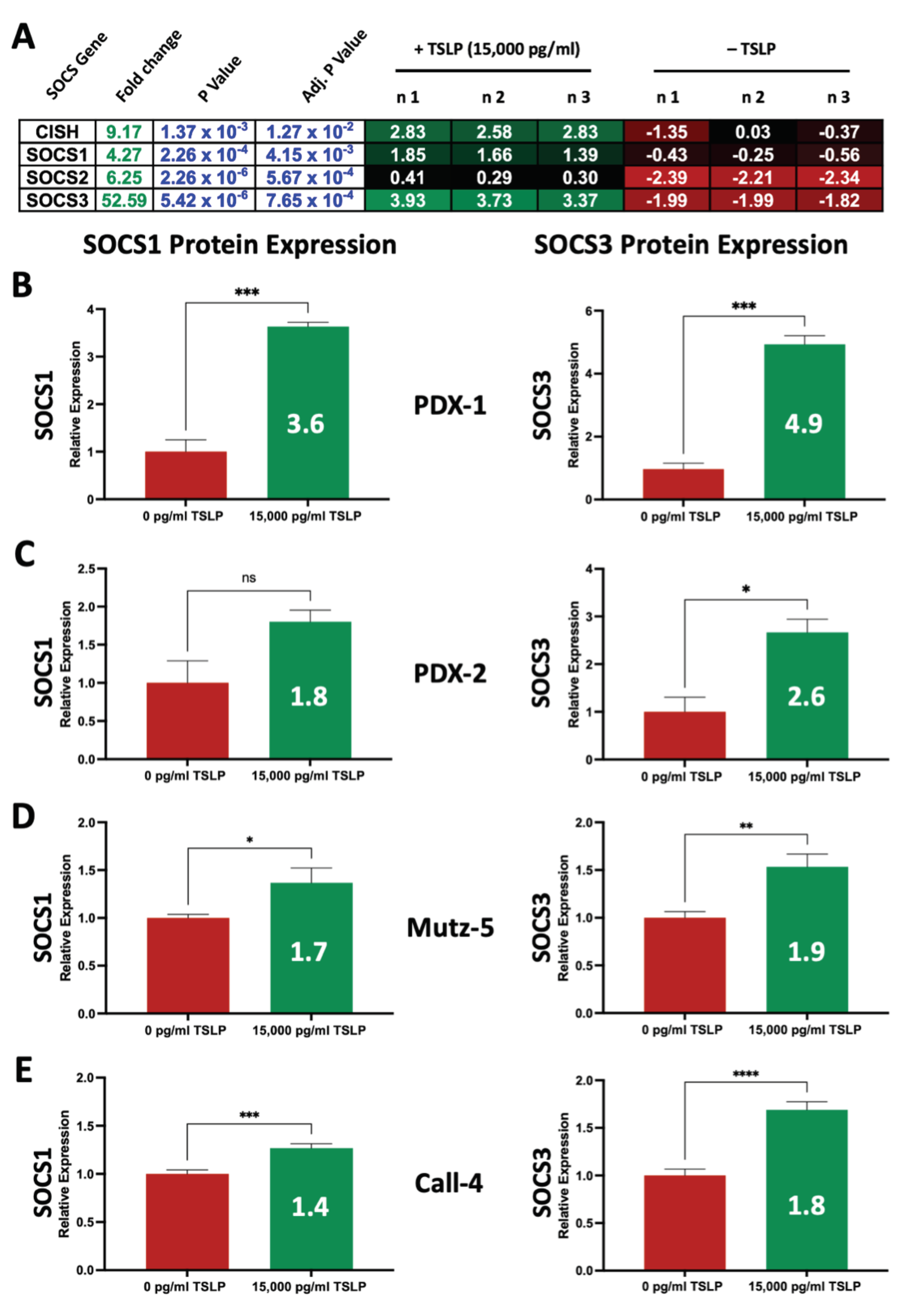
2.7. High Concentrations of TSLP Upregulate SOCS Genes and Proteins in CRLF2 B-ALL Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Primary Cells, and TSLP Treatment
4.2. Flow Cytometry
4.3. Phospho-Flow Cytometry Staining
4.4. Cell Death Studies
4.5. Western Blot
4.6. Xenograft Mouse Studies
4.7. IVIS Studies
4.8. Legend Plex Assay for TSLP
4.9. Whole-Genome Microarray Analysis of Gene Expression in CRLF2 B-ALL Cells
4.10. Flow Cytometry Antibodies
4.11. Statistical Analyses
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sasaki, K.; Jabbour, E.; Short, N.J.; Jain, N.; Ravandi, F.; Pui, C.-H.; Kantarjian, H. Acute lymphoblastic leukemia: A population-based study of outcome in the United States based on the surveillance, epidemiology, and end results (SEER) database, 1980–2017. Am. J. Hematol. 2021, 96, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Oriol, A.; Vives, S.; Hernández-Rivas, J.M.; Tormo, M.; Heras, I.; Rivas, C.; Bethencourt, C.; Moscardó, F.; Bueno, J.; Grande, C.; et al. Outcome after relapse of acute lymphoblastic leukemia in adult patients included in four consecutive risk-adapted trials by the PETHEMA Study Group. Haematologica 2010, 95, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.K.; Richards, S.M.; Chopra, R.; Lazarus, H.M.; Litzow, M.R.; Buck, G.; Durrant, I.J.; Luger, S.M.; Marks, D.I.; Franklin, I.M.; et al. Outcome of 609 adults after relapse of acute lymphoblastic leukemia (ALL); an MRC UKALL12/ECOG 2993 study. Blood 2007, 109, 944–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gökbuget, N.; Stanze, D.; Beck, J.; Diedrich, H.; Horst, H.A.; Hüttmann, A.; Kobbe, G.; Kreuzer, K.A.; Leimer, L.; Reichle, A.; et al. Outcome of relapsed adult lymphoblastic leukemia depends on response to salvage chemotherapy, prognostic factors, and performance of stem cell transplantation. Blood 2012, 120, 2032–2041. [Google Scholar] [CrossRef]
- Tavernier, E.; Boiron, J.M.; Huguet, F.; Bradstock, K.; Vey, N.; Kovacsovics, T.; Delannoy, A.; Fegueux, N.; Fenaux, P.; Stamatoullas, A.; et al. Outcome of treatment after first relapse in adults with acute lymphoblastic leukemia initially treated by the LALA-94 trial. Leukemia 2007, 21, 1907–1914. [Google Scholar] [CrossRef] [Green Version]
