
Lunasin and Its Epigenetic Impact in Cancer Chemoprevention

 , ,
, ,
Abstract
:1. Introduction
2. Characterization of Lunasin
2.1. Chemical Structure of Lunasin and Its Structure-Related Chemopreventive Activities
2.2. Lunasin Abundance in Soybeans and Other Plants
2.3. Digestion and Bioavailability of Lunasin
3. Lunasin and Its Effects on Normal Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Concentration | Time of Exposition | Type of Lunasin | Proliferation | Ref. |
|---|---|---|---|---|---|
| Human normal mammary epithelial MCF-10A | 1–320 µM | 24–48 h | Synthetic | No significant effect | [41] |
| Human normal mammary epithelial MCF-10A | 5–200 µM | 24–72 h | Synthetic | No significant effect | [42] |
| Normal bronchial epithelial HBE135-E6E7 BEAS-2B | 1–100 µM | 24–72 h | Natural | No significant effect | [43] |
| Normal colon fibroblast CCD-33Co | 1–100 µM | 24 h | Natural | No significant effect | [29,44] |
| Mouse fibroblast NIH-3T3 | 10 µM | 24–96 h | Synthetic | No significant effect | [39] |
| Mouse fibroblast NIH-3T3 | 0.01–10 µM | 24–72 h | Synthetic | No significant effect | [45] |
| Mouse macrophage RAW 264.7 | 10–50 µM | 24 h | Natural | No significant effect | [46] |
| Mouse macrophage RAW 264.7 | 0.2–200 μM | 24 h | Synthetic | No significant effect | [47] |
| Permanent endothelial EA.hy926 | 0.05–120 μM | 72 h | Recombinant | No significant effect | [40] |
4. Chemopreventive Properties of Lunasin
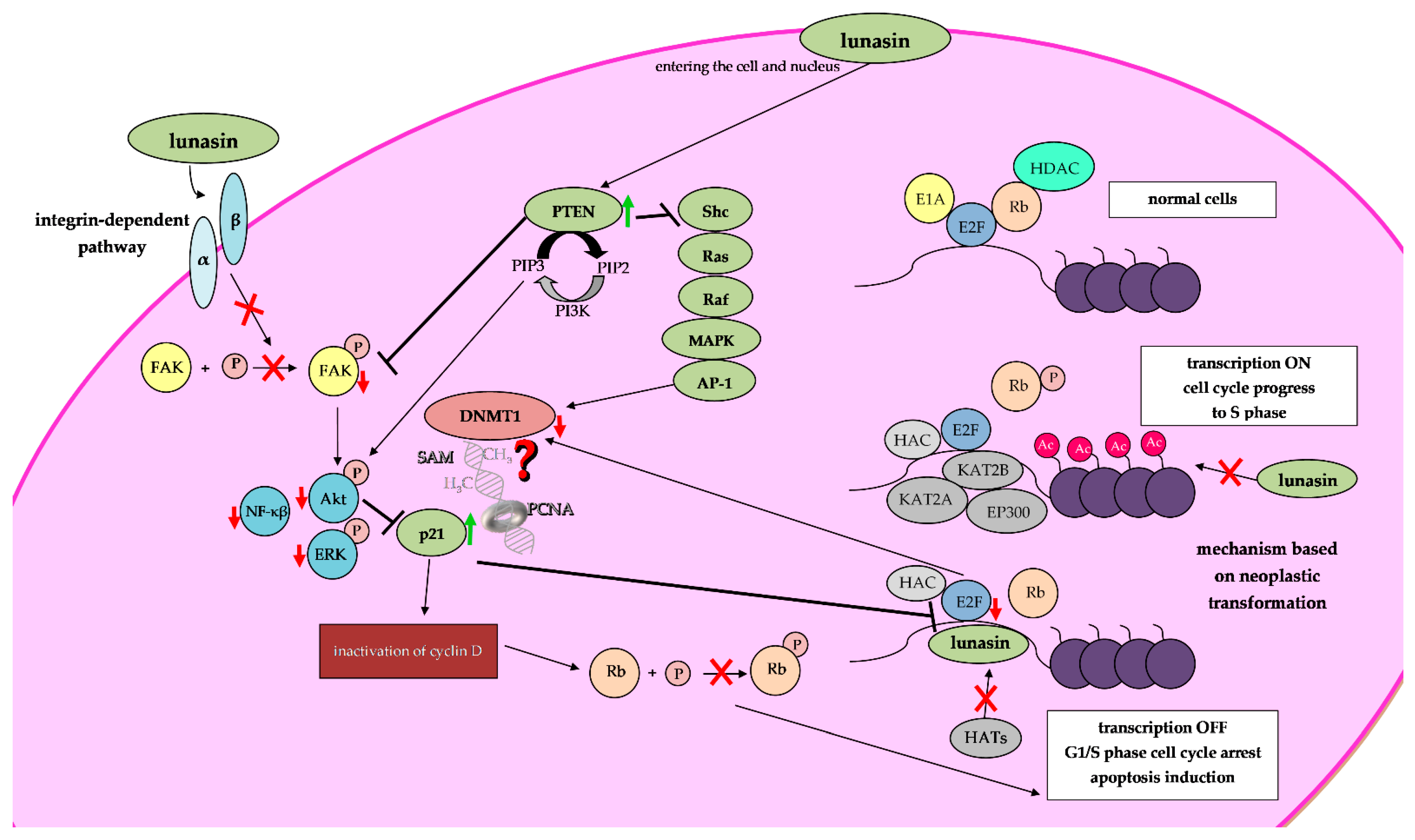
4.1. Epigenetic Mechanisms of Lunasin Anti-Cancer Activity
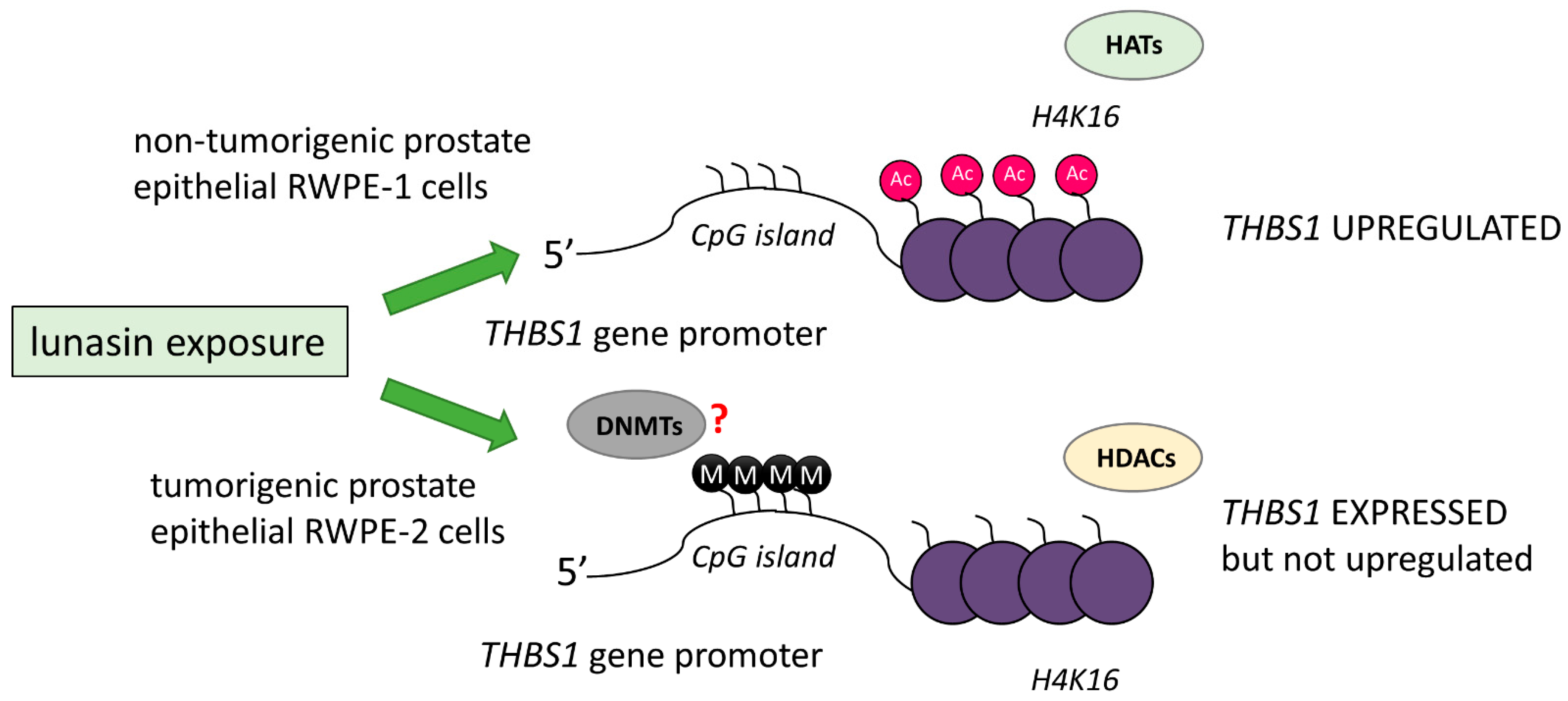
4.1.1. Inhibition of Histone Acetylation upon Lunasin Exposure
4.1.2. Lunasin and Its Potential to Modulate DNA Methylation
| Gene | Concentration | Exposure Time | Effects | Ref. |
|---|---|---|---|---|
| BAX | 1 µM | 24 h | 2.8-fold increase in expression | [44] |
| 10 µM | 6.6-fold increase in expression | |||
| 50 µM | 7.2-fold increase in expression | |||
| 10 µM | 24 h | 2.2-fold increase in expression | [29] | |
| BCL-2 | 1 µM | 24 h | 1.4-fold decrease in expression | [44] |
| 10 µM | 2-fold decrease in expression | |||
| 50 µM | 2.8-fold decrease in expression | |||
| 50 µM | 24 h | 2-fold decrease in expression | [75] | |
| 10 µM | 24 h | 2-fold decrease in expression | [29] | |
| CASP3 | 1 mg LES/mL (50 µM) | 24 h | 12-fold increase in expression | [30] |
| 10 µM | 24 h | 1.8-fold increase in expression | [29] | |
| 40 and 80 µM | 72 h | 1.9-fold increase in expression | [76] | |
| 1 µM | 24 h | 1.5-fold activity increase | [44] | |
| 10 µM | 1.6-fold activity increase | |||
| 50 µM | 1.8-fold activity increase | |||
| CASP8 | 1 mg LES/mL (50 µM) | 24 h | 5-fold increase in expression | [30] |
