
Effect of Secretion Efficiency of Mutant KRAS Neoantigen by Lactococcus lactis on the Immune Response of a Mucosal Vaccine Delivery Vehicle Targeting Colorectal Cancer
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. In Silico Analysis of KRAS Fusion Proteins
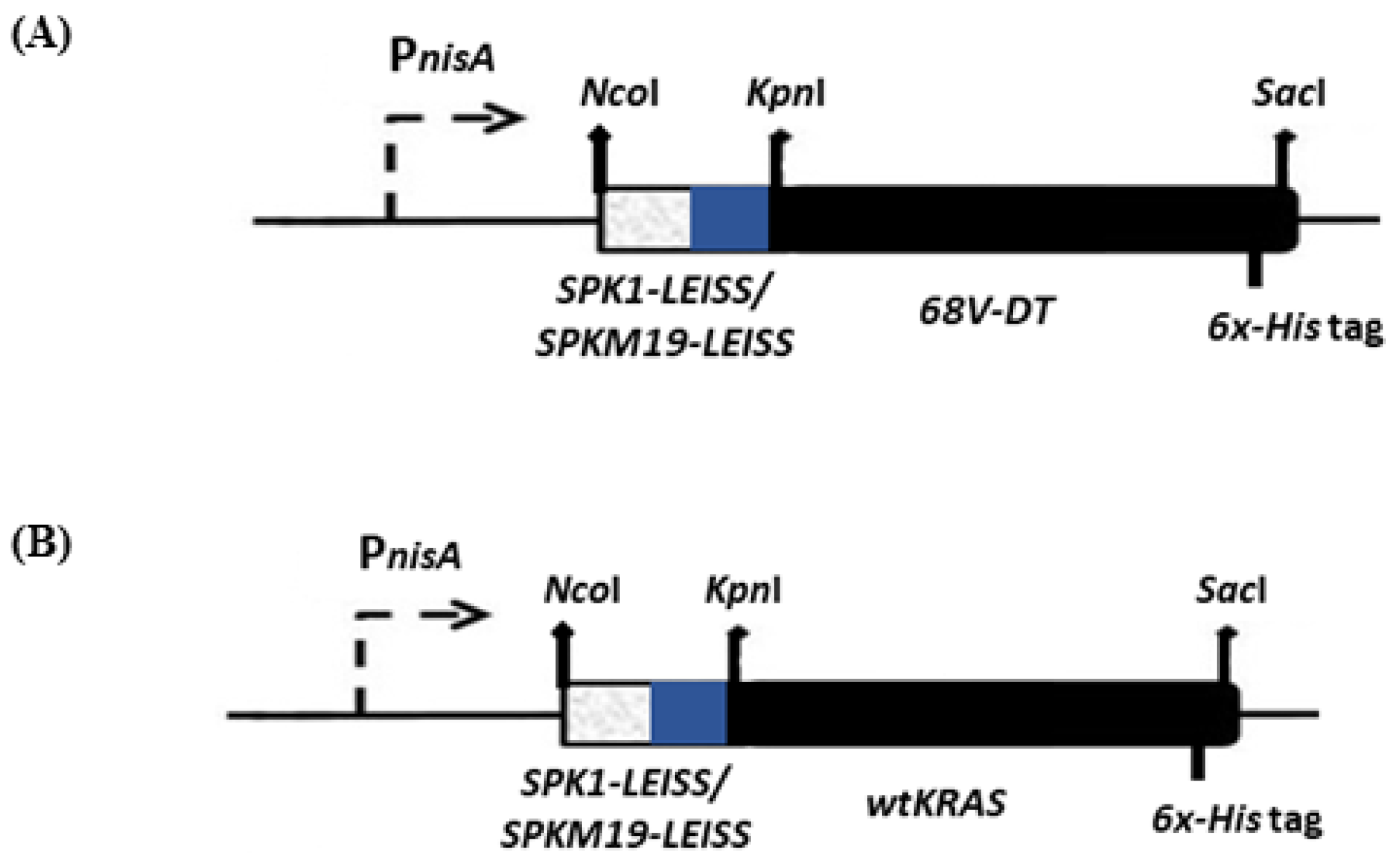
2.2. Lactococcal Recombinant Strains Secreting KRAS Mimotopes
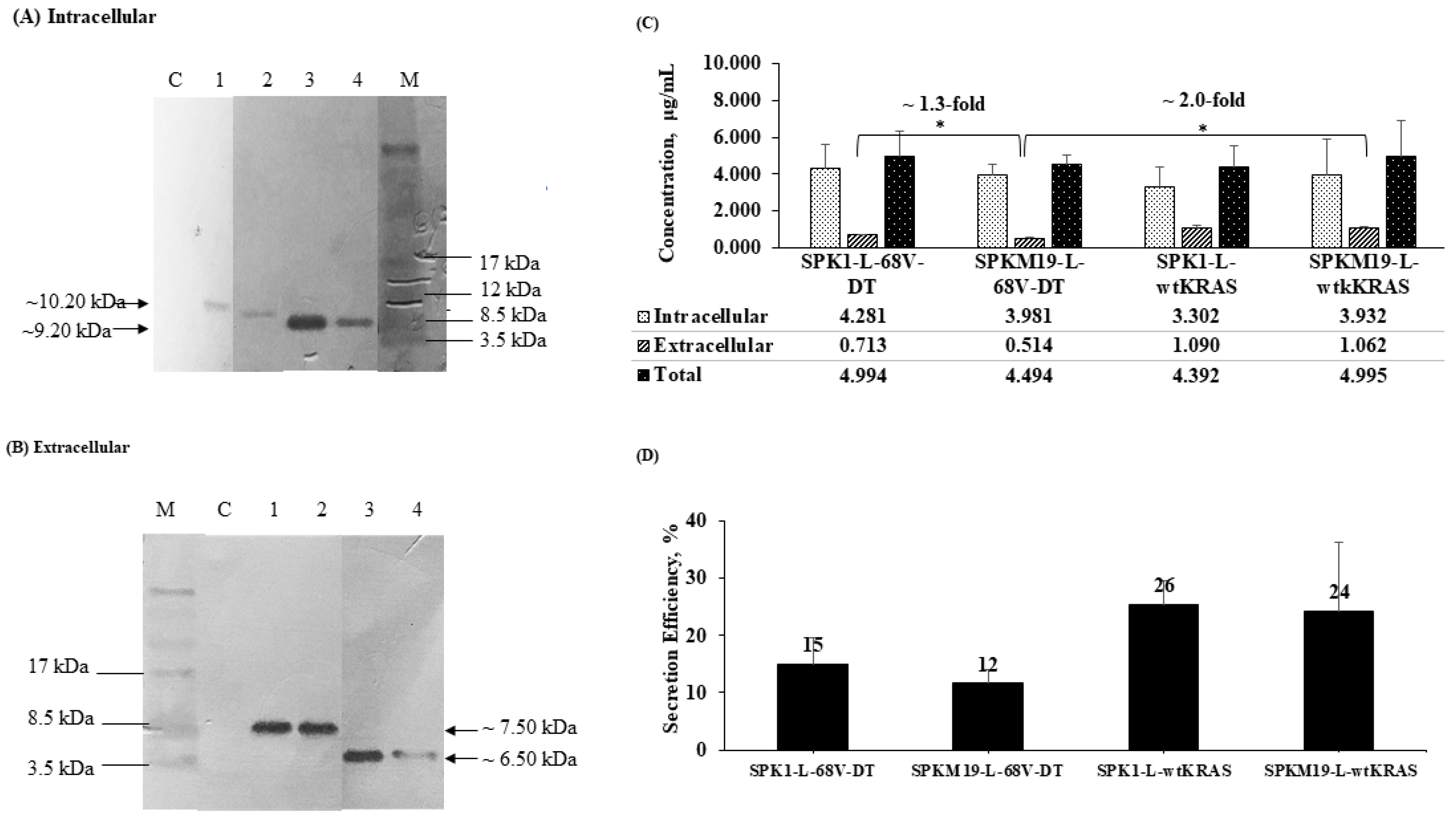
2.3. Yield and Secretion Efficiency of KRAS Mimotopes at 6 h Expression in L. lactis
2.4. Elicitation of KRAS-Specific sIgA following Oral Administration
3. Discussion
4. Materials and Methods
4.1. In Silico Characterization of Signal Peptide and Mature Proteins
4.2. Bacterial Strains, Plasmids, and Antibiotics
4.3. PCR Amplification of SPs and KRAS Mimotopes
4.4. Construction of Secretory Plasmids in L. lactis
4.5. Expression and Characterization of KRAS Mimotopes by Western Blotting
4.6. Quantification of KRAS Mimotopes by Indirect Enzyme-Linked Immunosorbent (ELISA) Assay
4.7. Double Coating of Recombinants L. lactis-Secreting KRASs into Edible Films for Oral Immunization
4.8. Animal, Immunization Schedule and Sample Extraction
4.9. Immunoglobulins (sIgA and IgG) Detection by Indirect ELISA
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Kanthan, R.; Senger, J.; Kanthan, S.C. Molecular Events in Primary and Metastatic Colorectal Carcinoma: A Review. Pathol. Res. Int. 2012, 2012, 597497. [Google Scholar] [CrossRef]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet 2014, 383, 1490–1502. [Google Scholar] [CrossRef]
- Fernández-Medarde, A.; Santos, E. Ras in cancer and developmental diseases. Genes Cancer 2011, 2, 344–358. [Google Scholar] [CrossRef]
- McCormick, F. KRAS as a Therapeutic Target. Clin. Cancer Res. 2015, 21, 1797–1801. [Google Scholar] [CrossRef]
- Arrington, A.K.; Heinrich, E.L.; Lee, W.; Duldulao, M.; Patel, S.; Sanchez, J.; Garcia-Aguilar, J.; Kim, J. Prognostic and predictive roles of KRAS mutation in colorectal cancer. Int. J. Mol. Sci. 2012, 13, 12153–12168. [Google Scholar] [CrossRef]
- Huang, D.; Sun, W.; Zhou, Y.; Li, P.; Chen, F.; Chen, H.; Xia, D.; Xu, E.; Lai, M.; Wu, Y.; et al. Mutations of key driver genes in colorectal cancer progression and metastasis. Cancer Metastasis Rev. 2018, 37, 173–187. [Google Scholar] [CrossRef]
