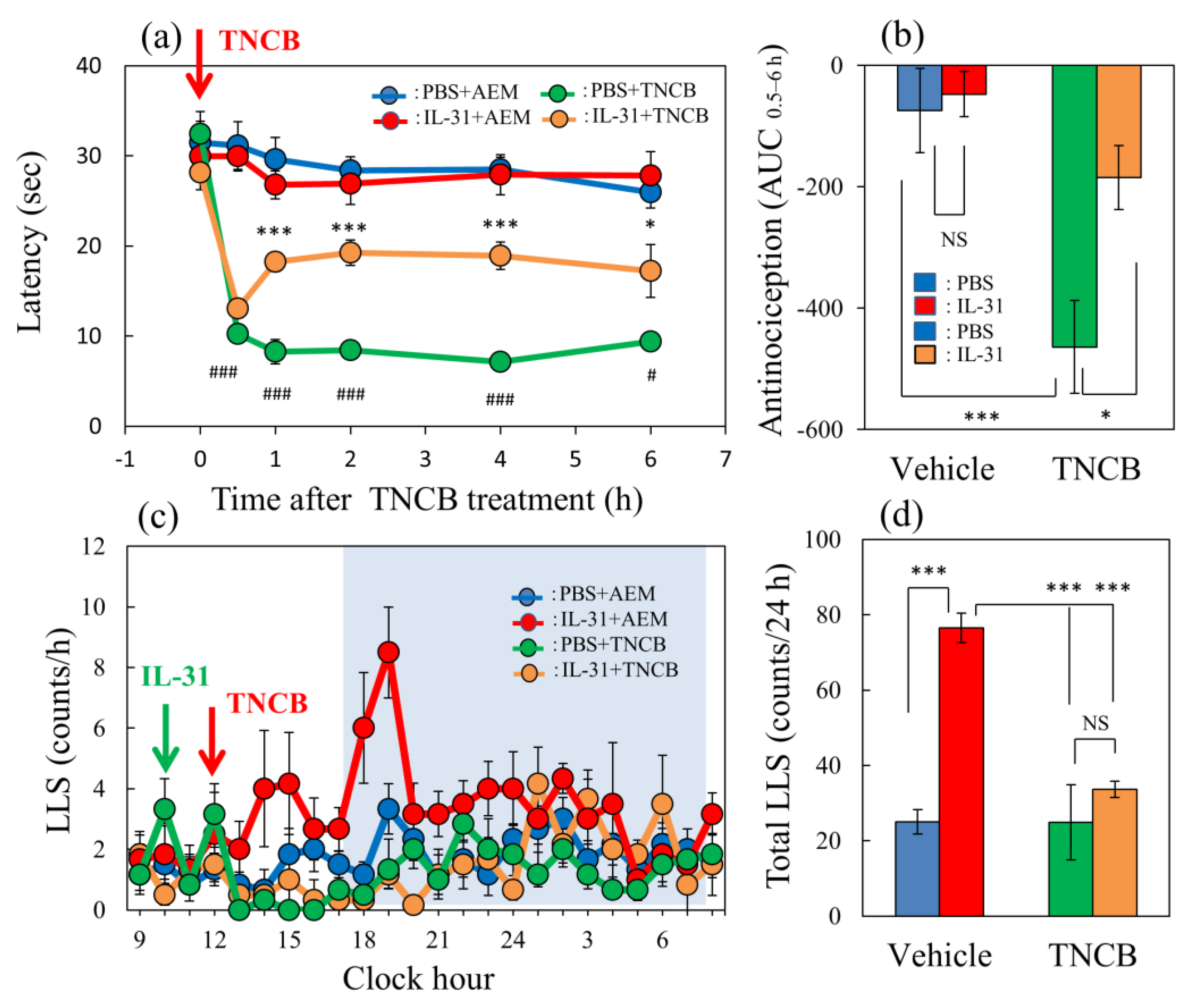
Figure 1.
Effect of IL-31 on TNCB-applied hot-plate test in BALB/c mice. (a) Effect of repeated pretreatment of IL-31 (50 μg/kg, intraperitoneally, every 12 h for three days) on TNCB (3%, 0.04 mL/each limb, total 0.16 mL/site)-applied hot-plate (45 °C) test. # p < 0.05, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. (b) Total antinociceptive index of IL-31 after 0.5 to 6 h (AUC0.5–6 h). (c) Time-course change of itch-associated scratching behavior (LLS, counts/h) after the application of TNCB (3%, 0.2 mL/dorsal site). The blue line indicates the PBS + AEM-treated group; the red line indicates IL-31 + AEM-treated group; the green line indicates PBS + TNCB-treated group; the yellow line indicates IL-31 + TNCB-treated group. The blue column indicates the PBS + AEM-treated group; the red column indicates the IL-31 + AEM-treated group; the green column indicates the PBS + TNCB-treated group; the yellow column indicates the IL-31 + TNCB-treated group. (a,b) * p < 0.05, *** p < 0.001 compared with the corresponding values in the PBS + TNCB-treated group. The lateral axis indicates the clock hour, and the shaded area represents nighttime (dark phase, 7:00 pm to 7:00 am). (d) Total LLS counts for 24 h. The blue column indicates the IL-31 + AEM-treated group; the red column indicates the IL-31 + TNCB-treated group; the green column indicates the PBS + TNCB-treated group; the yellow column indicates the IL-31 + TNCB-treated group. The red arrow indicates the AEM or TNCB application point. The blue arrow indicates the IL-31 (1 mg/kg, intraperitoneally) administration point. Each value represents the mean ± standard error (S.E.) from 6 mice (total 36 mice). *** p < 0.001 compared with the vehicle (PBS) + TNCB-treated group. Three-way ANOVA: group × treatment × time: F (5, 120) = 2.760, p = 0.0214, (a); group × treatment × time: F (23, 480) = 1.230, p = 0.2124, (c). Two-way ANOVA: group × treatment: F (1, 20) = 4.303, p = 0.0512, group: F (1, 20) = 18.68, p = 0.0003, treatment: F (1, 20) = 6.296, p = 0.0208, (b); group × treatment: F (1, 20) = 13.95, p = 0.0013, group: F (1, 20) = 14.16, p = 0.0012, treatment: F (1, 20) = 27.89, p < 0.0001, (d).
