
It Runs in the Bromodomain Family: Speckled Proteins (SP) Play a Role in the Antitumor Immune Response in Solid Tumors

 , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Results
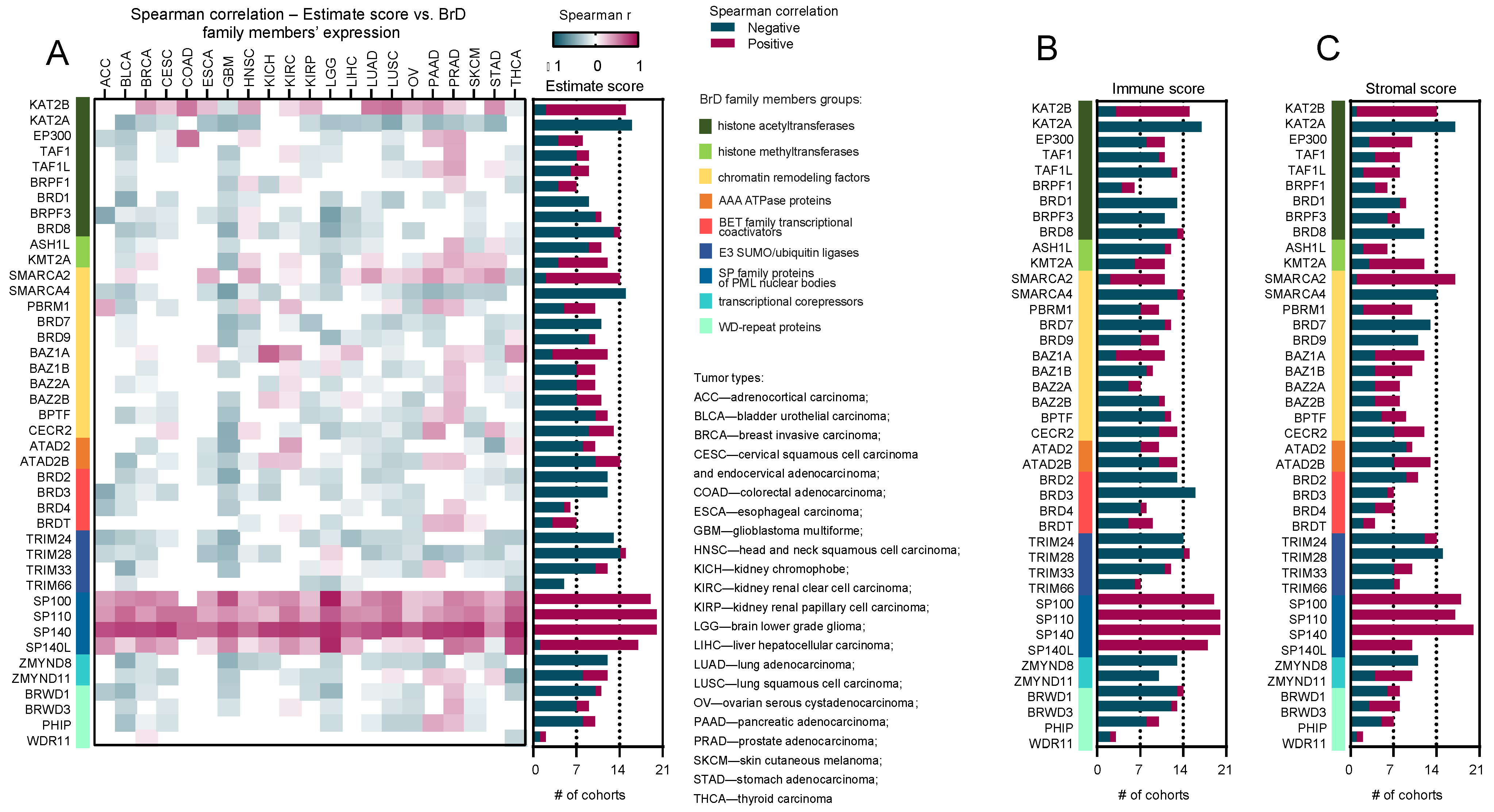
2.1. Correlation of the Expression of SP Family Members and Scores from the ESTIMATE Database Is Consistently Positive across All Tumor Types
2.2. Gene Set Cancer Analysis Shows a Strong Correlation between SP Family Expression and Immune Cell Infiltration
2.3. Significant Enrichment of Immune-Related Hallmark Gene Signatures in SP-Family-Associated Transcriptome Profiles
2.4. Better Survival Prognosis Is Linked with Immune Cells’ Infiltration Associated with SP-Family Expression
2.5. SP-Family Genes’ Expression Is Positively Correlated with Immunoinhibitors’ Abundance across 27 Tumor Types
3. Discussion
- BrD family members show a differential correlation between gene expression and immune-cell-infiltration status;
- SP-family genes (SP100, SP110, SP140, and SP140L) exhibit a consistently positive correlations with distinct immune scores, regardless of the tumor type;
- Based on the TIP database and GSCA data, the SP family shows a mostly positive correlation with T cells’ trafficking and infiltration into tumors. A high expression of SP-family genes is observed especially in solid tumors with great T-cell and macrophage abundance, and to a certain extent in tumors with enriched B-cell or neutrophil infiltrates;
- The transcriptome profiles associated with high expression of the SP-family genes are significantly enriched with allograft rejection, inflammatory response, IL2-STAT5, IL6-JAK-STAT3, TNF-α signaling via NF-κβ, and interferon-signaling signatures;
- SP genes’ expression might serve as a marker of immune cells’ invasion, while indirectly informing the chance of infiltration-dependent survival in various solid tumors.
- SP-family genes, especially SP140, show a highly positive correlation with the abundance of PDCD1 (PD-1), CTLA4, and CD274 (PD-L1) immunoinhibitors;
- As bromodomain present in SPs’ proteins is easily targetable, it may be an effective target for small molecule therapeutics to enhance immunotherapy outcomes.
4. Materials and Methods
4.1. TCGA Solid Tumor Types Selected for the Study
4.2. TCGA Genomic Data
4.3. Transcriptomic and Clinical Data
4.4. Estimate
4.5. Gene Set Cancer Analysis
4.6. Tracking Tumor Immunophenotype
4.7. Gene Set Enrichment Analysis
4.8. TISIDB
4.9. TIMER 2.0