| 10 µM | 24 h | 1.6-fold increase in expression | [49] | |
| 75 µM | ||||
| CASP9 | 1 mg LES/mL (50 µM) | 24 h | 6-fold increase in expression | [30] |
| p21 | 1 µM | 24 h | 3.6-fold increase in expression | [44] |
| 10 µM | 4.7-fold increase in expression | |||
| 50 µM | 7.3-fold increase in expression | |||
| 10 µM | 24 h | 2.2-fold increase in expression | [29] | |
| p27 | 10 µM | 24 h | 2.3-fold increase in expression | [29] |
| nCLU | 1 µM | 24 h | 3.7-fold increase in expression | [44] |
| 10 µM | 5.5-fold decrease in expression | |||
| 50 µM | 5.6-fold decrease in expression | |||
| 50 µM | 24 h | 2-fold decrease in expression | [75] | |
| MALT1 | 50 µM | 24 h | 1.9-fold decrease in expression | [75] |
| PTEN | 2 µM | 24 h | 2.4-fold increase in expression | [73] |
| 2 µM | 24 h | 2.5-fold increase in expression | [74] |
4.2. In Vitro and In Vivo Studies on Lunasin Chemopreventive Activity
| Cell Line | Concentration | Time of Exposure | Type of Lunasin | Effects | Ref. |
|---|---|---|---|---|---|
| Human breast cancer cell line MCF-7 | 10 µM | 72 h | Synthetic | No significant effect | [39] |
| Human breast cancer cell line MCF-7 MDA-MB-231 | 5–200 µM | 24–72 h | Synthetic | Decrease in cell proliferation IC50 MCF-7–(48 h) 232 μM IC50 MDA-MD-231–(48 h) 153 μM | [42] |
| Human breast cancer cell line MDA-MB-231 | 10–200 µM | 48 h | Synthetic | Decrease in cell proliferation IC50 MDA-MB-231–181 µM | [52] |
| Human breast cancer cell line MDA-MB-231 | 0.1–200 µM | 72 h | Synthetic | Decrease in cell proliferation IC50 MDA-MB-231–181µM | [49] |
| Human breast cancer cell line MCF-7MDA-MB-231 | 1–320 µM | 24, 48 h | Synthetic | Decrease in cell proliferation IC50 MCF-7–(24 h) 508.6 µM, (48 h) 431.9 µM IC50 MDA-MB-231–(24 h) 224.7 µM, (48 h) 194.9 µM | [41] |
| Human breast cancer cell line MDA-MB-231 | 1–100 µM | 48 h | Recombinant | Decrease in cell proliferation IC50 MDA-MB-231–56.73 µM | [78] |
| Human colon cancer cell line HT-29 | 1–100 µM | 24 h | Natural 90% | Decrease in cell proliferation IC50 HT-29–61.7 µM | [44] |
| Human colon cancer cell line: HT-29 KM12L4 RKO HCT-116 | 1–100 µM | 24 h | Natural >90% | Decrease in cell proliferation IC50 HT-29–61.7 µM IC50 KM12L4–13 µM IC50 RKO–21.6 µM IC50 HCT-116–26.3 µM | [29] |
| Human colon cancer cell line HT-29 Caco-2 | 10–200 µM | 24, 48, 72 h | Synthetic | Decrease in cell proliferation greatest for HT-29 cells after 72 h incubation at concentration of lunasin of 200 µM (23.8% of non-viable cells) No cytotoxicity effects in Caco-2 cells after 72 h exposure | [79] |
| Human colon cancer cell line HCT-116 | 2 µM | 48 h | Synthetic | Decrease in cell proliferation Increase in the number of apoptotic cells | [74] |