Figure 1.
Effect of IL-31 on TNCB-applied hot-plate test in BALB/c mice. (a) Effect of repeated pretreatment of IL-31 (50 μg/kg, intraperitoneally, every 12 h for three days) on TNCB (3%, 0.04 mL/each limb, total 0.16 mL/site)-applied hot-plate (45 °C) test. # p < 0.05, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. (b) Total antinociceptive index of IL-31 after 0.5 to 6 h (AUC0.5–6 h). (c) Time-course change of itch-associated scratching behavior (LLS, counts/h) after the application of TNCB (3%, 0.2 mL/dorsal site). The blue line indicates the PBS + AEM-treated group; the red line indicates IL-31 + AEM-treated group; the green line indicates PBS + TNCB-treated group; the yellow line indicates IL-31 + TNCB-treated group. The blue column indicates the PBS + AEM-treated group; the red column indicates the IL-31 + AEM-treated group; the green column indicates the PBS + TNCB-treated group; the yellow column indicates the IL-31 + TNCB-treated group. (a,b) * p < 0.05, *** p < 0.001 compared with the corresponding values in the PBS + TNCB-treated group. The lateral axis indicates the clock hour, and the shaded area represents nighttime (dark phase, 7:00 pm to 7:00 am). (d) Total LLS counts for 24 h. The blue column indicates the IL-31 + AEM-treated group; the red column indicates the IL-31 + TNCB-treated group; the green column indicates the PBS + TNCB-treated group; the yellow column indicates the IL-31 + TNCB-treated group. The red arrow indicates the AEM or TNCB application point. The blue arrow indicates the IL-31 (1 mg/kg, intraperitoneally) administration point. Each value represents the mean ± standard error (S.E.) from 6 mice (total 36 mice). *** p < 0.001 compared with the vehicle (PBS) + TNCB-treated group. Three-way ANOVA: group × treatment × time: F (5, 120) = 2.760, p = 0.0214, (a); group × treatment × time: F (23, 480) = 1.230, p = 0.2124, (c). Two-way ANOVA: group × treatment: F (1, 20) = 4.303, p = 0.0512, group: F (1, 20) = 18.68, p = 0.0003, treatment: F (1, 20) = 6.296, p = 0.0208, (b); group × treatment: F (1, 20) = 13.95, p = 0.0013, group: F (1, 20) = 14.16, p = 0.0012, treatment: F (1, 20) = 27.89, p < 0.0001, (d).

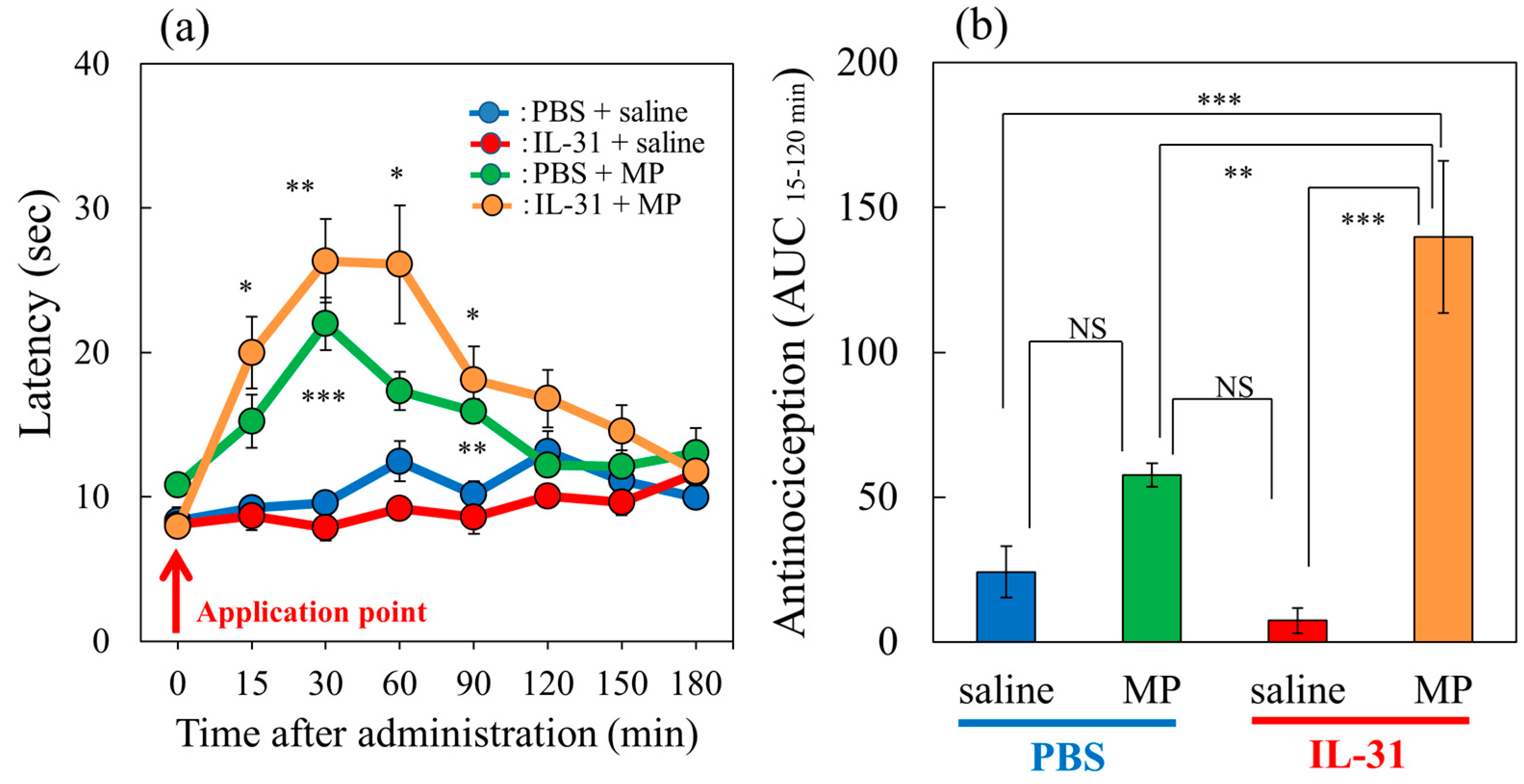
Figure 2.