- Caiazza, F.; Elliott, L.; Fennelly, D.; Sheahan, K.; Doherty, G.A.; Ryan, E.J. Targeting EGFR in metastatic colorectal cancer beyond the limitations of KRAS status: Alternative biomarkers and therapeutic strategies. Biomark. Med. 2015, 9, 363–375. [Google Scholar] [CrossRef]
- Bignucolo, A.; De Mattia, E.; Cecchin, E.; Roncato, R.; Toffoli, G. Pharmacogenomics of Targeted Agents for Personalization of Colorectal Cancer Treatment. Int. J. Mol. Sci. 2017, 18, 1522. [Google Scholar] [CrossRef]
- Carbone, D.P.; Ciernik, I.F.; Kelley, M.J.; Smith, M.C.; Nadaf, S.; Kavanaugh, D.; Maher, V.E.; Stipanov, M.; Contois, D.; Johnson, B.E.; et al. Immunization with mutant p53- and K-ras-derived peptides in cancer patients: Immune response and clinical outcome. J. Clin. Oncol. 2005, 23, 5099–5107. [Google Scholar] [CrossRef]
- Toubaji, A.; Achtar, M.; Provenzano, M.; Herrin, V.E.; Behrens, R.; Hamilton, M.; Bernstein, S.; Venzon, D.; Gause, B.; Marincola, F.; et al. Pilot study of mutant ras peptide-based vaccine as an adjuvant treatment in pancreatic and colorectal cancers. Cancer Immunol. Immunother. 2008, 57, 1413–1420. [Google Scholar] [CrossRef]
- Rahma, O.E.; Hamilton, J.M.; Wojtowicz, M.; Dakheel, O.; Bernstein, S.; Liewehr, D.J.; Steinberg, S.M.; Khleif, S.N. The immunological and clinical effects of mutated ras peptide vaccine in combination with IL-2, GM-CSF, or both in patients with solid tumors. J. Trans. Med. 2014, 12, 55. [Google Scholar] [CrossRef]
- Pan, J.; Zhang, Q.; Palen, K.; Wang, L.; Qiao, L.; Johnson, B.; Sei, S.; Shoemaker, R.H.; Lubet, R.A.; Wang, Y.; et al. Potentiation of Kras peptide cancer vaccine by avasimibe, a cholesterol modulator. eBioMedicine 2019, 49, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.W.R.; Tan, P.J.; Hoo, W.P.Y.; Liew, D.S.; Teo, M.Y.M.; Siak, P.Y.; Ng, S.M.; Tan, E.W.; Abdul Rahim, R.; Lim, R.L.H.; et al. In silico-guided sequence modifications of K-ras epitopes improve immunological outcome against G12V and G13D mutant KRAS antigens. PeerJ 2018, 7, e5056. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Joshi, M.D.; Singhania, S.; Ramsey, K.H.; Murthy, A.K. Peptide Vaccine: Progress and Challenges. Vaccines 2014, 2, 515–536. [Google Scholar] [CrossRef]