- Bhojwani, D.; Yang, J.J.; Pui, C.H. Biology of childhood acute lymphoblastic leukemia. Pediatr. Clin. 2015, 62, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Hunger, S.P.; Mullighan, C.G. Redefining ALL classification: Toward detecting high-risk ALL and implementing precision medicine. Blood 2015, 125, 3977–3987. [Google Scholar] [CrossRef] [Green Version]
- Tasian, S.K.; Loh, M.L. Understanding the biology of CRLF2-overexpressing acute lymphoblastic leukemia. Crit. Rev. Oncog. 2011, 16, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Russell, L.J.; Capasso, M.; Vater, I.; Akasaka, T.; Bernard, O.A.; Calasanz, M.J.; Chandrasekaran, T.; Chapiro, E.; Gesk, S.; Griffiths, M.; et al. Deregulated expression of cytokine receptor gene, CRLF2, is involved in lymphoid transformation in B-cell precursor acute lymphoblastic leukemia. Blood 2009, 114, 2688–2698. [Google Scholar] [CrossRef]
- Mullighan, C.G.; Collins-Underwood, J.R.; Phillips, L.A.; Loudin, M.G.; Liu, W.; Zhang, J.; Ma, J.; Coustan-Smith, E.; Harvey, R.C.; Willman, C.L.; et al. Rearrangement of CRLF2 in B-progenitor- and Down syndrome-associated acute lymphoblastic leukemia. Nat. Genet. 2009, 41, 1243–1246. [Google Scholar] [CrossRef]
- Yoda, A.; Yoda, Y.; Chiaretti, S.; Bar-Natan, M.; Mani, K.; Rodig, S.J.; West, N.; Xiao, Y.; Brown, J.R.; Mitsiades, C.; et al. Functional screening identifies CRLF2 in precursor B-cell acute lymphoblastic leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertzberg, L.; Vendramini, E.; Ganmore, I.; Cazzaniga, G.; Schmitz, M.; Chalker, J.; Shiloh, R.; Iacobucci, I.; Shochat, C.; Zeligson, S.; et al. Down syndrome acute lymphoblastic leukemia, a highly heterogeneous disease in which aberrant expression of CRLF2 is associated with mutated JAK2: A report from the International BFM Study Group. Blood 2010, 115, 1006–1017. [Google Scholar] [CrossRef] [Green Version]
- Cario, G.; Zimmermann, M.; Romey, R.; Gesk, S.; Vater, I.; Harbott, J.; Schrauder, A.; Moericke, A.; Izraeli, S.; Akasaka, T.; et al. Presence of the P2RY8-CRLF2 rearrangement is associated with a poor prognosis in non-high-risk precursor B-cell acute lymphoblastic leukemia in children treated according to the ALL-BFM 2000 protocol. Blood 2010, 115, 5393–5397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, R.C.; Mullighan, C.G.; Chen, I.M.; Wharton, W.; Mikhail, F.M.; Carroll, A.J.; Kang, H.; Liu, W.; Dobbin, K.K.; Smith, M.A.; et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood 2010, 115, 5312–5321. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Ozaki, K.; Baumann, H.; Levin, S.D.; Puel, A.; Farr, A.G.; Ziegler, S.F.; Leonard, W.J.; Lodish, H.F. Cloning of a receptor subunit required for signaling by thymic stromal lymphopoietin. Nat. Immunol. 2000, 1, 59–64. [Google Scholar] [CrossRef]