Effects of IL-31 and morphine on conventional hot-plate (51 °C) test in BALB/c mice. (a) Effect of repeated pretreatment of IL-31 (50 μg/kg, intraperitoneally, every 12 h for three days) and a single dose of morphine (MP, 3 mg/kg, subcutaneously) on conventional hot-plate (51 °C) test. The blue line indicates the phosphate-buffered saline (PBS) + saline-treated group; the red line indicates the IL-31 + saline-treated group; the green line indicates PBS + morphine-treated group; the yellow line indicates IL-31 + morphine-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (24 mice). * p < 0.05, ** p < 0.01, *** p < 0.001 compared with the corresponding values in the PBS + saline-treated group. (b) Total antinociceptive index of morphine after 15 to 120 min (AUC15–120 min). The blue column indicates PBS + saline-treated group; the green column indicates PBS + morphine-treated group. The red column indicates the IL-31 + saline-treated group; the yellow column indicates IL-31 + morphine-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (total 24 mice). * p < 0.05, ** p < 0.01, *** p < 0.001 compared with each group. Three-way ANOVA: group × treatment × time: F (7, 168) = 2.360, p = 0.0252, (a). Two-way ANOVA: group × treatment: F (1, 20) = 12.11, p = 0.0024, group: F (1, 20) = 34.12, p < 0.0001, treatment: F (1, 20) = 5.28, p = 0.0325, (b).
Figure 2.
Effects of IL-31 and morphine on conventional hot-plate (51 °C) test in BALB/c mice. (a) Effect of repeated pretreatment of IL-31 (50 μg/kg, intraperitoneally, every 12 h for three days) and a single dose of morphine (MP, 3 mg/kg, subcutaneously) on conventional hot-plate (51 °C) test. The blue line indicates the phosphate-buffered saline (PBS) + saline-treated group; the red line indicates the IL-31 + saline-treated group; the green line indicates PBS + morphine-treated group; the yellow line indicates IL-31 + morphine-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (24 mice). * p < 0.05, ** p < 0.01, *** p < 0.001 compared with the corresponding values in the PBS + saline-treated group. (b) Total antinociceptive index of morphine after 15 to 120 min (AUC15–120 min). The blue column indicates PBS + saline-treated group; the green column indicates PBS + morphine-treated group. The red column indicates the IL-31 + saline-treated group; the yellow column indicates IL-31 + morphine-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (total 24 mice). * p < 0.05, ** p < 0.01, *** p < 0.001 compared with each group. Three-way ANOVA: group × treatment × time: F (7, 168) = 2.360, p = 0.0252, (a). Two-way ANOVA: group × treatment: F (1, 20) = 12.11, p = 0.0024, group: F (1, 20) = 34.12, p < 0.0001, treatment: F (1, 20) = 5.28, p = 0.0325, (b).

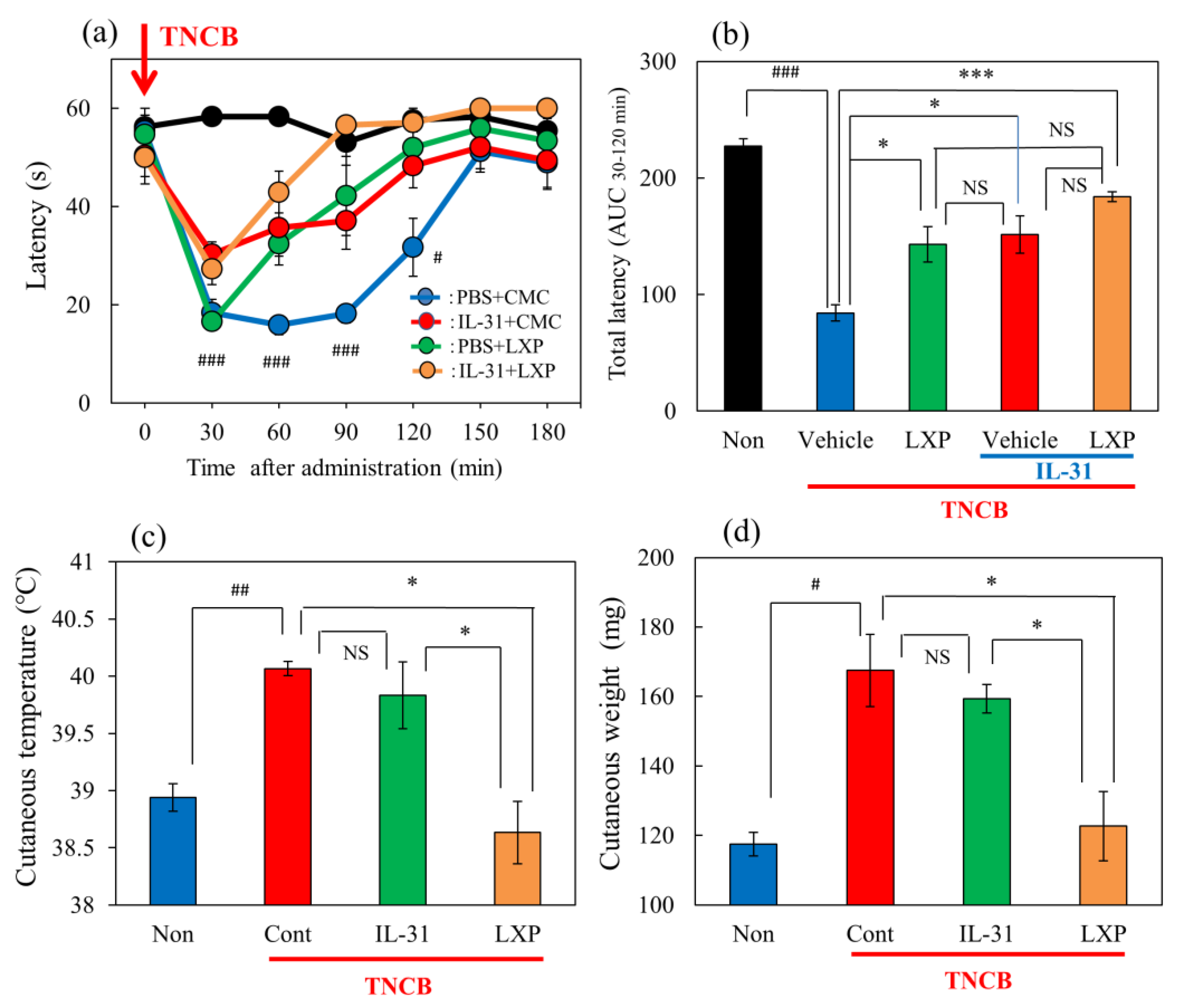
Figure 3.
Effects of IL-31 and loxoprofen (LXP) on TNCB–applied the hot-plate test in BALB/c mice. (a) Effects of IL-31 and loxoprofen (LXP) on TNCB-applied hot-plate (45 °C) test in mice. The black line and column indicate the non-TNCB-treated group; the blue line indicates phosphate-buffered saline (PBS) + vehicle (carboxymethyl cellulose sodium, CMC, 10 mL/kg, oral)-treated group; the red line indicates IL-31 + CMC-treated group; the green line indicates PBS + LXP-treated group; the yellow line indicate IL-31 + LXP treated group. The red arrow indicates the AEM or TNCB application point. # p < 0.05, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. (b) Total antinociceptive index of LXP after 30–120 min (AUC30–120 min). The black line and column indicate the non-treated group; the blue column indicates