Abstract
Multiple Sclerosis (MS) is a common immune-mediated disorder of the central nervous system that affects young adults and is characterized by demyelination and neurodegeneration. Recent studies have associated C9orf72 intermediate repeat expansions with MS. The objective of this study was to investigate whether C9orf72 repeat length is associated with MS or with a specific disease course in a monocentric Austrian MS cohort. Genotyping of 382 MS patients and 643 non-neurological controls for C9orf72 repeat expansions was performed. The study did not find a difference in the distribution of repeat numbers between controls and MS cases (median repeat units = 2; p = 0.39). Additionally, sub-analysis did not establish a link between intermediate repeats and MS (p = 0.23) and none of the patients with progressive disease course carried an intermediate allele (20–30 repeat units). Exploratory analysis for different cut-offs (of ≥7, ≥17, and ≥24) did not reveal any significant differences in allele frequencies between MS and controls. However, the study did identify a progressive MS patient with a pathogenic C9orf72 expansion and probable co-existing behavioral variant frontotemporal dementia (bvFTD) in a retrospective chart review. In conclusion, this study did not find evidence supporting an association between C9orf72 repeat length and MS or a specific disease course in the Austrian MS cohort. However, the identification of a progressive MS patient with a pathogenic C9orf72 expansion and probable co-existing with FTD highlights the complexity and challenges involved in recognizing distinct neurodegenerative diseases that may co-occur in MS patients.
1. Introduction
Multiple Sclerosis (MS) is the most common immune-mediated disorder of the central nervous system in young adults, characterized by demyelination and neurodegeneration. The conventional view of MS pathophysiology, supported by a plethora of available data, hypothesizes that auto-reactive cells are present in the immune repertoire of a susceptible host and overcome control mechanisms, e.g., autoimmune processes being elicited by environmental causes [1,2]. However, an alternative hypothesis that has been postulated is the so-called “inside out” model, with a primary neurodegenerative event followed by an immune response as a secondary consequence [3,4].
A GGGGCC hexanucleotide repeat expansion (HRE) in the first intron of the chromosome 9 open reading frame 72 (C9orf72) gene was discovered to be the most frequent cause of both frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS), as well as mixed FTD/ALS phenotype. While repeat units of hundreds or thousands are clearly disease-causing, a clear cut-off between normal and pathogenic repeat length has not been established in C9orf72 yet. In most studies, repeat numbers of >30 or >45 repeat units are considered pathogenic. In recent years, this variant has been linked to a broad clinical spectrum and multiple neurodegenerative diseases, as currently reviewed [5].
A possible correlation of C9orf72 repeat expansion with multiple sclerosis (MS) has been investigated previously. These studies, all performed in Italian patients, found no association of the pathogenic C9orf72 repeats expansion with MS pathogenesis [6,7,8]. Nevertheless, the pathogenic expansion was reported to occur in four of five patients with coexisting ALS and MS, showing a more rapid disease progression compared to patients with pure C9orf72 ALS. [6]. Although the concurrence of the two diseases is extremely rare, the authors suggested that MS might increase the risk of the C9orf72 mutation becoming penetrant and possibly modifies relevant pathogenic pathways. This was also discussed in a case report of a patient diagnosed with ALS carrying a C9orf72 repeat expansion and active brain demyelinating lesions, although the patient showed no MS symptoms [9].
Of note, a study examining a large cohort of Italian patients with progressive MS patients reported a significantly more frequent occurrence of alleles with intermediate repeat size between 20 and 30 repeat units [10]. The role of intermediate repeats in disease pathogenesis is to date not entirely clear, but they have been suggested to be a susceptibility risk factor for other neurodegenerative diseases and disease-modifier [11,12,13,14].
In this study, we screened a large Austrian MS cohort for the C9orf72 repeat expansion. Besides evaluating repeat length distribution and frequency of intermediate and pathogenic repeat alleles in MS patients, we describe a unique case with co-incidental pathogenic C9orf72 HRE in a patient with MS and probable co-existing FTD.
