Abstract
Waning vaccine-induced immunity, coupled with the emergence of SARS-CoV-2 variants, has inspired the widespread implementation of COVID-19 booster vaccinations. Here, we evaluated the potential of the GX-19N DNA vaccine as a heterologous booster to enhance the protective immune response to SARS-CoV-2 in mice primed with either an inactivated virus particle (VP) or an mRNA vaccine. We found that in the VP-primed condition, GX-19N enhanced the response of both vaccine-specific antibodies and cross-reactive T Cells to the SARS-CoV-2 variant of concern (VOC), compared to the homologous VP vaccine prime-boost. Under the mRNA-primed condition, GX-19N induced higher vaccine-induced T Cell responses but lower antibody responses than the homologous mRNA vaccine prime-boost. Furthermore, the heterologous GX-19N boost induced higher S-specific polyfunctional CD4+ and CD8+ T cell responses than the homologous VP or mRNA prime-boost vaccinations. Our results provide new insights into booster vaccination strategies for the management of novel COVID-19 variants.
1. Introduction
Unprecedented rates of vaccine development have occurred in response to the COVID-19 pandemic. As a result, 20 different types of COVID-19 vaccines have been approved for use, with 65.1% and 58.9% of the world’s population receiving at least one dose and full vaccination, respectively [1]. Approximately 4.6 and 3.2 billion doses of virus particle (VP) and mRNA vaccines were delivered globally, respectively [2].
Considering the increasing prevalence of COVID-19 and the duration of vaccine efficacy, many countries are implementing COVID-19 vaccine-booster programs. Although boosters have increased the vaccine-induced immune response (i.e., neutralizing the antibody response), the neutralizing antibody titers against the emerging variants were significantly lower than those against the SARS-CoV-2 wild-type [3,4,5,6]. Because the initial boosters were based on the SARS-CoV-2 wild-type sequence, their efficacy in inducing a potent neutralizing antibody response to mutated sequences is limited. Therefore, an additional booster shot is currently required approximately 6 months after the completion of a vaccination. To overcome the limitations of SARS-CoV-2 wild-type-based vaccine booster shots, variant-specific vaccines are continuously being developed [5,7]. Nevertheless, the rationale for the SARS-CoV-2 wild-type-based COVID-19 vaccine booster strategy is to prevent hospitalization due to infection rather than preventing infection itself, and the T Cell response, rather than the neutralizing the antibody response, is considered to be the main factor preventing hospitalization. Unlike neutralizing antibody responses, T cell recognition appears to be broadly cross-reactive against variants of concern (VOCs) [8].
Several vaccine platforms have been developed for COVID-19 vaccines, and clinical results have shown that different types of immune responses are induced by each. The COVID-19 protein subunit vaccine induces an antibody- rather than a T cell-associated immune response [9,10], the COVID-19 VP vaccine induces an antibody-associated immune response [11,12], and the COVID-19 viral vector vaccine induces both antibody- and T cell-associated immune responses [13,14]. Similarly, COVID-19 mRNA vaccines effectively induce antibody and T cell responses [15,16]. GX-19N, which is being developed as a COVID-19 DNA vaccine, effectively induces a T cell response with a marginal antibody response [17].
In this study, we evaluated the performance of GX-19N DNA vaccine booster-shot regimens in mice primed with either the COVID-19 mRNA or VP vaccine. We demonstrated for the first time that the heterologous GX-19N DNA-boosting vaccination induced a much stronger T cell response than either homologous mRNA or VP prime-boost vaccinations. Furthermore, we demonstrated that polyfunctional CD4+ and CD8+ T cell responses were also increased by the heterologous GX-19N DNA-boosting vaccination.
2. Result
2.1. Heterologous GX-19N DNA Boosting Vaccination Induced a Higher and Lower Antibody Response Than Homologous VP and mRNA Prime-Boost, Respectively
To investigate the potential of a heterologous GX-19N DNA booster vaccination, SARS-CoV-2 mRNA- or VP-primed BALB/c mice were vaccinated with either a homologous- or heterologous-primed GX-19N DNA booster (Figure 1A). The VP vaccine primed-GX-19N induced a much higher SRBD-specific antibody response than the homologous VP vaccine prime-boost regimen did, whereas the homologous mRNA vaccine prime-boost regimen induced a significantly higher SRBD-specific antibody response than the mRNA vaccine primed-GX-19N did (Figure 1B,C). The ratio of IgG2a to IgG1 antibody titer tended to increase after the GX-19N vaccination, indicating an enhancement of Th1-polarized immunity, consistent with previous reports (Figure 1D,E) [18,19].
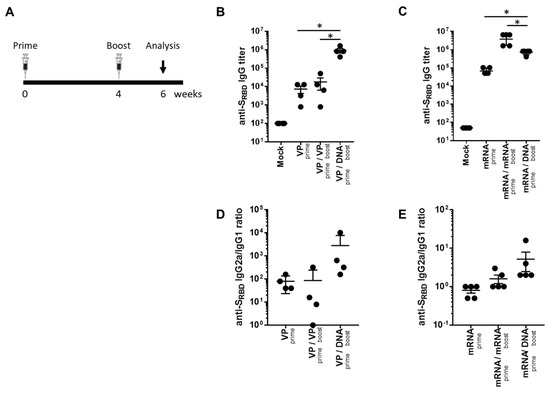
Figure 1.