the PBS + CMC-treated group; the green column indicates the PBS + LXP-treated group; the red column indicates the IL-31 + CMC treated-group; the yellow column indicates the IL-31 + LXP-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (total 54 mice). NS, not significant, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. * p < 0.05, *** p < 0.001 compared with the corresponding values in the vehicle + TNCB treated group; (c) Effects of IL-31 and LXP on TNCB-induced increasing cutaneous temperature (fever) after 24 h application of TNCB. (d) Effects of IL-31 and LXP on TNCB-induced increasing cutaneous weight (swelling) after 24 h application of TNCB. The blue column indicates saline + vehicle-treated group; the red column indicates vehicle + TNCB-treated group; the green column indicates IL-31 + TNCB-treated group; the yellow column indicates LXP + TNCB-treated group. (c,d) Each value represents the mean ± S.E. from 6 mice (total 24 mice), NS, not significant, # p < 0.05, ## p < 0.01 compared with the non-TNCB-treated group. * p < 0.05 when compared with each group. Two-way ANOVA: time × group: F (24, 175) = 5.382, p < 0.0001, time: F (6, 175) = 34.22, p < 0.0001, group: F (4, 175) = 37.29, p < 0.0001, (a). One-way ANOVA: F (4, 25) = 111, p < 0.0001, (b); F (3, 20) = 10.65, p = 0.0002, (c); F (3, 20) = 10.87, p = 0.0002, (d).
Figure 3.
Effects of IL-31 and loxoprofen (LXP) on TNCB–applied the hot-plate test in BALB/c mice. (a) Effects of IL-31 and loxoprofen (LXP) on TNCB-applied hot-plate (45 °C) test in mice. The black line and column indicate the non-TNCB-treated group; the blue line indicates phosphate-buffered saline (PBS) + vehicle (carboxymethyl cellulose sodium, CMC, 10 mL/kg, oral)-treated group; the red line indicates IL-31 + CMC-treated group; the green line indicates PBS + LXP-treated group; the yellow line indicate IL-31 + LXP treated group. The red arrow indicates the AEM or TNCB application point. # p < 0.05, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. (b) Total antinociceptive index of LXP after 30–120 min (AUC30–120 min). The black line and column indicate the non-treated group; the blue column indicates the PBS + CMC-treated group; the green column indicates the PBS + LXP-treated group; the red column indicates the IL-31 + CMC treated-group; the yellow column indicates the IL-31 + LXP-treated group. Each value represents the mean ± standard error (S.E.) from 6 mice (total 54 mice). NS, not significant, ### p < 0.001 compared with the corresponding values in the saline + vehicle-treated group. * p < 0.05, *** p < 0.001 compared with the corresponding values in the vehicle + TNCB treated group; (c) Effects of IL-31 and LXP on TNCB-induced increasing cutaneous temperature (fever) after 24 h application of TNCB. (d) Effects of IL-31 and LXP on TNCB-induced increasing cutaneous weight (swelling) after 24 h application of TNCB. The blue column indicates saline + vehicle-treated group; the red column indicates vehicle + TNCB-treated group; the green column indicates IL-31 + TNCB-treated group; the yellow column indicates LXP + TNCB-treated group. (c,d) Each value represents the mean ± S.E. from 6 mice (total 24 mice), NS, not significant, # p < 0.05, ## p < 0.01 compared with the non-TNCB-treated group. * p < 0.05 when compared with each group. Two-way ANOVA: time × group: F (24, 175) = 5.382, p < 0.0001, time: F (6, 175) = 34.22, p < 0.0001, group: F (4, 175) = 37.29, p < 0.0001, (a). One-way ANOVA: F (4, 25) = 111, p < 0.0001, (b); F (3, 20) = 10.65, p = 0.0002, (c); F (3, 20) = 10.87, p = 0.0002, (d).