| Human colon cancer cell line HCT-116 | 5–160 µM | 72 h | Synthetic | Decrease in cell proliferation IC50 HCT-116–107.5 µM | [76] |
| Human colon cancer cell line Caco-2 | 0.5–25 µM | 24 h | Synthetic | No significant effect | [81] |
| Human colon cancer cell line HCT-116 Human hepatoma cell line HepG2 | 1–100 µM | 48 h | Recombinant | Decrease in cell proliferation IC50 HCT-116–64.25 µM No cytotoxicity for HepG2 cells | [78] |
| Human hepatoma cell line HepG2 | 0.5–50 µM | 20 h | Synthetic | No significant effect | [80] |
| Mouse leukemia cell line L1210 | 1–80 µM | 24 h | Natural 98% | Decrease in cell proliferation IC50–13.9 µM | [82] |
| Mouse leukemia cell line L1210 | 1–100 µM | 24 h | Natural 98% 27% (LES) | Decrease in cell proliferation IC50–14 µM IC50–16 µM | [30] |
| Human non-small-cell lung cancer cell line: H661 H1299 H460 A549 | 1–100 µM | 24, 48, 72 h | Natural 99% | Decrease in cell proliferation (H661 cell line) IC50 H661 cells–(72 h) 63.9 µM No cytotoxicity for other cell lines | [43] |
| Human gastric adenocarcinoma cell line AGS | 10–200 µM | 24, 48, 72 h | Synthetic | Decrease in cell proliferation (AGS cells) at concentrations of 100 µM–7% and 200 µM–15% (average percentage) | [79] |
| Mouse skin cancer cell line B16-F10 Human skin cancer cell line A-375 | 0.03–550 µM | 24 h | Natural 40% | Decrease in cell proliferation IC50 B16-F10–330 µM IC50 A-375–370 µM | [77] |
| In Vivo Model | Type of Cancer | Lunasin | Dose of Lunasin (#, Groups) | Administration Method | Scheme of Treatment | Effects | References |
|---|---|---|---|---|---|---|---|
| 12-week-old female SENCAR mice | Mouse skin cancer initiated by DMBA (initiator) and TPA (promoter) | Synthetic | #1: control DMBA and TPA (n = 6); #2: solvents (n = 8); #3: 2.5 µg Lun/week (n = 9); #4: 25 µg Lun/week (n = 9); #5: 250 µg Lun/week (n = 9); | Percutaneous to the dorsal side, shaved | Before tumor induction 1 week 2 times a week After tumor induction 19 weeks 2 times a week | Reduction in skin tumor incidence by ~70%—dermal application of lunasin, 250 μg/week; decrease in tumor yield/mouse, and delay of the appearance of tumors by 2 weeks relative to the control 2.5 and 25 μg/week—decreasing trend in tumor incidence and yield (not statistically significant) | [23] |
| 6-week-old athymic NCr-nu/nu mice | Human breast cancer (1 × 107 MDA-MB-231 cells injected subcutaneously) | Natural | #1: control solvents; #2: Lun 20 mg/kg bw; #3: Lun 4 mg/kg bw (n = 8/group); | Intraperitoneal injection | Before cancer cells implantation 2 months 3 times a week | Decrease in tumor incidence by 49% at 20 mg/kg bw lunasin pretreated group and 33% at 4 mg/kg bw lunasin pretreated group compared with the vehicle-treated group | [83] |