Humoral response to SARS-CoV-2 SRBD antigen after homologous and heterologous prime-boost vaccination. (A) BALB/c mice (n = 4–5/group) were immunized at weeks 0 and 4; serum antibody responses were measured 2 weeks after the last immunization. Graphs show (B,C) SARS-CoV-2 SRBD-specific IgG titers and (D,E) ratios of SRBD-specific IgG2a to IgG1 titers. Individual mice are represented by a single data point. p-values were determined using a two-tailed Student’s t-test. * p < 0.05. DNA, GX-19N.
In addition, we evaluated neutralizing antibody responses to Wuhan and VOCs (B.1.351 and B.1.617.2), using the surrogate virus neutralization test (sVNT), which is highly correlated with the conventional VNT (cVNT) and pseudovirus-based VNT (pVNT) [20]. In the case of VP vaccine priming, the GX-19N induced a significantly higher neutralizing antibody titer than the homologous VP-boosting vaccination. The neutralizing antibody titer obtained through the heterogeneous regimen was 76.1-fold higher for the Wuhan (mean = 14 vs. 1076 sVNT20 titer), 53.8-fold higher for B.1.351 (mean = 10 vs. 538 sVNT20 titer), and 76.1-fold higher for B.1.617.2 (mean = 14 vs. 1076 sVNT20 titer) variants than that obtained with the homologous prime-boost regimen (Figure 2A–C). In the case of mRNA vaccine priming, the homologous mRNA-boosting vaccination induced significantly higher neutralizing antibody titers than the GX-19N vaccination. The neutralizing antibody titer obtained through the homologous prime-boost regimen was 2.2-fold higher for the Wuhan (mean = 1280 vs. 576 sVNT20 titer), 2.5-fold higher for B.1.351 (mean = 1280 vs. 512 sVNT20 titer), and 3.2-fold higher for B.1.617.2 (mean = 2688 vs. 848 sVNT20 titer) variants than that obtained with the heterologous prime-boost regimen (Figure 2D–F).
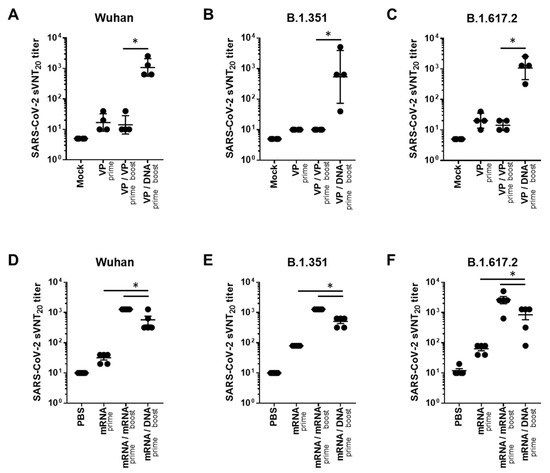
Figure 2.
Neutralizing antibody responses against SARS-CoV-2 variants after homologous and heterologous prime-boost vaccination. BALB/c mice (n = 4–5/group) were immunized at weeks 0 and 4; serum antibody responses were measured 2 weeks after the last immunization. Sera from vaccinated mice were tested for (A,D) sVNT20 titers against Wuhan, (B,E) B.1.351, (C,F) and B.1.617.2. Individual mice are represented by a single data point. p-values were determined using a two-tailed Student’s t-test. * p < 0.05. DNA, GX-19N.
Taken together, these data indicated that the heterologous GX-19N DNA booster vaccination showed different outcomes depending on the initial priming vaccine. When VP vaccine priming was used, the GX-19N booster induced a significantly higher SRBD-specific antibody response and neutralizing antibody titer compared to the homologous VP prime-boost regimen, while the homologous mRNA prime-boost regimen produced higher antibody responses than the GX-19N boost vaccination.
2.2. Heterologous GX-19N DNA Boosting Vaccination Induces a Higher Cross-Reactive T Cell Response against SARS-CoV-2 VOCs Than the Homologous VP or mRNA Prime-Boost Regimen
T cell responses were evaluated after homologous- or heterologous-primed GX-19N DNA boosting regimens in SARS-CoV-2 mRNA- or VP-primed BALB/c mice. Compared with the homologous VP prime-boost regimen, the GX-19N vaccination increased the T cell response by 21.1-fold (Figure 3A). As expected, we observed similar levels of cellular responses to B.1.351 (mean = 1644 SFUs/106 splenocytes) and B.1.617.2 (mean = 1753 SFUs/106 splenocytes) spike peptides (Figure 3B,D). This is consistent with previous findings of cellular immunity being relatively unimpaired by VOCs compared with neutralizing antibody responses [21]. Similar to the results for VP vaccine-primed mice, the GX-19N increased T cell responses by 2.3-fold compared to the homologous mRNA prime-boost vaccination (Figure 3C).