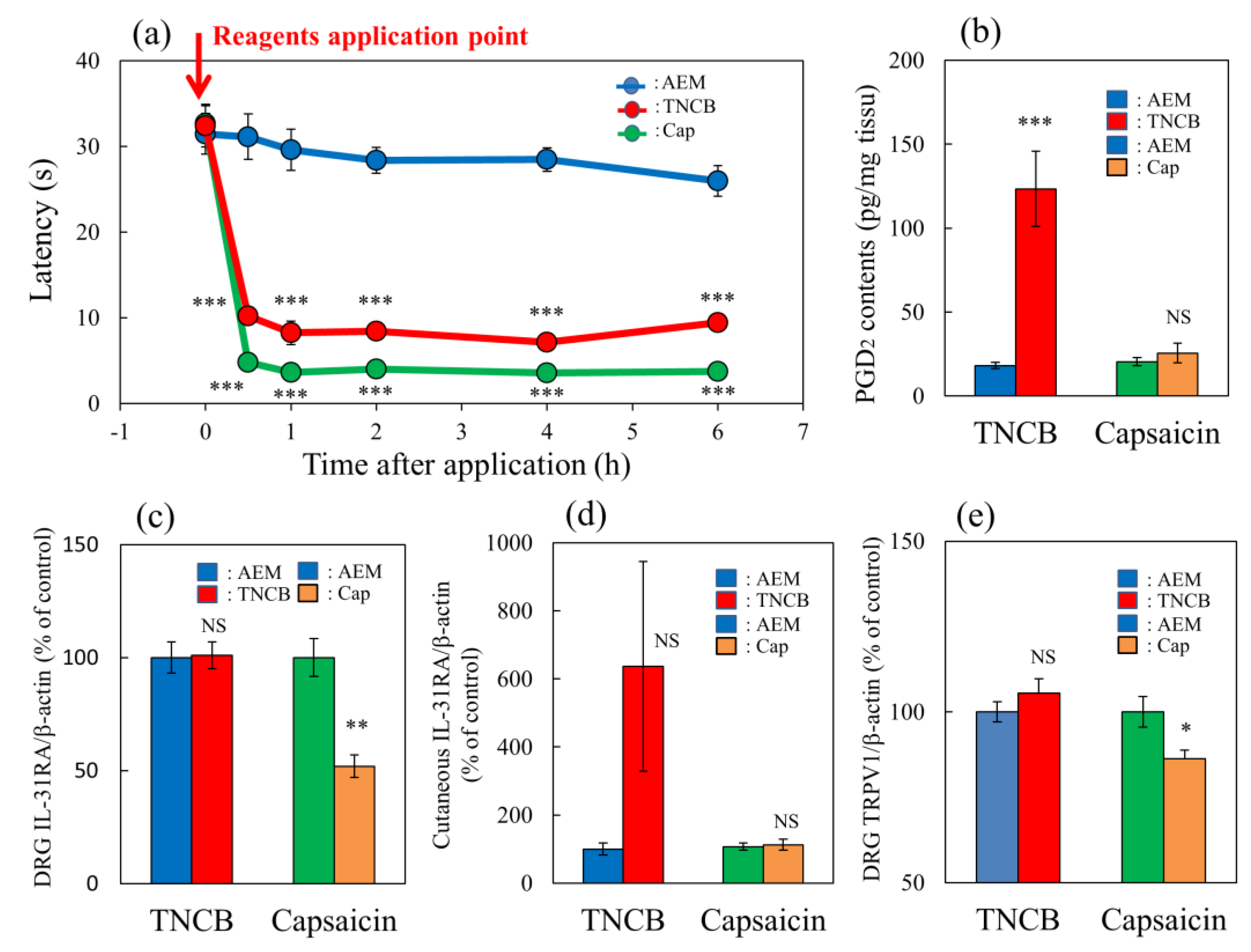
Figure 4.
Comparative effects of TNCB or capsaicin on latency in modified hot-plate test and several pain-related parameters in BALB/c mice. (a) Time-course changes of latency caused by topical application of TNCB (3%) or capsaicin (Cap, 1%) in modified hot-plate (45 °C) test. The red arrow indicates the vehicle (acetone-ethanol mixed liquor, AEM, 0.2 mL/site) or TNCB or capsaicin application point. The values represent the means ± standard error (S.E.) from 6 mice. *** p < 0.001 compared with the respective value of the AEM-treated group. (b) Effects of TNCB and capsaicin on cutaneous PGD2 contents 1 h after application. (c) Effects of TNCB or capsaicin on DRG neuronal IL-31RA mRNA expression 6 h after application. (d) Effects of TNCB or capsaicin on cutaneous IL-31 mRNA expression 1 h after application. € Effects of TNCB or capsaicin on DRG neuronal TRPV1 mRNA expression 6 h after application. The values represent the means ± standard error (S.E.) from 6 mice (total 42 mice). (a–d) NS, not significant, * p < 0.05, ** p < 0.01, *** p < 0.001 compared with the respective value of the AEM-treated group. Two-way ANOVA: time × group: F (10, 90) = 14.22, p < 0.0001, time: F (5, 90) = 74.75, p < 0.0001, group: F (2, 90) = 307.4, p < 0.0001, (a); group × treatment: F (1, 20) = 18.51, p = 0.0003, group: F (1, 20) = 22.51, p = 0.0001, treatment: F (1, 20) = 16.9, p = 0.0005, (b); group × treatment: F (1, 20) = 10.65, p = 0.0039, group: F (1, 20) = 9.817, p = 0.0052, treatment: F (1, 20) = 10.65, p = 0.0039, (c); group × treatment: F (1, 20) = 3.016, p = 0.0978, group: F (1, 20) = 3.014, p = 0.0979, treatment: F (1, 20) = 3.016, p = 0.0978, (d); group × treatment: F (1, 20) = 6.93, p = 0.0160, group: F (1, 20) = 1.273, p = 0.2725, treatment: F (1, 20) = 6.932, p = 0.0159, (e).