- Park, L.S.; Martin, U.; Garka, K.; Gliniak, B.; Di Santo, J.P.; Muller, W.; Largaespada, D.A.; Copeland, N.G.; Jenkins, N.A.; Farr, A.G.; et al. Cloning of the murine thymic stromal lymphopoietin (TSLP) receptor: Formation of a functional heteromeric complex requires interleukin 7 receptor. J. Exp. Med. 2000, 192, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlmann, A.; Sebastian, K.; Borowski, A.; Krause, S.; Friedrich, K. Signal transduction by the atopy-associated human thymic stromal lymphopoietin (TSLP) receptor depends on Janus kinase function. Biol. Chem. 2010, 391, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Rochman, Y.; Kashyap, M.; Robinson, G.W.; Sakamoto, K.; Gomez-Rodriguez, J.; Wagner, K.U.; Leonard, W.J. Thymic stromal lymphopoietin-mediated STAT5 phosphorylation via kinases JAK1 and JAK2 reveals a key difference from IL-7-induced signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 19455–19460. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Kim, M.S.; Chaerkady, R.; Wu, X.; Huang, T.C.; Getnet, D.; Mitchell, C.J.; Palapetta, S.M.; Sharma, J.; O’Meally, R.N.; et al. TSLP signaling network revealed by SILAC-based phosphoproteomics. Mol. Cell. Proteom. 2012, 11, M112.017764. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Sharma, J.; Raju, R.; Palapetta, S.M.; Prasad, T.S.; Huang, T.C.; Yoda, A.; Tyner, J.W.; van Bodegom, D.; Weinstock, D.M.; et al. TSLP signaling pathway map: A platform for analysis of TSLP-mediated signaling. Database 2014, 2014, bau007. [Google Scholar] [CrossRef]
- Tasian, S.K.; Doral, M.Y.; Borowitz, M.J.; Wood, B.L.; Chen, I.M.; Harvey, R.C.; Gastier-Foster, J.M.; Willman, C.L.; Hunger, S.P.; Mullighan, C.G.; et al. Aberrant STAT5 and PI3K/mTOR pathway signaling occurs in human CRLF2-rearranged B-precursor acute lymphoblastic leukemia. Blood 2012, 120, 833–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comeau, M.R.; Ziegler, S.F. The influence of TSLP on the allergic response. Mucosal Immunol. 2010, 3, 138–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, T.R.; Pesce, J.T.; Mentink-Kane, M.M.; Madala, S.; Cheever, A.W.; Comeau, M.R.; Ziegler, S.F.; Wynn, T.A. Regulation of helminth-induced Th2 responses by thymic stromal lymphopoietin. J. Immunol. 