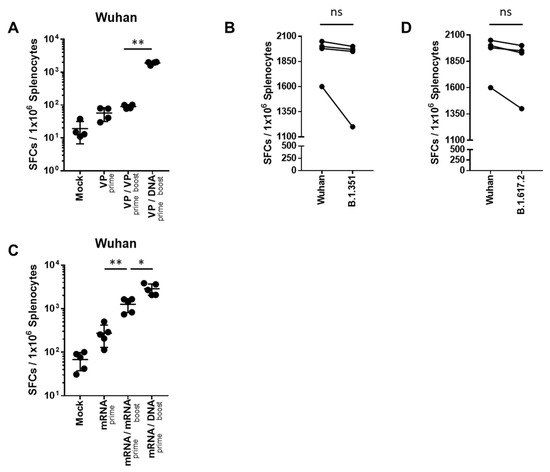
Figure 3.
SARS-CoV-2-spike-specific T cell response against SARS-CoV-2 variants after homologous and heterologous prime-boost vaccination. BALB/c mice (n = 4–5/group) were immunized at weeks 0 and 4; T Cell response was measured by IFN-γ ELIspot in splenocytes stimulated with peptide pools spanning the SARS-CoV-2 spike protein of the Wuhan (A,C). T Cell response was measured by IFN-γ ELispot in splenocytes from heterologous VP prime-DNA boost mice stimulated with peptide pools spanning the spike proteins of (B) B.1.351 and (D) B.1.6172. Individual mice are represented by a single data point. Shown are the average spot-forming cells (SFCs) per 106 splenocytes in triplicate wells against peptide pools, after subtracting the background number of spots. p-values were determined using a two-tailed Student’s t-test. ns—not significant; * p < 0.05; ** p < 0.01. DNA, GX-19N.
In summary, the heterologous GX-19N DNA booster vaccination significantly enhanced T cell responses compared to both homologous VP and mRNA prime-boost regimens. Moreover, comparable levels of T Cell responses to Wuhan and VOCs were observed, indicating that cellular immunity is relatively unaffected by the variant spike proteins compared to neutralizing antibody responses.
2.3. Heterologous GX-19N DNA Boosting Vaccination Induces a Superior Polyfunctional T Cell Response Than the Homologous mRNA Prime-Boost
The quality of the S-specific T cell responses was characterized by analyzing the pattern of cytokine production (i.e., IFN-γ, TNF-α, and/or IL-2) (Figure 4A and Figure 5A). In the S-specific CD4+ T cell response, higher IFN-γ+TNF-α+ or IFN-γ+TNF-α+IL-2+ polyfunctional T cells were induced by the GX-19N than by homologous mRNA prime-boost regimens. The proportion of polyfunctional T cells among cytokine-producing cells was also increased by the GX-19N regimen (mRNA/mRNA, 65% vs. mRNA/GX-19N DNA, 87%) (Figure 4B). An S-specific multifunctional CD8+ T cell response was also generated. Similar to CD4+ T cell responses, the proportion of multifunctional CD8+ T cells among cytokine-producing cells was increased by the GX-19N regimen (mRNA/mRNA, 48% vs. mRNA/GX-19N DNA, 80%) (Figure 5B).
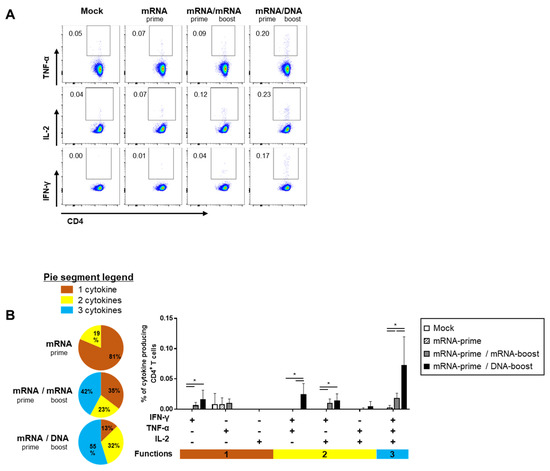
Figure 4.
Polyfunctionality of vaccine-induced CD4+ T cell response after homologous and heterologous prime-boost vaccination. BALB/c mice (n = 4/group) were immunized at weeks 0 and 4; CD4+ T Cell response was measured by ICS in splenocytes stimulated with peptide pools spanning the SARS-CoV-2 spike protein. (A) Vaccine-induced CD4+ T cell response. (B) Polyfunctionality of vaccine-induced CD4+ T cell responses based on every possible combination of functions. Pie graph sections represent the fraction of T Cells positive for a given number of functions. The three horizontal bars of different colors below x axis depict the population of three, two, or one functional responses. p-values were determined using a two-tailed Student’s t-test. * p < 0.05. DNA, GX-19N.
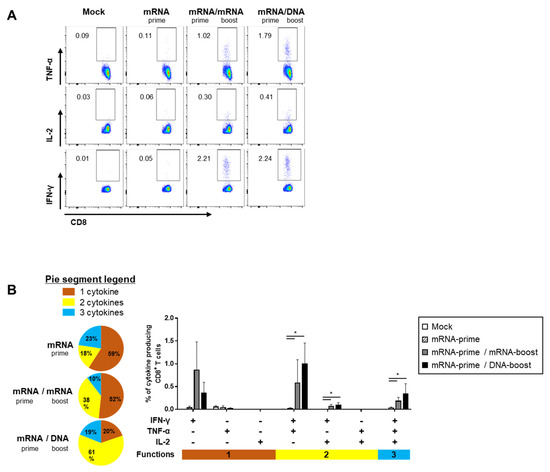
Figure 5.