Figure 4.
Comparative effects of TNCB or capsaicin on latency in modified hot-plate test and several pain-related parameters in BALB/c mice. (a) Time-course changes of latency caused by topical application of TNCB (3%) or capsaicin (Cap, 1%) in modified hot-plate (45 °C) test. The red arrow indicates the vehicle (acetone-ethanol mixed liquor, AEM, 0.2 mL/site) or TNCB or capsaicin application point. The values represent the means ± standard error (S.E.) from 6 mice. *** p < 0.001 compared with the respective value of the AEM-treated group. (b) Effects of TNCB and capsaicin on cutaneous PGD2 contents 1 h after application. (c) Effects of TNCB or capsaicin on DRG neuronal IL-31RA mRNA expression 6 h after application. (d) Effects of TNCB or capsaicin on cutaneous IL-31 mRNA expression 1 h after application. € Effects of TNCB or capsaicin on DRG neuronal TRPV1 mRNA expression 6 h after application. The values represent the means ± standard error (S.E.) from 6 mice (total 42 mice). (a–d) NS, not significant, * p < 0.05, ** p < 0.01, *** p < 0.001 compared with the respective value of the AEM-treated group. Two-way ANOVA: time × group: F (10, 90) = 14.22, p < 0.0001, time: F (5, 90) = 74.75, p < 0.0001, group: F (2, 90) = 307.4, p < 0.0001, (a); group × treatment: F (1, 20) = 18.51, p = 0.0003, group: F (1, 20) = 22.51, p = 0.0001, treatment: F (1, 20) = 16.9, p = 0.0005, (b); group × treatment: F (1, 20) = 10.65, p = 0.0039, group: F (1, 20) = 9.817, p = 0.0052, treatment: F (1, 20) = 10.65, p = 0.0039, (c); group × treatment: F (1, 20) = 3.016, p = 0.0978, group: F (1, 20) = 3.014, p = 0.0979, treatment: F (1, 20) = 3.016, p = 0.0978, (d); group × treatment: F (1, 20) = 6.93, p = 0.0160, group: F (1, 20) = 1.273, p = 0.2725, treatment: F (1, 20) = 6.932, p = 0.0159, (e).

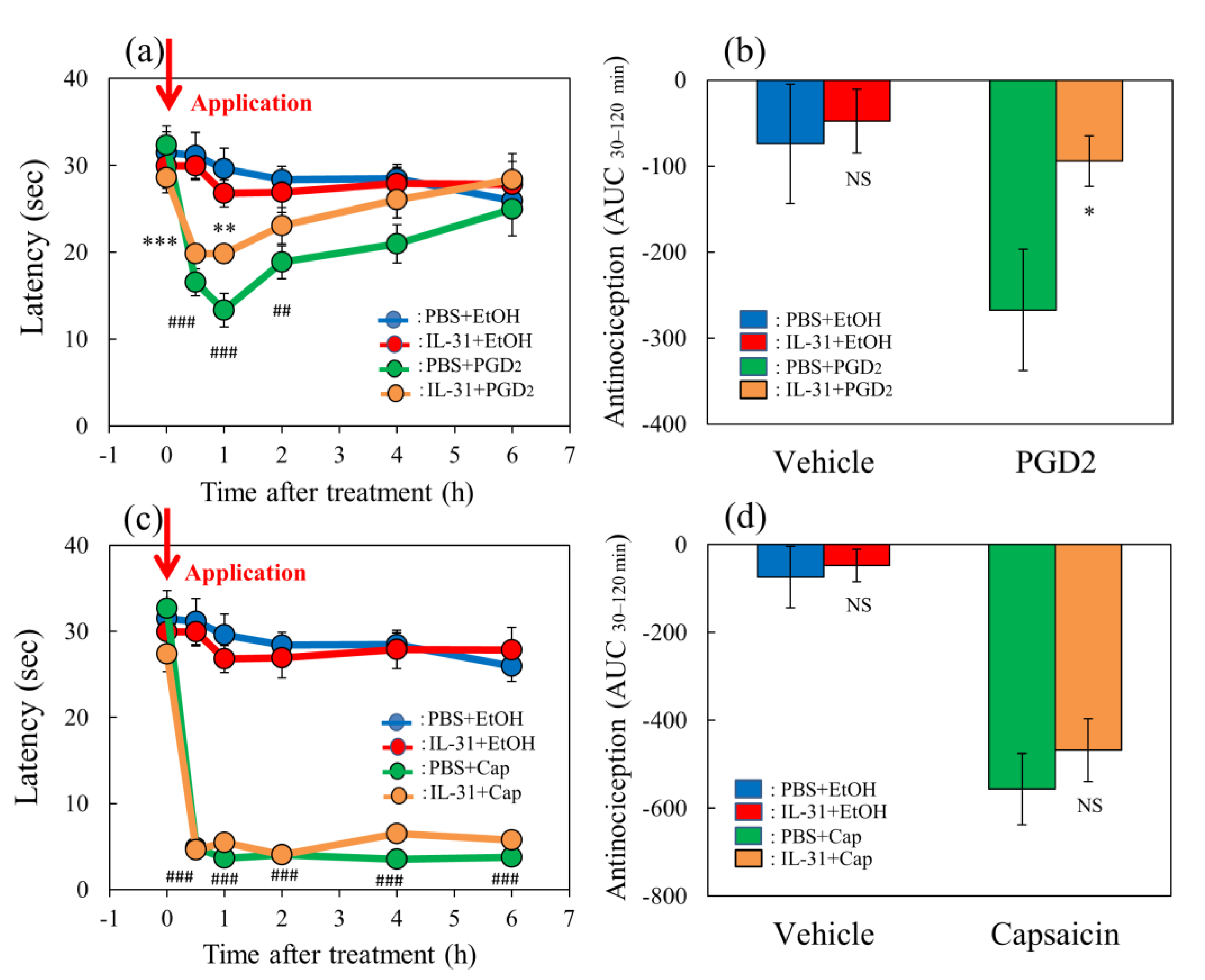
Figure 5.