2. Results
2.1. C9orf72 Repeat Length Distribution in Multiple Sclerosis
In the first step, the C9orf72 repeat length distribution of the patients was compared to healthy controls. Repeat ranges of 2–27 was observed in the neurologically healthy control (HC) cohort (n = 643) and a range of 2–22 in the multiple sclerosis (MS) cohort (n = 382). Similarly, to the distribution reported previously, a characteristic trimodal pattern was observed with peaks at two, five, and eight repeat units, as shown in Figure 1. There was no difference between repeat number distribution in controls and MS cases (p = 0.39). Notably, we identified one patient diagnosed with secondary progressive multiple sclerosis (SPMS) carrying a repeat expansion of approximately 1300 repeat units, reported in detail below.
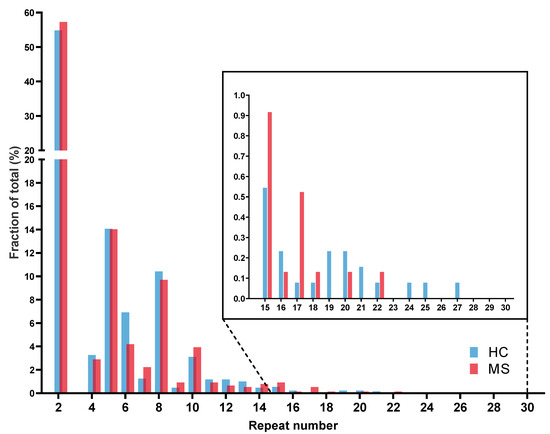
Figure 1.
C9orf72 repeat allele length in multiple sclerosis patients.
The histogram shows the relative frequency of repeat lengths in HC (n = 643) and MS (n = 382) cohorts. Both cohorts show a characteristic trimodal pattern with high frequency of alleles of two, five, and eight repeats. Cutout shows repeat lengths between 15 and 30 repeat units. There was no significant difference in allele frequency (Mann–Whitney U-test, p = 0.39).
Abbr.: HC = healthy controls; MS = multiple sclerosis.
2.2. C9orf72 Intermediate Allele Frequency
Considering a previous study suggesting a correlation between intermediate repeats (defined as 20–30 repeats) and primary progressive multiple sclerosis (PPMS) [10], we took a more detailed look at the occurrence of intermediate alleles and different sub-phenotypes in our cohort. Accordingly, we have classified repeat length as small (S; <20 repeats); intermediate (I; 20–30 repeats) and expanded (E; >30 repeats) alleles. Again, there was no difference between repeat lengths using these cut-offs.
Only 11 carriers of alleles ≥20 repeat units were identified, nine in the group of HC (0.7%) and two in the SPMS cohort (1.6%) without significant difference (p = 0.257). Mean repeat lengths and allele frequencies are shown in Table 1. Notably, the SPMS patient who was a carrier of a pathogenic C9orf72 expansion allele was also a carrier of an intermediate repeat allele.

Table 1.
Frequency of C9orf72 repeat lengths in MS sub-cohorts.
Additionally, other thresholds were tested, as the cut-off for the intermediate repeat length is not well established. We, therefore, assessed cut-offs of ≥7, ≥17 and ≥24, suggested in the literature based on specific risk haplotypes, studies on DNA methylation, and gene expression and association with increased risk for specific diseases [15,16,17,18,19,20]. Again, there were no significant differences in allele frequencies at those cut-offs (shown in Table 2).
Table 2.
Intermediate repeat lengths.
Fisher’s exact test was used to compare frequencies of intermediate repeat alleles at different cut-offs using Holm–Sidak correction for multiple testing.
2.3. C9orf72 Hexanucleotide Repeat Expansion Carrier
Remarkably, we found one patient with a SPMS disease course carrying a pathogenic C9orf72 repeat expansion. Southern blotting revealed an expanded allele of approximately 1300 units and an intermediate repeat of 20 units on the other allele (see Supplementary Figure S1).