Polyfunctionality of vaccine-induced CD8+ T cell response after homologous and heterologous prime-boost vaccination. BALB/c mice (n = 4/group) were immunized at weeks 0 and 4; CD8+ T Cell response was measured by ICS in splenocytes stimulated with peptide pools spanning the SARS-CoV-2 spike protein. (A) Vaccine-induced CD8+ T cell response. (B) Polyfunctionality of vaccine-induced CD8+ T cell responses based on every possible combination of functions. Pie graph sections represent the fraction of T Cells positive for a given number of functions. The three horizontal bars of different colors below x axis depict the population of three, two, or one functional responses. p-values were determined using a two-tailed Student’s t-test. * p < 0.05. DNA, GX-19N.
In summary, the heterologous GX-19N boost regimen induced higher levels of polyfunctional T cells in both S-specific CD4+ and CD8+ T cell responses, indicating the superior quality of T Cell responses elicited by the heterologous GX-19N boost vaccination.
3. Discussion
The rapid decline of vaccine-induced immunity against SARS-CoV-2, especially that of humoral immunity against its variants, has led to an increase in infection rates, which has prompted the development of vaccine booster doses. As a result of many clinical studies on homologous or heterologous booster doses using current COVID-19 vaccines, the use of booster shots for COVID-19 mRNA has been approved [3,6,22]. The COVID-19 booster vaccine has the advantage of providing substantially increased protection against severe infections and has reduced the rates of hospitalization and death globally [23]. However, the efficacy of the COVID-19 booster dose against SARS-CoV-2 infection remains low (30% or less) [4]. T cells are expected to be particularly important in enhancing protection against severe COVID-19 infections after booster doses. Mounting evidence suggests that T Cell contributions to the host immune response are required for early, broad, and durable protection from SARS-CoV-2, especially with regard to new VOCs [24,25,26,27,28,29]. However, taking two or more booster shots with COVID-19 mRNA vaccines every few months may pose potential risks, including anaphylaxis. Reactions with anaphylactic features following administration of both COVID-19 mRNA vaccines have been reported in the United Kingdom, United States, Japan, and elsewhere [30,31,32,33]. The incidence of these reactions may be higher with these than with other vaccines, such as protein subunit or VP vaccines. Polyethylene glycol (PEG), one of the lipid components of the COVID-19 mRNA vaccine, is hypothesized to be the cause of IgE-mediated anaphylactic reactions to medications, bowel preps, or laxatives containing PEG [34,35]. In addition, it is presumed that the cause of anaphylaxis is a complement activation-related pseudoallergy (CARPA), in which the pre-existing IgG or IgM antibody to PEG activates complementarily, generating anaphylatoxins (C3a, C4a, and C5a), and causing mast cell degranulation [36]. There is also an increased risk of myocarditis. Based on reports of passive surveillance in the United States, the risk of myocarditis after vaccination with COVID-19 mRNA increased across multiple age and sex strata, and was more prevalent after the second dose of the vaccination [37]. This suggests that repeated booster doses of the COVID-19 mRNA vaccine increase the risk of developing myocarditis. Finally, there is an increased likelihood of antibody-dependent enhancement (ADE). Although no serious ADE concerns have arisen with the COVID-19 vaccine, studies on COVID-19 patients have reported a high potential for ADE [38,39]. Repeated booster shots of the SARS-CoV-2 wild-type sequence-based COVID-19 mRNA vaccine, which can induce strong humoral and T cell responses, can increase non-neutralizing antibody responses to SARS-CoV-2 variants. Given that ADE are mediated by non-neutralizing antibody responses, the likelihood of developing ADEs may gradually increase. Taken together, these findings suggest that an ideal COVID-19 vaccine booster should be safe and T cell-oriented. In this study, we investigated the performance of the GX-19N DNA vaccine, which demonstrated an excellent safety profile and T cell-biased immune response in a previous clinical study [17], as a booster for mRNA or VP vaccines.
Interestingly, the heterologous GX-19N DNA booster vaccination significantly increased the vaccine-induced T cell response, more so than the homologous mRNA booster. In contrast, the induction of an antibody response was lower in the GX-19N than in the homologous mRNA booster regimen. This may be due to the different responses between Th1 (type 1 T helper) and Th2 among the different vaccines used in this study. It is known that the Th1 response induces a cell-mediated response, whereas the Th2 response is related to the humoral immune response [40]. COVID-19 mRNA vaccines can induce an immune response that is unbiased for Th1 or Th2 responses in mice [41,42]. In contrast, the GX-19N DNA vaccine induced a Th1-predominant response in mice [43]. Considering that Th1 and Th2 responses are distinctly related to cellular and humoral immune responses, the immune response induced by the mRNA vaccine prime vaccination would be biased toward the Th1 response induced by the GX-19N boost vaccination. As a result, the heterologous GX-19N DNA booster induced a high T cell response but a low antibody response compared to the mRNA booster vaccination. The higher ratio of IgG2a to IgG1 antibody titers observed using the GX-19N booster, compared to the homologous mRNA prime-boost, support the above explanation.