Effects of IL-31 on PGD2- or capsaicin-applied hot-plate test in BALB/c mice. (a) Effect of IL-31 on PGD2-induced decreasing latency on hot–plate (45 °C) test. ** p < 0.01, *** p < 0.001 compared with the corresponding values in the vehicle (ethanol, EtOH, 0.2 mL/site)-treated group; ## p < 0.01, ### p < 0.001 compared with the corresponding values in the vehicle-treated group. (b) Total antinociceptive index from 30 to 120 min (AUC30–120 min) after PGD2 application. NS, not significant, * p < 0.05 compared with the corresponding values in the vehicle-treated group. (c) Effect of IL-31 on capsaicin (Cap)-induced decreasing latency on modified hot-plate (45 °C) test. ### p < 0.001 compared with the corresponding values in the vehicle-treated group. (d) After capsaicin application, the total antinociceptive index changes from 30 to 120 min (AUC30–120 min). The blue line indicates the vehicle (phosphate-buffered saline, PBS, intraperitoneal, every 12 h for 3 days) + EtOH-group; the red line indicates the IL-31-pretreated (50 μg/kg, intraperitoneal, every 12 h for 3 days) + EtOH-treated group; the green line indicates PBS + PGD2 (0.1%, 0.04 mL/each limb)- or capsaicin (3%, 0.04 mL/each limb)-treated group; the yellow line indicates IL-31 + PGD2- or capsaicin-treated group. The red arrow indicates the vehicle (ethanol, EtOH)- or PGD2 or capsaicin application point. Values represent the mean ± S.E. from 6 mice (total of 48 mice). NS, not significant, compared with the respective values of the vehicle-treated group. Three-way ANOVA: group × treatment × time: F (5, 120) = 0.885, p = 0.4934, group × treatment: F (1, 120) = 5.631, p = 0.0192, group: F (1, 120) = 47.782, p < 0.0001, treatment: F (1, 120) = 1.591, p = 0.2096, time: F (5, 120) = 7.123, p < 0.0001, (a); group × treatment × time: F (5, 126) = 0.824, p = 0.5347, group: F (1, 126) = 903.432, p < 0.0001, treatment: F (1, 126) = 0.315, p = 0.5759, time: F (5, 126) = 50.026, p < 0.0001, (c). Two-way ANOVA: group × treatment: F (1, 20) = 1.79, p = 0.1959, group: F (1, 20) = 3.317, p = 0.0835, treatment: F (1, 20) = 4.791, p = 0.0406, (b); group × treatment: F (1, 21) = 0.2122, p = 0.6498, group: F (1, 21) = 44.57, p < 0.0001, treatment: F (1, 21) = 0.727, p = 0.4035, (d).
Figure 5.
Effects of IL-31 on PGD2- or capsaicin-applied hot-plate test in BALB/c mice. (a) Effect of IL-31 on PGD2-induced decreasing latency on hot–plate (45 °C) test. ** p < 0.01, *** p < 0.001 compared with the corresponding values in the vehicle (ethanol, EtOH, 0.2 mL/site)-treated group; ## p < 0.01, ### p < 0.001 compared with the corresponding values in the vehicle-treated group. (b) Total antinociceptive index from 30 to 120 min (AUC30–120 min) after PGD2 application. NS, not significant, * p < 0.05 compared with the corresponding values in the vehicle-treated group. (c) Effect of IL-31 on capsaicin (Cap)-induced decreasing latency on modified hot-plate (45 °C) test. ### p < 0.001 compared with the corresponding values in the vehicle-treated group. (d) After capsaicin application, the total antinociceptive index changes from 30 to 120 min (AUC30–120 min). The blue line indicates the vehicle (phosphate-buffered saline, PBS, intraperitoneal, every 12 h for 3 days) + EtOH-group; the red line indicates the IL-31-pretreated (50 μg/kg, intraperitoneal, every 12 h for 3 days) + EtOH-treated group; the green line indicates PBS + PGD2 (0.1%, 0.04 mL/each limb)- or capsaicin (3%, 0.04 mL/each limb)-treated group; the yellow line indicates IL-31 + PGD2- or capsaicin-treated group. The red arrow indicates the vehicle (ethanol, EtOH)- or PGD2 or capsaicin application point. Values represent the mean ± S.E. from 6 mice (total of 48 mice). NS, not significant, compared with the respective values of the vehicle-treated group. Three-way ANOVA: group × treatment × time: F (5, 120) = 0.885, p = 0.4934, group × treatment: F (1, 120) = 5.631, p = 0.0192, group: F (1, 120) = 47.782, p < 0.0001, treatment: F (1, 120) = 1.591, p = 0.2096, time: F (5, 120) = 7.123, p < 0.0001, (a); group × treatment × time: F (5, 126) = 0.824, p = 0.5347, group: F (1, 126) = 903.432, p < 0.0001, treatment: F (1, 126) = 0.315, p = 0.5759, time: F (5, 126) = 50.026, p < 0.0001, (c). Two-way ANOVA: group × treatment: F (1, 20) = 1.79, p = 0.1959, group: F (1, 20) = 3.317, p = 0.0835, treatment: F (1, 20) = 4.791, p = 0.0406, (b); group × treatment: F (1, 21) = 0.2122, p = 0.6498, group: F (1, 21) = 44.57, p < 0.0001, treatment: F (1, 21) = 0.727, p = 0.4035, (d).