The male patient was diagnosed with RRMS at the age of 28 years with no reported family history of neurodegenerative diseases. The patient was followed up regularly at the outpatient clinic of the Department of Neurology of the MUV from the age of 38 years. At the age of 45, acute worsening of symptoms occurred. The patient was admitted to the hospital after he was found completely immobile at home. At this time, a progressive deterioration of walking ability within the last two weeks was reported. The patient stated that he had recently been unable to perform hygiene adequately and that he had not continued his established Betaferon therapy for the past two weeks. The patient himself associated his symptomatology with the cold wind (“draught”). At the time of admission, clinical neurological examination revealed spastic tetraparesis and ataxia, corresponding to an EDSS (expanded disability status scale) score of 8.0. During the admission, MRI imaging of brain and spinal cord was performed, revealing multiple spinal hyperintensities as well as marked atrophy of the cervical and thoracic spinal cord. Likewise, numerous non-enhancing T2 lesions were present supratentorially on both hemispheres with generalized supratentorial and perisylvian accentuated signs of atrophy. In the neuropsychiatric examination, below-average scores were observed in Rey–Osterrieth Complex Figure Test (testing perception) and tests for divided attention. At the Wechsler Memory Scale (WMS-IV), the patient achieved below-average values in the areas of logical short- and long-term memory. Moreover, verbal fluency in terms of formal-lexical as well as semantic-categorical (Regensburger Wortflüssigkeitstest) and non-verbal fluency (HAMASCH test) was below-average. In summary, deficits in the cognitive domains: attention, short- and longer-term verbal memory, and executive functions were present. Interestingly, the patient displayed perceptual changes and mild paranoid thoughts during the inpatient stay. At the age of 46, SPMS was diagnosed. The patient committed suicide at 50 years of age, no postmortem material was available and no biomarker testing such as PET or CSF analysis specific for dementia was performed.
3. Discussion
In our study, we determined the C9orf72 repeat length in an Austrian MS cohort in order to investigate the frequency of intermediate alleles ≥20 repeat units, as they were suggested to occur in higher frequency in patients diagnosed with primary progressive MS (PPMS) [10].
We did not observe a correlation of the C9orf72 repeat length with MS or a specific disease course in our cohort. Likewise, when considering different cut-offs for intermediate repeats as previously suggested, repeat sizes between MS patients and control groups did not differ significantly in our cohort [10,21,22]. Notably, we identified one patient with a progressive disease course who was a carrier of the pathogenic C9orf72 repeat expansion. The retrospective chart review of the medical history revealed a probable psychiatric symptomatology with the occurrence of psychotic symptoms and the committing of suicide, which is suggestive of a co-existing bvFTD pathology according to criteria of Rascovsky et al., although no post-mortem material was available [23]. The symptoms of the patient fit psychological features enriched in C9orf72 repeat expansion carriers, which commonly include delusions and hallucinations, mood disorders, obsessive compulsive disorder and catatonia [24,25,26,27] and were reported to occur in some patients several years before characteristic ALS or FTD onset [24,27]. Atrophy patterns observed in MRI imaging of the patient were also seen in C9orf72 patients diagnosed with FTD [28].
Pathogenic C9orf72 repeat expansions were previously identified in patients with very rare concurrence of MS and amyotrophic lateral sclerosis (ALS), but not in patients without concomitant disease [6]. There are no reports on C9orf72 HRE in patients with MS and coexisting FTD, although neurodegenerative dementias become more common in this diagnostic group as persons diagnosed with MS have higher life expectancy today [29,30]. However, cognitive impairment is also observed in MS patients, more pronounced at later disease stages. Cognitive impairment related to MS has shown a strong involvement of information processing speed, while other domains are less affected, including characteristic changes in neurodegenerative dementias [31]. As there are overlaps between typically affected domains, distinguishing cognitive impairment due to MS from neurodegenerative dementia co-occurring with MS based only on neuropsychiatric examination is difficult [32,33,34]. Therefore, a major limitation of this study is the lack of more detailed diagnostic work up and post-mortem tissue, as the definite diagnosis of FTD and other neurodegenerative disorders still depends on neuropathological findings in autopsy. Thus, we cannot say with certainty whether the symptoms and changes in MRI described are definitely due to the C9orf72 repeat expansion mutation.
Furthermore, the lack of association between intermediate allele size and PPMS in our cohort could also be due to a low patient number of PPMS patients in our cohort.
In summary, our findings suggest that there is no significant contribution of intermediate C9orf72 repeat alleles to the pathogenesis of the MS clinical spectrum in Austrian patients. Furthermore, we describe a patient with MS and pronounced psychiatric symptoms who is also a carrier of the pathogenic C9orf72 HRE which we interpret most likely as coincidental. However, this case is exemplary of the increasing evidence and recognition of mixed neurodegenerative pathologies as patients get older these days [35,36]. The cases described in this study underline the need for further research on specific in vivo biomarkers and clinical red flags and, eventually, the importance to consider additional diagnoses in the context of atypical or rapid disease progression. Deciphering whether cognitive impairment is due to neurodegenerative changes in the course of MS itself or comorbid occurrence of neurodegenerative dementia will become increasingly important in order to provide optimal patient counseling and therapy.