It is important to acknowledge the limitations of the study. The first limitation is the absence of a DNA-vaccine-only control. Despite the fact that most of the population has received at least one dose of the VP vaccine or mRNA vaccines, this still limits the applicability of the analysis. Additionally, we did not perform an analysis of an mRNA vaccine prime-VP vaccine boost or vice versa. It is expected that the homologous prime-boost effect would be weak, but it is unknown whether the effect of a heterologous prime-boost using an mRNA vaccine or VP vaccine is the same as that of a DNA vaccine boost. Further studies are needed for a comprehensive comparative evaluation. The second limitation of the study is that it only evaluated antibody responses and T Cell responses in the blood as indicators of vaccine efficacy. While many other studies also utilize adaptive immune responses in the blood as indicators of vaccine efficacy, the actual contribution of the protective effectiveness of the vaccine varies among studies [44,45]. Moreover, it is believed that the adaptive immune response has limited access to the respiratory tract, due to the tolerance of many antigenic interactions in the respiratory tract. Therefore, the antibody and T cell responses measured in the blood may not predict the protective efficacy of vaccines against respiratory tract infections. Further studies evaluating adaptive immune responses in the respiratory tract, or studies on the protective efficacy using SARS-CoV-2 infection model are needed.
4. Methods and Materials
4.1. Vaccines
The COVID-19 GX-19N DNA vaccine, consisting of GX-19 and GX-21 at a ratio of 1:2, was constructed by inserting the antigen genes of SARS-CoV-2 into a pGX27 vector [43]. GX-19 (pGX27-SΔTM/IC) contains the SARS-CoV-2 spike (S) gene lacking the transmembrane (TM)/intracellular (IC) domain, and GX-21 (pGX27-SRBD-F/NP) is designed to express the fusion protein of the receptor-binding domain (RBD) of the spike protein, the T4 fibritin C-terminal Foldon (SRBD-Foldon), and the nucleocapsid protein (N). S, SRBD-Foldon, and N are preceded by the secretory signal sequence of tissue plasminogen activation (tPA). The inactivated SARS-CoV-2 vaccine produced from Vero cells contained 4 μg of viral antigens and 0.225 mg of aluminum hydroxide adjuvant in a 0.5-mL dose. The mRNA vaccine was encapsulated in a lipid nanoparticle through a modified ethanol-drop nanoprecipitation process as previously described. Briefly, ionizable, structural, helper, and PEG lipids were mixed with mRNA in acetate buffer at a ratio of 2.5:1 (lipids:mRNA). The mixture was neutralized with Tris-Cl, sucrose was added as a cryoprotectant, and the final solution was sterile filtered and stored frozen at −70 °C until further use [42].
4.2. Mouse Immunizations
Female BALB/c mice aged 6–8 weeks (Central Lab Animal) were intramuscularly immunized with 0.4 μg/animal VP vaccine (total volume of 50 μL) or 1 ug/animal mRNA (total volume of 50 uL) at week 0. At week 4, the same mice were injected with homologous vaccine booster or 12 μg/animal GX-19N booster (total volume of 50 μL, adjusted with PBS) into the tibialis anterior muscle with in vivo electroporation using an OrbiJector® system (SL VaxiGen Inc., Seongnam, Republic of Korea). Mice were sacrificed two weeks after the final immunization.
4.3. Antigen Binding ELISA
The serum collected at each time point was evaluated for binding titers. In this assay, 96-well ELISA plates (NUNC) were coated with 1 μg/mL recombinant SARS-CoV-2 spike RBD-His protein (Sino Biological, 40592-V08B, Beijing, China) in PBS, overnight at 4 °C. The plates were washed three times with 0.05% PBST (Tween 20 in PBS) and blocked with 5% skim milk in 0.05% PBST (SM) for 2–3 h at room temperature. The sera were serially diluted in 5% SM, added to the wells, and incubated for 2 h at 37 °C. Following incubation, the plates were washed five times with 0.05% PBST and then incubated with horseradish peroxidase (HRP)-conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories, 115-035-003, West Grove, PA, USA), IgG1 (Jackson ImmunoResearch Laboratories 115-035-205), or IgG2a (Jackson ImmunoResearch Laboratories 115-035-206) for 1 h at 37 °C. After the final wash, the plates were developed using TMB solution (Surmodics, TMBW-1000-01, Eden Prairie, MN, USA), and the reaction was stopped with 2N H2SO4. The plates were analyzed at 450 nm using a SpectraMax Plus384 (Molecular Devices, San Jose, CA, USA).
4.4. Surrogate Virus-Neutralization Assay
The sVNT was used to analyze the binding ability of RBD to ACE2 after neutralizing RBD with antibodies in the serum. Serum collected two weeks after the final immunization was quantified according to the manufacturer’s instructions (Sugentech, CONE001E, Daejeon, Republic of Korea). Briefly, sera were serially diluted in dilution buffer and treated with HRP-conjugated RBD for 30 min at 37 °C. The samples were added to a plate coated with the human ACE2 protein and incubated for 15 min at 37 °C. Following incubation, the plates were washed five times with the wash solution. After the final wash, the plates were developed using TMB solution, and the reaction was arrested with a stop solution. The plates were analyzed at 450 nm using the SpectraMax Plus384 (Molecular Devices). The reciprocal of the dilution that resulted in a binding inhibition rate of 20% or more (PI20) was defined as the neutralizing antibody titer.