- Wells, J.M.; Mercenier, A. Mucosal delivery of therapeutic and prophylactic molecules using Lactic Acid Bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Nouaille, S.; Ribeiro, L.A.; Miyoshi, A.; Pontes, D.; Le Loir, Y.; Oliveira, S.C.; Langella, P.; Azevedo, V. Heterologous protein production and delivery systems for Lactococcus lactis. Genet. Mol. Res. 2003, 2, 102–111. [Google Scholar]
- Wyszyńska, A.; Kobierecka, P.; Bardowski, J.; Jagusztyn-Krynicka, E.K. Lactic acid bacteria--20 years exploring their potential as live vectors for mucosal vaccination. Appl. Microbiol. Biotehncol. 2015, 99, 2967–2977. [Google Scholar] [CrossRef]
- Vilander, A.C.; Dean, G.A. Adjuvant Strategies for Lactic Acid Bacterial Mucosal Vaccines. Vaccines 2019, 7, 150. [Google Scholar] [CrossRef]
- Song, A.A.; In LL, A.; Lim SH, E.; Rahim, R.A. A review on Lactococcus lactis: From food to factory. Microbial Cell. Factories 2017, 16, 55. [Google Scholar] [CrossRef]
- Alias, N.A.R.; Song, A.A.; Alitheen, N.B.; Rahim, R.A.; Othman, S.S.; In, L.L.A. Optimization of signal peptide via site-directed mutagenesis for enhanced secretion of heterologous proteins in Lactococcus Lactis. Int. J. Mol. Sci. 2022, 23, 10044. [Google Scholar] [CrossRef] [PubMed]
- Le Loir, Y.; Gruss, A.; Ehrlich, S.D.; Langella, P. A nine-residue synthetic propeptide enhances secretion efficiency of heterologous proteins in Lactococcus lactis. J. Bacteriol. 1998, 180, 1895–1903. [Google Scholar] [CrossRef]
- Le Loir, Y.; Nouaille, S.; Commissaire, J.; Brétigny, L.; Gruss, A.; Langella, P. Signal Peptide and propeptide optimization for heterologous protein secretion in Lactococcus lactis. Appl. Environ. Microbiol. 2001, 67, 4119–4127. [Google Scholar] [CrossRef]
- Choo, K.H.; Ranganathan, S. Flanking signal and mature peptide residues influence signal peptide cleavage. BMC Bioinform. 2008, 9, S15. [Google Scholar] [CrossRef] [PubMed]
- Kajava, A.V.; Zolov, S.N.; Kalinin, A.E.; Nesmeyanova, M.A. The net charge of the first 18 residues of the mature sequence affects protein translocation across the cytoplasmic membrane of Gram-negative bacteria. J. Bacteriol. 2000, 182, 2163–2169. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S.; Piao, D.C.; Jiang, T.; Bok, J.D.; Cho, C.S.; Lee, Y.S.; Kang, S.K.; Choi, Y.J. Recombinant interleukin 6 with M cell-targeting moiety produced in Lactococcus lactis IL1403 as a potent mucosal adjuvant for peroral immunization. Vaccine 2015, 33, 1959–1967. [Google Scholar] [CrossRef]