4. Materials and Methods
4.1. Patient Cohort
DNA samples were derived from blood leukocytes of 382 unrelated patients diagnosed with MS from the biobank of the Department of Neurology of the Medical University of Vienna (MUV). A total of 643 DNA samples of neurologically unaffected individuals obtained from the biobank of the Department of Neurology of the MUV (n = 489) or acquired from the VITA collective (n = 154) served as healthy controls (HC). Written informed consent was obtained from all participants and ethical approval is available for all collectives. The study was approved by the ethics committee of the Medical University of Vienna (EK Nr 1220/2018). MS patients were clinically diagnosed based on established criteria [37] and assigned to sub-cohorts PPMS (n = 19), SPMS (n = 63) or RRMS (n = 269) according to their clinical phenotype [38]. A total of 31 patients could not be assigned to a distinct phenotype due to loss of follow-up. Information on patient characteristics is shown in Table 3.
Table 3.
Characteristics of study participants.
4.2. C9orf72 Genotyping
DNA was isolated from peripheral whole blood using standard protocols. C9orf72 genotyping was performed as described before [14]. In short, a two-step PCR assay was used to amplify the repeat sequence: In the first step, the complete 5′UTR of the C9orf72 gene was amplified using primers flanking the pathological repeat region, followed by repeat primed PCR (RP-PCR). To validate C9orf72 repeat expansions in individuals showing a characteristic chainsaw pattern in the RP-PCR analysis and to estimate the repeat length, Southern blotting was performed (see Supplementary Figure S1).
4.3. Statistics
For non-pathogenic range (2–30 repeats), we performed a Mann–Whitney U-Test to evaluate repeat length distribution between cohorts. Fisher’s exact test was used to compare frequencies of intermediate repeat alleles at different cut-offs (≥7, ≥17, ≥20, and ≥24). Correction for multiple testing was performed using Dunn’s multiple comparison test and Holm–Sidak correction for contingency analysis. Statistical analyses were performed using the software GraphPad Prism v8.0.0. A p-value of <0.05 was considered statistically significant.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241411254/s1.
Author Contributions
Conceptualization, T.K. and T.Z.; methodology, T.K., F.L., A.Z., C.S., E.S. and T.Z.; validation, E.S., F.L. and T.Z.; formal analysis, T.K. and T.Z.; resources, T.B. and E.S.; data curation, T.K. and T.Z.; writing—original draft preparation, T.K. and T.Z.; writing—review and editing, T.K., F.L., A.Z., T.B., C.S., E.S. and T.Z.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study has been performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. The study was approved by the ethics committee of the Medical University of Vienna (EK Nr 1220/2018).
Informed Consent Statement
Written informed consent was obtained from all participants and ethical approval is available for all collectives.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author, E.S.