2009, 182, 6452–6459. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.F.; Roan, F.; Bell, B.D.; Stoklasek, T.A.; Kitajima, M.; Han, H. The biology of thymic stromal lymphopoietin (TSLP). Adv. Pharm. 2013, 66, 129–155. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.B.; Kim, K.W.; Hong, J.Y.; Jee, H.M.; Sohn, M.H.; Kim, K.-E. Increased serum thymic stromal lymphopoietin in children with atopic dermatitis. Pediatr. Allergy Immunol. 2010, 21, e457–e460. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T.; Sugaya, M.; Fujita, H.; Saeki, H.; Tamaki, K. Increased serum thymic stromal lymphopoietin levels in patients with cutaneous T cell lymphoma. Clin. Exp. Dermatol. 2009, 34, 539–540. [Google Scholar] [CrossRef]
- Glück, J.; Rymarczyk, B.; Kasprzak, M.; Rogala, B. Increased Levels of Interleukin-33 and Thymic Stromal Lymphopoietin in Exhaled Breath Condensate in Chronic Bronchial Asthma. Int. Arch. Allergy Immunol. 2016, 169, 51–56. [Google Scholar] [CrossRef]
- Milford, T.A.; Su, R.J.; Francis, O.L.; Baez, I.; Martinez, S.R.; Coats, J.S.; Weldon, A.J.; Calderon, M.N.; Nwosu, M.C.; Botimer, A.R.; et al. TSLP or IL-7 provide an IL-7Rα signal that is critical for human B lymphopoiesis. Eur. J. Immunol. 2016, 46, 2155–2161. [Google Scholar] [CrossRef] [Green Version]
- Francis, O.L.; Milford, T.A.; Martinez, S.R.; Baez, I.; Coats, J.S.; Mayagoitia, K.; Concepcion, K.R.; Ginelli, E.; Beldiman, C.; Benitez, A.; et al. A novel xenograft model to study the role of TSLP-induced CRLF2 signals in normal and malignant human B lymphopoiesis. Haematologica 2016, 101, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Downes, C.E.; McClure, B.J.; McDougal, D.P.; Heatley, S.L.; Bruning, J.B.; Thomas, D.; Yeung, D.T.; White, D.L. JAK2 Alterations in Acute Lymphoblastic Leukemia: Molecular Insights for Superior Precision Medicine Strategies. Front. Cell Dev. Biol. 2022, 10, 942053. [Google Scholar] [CrossRef]
- Roll, J.D.; Reuther, G.W. CRLF2 and JAK2 in B-Progenitor Acute Lymphoblastic Leukemia: A Novel Association in Oncogenesis. Cancer Res. 2010, 70, 7347–7352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobah, M.L.; Liongue, C.; Ward, A.C. SOCS Proteins in Immunity, Inflammatory Diseases, and Immune-Related Cancer. Front. Med. 2021, 8, 727987. [Google Scholar] [CrossRef] [PubMed]
- Leibniz Institute DSMZ-MUTZ-5. Available online: https://www.dsmz.de/collection/catalogue/details/culture/ACC-490 (accessed on 11 July 2022).