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa-Pinheiro, P.; Montezuma, D.; Henrique, R.; Jerónimo, C. Diagnostic and Prognostic Epigenetic Biomarkers in Cancer. Epigenomics 2015, 7, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbioso, A.; Tambaro, F.P.; Dell’Aversana, C.; Altucci, L. Cancer Epigenetics: Moving Forward. PLoS Genet. 2018, 14, e1007362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flavahan, W.A.; Gaskell, E.; Bernstein, B.E. Epigenetic Plasticity and the Hallmarks of Cancer. Science 2017, 357, eaal2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaware, N.; Zhou, M.M. Bromodomain Biology and Drug Discovery. Nat. Struct. Mol. Biol. 2019, 26, 870–879. [Google Scholar] [CrossRef]
- Boyson, S.P.; Gao, C.; Quinn, K.; Boyd, J.; Paculova, H.; Frietze, S.; Glass, K.C. Functional Roles of Bromodomain Proteins in Cancer. Cancers 2021, 13, 3606. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Qi, J.; Picaud, S.; Shen, Y.; Smith, W.B.; Fedorov, O.; Morse, E.M.; Keates, T.; Hickman, T.T.; Felletar, I.; et al. Selective Inhibition of BET Bromodomains. Nature 2010, 468, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Filippakopoulos, P. Functions of Bromodomain-Containing Proteins and Their Roles in Homeostasis and Cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 246–262. [Google Scholar] [CrossRef]
- Fraschilla, I.; Jeffrey, K.L. The Speckled Protein (SP) Family: Immunity’s Chromatin Readers. Trends Immunol. 2020, 41, 572. [Google Scholar] [CrossRef]
- Bottomley, M.J.; Collard, M.W.; Huggenvik, J.I.; Liu, Z.; Gibson, T.J.; Sattler, M. The SAND Domain Structure Defines a Novel DNA-Binding Fold in Transcriptional Regulation. Nat. Struct. Biol. 2001, 8, 626–633. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Picaud, S.; Mangos, M.; Keates, T.; Lambert, J.P.; Barsyte-Lovejoy, D.; Felletar, I.; Volkmer, R.; Müller, S.; Pawson, T.; et al. Histone Recognition and Large-Scale Structural Analysis of the Human Bromodomain Family. Cell 2012, 149, 214–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, D.; Xiong, X.; He, Z.; Li, H. Multifaceted Histone H3 Methylation and Phosphorylation Readout by the Plant Homeodomain Finger of Human Nuclear Antigen Sp100C. J. Biol. Chem. 2016, 291, 12786–12798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perniola, R.; Musco, G. The Biophysical and Biochemical Properties of the Autoimmune Regulator (AIRE) Protein. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2014, 1842, 326–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.J.; Fernández-Montalván, A.E.; Badock, V.; Ott, C.J.; Holton, S.J.; von Ahsen, O.; Toedling, J.; Vittori, S.; Bradner, J.E.; Gorjánácz, M. ATAD2 Is an Epigenetic Reader of Newly Synthesized Histone Marks during DNA Replication. Oncotarget 2016, 7, 70323–70335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Cai, Z.; Kong, M.; Wu, A.; Hu, Z.; Wang, F.; Wang, H. Prognostic Significance of TRIM28 Expression in Patients with Breast Carcinoma. Open Med. 2021, 16, 472. [Google Scholar] [CrossRef]
- Jung, M.; Gelato, K.A.; Fernández-Montalván, A.; Siegel, S.; Haendler, B. Targeting BET Bromodomains for Cancer Treatment. Epigenomics 2015, 7, 487–501. [Google Scholar] [CrossRef]