4.5. IFN-γ ELISPOT
A mouse IFN-γ ELISPOT set (BD 551083) was used as directed by the manufacturer. The ELISPOT plates were coated with purified anti-mouse IFN-γ capture antibody and incubated overnight at 4 °C. Plates were washed and blocked for 2 h with RPMI + 10% FBS (R10 medium), and 5 × 105 splenocytes were added to each well and stimulated for 24 h at 37 °C in 5% CO2 with R10 medium (negative control), concanavalin A (positive control), or specific peptide pools (2 μg/mL). Peptide pools consisted of 15-mer peptides overlapped by 11 amino acids and spanning the entire S proteins of SARS-CoV-2 (GenScript, Nanjing, China). After stimulation, the plates were washed and spots were developed according to the manufacturer’s instructions. The plates were scanned and counted using an AID ELISPOT reader. Spot-forming units (SFU) per million cells were calculated by subtracting the number of negative control wells.
4.6. Intracellular Cytokine Staining
Splenocytes were stimulated in R10 media with specific peptide pools or medium alone (DMSO control) for 12 h. After stimulation, cells were washed with PBS for subsequent immunostaining. Antibodies for staining cells were CD8 FITC (Biolegend, 100706, San Diego, CA, USA), IL-2 PE (Biolegend 503808), CD4 PE-Cy7 (Biolegend 100528), IFN-γ APC (Biolegend 505810), TNF-α (Biolegend 506328), CD3 BV605 (Biolgend 100351), and Live/dead IR (Invitrogen L10119). Fluorescence-activated cell sorting analysis was accomplished using a Fortessa flow cytometer (BD bioscience, San Jose, CA, USA), and the data were analyzed using FlowJo software (v10.8.1). Background cytokine expression in the DMSO-controls was subtracted from that measured in the S peptide pools.
4.7. Statistical Analysis
Data analyses were performed using GraphPad Prism 7 (GraphPad Software). Comparisons between groups were performed using two-tailed Student’s t-tests. Statistical significance was set at p < 0.05.
Author Contributions
Y.B.S. contributed to study design, oversaw the planning of the project, analyzed the data, and wrote the manuscript. A.K., D.S., J.K., J.N., J.I.R., S.L. and M.J.O. performed experiments and data analysis, including sample processing, ELISA, IFN-γ ELISPOT assays and the ICS assay. Y.S.S. contributed to study coordination. Y.C.S. oversaw the planning and direction of the project, including analysis and interpretation of the data and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of Genenbio (IACUC permit number GN-IACUC-22-04-09).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
Y.B.S., A.K., J.K., J.I.R., S.L. and M.J.O. are employees of SL VaxiGen Inc. D.S., Y.S.S., J.N. and Y.C.S. are employees of Genexine Inc. Y.B.S. is an inventor of multiple patent applications related to GX-19N DNA vaccine antigen constructs and vaccines. Y.C.S serves on the Scientific Advisory Board of SL VaxiGen.
Abbreviations
| ACE2 | Angiotensin-converting enzyme 2 |
| ADE | antibody-dependent enhancement |
| COVID-19 | coronavirus disease-2019 |
| cVNT | conventional virus neutralization test |
| ELISA | enzyme-linked immunosorbent assay |
| ELISpot | Enzyme-Linked ImmunoSpot |
| IC | intracellular |
| IFN-γ | interferon-γ |
| IL-2 | Interleukin-2 |
| N | nucleocapsid protein |
| pVNT | pseudovirus-based virus neutralization test |
| RBD | receptor-binding domain |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| SFCs | spot-forming cells |
| sVNT | surrogate virus neutralization test |
| TM | transmembrane |
| TNF-α | tumor necrosis factor-α |
| tPA | tissue plasminogen activation |
| VOC | variant of concern |
| VP | virus particle |
References
- Coronavirus (COVID-19) Vaccinations. Our World in Data: 2022. Available online: https://ourworldindata.org/covid-vaccinations (accessed on 13 May 2022).