| 7- to 8-week-old male outbred homozygous nude mice | Human non-small-cell lung cancer (H1299 cells 2 × 106 injected subcutaneously) | Natural | #1: control solvents; #2: Lun 30 mg/kg bw (n = 10/group); | Intraperitoneal injection | After cancer cells implantation 32 days 1 time a day | Reduction in tumor volume by 63% compared to the control group | [43] |
| 6- to 8-week-old male athymic nude mice (Jackson #002019) | Human melanoma (A375 cells 2.5 × 106 injected subcutaneously) | Natural | #1: control solvent (n = 8); #2: Lun 30 mg/kg bw (n = 10); | Intraperitoneal injection | After cancer cells implantation 34 days 1 time a day | Reduction in tumor volume by 55% and wet tumor weight by 46% | [55] |
| 6-week-old male C57BL/6 mice | Mouse lung carcinoma (LLC cells 1 × 105 injected subcutaneously) | Natural | #1: control solvents; #2: Lun 10 mg/kg bw; #3: Lun 30 mg/kg bw (n = 6–10/group); | Intraperitoneal injection | After cancer cells implantation 22 days 1 time a day | Inhibition of tumor growth initiated by LLC cells at 30 mg/kg dose of lunasin by 55% | [56] |
| Mouse melanoma (B16-F0 cells 1 × 106 injected subcutaneously) | #1: control solvents; #2: Lun 10 mg/kg bw; #3: Lun 30 mg/kg bw (n = 6–10/group); | Inhibition of tumor growth initiated by B16-F0 cells at the 30 mg/kg dose of lunasin by 60% | |||||
| 6-week-old female Sprague Dawley (SD) rats | Breast cancer rat model induced by DMBA | Natural | #1: control solvent (n = 4); #2: control DMBA (n = 4); #3: tamoxifen 10 mg/kg bw (n = 4); #4: Lun 500 mg/kg bw (n = 4); #5: tamoxifen + Lun (10 + 500 mg/kg bw) (n = 4); | Intraperitoneal injection | After tumor induction 8 weeks | 8 weeks of treatment shown that tamoxifen, lunasin and a combination of tamoxifen and lunasin could reduce tumor volume (>50%) and tumor weight when compared to control (DMBA) | [84] |
| 6- to 8-week-old mice | Human colon cancer cells (spleen implantation of 1 × 106 KM12L4 human colon cancer cells) | Natural | #1: 100 μL of PBS (n = 10); #2: Lun 4 mg/kg bw (n = 9); | Intraperitoneal injection By oral gavage | After cancer cells implantation 28 days 1 time a day | Reduction in CRC liver metastasis by 50% compared to the control group and the liver weight/body weight ratio by 23% | [85] |
| #1: 100 μL PBS (n = 6); #2: Lun 8 mg/kg bw (n = 5); #3: Lun 20 mg/kg bw (n = 3); | Oral administration—reduction in the number of liver metastasis (not statistically significant) |
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cancer Today. Available online: http://gco.iarc.fr/today/home (accessed on 2 June 2022).