- Zou, W. Immunosuppressive Networks in the Tumour Environment and Their Therapeutic Relevance. Nat. Rev. Cancer 2005, 5, 263–274. [Google Scholar] [CrossRef]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T Cells, Recurrence, and Survival in Epithelial Ovarian Cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Ogino, S.; Nosho, K.; Irahara, N.; Meyerhardt, J.A.; Baba, Y.; Shima, K.; Glickman, J.N.; Ferrone, C.R.; Mino-Kenudson, M.; Tanaka, N.; et al. Lymphocytic Reaction to Colorectal Cancer Is Associated with Longer Survival, Independent of Lymph Node Count, Microsatellite Instability, and CpG Island Methylator Phenotype. Clin. Cancer Res. 2009, 15, 6412–6420. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.K.; Donskov, F.; Marcussen, N.; Nordsmark, M.; Lundbeck, F.; Von Der Maase, H. Presence of Intratumoral Neutrophils Is an Independent Prognostic Factor in Localized Renal Cell Carcinoma. J. Clin. Oncol. 2009, 27, 4709–4717. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring Tumour Purity and Stromal and Immune Cell Admixture from Expression Data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.R.; Zhang, Q.; Lei, Q.; Luo, M.; Xie, G.Y.; Wang, H.; Guo, A.Y. ImmuCellAI: A Unique Method for Comprehensive T-Cell Subsets Abundance Prediction and Its Application in Cancer Immunotherapy. Adv. Sci. 2020, 7, 1902880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Tayyari, F.; Pinnaduwage, D.; Bayani, J.; Bartlett, J.M.S.; Mulligan, A.M.; Bull, S.B.; Andrulis, I.L. Tumoral BRD4 Expression in Lymph Node-Negative Breast Cancer: Association with T-Bet+ Tumor-Infiltrating Lymphocytes and Disease-Free Survival. BMC Cancer 2018, 18, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.; Shao, S.; Zhang, Y.; Luo, A.; Zhao, L.; Zhang, L.; Gu, S.; Zhao, X. BRD4 inhibition suppresses PD-L1 expression in triple-negative breast cancer. Exp Cell Res 2020, 392, 112034. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Ouyang, S.S.; Chen, Y.L.; Li, P.; Xu, H.W.; Zhu, S.L. BRD4/8/9 Are Prognostic Biomarkers and Associated with Immune Infiltrates in Hepatocellular Carcinoma. Aging 2020, 12, 17541–17567. [Google Scholar] [CrossRef]
- Adeegbe, D.O.; Liu, Y.; Lizotte, P.H.; Kamihara, Y.; Aref, A.R.; Almonte, C.; Dries, R.; Li, Y.; Liu, S.; Wang, X.; et al. Synergistic Immunostimulatory Effects and Therapeutic Benefit of Combined Histone Deacetylase and Bromodomain Inhibition in Non-Small Cell Lung Cancer. Cancer Discov. 2017, 7, 852–867. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.; Keany, M.P.; Dong, H.; Al-Alem, L.F.; Pandya, U.M.; Lazo, S.; Boehnke, K.; Lynch, K.N.; Xu, R.; Zarrella, D.T.; et al. Enhanced Efficacy of Simultaneous PD-1 and PD-L1 Immune Checkpoint Blockade in High-Grade Serous Ovarian Cancer. Cancer Res. 2021, 81, 158–173. [Google Scholar] [CrossRef]