Conflicts of Interest
T.K., F.L., A.Z., C.S. and T.Z. have nothing to disclose. T.B. has received honoraria and consulting fees from Almirall, Bayer, Biogen, Biologix, Bionorica, Celgene/BMS, Genesis, GSK, Horizon, Janssen-Cilag, Jazz, MedDay, Merck, Novartis, Octapharma, Roche, Sandoz, Sanofi-Genzyme, TEVA, TG Therapeutics, and UCB. His institution has received financial support by unrestricted research grants from Almirall, Biogen, Bayer, Celgene/BMS, Merck, Novartis, Roche, Sanofi-Genzyme and TEVA, and honoraria for participation in clinical trials sponsored by Alexion, Bayer, Biogen, Celgene/BMS, Merck, Novartis, Octapharma, Roche, Sanofi, and TEVA. In the past 36 months, E.S. received grants from Roche, Eisai, FFG/AAL, Horizon2020 and the Austrian Alzheimer Association (all to the institution). E.S. received consulting fees from Biogen and received support for attending meetings and/or travel from Roche. E.S. received for lectures, presentations, speakers bureaus, manuscript writing, or educational events payment by Biogen, Roche, Eisai, and Novartis. E. Stögmann participated on an advisory board (Biogen, Roche, Eisai) and held leadership or a fiduciary role in scientific societies (Austrian Alzheimer Association, the EAN scientific panel dementia). The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Attfield, K.E.; Jensen, L.T.; Kaufmann, M.; Friese, M.A.; Fugger, L. The immunology of multiple sclerosis. Nat. Rev. Immunol. 2022, 22, 734–750. [Google Scholar] [CrossRef]
- Bar-Or, A.; Li, R. Cellular immunology of relapsing multiple sclerosis: Interactions, checks, and balances. Lancet Neurol. 2021, 20, 470–483. [Google Scholar] [CrossRef] [PubMed]
- ’t Hart, B.A.; Luchicchi, A.; Schenk, G.J.; Stys, P.K.; Geurts, J.J.G. Mechanistic underpinning of an inside-out concept for autoimmunity in multiple sclerosis. Ann. Clin. Transl. Neurol. 2021, 8, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Stys, P.K.; Zamponi, G.W.; van Minnen, J.; Geurts, J.J. Will the real multiple sclerosis please stand up? Nat. Rev. Neurosci. 2012, 13, 507–514. [Google Scholar] [CrossRef] [PubMed]
- van der Ende, E.L.; Jackson, J.L.; White, A.; Seelaar, H.; van Blitterswijk, M.; Van Swieten, J.C. Unravelling the clinical spectrum and the role of repeat length in C9ORF72 repeat expansions. J. Neurol. Neurosurg. Psychiatry 2021, 92, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Cooper-Knock, J.; Highley, J.R.; Milano, A.; Kirby, J.; Goodall, E.; Lowe, J.; Scott, I.; Constantinescu, C.S.; Walters, S.J.; et al. Concurrence of multiple sclerosis and amyotrophic lateral sclerosis in patients with hexanucleotide repeat expansions of C9ORF72. J. Neurol. Neurosurg. Psychiatry 2013, 84, 79–87. [Google Scholar] [CrossRef]
- Lorefice, L.; Murru, M.R.; Fenu, G.; Corongiu, D.; Frau, J.; Cuccu, S.; Coghe, G.C.; Tranquilli, S.; Cocco, E.; Marrosu, M.G. A genetic association study of two genes linked to neurodegeneration in a Sardinian multiple sclerosis population: The TARDBP Ala382Thr mutation and C9orf72 expansion. J. Neurol. Sci. 2015, 357, 229–234. [Google Scholar] [CrossRef]
- Fenoglio, C.; De Riz, M.; Villa, C.; Serpente, M.; Ridolfi, E.; Bonsi, R.; Cioffi, S.M.; Barone, C.; Pietroboni, A.; Calvi, A.; et al. C9ORF72 repeat expansion not detected in patients with multiple sclerosis. Neurobiol. Aging 2014, 35, 1213.e1–1213.e2. [Google Scholar] [CrossRef]
- Oliveira Santos, M.; Caldeira, I.; Gromicho, M.; Pronto-Laborinho, A.; de Carvalho, M. Brain white matter demyelinating lesions and amyotrophic lateral sclerosis in a patient with C9orf72 hexanucleotide repeat expansion. Mult. Scler. Relat. Disord. 2017, 17, 1–4. [Google Scholar] [CrossRef]