- Bermúdez-Humarán, L.G.; Langella, P.; Commissaire, J.; Gilbert, S.; Le Loir, Y.; L’Haridon, R.; Corthier, G. Controlled intra- or extracellular production of staphylococcal nuclease and ovine omega interferon in Lactococcus lactis. FEMS Microbiol. Lett. 2003, 224, 307–313. [Google Scholar] [CrossRef]
- Steidler, L.; Robinson, K.; Chamberlain, L.; Schofield, K.M.; Remaut, E.; Le Page, R.W.; Wells, J.M. Mucosal delivery of murine interleukin-2 (IL-2) and IL-6 by recombinant strains of Lactococcus lactis coexpressing antigen and cytokine. Infect. Immun. 1998, 66, 3183–3189. [Google Scholar] [CrossRef]
- Steidler, L.; Hans, W.; Schotte, L.; Neirynck, S.; Obermeier, F.; Falk, W.; Fiers, W.; Remaut, E. Treatment of murine colitis by Lactococcus lactis secreting interleukin-10. Science 2000, 289, 1352–1355. [Google Scholar] [CrossRef]
- Diethelm-Okita, B.M.; Okita, D.K.; Banaszak, L.; Conti-Fine, B.M. Universal epitopes for human CD4+ cells on tetanus and diphtheria toxins. J. Infect. Dis. 2000, 181, 1001–1009. [Google Scholar] [CrossRef]
- Bramwell, V.W.; Somavarapu, S.; Outschoorn, I.; Alpar, H.O. Adjuvant action of melittin following intranasal immunisation with tetanus and diphtheria toxoids. J. Drug Target. 2003, 11, 525–530. [Google Scholar] [CrossRef]
- Alias, N.A.R.; Song, A.A.; Alitheen, N.B.; Othman, S.S.; In, L.L.A.; Rahim, R.A. Incorporation of propeptide LEISSTCDA for enhanced protein secretion in Lactococcus lactis. In Proceedings of the Multidisciplinary International Conference 2023, Selangor, Malaysia, 7 March 2023. [Google Scholar]
- Le Loir, Y.; Azevedo, V.; Oliveira, S.C.; Freitas, D.A.; Miyoshi, A.; Bermúdez-Humarán, L.G.; Nouaille, S.; Ribeiro, L.A.; Leclercq, S.; Gabriel, J.E.; et al. Protein secretion in Lactococcus lactis: An efficient way to increase the overall heterologous protein production. Microb. Cell Factories 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhong, J.; Liang, X. Improvement of human interferon alpha secretion by Lactococcus lactis. Biotechnol. Lett. 2010, 32, 1271–1277. [Google Scholar] [CrossRef]