- Dolgin, E. Omicron thwarts some of the world’s most-used COVID vaccines. Nature 2022, 601, 311. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; St Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. Cell 2022, 185, 457–466.e4. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Gonen, T.; Gilboa, M.; Mandelboim, M.; Indenbaum, V.; Amit, S.; Meltzer, L.; Asraf, K.; Cohen, C.; Fluss, R.; et al. Efficacy of a Fourth Dose of COVID-19 mRNA Vaccine against Omicron. N. Engl. J. Med. 2022, 386, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Pajon, R.; Doria-Rose, N.A.; Shen, X.; Schmidt, S.D.; O’Dell, S.; McDanal, C.; Feng, W.; Tong, J.; Eaton, A.; Maglinao, M.; et al. SARS-CoV-2 Omicron Variant Neutralization after mRNA-1273 Booster Vaccination. N. Engl. J. Med. 2022, 386, 1088–1091. [Google Scholar] [CrossRef]
- Um, J.; Choi, Y.Y.; Kim, G.; Kim, M.K.; Lee, K.S.; Sung, H.K.; Kim, B.C.; Lee, Y.K.; Jang, H.C.; Bang, J.H.; et al. Booster BNT162b2 COVID-19 Vaccination Increases Neutralizing Antibody Titers Against the SARS-CoV-2 Omicron Variant in Both Young and Elderly Adults. J. Korean Med. Sci. 2022, 37, e70. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.; Koch, M.; Wu, K.; Chu, L.; Ma, L.; Hill, A.; Nunna, N.; Huang, W.; Oestreicher, J.; Colpitts, T.; et al. Safety and immunogenicity of SARS-CoV-2 variant mRNA vaccine boosters in healthy adults: An interim analysis. Nat. Med. 2021, 27, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1-2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef]
- Ella, R.; Vadrevu, K.M.; Jogdand, H.; Prasad, S.; Reddy, S.; Sarangi, V.; Ganneru, B.; Sapkal, G.; Yadav, P.; Abraham, P.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: A double-blind, randomised, phase 1 trial. Lancet Infect. Dis. 2021, 21, 637–646. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 2020, 396, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): A single-blind, randomised, controlled, phase 2/3 trial. Lancet 2021, 396, 1979–1993. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021, 595, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Lee, J.; Suh, Y.S.; Song, Y.G.; Choi, Y.J.; Lee, K.H.; Seo, S.H.; Song, M.; Oh, J.W.; Kim, M.; et al. Safety and immunogenicity of two recombinant DNA COVID-19 vaccines containing the coding regions of the spike or spike and nucleocapsid proteins: An interim analysis of two open-label, non-randomised, phase 1 trials in healthy adults. Lancet Microbe 2022, 3, e173–e183. [Google Scholar] [CrossRef]
- Lim, Y.S.; Kang, B.Y.; Kim, E.J.; Kim, S.H.; Hwang, S.Y.; Kim, T.S. Potentiation of antigen-specific, Th1 immune responses by multiple DNA vaccination with an ovalbumin/interferon-gamma hybrid construct. Immunology 1998, 94, 135–141. [Google Scholar] [CrossRef]
- Song, M.K.; Lee, S.W.; Suh, Y.S.; Lee, K.J.; Sung, Y.C. Enhancement of immunoglobulin G2a and cytotoxic T-lymphocyte responses by a booster immunization with recombinant hepatitis C virus E2 protein in E2 DNA-primed mice. J. Virol. 2000, 74, 2920–2925. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Tarke, A.; Sidney, J.; Methot, N.; Yu, E.D.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; Wang, E.; et al. Impact of SARS-CoV-2 variants on the total CD4(+) and CD8(+) T cell reactivity in infected or vaccinated individuals. Cell Rep. Med. 2021, 2, 100355. [Google Scholar] [CrossRef]
- Perez-Then, E.; Lucas, C.; Monteiro, V.S.; Miric, M.; Brache, V.; Cochon, L.; Vogels, C.B.F.; Malik, A.A.; De la Cruz, E.; Jorge, A.; et al. Neutralizing antibodies against the SARS-CoV-2 Delta and Omicron variants following heterologous CoronaVac plus BNT162b2 booster vaccination. Nat. Med. 2022, 28, 481–485. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Sachdeva, R.; Gower, C.; Ramsay, M.; Lopez Bernal, J. Effectiveness of COVID-19 booster vaccines against COVID-19-related symptoms, hospitalization and death in England. Nat. Med. 2022, 28, 831–837. [Google Scholar] [CrossRef]
- Pulliam, J.R.C.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased risk of SARS-CoV-2 reinfection associated with emergence of Omicron in South Africa. Science 2022, 376, eabn4947. [Google Scholar] [CrossRef]
- Feng, C.; Shi, J.; Fan, Q.; Wang, Y.; Huang, H.; Chen, F.; Tang, G.; Li, Y.; Li, P.; Li, J.; et al. Protective humoral and cellular immune responses to SARS-CoV-2 persist up to 1 year after recovery. Nat. Commun. 2021, 12, 4984. [Google Scholar] [CrossRef] [PubMed]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Tan, A.T.; Linster, M.; Tan, C.W.; Le Bert, N.; Chia, W.N.; Kunasegaran, K.; Zhuang, Y.; Tham, C.Y.L.; Chia, A.; Smith, G.J.D.; et al. Early induction of functional SARS-CoV-2-specific T cells associates with rapid viral clearance and mild disease in COVID-19 patients. Cell Rep. 2021, 34, 108728. [Google Scholar] [CrossRef] [PubMed]