- Vineis, P.; Wild, C.P. Global cancer patterns: Causes and prevention. Lancet 2013, 383, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean Bioactive Peptides and Their Functional Properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, A.J.; Fleshner, N.; Casselman, B.; Trachtenberg, J. Nutritional aspects of prostate cancer: A review. Can. J. Urol. 2000, 7, 927–935. [Google Scholar] [PubMed]
- Greenwald, P.; Clifford, C.; Milner, J. Diet and cancer prevention. Eur. J. Cancer 2001, 37, 948–965. [Google Scholar] [CrossRef] [PubMed]
- Pollard, M.; Wolter, W. Prevention of spontaneous prostate-related cancer in Lobund-Wistar rats by a soy protein isolate/isoflavone diet. Prostate 2000, 45, 101–105. [Google Scholar] [CrossRef]
- Odani, S.; Koide, T.; Ono, T. Amino acid sequence of a soybean (Glycine max) seed polypeptide having a poly(L-aspartic acid) structure. J. Biol. Chem. 1987, 262, 10502–10505. [Google Scholar] [CrossRef]
- Galvez, A.F.; de Lumen, B.O. A soybean cDNA encoding a chromatin-binding peptide inhibits mitosis of mammalian cells. Nat. Biotechnol. 1999, 17, 495–500. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B. Lunasin, a novel seed peptide for cancer prevention. Peptides 2009, 30, 426–430. [Google Scholar] [CrossRef]
- Eden, S.; Hashimshony, T.; Keshet, I.; Cedar, H.; Thorne, A.W. DNA methylation models histone acetylation. Nature 1998, 394, 842. [Google Scholar] [CrossRef]
- Nan, X.; Campoy, F.J.; Bird, A. MeCP2 Is a Transcriptional Repressor with Abundant Binding Sites in Genomic Chromatin. Cell 1997, 88, 471–481. [Google Scholar] [CrossRef]
- D’alessio, A.C.; Szyf, M. Epigenetic tête-à-tête: The bilateral relationship between chromatin modifications and DNA methylation. Biochem. Cell Biol. 2006, 84, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Kaufman-Szymczyk, A.; Majewski, G.; Lubecka-Pietruszewska, K.; Fabianowska-Majewska, K. The Role of Sulforaphane in Epigenetic Mechanisms, Including Interdependence between Histone Modification and DNA Methylation. Int. J. Mol. Sci. 2015, 16, 29732–29743. [Google Scholar] [CrossRef] [PubMed]
- Lubecka-Pietruszewska, K.; Kaufman-Szymczyk, A.; Stefanska, B.; Cebula-Obrzut, B.; Smolewski, P.; Fabianowska-Majewska, K. Sulforaphane Alone and in Combination with Clofarabine Epigenetically Regulates the Expression of DNA Methylation-Silenced Tumour Suppressor Genes in Human Breast Cancer Cells. J. Nutr. Nutr. 2015, 8, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Patel, S.N.; Tollefsbol, T.O. Sulforaphane Causes Epigenetic Repression of hTERT Expression in Human Breast Cancer Cell Lines. PLoS ONE 2010, 5, e11457. [Google Scholar] [CrossRef]
- Yoon, H.-G.; Lee, Y.-H.; Kwak, J.; Choi, H.-K.; Choi, K.-C.; Kim, S.; Lee, J.; Jun, W.; Park, H.-J.; Yoon, H.G. EGCG suppresses prostate cancer cell growth modulating acetylation of androgen receptor by anti-histone acetyltransferase activity. Int. J. Mol. Med. 2012, 30, 69–74. [Google Scholar] [CrossRef]
- Lubecka, K.; Kaufman-Szymczyk, A.; Cebula-Obrzut, B.; Smolewski, P.; Szemraj, J.; Fabianowska-Majewska, K. Novel Clofarabine-Based Combinations with Polyphenols Epigenetically Reactivate Retinoic Acid Receptor Beta, Inhibit Cell Growth, and Induce Apoptosis of Breast Cancer Cells. Int. J. Mol. Sci. 2018, 19, 3970. [Google Scholar] [CrossRef] [PubMed]
- Lubecka, K.; Kurzava, L.; Flower, K.; Buvala, H.; Zhang, H.; Teegarden, D.; Camarillo, I.; Suderman, M.; Kuang, S.; Andrisani, O.; et al. Stilbenoids remodel the DNA methylation patterns in breast cancer cells and inhibit oncogenic NOTCH signaling through epigenetic regulation of MAML2 transcriptional activity. Carcinogenesis 2016, 37, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Galvez, A.F.; Huang, L.; Magbanua, M.M.J.; Dawson, K.; Rodriguez, R.L. Differential Expression of Thrombospondin (THBS1) in Tumorigenic and Nontumorigenic Prostate Epithelial Cells in Response to a Chromatin-Binding Soy Peptide. Nutr. Cancer 2011, 63, 623–636. [Google Scholar] [CrossRef]
- Wan, X.; Liu, H.; Sun, Y.; Zhang, J.; Chen, X.; Chen, N. Lunasin: A promising polypeptide for the prevention and treatment of cancer. Oncol. Lett. 2017, 13, 3997–4001. [Google Scholar] [CrossRef]
- Galvez, A.F.; Revilleza, M.J.R.; de Lumen, B.O. A novel methionine-rich protein from soybean cotyledon: Cloning and characterization of cDNA (accession no. AF005030). Plant Gene Register #PGR97-103. Plant Physiol. 1997, 114, 1567–1569. [Google Scholar]
- Lule, V.K.; Garg, S.; Pophaly, S.D.; Hitesh; Tomar, S.K. Potential Health Benefits of Lunasin: A Multifaceted Soy-Derived Bioactive Peptide. J. Food Sci. 2015, 80, R485–R494. [Google Scholar] [CrossRef] [PubMed]
- Galvez, A.F.; Chen, N.; Macasieb, J.; de Lumen, B.O. Chemopreventive property of a soybean peptide (lunasin) that binds to deacetylated histones and inhibits acetylation. Cancer Res. 2001, 61, 7473–7478. [Google Scholar] [PubMed]