- Freitas, D.A.; Leclerc, S.; Miyoshi, A.; Oliveira, S.C.; Sommer, P.S.; Rodrigues, L.; Correa Junior, A.; Gautier, M.; Langella, P.; Azevedo, V.A.; et al. Secretion of Sterptomyces tandae antifungal protein 1 by Lactococcus lactis. Braz. J. Med. Biol. Res. 2005, 38, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.W.; Tan, K.Y.; Phang, L.V.; Kumar, P.V.; In, L.L.A. Enhanced gastrointestinal survivability of recombinant Lactococcus lactis using a double coated mucoadhesive film approach. PLoS ONE 2019, 14, e0219912. [Google Scholar] [CrossRef]
- Sachan, N.; Pushkar, S.; Jha, A.; Bhattcharya, A. Sodium alginate: The wonder polymer for controlled drug delivery. J. Pharm. Res. 2009, 2, 1191–1199. [Google Scholar]
- Hariyadi, D.M.; Islam, N. Current Status of Alginate in Drug Delivery. Adv. Pharmacol. Pharm. Sci. 2020, 2020, 8886095. [Google Scholar] [CrossRef]
- Wu, Y.; Weller, C.L.; Hamouz, F.; Cuppett, S.; Schnepf, M. Moisture loss and lipid oxidation for precooked ground-beef patties packaged in edible starch-alginate-based composite films. J. Food Sci. 2001, 66, 486–493. [Google Scholar] [CrossRef]
- Fazilah, N.F.; Hamidon, N.H.; Ariff, A.B.; Khayat, M.E.; Wasoh, H.; Halim, M. Microencapsulation of Lactococcus lactis Gh1 with Gum Arabic and Synsepalum dulcificum via spray drying for potential inclusion in functional yogurt. Molecules 2019, 24, 1422. [Google Scholar] [CrossRef]
- Divya, J.B.; Nampoothiri, K.M. Encapsulated Lactococcus lactis with enhanced gastrointestinal survival for the development of folate enriched functional foods. Bioresour. Technol. 2015, 188, 226–230. [Google Scholar] [CrossRef]
- McConnell, E.L.; Basit, A.W.; Murdan, S. Measurements of rat and mouse gastrointestinal pH, fluid and lymphoid tissue, and implications for in-vivo experiments. J. Pharm. Pharmacol. 2008, 60, 63–70. [Google Scholar] [CrossRef]
- Koido, S.; Ohkusa, T.; Homma, S.; Namiki, Y.; Takakura, K.; Saito, K.; Ito, Z.; Kobayashi, H.; Kajihara, M.; Uchiyama, K.; et al. Immunotherapy for colorectal cancer. World J. Gastroenterol. 2013, 19, 8531–8542. [Google Scholar] [CrossRef] [PubMed]