- Le Bert, N.; Clapham, H.E.; Tan, A.T.; Chia, W.N.; Tham, C.Y.L.; Lim, J.M.; Kunasegaran, K.; Tan, L.W.L.; Dutertre, C.A.; Shankar, N.; et al. Highly functional virus-specific cellular immune response in asymptomatic SARS-CoV-2 infection. J. Exp. Med. 2021, 218, e20202617. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro, T.; Nair, N. Allergic Reactions Including Anaphylaxis After Receipt of the First Dose of Pfizer-BioNTech COVID-19 Vaccine. JAMA 2021, 325, 780–781. [Google Scholar] [CrossRef] [PubMed]
- CDC COVID-19 Response Team; Food and Drug Administration. Allergic Reactions Including Anaphylaxis After Receipt of the First Dose of Moderna COVID-19 Vaccine—United States, December 21, 2020–January 10, 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 125–129. [Google Scholar] [CrossRef]
- Greenhawt, M.; Abrams, E.M.; Shaker, M.; Chu, D.K.; Khan, D.; Akin, C.; Alqurashi, W.; Arkwright, P.; Baldwin, J.L.; Ben-Shoshan, M.; et al. The Risk of Allergic Reaction to SARS-CoV-2 Vaccines and Recommended Evaluation and Management: A Systematic Review, Meta-Analysis, GRADE Assessment, and International Consensus Approach. J. Allergy Clin. Immunol. Pract. 2021, 9, 3546–3567. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, T.; Umeda, H.; Kojima, M.; Kanno, Y.; Tanaka, Y.; Kinoshita, N.; Sato, D. Cumulative Adverse Event Reporting of Anaphylaxis After mRNA COVID-19 Vaccine (Pfizer-BioNTech) Injections in Japan: The First-Month Report. Drug Saf. 2021, 44, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Stone, C.A., Jr.; Liu, Y.; Relling, M.V.; Krantz, M.S.; Pratt, A.L.; Abreo, A.; Hemler, J.A.; Phillips, E.J. Immediate Hypersensitivity to Polyethylene Glycols and Polysorbates: More Common Than We Have Recognized. J. Allergy Clin. Immunol. Pract. 2019, 7, 1533–1540.e8. [Google Scholar] [CrossRef]
- Wenande, E.; Garvey, L.H. Immediate-type hypersensitivity to polyethylene glycols: A review. Clin. Exp. Allergy 2016, 46, 907–922. [Google Scholar] [CrossRef]
- Klimek, L.; Novak, N.; Cabanillas, B.; Jutel, M.; Bousquet, J.; Akdis, C.A. Allergenic components of the mRNA-1273 vaccine for COVID-19: Possible involvement of polyethylene glycol and IgG-mediated complement activation. Allergy 2021, 76, 3307–3313. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.E.; Shay, D.K.; Su, J.R.; Gee, J.; Creech, C.B.; Broder, K.R.; Edwards, K.; Soslow, J.H.; Dendy, J.M.; Schlaudecker, E.; et al. Myocarditis Cases Reported After mRNA-Based COVID-19 Vaccination in the US From December 2020 to August 2021. JAMA 2022, 327, 331–340. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, Q.; Wang, H.; Liu, W.; Liao, X.; Su, Y.; Wang, X.; Yuan, J.; Li, T.; Li, J.; et al. Antibody Responses to SARS-CoV-2 in Patients With Novel Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yan, R.; Liu, M.; Liu, Z.; Wang, Y.; Luan, D.; Wu, K.; Song, Z.; Sun, T.; Ma, Y.; et al. Antibody-dependent enhancement (ADE) of SARS-CoV-2 infection in recovered COVID-19 patients: Studies based on cellular and structural biology analysis. Medrxiv 2020. [Google Scholar] [CrossRef]
- Constant, S.L.; Bottomly, K. Induction of Th1 and Th2 CD4+ T cell responses: The alternative approaches. Annu. Rev. Immunol. 1997, 15, 297–322. [Google Scholar] [CrossRef]
- Rauch, S.; Roth, N.; Schwendt, K.; Fotin-Mleczek, M.; Mueller, S.O.; Petsch, B. mRNA-based SARS-CoV-2 vaccine candidate CVnCoV induces high levels of virus-neutralising antibodies and mediates protection in rodents. NPJ Vaccines 2021, 6, 57. [Google Scholar] [CrossRef]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schafer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef]
- Seo, Y.B.; Suh, Y.S.; Ryu, J.I.; Jang, H.; Oh, H.; Koo, B.S.; Seo, S.H.; Hong, J.J.; Song, M.; Kim, S.J.; et al. Soluble Spike DNA Vaccine Provides Long-Term Protective Immunity against SARS-CoV-2 in Mice and Nonhuman Primates. Vaccines 2021, 9, 307. [Google Scholar] [CrossRef]
- Nasreen, S.; Febriani, Y.; Velasquez Garcia, H.A.; Zhang, G.; Tadrous, M.; Buchan, S.A.; Righolt, C.H.; Mahmud, S.M.; Janjua, N.Z.; Krajden, M.; et al. Effectiveness of Coronavirus Disease 2019 Vaccines Against Hospitalization and Death in Canada: A Multiprovincial, Test-Negative Design Study. Clin. Infect. Dis. 2023, 76, 640–648. [Google Scholar] [CrossRef]
- Takheaw, N.; Liwsrisakun, C.; Chaiwong, W.; Laopajon, W.; Pata, S.; Inchai, J.; Duangjit, P.; Pothirat, C.; Bumroongkit, C.; Deesomchok, A.; et al. Correlation Analysis of Anti-SARS-CoV-2 RBD IgG and Neutralizing Antibody against SARS-CoV-2 Omicron Variants after Vaccination. Diagnostics 2022, 12, 1315. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).