- Matsuki, K.; Sasho, T.; Nakagawa, K.; Tahara, M.; Sugioka, K.; Ochiai, N.; Ogino, S.; Wada, Y.; Moriya, H. RGD peptide-induced cell death of chondrocytes and synovial cells. J. Orthop. Sci. 2008, 13, 524–532. [Google Scholar] [CrossRef] [PubMed]
- De Souza, S.M.A.; Fernandes, T.V.A.; Kalume, D.E.; Lima, L.M.T.R.; Pascutti, P.G.; de Souza, T.L.F. Physicochemical and structural properties of lunasin revealed by spectroscopic, chromatographic and molecular dynamics approaches. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140440. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P. The interplay between structure and function in intrinsically unstructured proteins. FEBS Lett. 2005, 579, 3346–3354. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef]
- Anuhadra, C.D.; Kanno, S.; Hirano, S. RGD peptide-induced apoptosis in human leukemia HL-60 cells required caspase-3 activation. Cell Biol. Toxic. 2000, 16, 275–283. [Google Scholar]
- Dia, V.P.; Gonzalez de Mejia, E. Lunasin induces apoptosis and modifies the expression of genes associated with extracellular matrix and cell adhesion in human metastatic colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef]
- De Mejia, E.G.; Wang, W.; Dia, V.P. Lunasin, with an arginine-glycine-aspartic acid motif, causes apoptosis to L1210 leukemia cells by activation of caspase-3. Mol. Nutr. Food Res. 2010, 54, 406–414. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lam, Y.; de Lumen, B.O. Barley Lunasin Suppresses ras-Induced Colony Formation and Inhibits Core Histone Acetylation in Mammalian Cells. J. Agric. Food Chem. 2002, 50, 5903–5908. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; Park, J.H.; Lee, J.B.; Kweon, D.H.; Chung, G.Y.; Seo, E.W.; de Lumen, B.O. The cancer preventive peptide lunasin from wheat inhibits core histone acetylation. Cancer Lett. 2007, 255, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Lee, J.R.; Jeong, J.B.; Park, J.H.; Cheong, Y.-K.; de Lumen, B.O. The Cancer Preventive Seed Peptide Lunasin From Rye Is Bioavailable and Bioactive. Nutr. Cancer 2009, 61, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Alaswad, A.A.; Krishnan, H.B. Immunological Investigation for the Presence of Lunasin, a Chemopreventive Soybean Peptide, in the Seeds of Diverse Plants. J. Agric. Food Chem. 2016, 64, 2901–2909. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Huerta, E.; Fernández-Tomé, S.; Arques, M.C.; Amigo, L.; Recio, I.; Clemente, A.; Hernández-Ledesma, B. The protective role of the Bowman-Birk protease inhibitor in soybean lunasin digestion: The effect of released peptides on colon cancer growth. Food Funct. 2015, 6, 2626–2635. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H. Prevention of cancer and inflammation by soybean protease inhibitors. Front. Biosci. 2013, 5, 966–973. [Google Scholar] [CrossRef]
- Gitlin-Domagalska, A.; Maciejewska, A.; Dębowski, D. Bowman-Birk Inhibitors: Insights into Family of Multifunctional Proteins and Peptides with Potential Therapeutical Applications. Pharmaceuticals 2020, 13, 421. [Google Scholar] [CrossRef]
- Dia, V.P.; Torres, S.; De Lumen, B.O.; Erdman, J.W., Jr.; De Mejia, E.G. Presence of Lunasin in Plasma of Men after Soy Protein Consumption. J. Agric. Food Chem. 2009, 57, 1260–1266. [Google Scholar] [CrossRef]
- Lam, Y.; Galvez, A.; de Lumen, B.O. Lunasin suppresses E1A-mediated transformation of mammalian cells but does not inhibit growth of immortalized and established cancer cell lines. Nutr. Cancer 2003, 47, 88–94. [Google Scholar] [CrossRef]
- Gu, L.; Ye, P.; Li, H.; Wang, Y.; Xu, Y.; Tian, Q.; Lei, G.; Zhao, C.; Gao, Z.; Zhao, W.; et al. Lunasin attenuates oxidant-induced endothelial injury and inhibits atherosclerotic plaque progression in ApoE−/−mice by up-regulating heme oxygenase-1viaPI3K/Akt/Nrf2/ARE pathway. FASEB J. 2019, 33, 4836–4850. [Google Scholar] [CrossRef]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D.; Li, H. Lunasin suppresses the migration and invasion of breast cancer cells by inhibiting matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-κB signaling pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Wu, C.-H.; Peng, S.-H.; Chang, C.-H. Seed-derived peptide lunasin suppressed breast cancer cell growth by regulating inflammatory mediators, aromatase, and estrogen receptors. Food Nutr. Res. 2023, 67, 8991. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.J.; Devapatla, B.; Yaddanapudi, K.; Davis, K.R. The soybean-derived peptide lunasin inhibits non-small cell lung cancer cell proliferation by suppressing phosphorylation of the retinoblastoma protein. Oncotarget 2015, 6, 4649–4662. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin promotes apoptosis in human colon cancer cells by mitochondrial pathway activation and induction of nuclear clusterin expression. Cancer Lett. 2010, 295, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Hernández-Ledesma, B.; De Lumen, B.O. Lunasin–Aspirin Combination Against NIH/3T3 Cells Transformation Induced by Chemical Carcinogens. Plant Foods Hum. Nutr. 2011, 66, 107–113. [Google Scholar] [CrossRef]