- Mao, W.; Ghasemzadeh, A.; Freeman, Z.T.; Obradovic, A.; Chaimowitz, M.G.; Nirschl, T.R.; McKiernan, E.; Yegnasubramanian, S.; Drake, C.G. Immunogenicity of Prostate Cancer Is Augmented by BET Bromodomain Inhibition. J. Immunother Cancer 2019, 7, 277. [Google Scholar] [CrossRef]
- Leal, A.S.; Liu, P.; Krieger-Burke, T.; Ruggeri, B.; Liby, K.T. The Bromodomain Inhibitor, INCB057643, Targets Both Cancer Cells and the Tumor Microenvironment in Two Preclinical Models of Pancreatic Cancer. Cancers 2020, 13, 96. [Google Scholar] [CrossRef]
- Lou, W.; Gao, K.; Xu, C.; Li, Q. Bromodomain-Containing Protein 9 Is a Prognostic Biomarker Associated with Immune Infiltrates and Promotes Tumor Malignancy through Activating Notch Signaling Pathway in Negative HIF-2α Clear Cell Renal Cell Carcinoma. IUBMB Life 2021, 73, 1334–1347. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Deng, C.; Pang, B.; Zhang, X.; Liu, W.; Liao, G.; Yuan, H.; Cheng, P.; Li, F.; Long, Z.; et al. TIP: A Web Server for Resolving Tumor Immunophenotype Profiling. Cancer Res. 2018, 78, 6575–6580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for Analysis of Tumor-Infiltrating Immune Cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Zhang, S.-C.; Hu, Z.-Q.; Long, J.-H.; Zhu, G.-M.; Wang, Y.; Jia, Y.; Zhou, J.; Ouyang, Y.; Zeng, Z. Clinical Implications of Tumor-Infiltrating Immune Cells in Breast Cancer. J. Cancer 2019, 10, 6175–6184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, S.; Wei, M.; Wang, S.; Dong, J.; Wei, J. Pan-Cancer Analysis of Immune Cell Infiltration Identifies a Prognostic Immune-Cell Characteristic Score (ICCS) in Lung Adenocarcinoma. Front. Immunol. 2020, 11, 1218. [Google Scholar] [CrossRef]
- Zhong, L.; Yang, Z.; Lei, D.; Li, L.; Song, S.; Cao, D.; Liu, Y. Bromodomain 4 Is a Potent Prognostic Marker Associated with Immune Cell Infiltration in Breast Cancer. Basic Clin. Pharmacol. Toxicol. 2021, 128, 169–182. [Google Scholar] [CrossRef]
- Czerwinska, P.; Jaworska, A.M.; Wlodarczyk, N.A.; Cisek, M.; Karwacka, M.; Lipowicz, J.; Ostapowicz, J.; Rosochowicz, M.; Mackiewicz, A.A. The Association between Bromodomain Proteins and Cancer Stemness in Different Solid Tumor Types. Int. J. Cancer 2022, 150, 1838. [Google Scholar] [CrossRef]
- Miranda, A.; Hamilton, P.T.; Zhang, A.W.; Pattnaik, S.; Becht, E.; Mezheyeuski, A.; Bruun, J.; Micke, P.; de Reynies, A.; Nelson, B.H. Cancer Stemness, Intratumoral Heterogeneity, and Immune Response across Cancers. Proc. Natl. Acad. Sci. USA 2019, 116, 9020–9029. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Alexander Cronkite, D.; Basavappa, M.; Saunders, T.L.; Adiliaghdam, F.; Amatullah, H.; Morrison, S.A.; Pagan, J.D.; Anthony, R.M.; Tonnerre, P.; et al. Maintenance of Macrophage Transcriptional Programs and Intestinal Homeostasis by Epigenetic Reader SP140. Sci. Immunol. 2017, 2, eaag3160. [Google Scholar] [CrossRef] [Green Version]