- Aggerbeck, H.; Gizurarson, S.; Wantzin, J.; Heron, I. Intranasal booster vaccination against diphtheria and tetanus in man. Exploration 1997, 1, 26. [Google Scholar] [CrossRef] [PubMed]
- Qiao, N.; Du, G.; Zhong, X.; Sun, X. Recombinant lactic acid bacteria as promising vectors for mucosal vaccination. Nat. Methods 2021, 8, 785–786. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- De Ruyter, P.G.; Kuipers, O.P.; de Vos, W.M. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl. Environ. Microbiol. 1996, 62, 3662–3667. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, O.P.; de Ruyter PG, G.A.; Kleerebezem, M.; de Vos, W.M. Quorum sensing-controlled gene expression in lactic acid bacteria. J. Biotechnol. 1998, 64, 15–21. [Google Scholar] [CrossRef]
- Hoo WP, Y.; Siak, P.Y.; Alias NA, R.; Wong, J.J.; Tan, E.W.; Song AA, L.; Rahim, R.A.; In, L.L.A. K-ras peptide mimotope induces a humoral immune response against G12V K-ras antigen in balb/c mice. Asia-Pac. J. Mol. Biol. Biotechnol. 2020, 28, 22–35. [Google Scholar]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus lactis subsp. cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar]
- Koontz, L. TCA precipitation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2014; Volume 541, pp. 3–10. [Google Scholar]
- Carbone, L.; Carbone, E.T.; Yi, E.M.; Bauer, D.B.; Lindstrom, K.A.; Parker, J.M.; Austin, J.A.; Seo, Y.; Gandhi, A.D.; Wilkerson, J.D. Assessing cervical dislocation as a humane euthanasia method in mice. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 352–356. [Google Scholar] [PubMed]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93. [Google Scholar] [PubMed]
- Siak, P.Y.; Wong, K.Y.; Song, A.A.L.; Rahim, R.A.; In, L.L.A. K-Ras peptide mimotope induces antigen specific Th1 and B-cell immune responses against G12A-mutated K-ras antigen in Balb/c mice. Vaccines 2021, 9, 195. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Fusion Genes | SP Length (aa) | D-Score | SPase I Cleavage Site (−3 to −1) | Net Charge of SP | First 10 Residues of MP (+1 to +10) | Net Charge of First 10 Residues of MP |
|---|---|---|---|---|---|---|
| SPK1-LEISS-68V-DT | 23 | 0.710 | VHA | +3 | LEISSTCDAG | −1 |
| SPKM19-LEISS-68V-DT | 23 | 0.815 | AHA | +2 | ALEISSTCDA | −1 |