- Tiloca, C.; Sorosina, M.; Esposito, F.; Peroni, S.; Colombrita, C.; Ticozzi, N.; Ratti, A.; Martinelli-Boneschi, F.; Silani, V. No C9orf72 repeat expansion in patients with primary progressive multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 25, 192–195. [Google Scholar] [CrossRef]
- Ng, A.S.L.; Tan, E.K. Intermediate C9orf72 alleles in neurological disorders: Does size really matter? J. Med. Genet. 2017, 54, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Serpente, M.; Fenoglio, C.; Arighi, A.; Fumagalli, G.G.; Arcaro, M.; Sorrentino, F.; Visconte, C.; Scarpini, E.; Galimberti, D. Analysis of C9orf72 Intermediate Alleles in a Retrospective Cohort of Neurological Patients: Risk Factors for Alzheimer’s Disease? J. Alzheimer’s Dis. JAD 2021, 81, 1445–1451. [Google Scholar] [CrossRef]
- Kaivola, K.; Kiviharju, A.; Jansson, L.; Rantalainen, V.; Eriksson, J.G.; Strandberg, T.E.; Laaksovirta, H.; Renton, A.E.; Traynor, B.J.; Myllykangas, L.; et al. C9orf72 hexanucleotide repeat length in older population: Normal variation and effects on cognition. Neurobiol. Aging 2019, 84, 242.e7–242.e12. [Google Scholar] [CrossRef]
- Konig, T.; Wurm, R.; Parvizi, T.; Silvaieh, S.; Hotzy, C.; Cetin, H.; Klotz, S.; Gelpi, E.; Bancher, C.; Benke, T.; et al. C9orf72 repeat length might influence clinical sub-phenotypes in dementia patients. Neurobiol. Dis. 2022, 175, 105927. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Tortosa, E.; Gallego, J.; Guerrero-Lopez, R.; Marcos, A.; Gil-Neciga, E.; Sainz, M.J.; Diaz, A.; Franco-Macias, E.; Trujillo-Tiebas, M.J.; Ayuso, C.; et al. C9ORF72 hexanucleotide expansions of 20-22 repeats are associated with frontotemporal deterioration. Neurology 2013, 80, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Nuytemans, K.; Bademci, G.; Kohli, M.M.; Beecham, G.W.; Wang, L.; Young, J.I.; Nahab, F.; Martin, E.R.; Gilbert, J.R.; Benatar, M.; et al. C9ORF72 intermediate repeat copies are a significant risk factor for Parkinson disease. Ann. Hum. Genet. 2013, 77, 351–363. [Google Scholar] [CrossRef]
- van der Zee, J.; Gijselinck, I.; Dillen, L.; Van Langenhove, T.; Theuns, J.; Engelborghs, S.; Philtjens, S.; Vandenbulcke, M.; Sleegers, K.; Sieben, A.; et al. A pan-European study of the C9orf72 repeat associated with FTLD: Geographic prevalence, genomic instability, and intermediate repeats. Hum. Mutat. 2013, 34, 363–373. [Google Scholar] [CrossRef]
- Gijselinck, I.; Van Mossevelde, S.; van der Zee, J.; Sieben, A.; Engelborghs, S.; De Bleecker, J.; Ivanoiu, A.; Deryck, O.; Edbauer, D.; Zhang, M.; et al. The C9orf72 repeat size correlates with onset age of disease, DNA methylation and transcriptional downregulation of the promoter. Mol. Psychiatry 2016, 21, 1112–1124. [Google Scholar] [CrossRef] [PubMed]
- Cali, C.P.; Patino, M.; Tai, Y.K.; Ho, W.Y.; McLean, C.A.; Morris, C.M.; Seeley, W.W.; Miller, B.L.; Gaig, C.; Vonsattel, J.P.G.; et al. C9orf72 intermediate repeats are associated with corticobasal degeneration, increased C9orf72 expression and disruption of autophagy. Acta Neuropathol. 2019, 138, 795–811. [Google Scholar] [CrossRef]
- Iacoangeli, A.; Al Khleifat, A.; Jones, A.R.; Sproviero, W.; Shatunov, A.; Opie-Martin, S.; Alzheimer’s Disease Neuroimaging, I.; Morrison, K.E.; Shaw, P.J.; Shaw, C.E.; et al. C9orf72 intermediate expansions of 24–30 repeats are associated with ALS. Acta Neuropathol. Commun. 2019, 7, 115. [Google Scholar] [CrossRef]
- Biasiotto, G.; Zanella, I. The effect of C9orf72 intermediate repeat expansions in neurodegenerative and autoimmune diseases. Mult. Scler. Relat. Disord. 2019, 27, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Tiloca, C.; Sorosina, M.; Esposito, F.; Peroni, S.; Colombrita, C.; Ticozzi, N.; Ratti, A.; Martinelli-Boneschi, F.; Silani, V. Response to the commentary “The effect of C9orf72 intermediate repeat expansions in neurodegenerative and autoimmune diseases” by Biasiotto G and Zanella I. Mult. Scler. Relat. Disord. 2019, 27, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Rascovsky, K.; Hodges, J.R.; Knopman, D.; Mendez, M.F.; Kramer, J.H.; Neuhaus, J.; van Swieten, J.C.; Seelaar, H.; Dopper, E.G.; Onyike, C.U.; et al. Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain 2011, 134 Pt 9, 2456–2477. [Google Scholar] [CrossRef] [PubMed]