- Collados Rodríguez, M. The Fate of Speckled Protein 100 (Sp100) During Herpesviruses Infection. Front. Cell. Infect. Microbiol. 2021, 10, 900. [Google Scholar] [CrossRef]
- Bechter, O.; Schöffski, P. Make Your Best BET: The Emerging Role of BET Inhibitor Treatment in Malignant Tumors. Pharmacol. Ther. 2020, 208, 107479. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pan, X.; Zhang, W.; Guo, H.; Cheng, S.; He, Q.; Yang, B.; Ding, L. Epigenetic Strategies Synergize with PD-L1/PD-1 Targeted Cancer Immunotherapies to Enhance Antitumor Responses. Acta Pharm. Sin. B 2020, 10, 723. [Google Scholar] [CrossRef] [PubMed]
- Spivey, T.L.; Uccellini, L.; Ascierto, M.L.; Zoppoli, G.; De Giorgi, V.; Delogu, L.G.; Engle, A.M.; Thomas, J.M.; Wang, E.; Marincola, F.M.; et al. Gene Expression Profiling in Acute Allograft Rejection: Challenging the Immunologic Constant of Rejection Hypothesis. J. Transl. Med. 2011, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, K.; Saio, M.; Umemura, N.; Kikuchi, A.; Takahashi, T.; Osada, S.; Yoshida, K. Th1 Polarization in the Tumor Microenvironment Upregulates the Myeloid-Derived Suppressor-like Function of Macrophages. Cell. Immunol. 2021, 369, 104437. [Google Scholar] [CrossRef] [PubMed]
- Jhunjhunwala, S.; Hammer, C.; Delamarre, L. Antigen Presentation in Cancer: Insights into Tumour Immunogenicity and Immune Evasion. Nat. Rev. Cancer 2021, 21, 298–312. [Google Scholar] [CrossRef]
- Jones, D.M.; Read, K.A.; Oestreich, K.J. Dynamic Roles for IL-2-STAT5 Signaling in Effector and Regulatory CD4+ T Cell Populations. J. Immunol. (Baltim. Md. 1950) 2020, 205, 1721–1730. [Google Scholar] [CrossRef]
- Yu, H.; Pardoll, D.; Jove, R. STATs in Cancer Inflammation and Immunity: A Leading Role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Arce Vargas, F.; Furness, A.J.S.; Solomon, I.; Joshi, K.; Mekkaoui, L.; Lesko, M.H.; Miranda Rota, E.; Dahan, R.; Georgiou, A.; Sledzinska, A.; et al. Fc-Optimized Anti-CD25 Depletes Tumor-Infiltrating Regulatory T Cells and Synergizes with PD-1 Blockade to Eradicate Established Tumors. Immunity 2017, 46, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 Signalling Axis in Cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Zinatizadeh, M.R.; Schock, B.; Chalbatani, G.M.; Zarandi, P.K.; Jalali, S.A.; Miri, S.R. The Nuclear Factor Kappa B (NF-KB) Signaling in Cancer Development and Immune Diseases. Genes Dis. 2021, 8, 287. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schober, P.; Boer, C.; Schwarte, L.A. Correlation Coefficients: Appropriate Use and Interpretation. Anesth. Analg. 2018, 126, 1763. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Hu, F.F.; Xia, M.X.; Han, L.; Zhang, Q.; Guo, A.Y. GSCALite: A Web Server for Gene Set Cancer Analysis. Bioinformatics 2018, 34, 3771–3772. [Google Scholar] [CrossRef]