- Leibniz Institute DSMZ-MHH-CALL-4. Available online: https://www.dsmz.de/collection/catalogue/details/culture/ACC-337 (accessed on 11 July 2022).
- Meyer, C.; MacLeod, R.A.F.; Quentmeier, H.; Janssen, J.W.G.; Coignet, L.J.; Dyer, M.J.S.; Drexler, H.G. Establishment of the B cell precursor acute lymphoblastic leukemia cell line MUTZ-5 carrying a (12;13) translocation. Leukemia 2001, 15, 1471–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, W.; Lin, Y.; Yang, X.; Li, B.; Liu, J.; He, R. Thymic stromal lymphopoietin (TSLP) inhibits human colon tumor growth by promoting apoptosis of tumor cells. Oncotarget 2016, 7, 16840–16854. [Google Scholar] [CrossRef] [Green Version]
- Liau, N.P.D.; Laktyushin, A.; Lucet, I.S.; Murphy, J.M.; Yao, S.; Whitlock, E.; Callaghan, K.; Nicola, N.A.; Kershaw, N.J.; Babon, J.J. The molecular basis of JAK/STAT inhibition by SOCS1. Nat. Commun. 2018, 9, 1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, C.E.; Whyte, C.S.; Gordon, P.; Barker, R.N.; Rees, A.J.; Wilson, H.M. A critical role for suppressor of cytokine signalling 3 in promoting M1 macrophage activation and function in vitro and in vivo. Immunology 2014, 141, 96–110. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Source of Cells | Ex Vivo Treatment |
|---|---|---|
| A1, A2, A3 | CRLF2 B-ALL cells from + T mice (low physiological TSLP levels) | 15,000 pg/mL TSLP for 3 days |
| B1, B2, B3 | CRLF2 B-ALL cells from + T mice (low physiological TSLP levels) | No TSLP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkashgari, H.R.; Ruiz-Jimenez, C.; Stoian, C.; Coats, J.S.; Baez, I.; Chirshev, E.; Martinez, S.R.; Dovat, S.; Francis-Boyle, O.L.; Casiano, C.A.; et al. TSLP as a Potential Therapy in the Treatment of CRLF2 B Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2023, 24, 474. https://doi.org/10.3390/ijms24010474
Alkashgari HR, Ruiz-Jimenez C, Stoian C, Coats JS, Baez I, Chirshev E, Martinez SR, Dovat S, Francis-Boyle OL, Casiano CA, et al. TSLP as a Potential Therapy in the Treatment of CRLF2 B Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2023; 24(1):474. https://doi.org/10.3390/ijms24010474
Chicago/Turabian StyleAlkashgari, Hossam R., Caleb Ruiz-Jimenez, Cornelia Stoian, Jacqueline S. Coats, Ineavely Baez, Evgeny Chirshev, Shannalee R. Martinez, Sinisa Dovat, Olivia L. Francis-Boyle, Carlos A. Casiano, and et al. 2023. "TSLP as a Potential Therapy in the Treatment of CRLF2 B Cell Acute Lymphoblastic Leukemia" International Journal of Molecular Sciences 24, no. 1: 474. https://doi.org/10.3390/ijms24010474
APA StyleAlkashgari, H. R., Ruiz-Jimenez, C., Stoian, C., Coats, J. S., Baez, I., Chirshev, E., Martinez, S. R., Dovat, S., Francis-Boyle, O. L., Casiano, C. A., & Payne, K. J. (2023). TSLP as a Potential Therapy in the Treatment of CRLF2 B Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences, 24(1), 474. https://doi.org/10.3390/ijms24010474