- De Mejia, E.G.; Dia, V.P. Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides 2009, 30, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Chou, M.-J.; Wang, C.-H. Lunasin attenuates obesity-related inflammation in RAW264.7 cells and 3T3-L1 adipocytes by inhibiting inflammatory cytokine production. PLoS ONE 2017, 12, e0171969. [Google Scholar] [CrossRef]
- Jeong, H.J.; Park, J.H.; Lam, Y.; de Lumen, B.O. Characterization of lunasin isolated from soybean. J. Agric. Food Chem. 2003, 51, 7901–7906. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin, a novel seed peptide, sensitizes human breast cancer MDA-MB-231 cells to aspirin-arrested cell cycle and induced apoptosis. Chem. Biol. Interact. 2010, 186, 127–134. [Google Scholar] [CrossRef]
- Hashimoto, H.; Vertino, P.M.; Cheng, X.; Migliori, V.; Phalke, S.; Bezzi, M.; Guccione, E.; Wei, Y.; Gañán-Gómez, I.; Salazar-Dimicoli, S.; et al. Molecular coupling of DNA methylation and histone methylation. Epigenomics 2010, 2, 657–669. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Relationship between lunasin’s sequence and its inhibitory activity of histones H3 and H4 acetylation. Mol. Nutr. Food Res. 2011, 55, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; de Lumen, B.O. Inhibition of Core Histone Acetylation by the Cancer Preventive Peptide Lunasin. J. Agric. Food Chem. 2007, 55, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Vuyyuri, S.B.; Shidal, C.; Davis, K.R. Development of the plant-derived peptide lunasin as an anticancer agent. Curr. Opin. Pharmacol. 2018, 41, 27–33. [Google Scholar] [CrossRef]
- Shidal, C.; Al-Rayyan, N.; Yaddanapudi, K.; Davis, K.R. Lunasin is a novel therapeutic agent for targeting melanoma cancer stem cells. Oncotarget 2016, 7, 84128–84141. [Google Scholar] [CrossRef] [PubMed]
- Devapatla, B.; Shidal, C.; Yaddanapudi, K.; Davis, K.R. Validation of syngeneic mouse models of melanoma and non-small cell lung cancer for investigating the anticancer effects of the soy-derived peptide Lunasin. F1000Research 2016, 5, 2432. [Google Scholar] [CrossRef]
- Shidal, C.; Inaba, J.-I.; Yaddanapudi, K.; Davis, K.R. The soy-derived peptide Lunasin inhibits invasive potential of melanoma initiating cells. Oncotarget 2017, 8, 25525–25541. [Google Scholar] [CrossRef]
- Inaba, J.; McConnell, E.J.; Davis, K.R. Lunasin Sensitivity in Non-Small Cell Lung Cancer Cells Is Linked to Suppression of Integrin Signaling and Changes in Histone Acetylation. Int. J. Mol. Sci. 2014, 15, 23705–23724. [Google Scholar] [CrossRef]
- Taylor, G.C.; Eskeland, R.; Hekimoglu-Balkan, B.; Pradeepa, M.M.; Bickmore, W.A. H4K16 acetylation marks active genes and enhancers of embryonic stem cells, but does not alter chromatin compaction. Genome Res. 2013, 23, 2053–2065. [Google Scholar] [CrossRef]
- Moreno-Layseca, P.; Streuli, C.H. Signalling pathways linking integrins with cell cycle progression. Matrix Biol. 2014, 34, 144–153. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, G.E. Lunasin potentiates the effect of oxaliplatin preventing outgrowth of colon cancer metastasis, binds to α5β1 integrin and suppresses FAK/ERK/NF-κB signaling. Cancer Lett. 2011, 313, 167–180. [Google Scholar] [CrossRef]
- Cam, A.; de Mejia, E.G. RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Mol. Nutr. Food Res. 2012, 56, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, B.; Rudnicka, K.; Bednarek, A.; Fabianowska-Majewska, K. Hypomethylation and induction of retinoic acid receptor beta 2 by concurrent action of adenosine analogues and natural compounds in breast cancer cells. Eur. J. Pharmacol. 2010, 638, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Stefanska, B.; Salamé, P.; Bednarek, A.; Fabianowska-Majewska, K. Comparative effects of retinoic acid, vitamin D and resveratrol alone and in combination with adenosine analogues on methylation and expression of phosphatase and tensin homologue tumour suppressor gene in breast cancer cells. Br. J. Nutr. 2012, 107, 781–790. [Google Scholar] [CrossRef]
- Kaufman-Szymczyk, A.; Majda, K.; Szuławska-Mroczek, A.; Fabianowska-Majewska, K.; Lubecka, K. Clofarabine-phytochemical combination exposures in CML cells inhibit DNA methylation machinery, upregulate tumor suppressor genes and promote caspase-dependent apoptosis. Mol. Med. Rep. 2019, 20, 3597–3608. [Google Scholar] [CrossRef]
- Chuang, L.S.; Ian, H.I.; Koh, T.W.; Ng, H.H.; Xu, G.; Li, B.F. Human DNA-(cytosine-5)-methyl-transferase-PCNA complex as a target for p21WAF1. Science 1997, 277, 1996–2000. [Google Scholar] [CrossRef]
- Iida, T.; Suetake, I.; Tajima, S.; Morioka, H.; Ohta, S.; Obuse, C.; Tsurimoto, T. PCNA clamp facilitates action of DNA cytosine methyltransferase 1 on hemimethylated DNA. Genes Cells 2002, 7, 997–1007. [Google Scholar] [CrossRef]
- Delavaine, L.; La Thangue, N. Control of E2F activity by p21Waf1/Cip1. Oncogene 1999, 18, 5381–5392. [Google Scholar] [CrossRef]