- Janssens, A.C.J.W.; Martens, F.K. Reflection on Modern Methods: Revisiting the Area under the ROC Curve. Int. J. Epidemiol. 2020, 49, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular Signatures Database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef]
- Ru, B.; Wong, C.N.; Tong, Y.; Zhong, J.Y.; Zhong, S.S.W.; Wu, W.C.; Chu, K.C.; Wong, C.Y.; Lau, C.Y.; Chen, I.; et al. TISIDB: An Integrated Repository Portal for Tumor-Immune System Interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Number | Function | Proteins |
|---|---|---|
| I | Histone acetyltransferase | PCAF, GCN5L2, p300/CBP, TAF1, TAF1L, BRPF1A/B, BRPF2 (BRD1), BRPF3, BRD8 (SMAP) |
| II | Histone methyltransferase | ASH1L, MLL |
| III | Chromatin remodeling factor | SMARCA2A/2B, SMARCA4, PBRM1, BRD7, BRD9, BAZ1A, BAZ1B, BAZ2A, BAZ2B, BPTF, CECR2 |
| IV | AAA ATPase protein | ATAD2, ATAD2B |
| V | BET family transcriptional coactivator | BRD2, BRD3, BRD4, BRDT |
| VI | E3 SUMO/ubiquitin ligase | TRIM24, TRIM28, TRIM33A/B, TRIM66 |
| VII | SP-family proteins of PML nuclear bodies | SP100, SP110, SP140, SP140L |
| VIII | Transcriptional corepressor | ZMYND8, ZMYND11 |
| IX | WD-repeat protein | BRWD1, BRWD3, PHIP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosochowicz, M.A.; Lipowicz, J.M.; Karwacka, M.I.; Ostapowicz, J.; Cisek, M.; Mackiewicz, A.A.; Czerwinska, P. It Runs in the Bromodomain Family: Speckled Proteins (SP) Play a Role in the Antitumor Immune Response in Solid Tumors. Int. J. Mol. Sci. 2023, 24, 549. https://doi.org/10.3390/ijms24010549
Rosochowicz MA, Lipowicz JM, Karwacka MI, Ostapowicz J, Cisek M, Mackiewicz AA, Czerwinska P. It Runs in the Bromodomain Family: Speckled Proteins (SP) Play a Role in the Antitumor Immune Response in Solid Tumors. International Journal of Molecular Sciences. 2023; 24(1):549. https://doi.org/10.3390/ijms24010549
Chicago/Turabian StyleRosochowicz, Monika Anna, Julia Maria Lipowicz, Marianna Iga Karwacka, Julia Ostapowicz, Malgorzata Cisek, Andrzej Adam Mackiewicz, and Patrycja Czerwinska. 2023. "It Runs in the Bromodomain Family: Speckled Proteins (SP) Play a Role in the Antitumor Immune Response in Solid Tumors" International Journal of Molecular Sciences 24, no. 1: 549. https://doi.org/10.3390/ijms24010549
APA StyleRosochowicz, M. A., Lipowicz, J. M., Karwacka, M. I., Ostapowicz, J., Cisek, M., Mackiewicz, A. A., & Czerwinska, P. (2023). It Runs in the Bromodomain Family: Speckled Proteins (SP) Play a Role in the Antitumor Immune Response in Solid Tumors. International Journal of Molecular Sciences, 24(1), 549. https://doi.org/10.3390/ijms24010549