| SPK1-LEISS-wtKRAS | 23 | 0.711 | VHA | +3 | LEISSTCDAG | −1 |
| SPKM19-LEISS-wtKRAS | 23 | 0.808 | AHA | +2 | ALEISSTCDA | −1 |
| Fusion Genes | MP Length | MP Size (kDa) | Charge of First 10 MP | Charge of First 18 MP | MP Total Charge | MP pI | MP Gravy Index | MP Aliphatic Index | MP Instability Index |
|---|---|---|---|---|---|---|---|---|---|
| SPK1-L-68V-DT | 61 | 6.53 | −1 | −2 | −2 | 6.65 | −0.02 | 95.90 | 21.24 |
| SPKM19-L-68V-DT | 62 | 6.59 | −1 | −2 | −2 | 6.66 | 0.01 | 95.97 | 21.06 |
| SPK1-L-wtKRAS | 67 | 7.43 | −1 | −2 | −6 | 5.46 | −0.35 | 87.16 | 25.97 |
| SPKM19-L-wtKRAS | 68 | 7.50 | −1 | −2 | −6 | 5.46 | −0.32 | 87.35 | 25.73 |
| SPK1-L-NUC | 186 | 20.80 | −2 | −3 | +3 | 8.78 | −0.86 | 65.06 | 33.77 |
| SPKM19-L-NUC | 187 | 20.87 | −2 | −3 | +3 | 8.78 | −0.85 | 65.24 | 33.64 |
| Bacterial Strains and Plasmids | Relevant Features | Reference |
|---|---|---|
| Strains | ||
| E. coli TOP 10 | Propagation host for pCR™-Blunt II-TOPO® | Invitrogen, Waltham, MA, USA |
| L. lactis NZ9000 | Expression host for nisin-inducible plasmid pNZ8048 | De Ruyter et al. (1996) [49] |
| Plasmids | ||
| pCR™-Blunt II-TOPO® | E. coli cloning plasmid containing pUC ori, Knr, lacZα genes; 3.5 kb | Invitrogen, Waltham, MA, USA |
| pNZ8048 | L. lactis nisin inducible expression plasmid; containing PnisA promoter and Cmr; 3.3 kb | Kuipers et al. (1998) [50] |
| pNZ-SPKM19-LEISS-wtKRAS | pNZ8048 harboring SPSPKM19-LEISS-wild-type KRAS cassettes with C-terminal six-histidine tag; 4.0 kb | This study |
| pNZ-SPK1-LEISS-wtKRAS | pNZ8048 carrying SPSPK1-LEISS-wild-type KRAS cassettes with C-terminal six-histidine tag; 3.97 kb | This study |
| pNZ-SPKM19-LEISS-68V-DT | pNZ8048 carrying SPSPKM19-LEISS-68V-DT cassettes with C-terminal six-histidine tag; 3.92 kb | This study |
| pNZ-SPK1-LEISS-68V-DT | pNZ8048 carrying SPSPK1-LEISS-68V-DT cassettes with C-terminal six histidine tag; 3.98 kb | This study |
| Gene (SP-MP)/Plasmid | Primer | Sequence from 5′ to 3′ | Ta (°C) | Amplicon Size (bp) |
|---|---|---|---|---|
| SPKM19-LEISS | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 47 | 119 |
| (2) R-M19LEISS | GGTACCTGCATCACAAGTCGACGATATTTCGAGAGCATGTACTG | |||
| SPK1-LEISS | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 47 | 110 |