- Snowden, J.S.; Rollinson, S.; Thompson, J.C.; Harris, J.M.; Stopford, C.L.; Richardson, A.M.; Jones, M.; Gerhard, A.; Davidson, Y.S.; Robinson, A.; et al. Distinct clinical and pathological characteristics of frontotemporal dementia associated with C9ORF72 mutations. Brain 2012, 135 Pt 3, 693–708. [Google Scholar] [CrossRef]
- Solje, E.; Aaltokallio, H.; Koivumaa-Honkanen, H.; Suhonen, N.M.; Moilanen, V.; Kiviharju, A.; Traynor, B.; Tienari, P.J.; Hartikainen, P.; Remes, A.M. The Phenotype of the C9ORF72 Expansion Carriers According to Revised Criteria for bvFTD. PLoS ONE 2015, 10, e0131817. [Google Scholar] [CrossRef]
- Bieniek, K.F.; van Blitterswijk, M.; Baker, M.C.; Petrucelli, L.; Rademakers, R.; Dickson, D.W. Expanded C9ORF72 hexanucleotide repeat in depressive pseudodementia. JAMA Neurol. 2014, 71, 775–781. [Google Scholar] [CrossRef]
- Block, N.R.; Sha, S.J.; Karydas, A.M.; Fong, J.C.; De May, M.G.; Miller, B.L.; Rosen, H.J. Frontotemporal Dementia and Psychiatric Illness: Emerging Clinical and Biological Links in Gene Carriers. Am. J. Geriatr. Psychiatry Off. J. Am. Assoc. Geriatr. Psychiatry 2016, 24, 107–116. [Google Scholar] [CrossRef]
- Saracino, D.; Geraudie, A.; Remes, A.M.; Ferrieux, S.; Nogues-Lassiaille, M.; Bottani, S.; Cipriano, L.; Houot, M.; Funkiewiez, A.; Camuzat, A.; et al. Primary progressive aphasias associated with C9orf72 expansions: Another side of the story. Cortex 2021, 145, 145–159. [Google Scholar] [CrossRef]
- Martinelli, V.; Rodegher, M.; Moiola, L.; Comi, G. Late onset multiple sclerosis: Clinical characteristics, prognostic factors and differential diagnosis. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2004, 25 (Suppl. 4), S350–S355. [Google Scholar] [CrossRef]
- Goodin, D.S. Disease-modifying therapy in multiple sclerosis: Update and clinical implications. Neurology 2008, 71 (Suppl. 3), S8–S13. [Google Scholar] [CrossRef]
- Branco, M.; Ruano, L.; Portaccio, E.; Goretti, B.; Niccolai, C.; Patti, F.; Chisari, C.; Gallo, P.; Grossi, P.; Ghezzi, A.; et al. Aging with multiple sclerosis: Prevalence and profile of cognitive impairment. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2019, 40, 1651–1657. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, P.; Penner, I.K. Cognitive dysfunctions in multiple sclerosis—A “multiple disconnection syndrome”? J. Neurol. 2007, 254 (Suppl. 2), II18–II21. [Google Scholar] [CrossRef]
- Chiaravalloti, N.D.; DeLuca, J. Cognitive impairment in multiple sclerosis. Lancet Neurol. 2008, 7, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Londono, D.P.; Arumaithurai, K.; Constantopoulos, E.; Basso, M.R.; Reichard, R.R.; Flanagan, E.P.; Keegan, B.M. Diagnosis of coexistent neurodegenerative dementias in multiple sclerosis. Brain Commun. 2022, 4, fcac167. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G.; Alafuzoff, I.; Al-Sarraj, S.; Arzberger, T.; Bogdanovic, N.; Capellari, S.; Ferrer, I.; Gelpi, E.; Kovari, V.; Kretzschmar, H.; et al. Mixed brain pathologies in dementia: The BrainNet Europe consortium experience. Dement. Geriatr. Cogn. Disord. 2008, 26, 343–350. [Google Scholar] [CrossRef]
- Rahimi, J.; Kovacs, G.G. Prevalence of mixed pathologies in the aging brain. Alzheimer’s Res. Ther. 2014, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sorensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).