- Cantley, L.C.; Neel, B.G. New insights into tumor suppression: PTEN suppresses tumor formation by restraining the phosphoinositide 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 4240–4245. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Tamura, M.; Yamada, K.M. Tumor suppressor PTEN inhibits integrin- and growth factor-mediated mitogen-activated protein (MAP) kinase signaling pathways. J. Cell Biol. 1998, 143, 1375–1383. [Google Scholar] [CrossRef]
- Bigey, P.; Ramchandani, S.; Theberge, J.; Araujo, F.D.; Szyf, M. Transcriptional regulation of the human DNA Methyltransferase (DNMT1) gene. Gene 2000, 242, 407–418. [Google Scholar] [CrossRef]
- Qin, W.; Leonhardt, H.; Pichler, G. Regulation of DNA methyltransferase 1 by interactions and modifications. Nucleus 2011, 2, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Pabona, J.M.; Dave, B.; Su, Y.; Montales, M.T.; de Lumen, B.O.; de Mejia, E.G.; Rahal, O.M.; Simmen, R.C. The soybean peptide lunasin promotes apoptosis of mammary epithelial cells via induction of tumor suppressor PTEN: Similarities and distinct actions from soy isoflavone genistein. Genes Nutr. 2013, 8, 79–90. [Google Scholar] [CrossRef]
- Montales, M.T.; Simmen, R.C.; Ferreira, E.S.; Neves, V.A.; Simmen, F.A. Metformin and soybean-derived bioactive molecules attenuate the expansion of stem cell-like epithelial subpopulation and confer apoptotic sensitivity in human colon cancer cells. Genes Nutr. 2015, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; de Mejia, E.G. Differential gene expression of RAW 264.7 macrophages in response to the RGD peptide lunasin with and without lipopolysaccharide stimulation. Peptides 2011, 32, 1979–1988. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Xu, F.; Han, Y.; Hernández-Ledesma, B.; Xiao, H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation. Int. J. Mol. Sci. 2020, 21, E537. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez de Mejia, E.; Castañeda-Reyes, E.D.; Mojica, L.; Dia, V.; Wang, H.; Wang, T.; Johnson, L.A. Potential Health Benefits Associated with Lunasin Concentration in Dietary Supplements and Lunasin-Enriched Soy Extract. Nutrients 2021, 13, 1618. [Google Scholar] [CrossRef]
- Setrerrahmane, S.; Zhang, Y.; Dai, G.; Lv, J.; Tan, S. Efficient production of native lunasin with correct N-terminal processing by using the pH-induced self-cleavable Ssp DnaB mini-intein system in Escherichia coli. Appl. Biochem. Biotechnol. 2014, 174, 612–622. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Sanchón, J.; Recio, I.; Hernández-Ledesma, B. Transepithelial transport of lunasin and derived peptides: Inhibitory effects on the gastrointestinal cancer cells viability. J. Food Compos. Anal. 2018, 68, 101–110. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Ramos, S.; Cordero-Herrera, I.; Recio, I.; Goya, L.; Hernández-Ledesma, B. In vitro chemo-protective effect of bioactive peptide lunasin against oxidative stress in human HepG2 cells. Food Res. Int. 2014, 62, 793–800. [Google Scholar] [CrossRef]
- García-Nebot, M.J.; Recio, I.; Hernández-Ledesma, B. Antioxidant activity and protective effects of peptide lunasin against oxidative stress in intestinal Caco-2 cells. Food Chem. Toxicol. 2014, 65, 155–161. [Google Scholar] [CrossRef]
- Wang, W.; Bringe, N.A.; Berhow, M.A.; Gonzalez de Mejia, E. β-Conglycinins among Sources of Bioactives in Hydrolysates of Different Soybean Varieties That Inhibit Leukemia Cells in Vitro. J. Agric. Food Chem. 2008, 56, 4012–4020. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Hernández-Ledesma, B.; Jeong, H.J.; Park, J.H.; de Lumen, B.O. Complementary roles in cancer prevention: Protease inhibitor makes the cancer preventive peptide lunasin bioavailable. PLoS ONE 2010, 5, e8890. [Google Scholar] [CrossRef] [PubMed]
- Rusdi, N.K.; Purwaningsih, E.H.; Hestiantoro, A.; Elya, B.; Kusmardi, K. In Vivo Antimammary Tumor Effects of Soybean Extract with Targeted Lunasin (ET-Lun). Pharm. J. 2021, 13, 1269–1276. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Potential of lunasin orally-administered in comparison to intraperitoneal injection to inhibit colon cancer metastasis in vivo. J. Cancer Ther. 2013, 4, 34–43. [Google Scholar] [CrossRef]


| Plant | Content of Lunasin (mg per g of Seeds) |
|---|---|
| Soybean | 0.5–8.1 |
| Barley | 0.01–0.02 |
| Wheat | 0.2–0.3 |
| Rye | 0.045–0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaufman-Szymczyk, A.; Kaczmarek, W.; Fabianowska-Majewska, K.; Lubecka-Gajewska, K. Lunasin and Its Epigenetic Impact in Cancer Chemoprevention. Int. J. Mol. Sci. 2023, 24, 9187. https://doi.org/10.3390/ijms24119187
Kaufman-Szymczyk A, Kaczmarek W, Fabianowska-Majewska K, Lubecka-Gajewska K. Lunasin and Its Epigenetic Impact in Cancer Chemoprevention. International Journal of Molecular Sciences. 2023; 24(11):9187. https://doi.org/10.3390/ijms24119187
Chicago/Turabian StyleKaufman-Szymczyk, Agnieszka, Wiktoria Kaczmarek, Krystyna Fabianowska-Majewska, and Katarzyna Lubecka-Gajewska. 2023. "Lunasin and Its Epigenetic Impact in Cancer Chemoprevention" International Journal of Molecular Sciences 24, no. 11: 9187. https://doi.org/10.3390/ijms24119187