| (3) R-Spk1LEISS | GGTACCTGCATCACAAGTCGACGATATTTCGAGAGCATGTACATTC | |||
| KRAS 68V-DTD | (4) F-68V-DT | GGTACCATGTATAAATTAGATGTTGTTG | 48 | 165 |
| (5) R-68V-DT | GAGCTCCTAATGATGATGATGATGATGT | |||
| KRAS wild-type (wtKRAS) | (6) F-wtKRAS | GGTACCATGACTGAATATAAACTTGTGGTAGTT | 52 | 183 |
| (7) R-wtKRAS | GAGCTCCTAATGATGATGATGATGATGG | |||
| SPKM19-LEISS-wtKRAS | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 53 | 292 |
| (7) R-wtKRAS | GAGCTCCTAATGATGATGATGATGATGG | |||
| SPK1-LEISS-wtKRAS | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 53 | 289 |
| (7) R-wtKRAS | GAGCTCCTAATGATGATGATGATGATGG | |||
| SPKM19-LEISS-68V-DT | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 46 | 277 |
| (5) R-68V-DT | GAGCTCCTAATGATGATGATGATGATGT | |||
| SPK1-LEISS-68V-DT | (1) F-Spk1 | CCATGGCTATGAAAAAAATATTAAC | 51 | 274 |
| (5) R-68V-DT | GAGCTCCTAATGATGATGATGATGATGT | |||
| pNZ8048 | (8) F-pNZ8048 | TATTGTCGATAACGCGAGCA | 55 | Varies with insert size |
| (9) R-pNZ8048 | CGTTTCAAGCCTTGGTTTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alias, N.A.R.; Hoo, W.P.Y.; Siak, P.Y.; Othman, S.S.; Mohammed Alitheen, N.B.; In, L.L.A.; Abdul Rahim, R.; Song, A.A.-L. Effect of Secretion Efficiency of Mutant KRAS Neoantigen by Lactococcus lactis on the Immune Response of a Mucosal Vaccine Delivery Vehicle Targeting Colorectal Cancer. Int. J. Mol. Sci. 2023, 24, 8928. https://doi.org/10.3390/ijms24108928
Alias NAR, Hoo WPY, Siak PY, Othman SS, Mohammed Alitheen NB, In LLA, Abdul Rahim R, Song AA-L. Effect of Secretion Efficiency of Mutant KRAS Neoantigen by Lactococcus lactis on the Immune Response of a Mucosal Vaccine Delivery Vehicle Targeting Colorectal Cancer. International Journal of Molecular Sciences. 2023; 24(10):8928. https://doi.org/10.3390/ijms24108928
Chicago/Turabian StyleAlias, Nur Aqlili Riana, Winfrey Pui Yee Hoo, Pui Yan Siak, Siti Sarah Othman, Noorjahan Banu Mohammed Alitheen, Lionel Lian Aun In, Raha Abdul Rahim, and Adelene Ai-Lian Song. 2023. "Effect of Secretion Efficiency of Mutant KRAS Neoantigen by Lactococcus lactis on the Immune Response of a Mucosal Vaccine Delivery Vehicle Targeting Colorectal Cancer" International Journal of Molecular Sciences 24, no. 10: 8928. https://doi.org/10.3390/ijms24108928