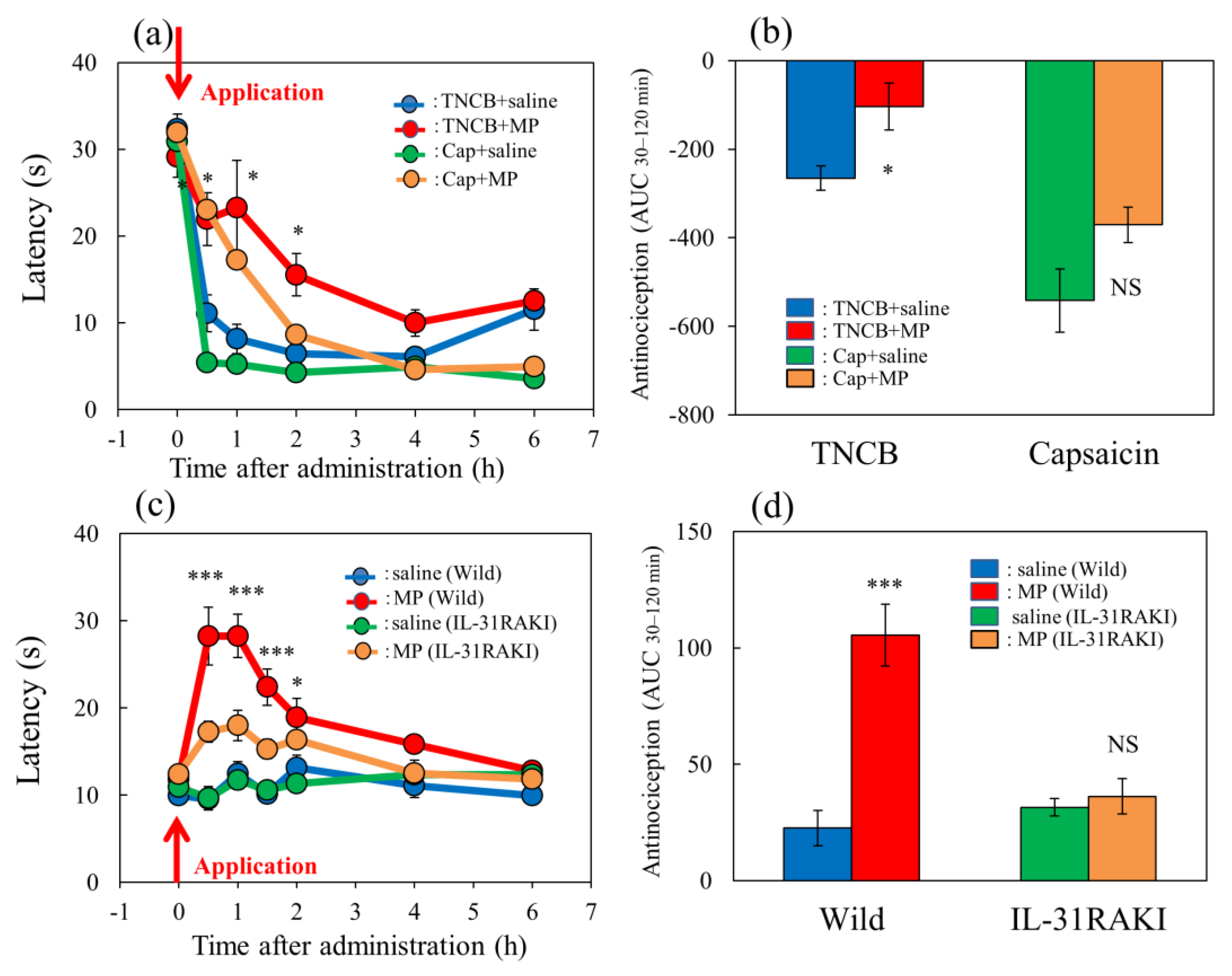
Figure 6.
Effect of morphine on TNCB- or capsaicin-applied hot-plate test in wild-type or IL-31RAKI mice. (a) Effect of morphine (3 mg/kg, subcutaneous) on TNCB- or capsaicin-applied hot-plate (45 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.) from 6 mice * p < 0.05 compared with the respective values in vehicle (saline)-treated mice. (b) Total antinociceptive index of morphine after 30 to 120 min (AUC30–120 min) as per the TNCB or capsaicin-treated hot-plate test. NS, not significant, * p < 0.05 compared with the respective values in vehicle-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + morphine-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + morphine-treated group. The red arrow indicates the TNCB or capsaicin application point. (c) Effect of morphine on conventional hot-plate (51 °C) test in wild-type (C57BL/6) and IL-31RAKI (C57BL/6 genetic background) mice. * p < 0.05, and *** p < 0.001 compared with the respective values in vehicle-treated mice. (d) Total antinociceptive index of morphine after 30 to 120 min (AUC30–120 min) of wild-type and IL-31RAKI mice. The blue line and column indicate the vehicle (saline)-treated group in wild-type mice; the red line and column indicates the morphine-treated group in wild-type mice; the green line and column indicates the saline-treated group in IL-31RAKI mice; the yellow line and column indicates the morphine-treated group in IL-31RAKI mice. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). *** p < 0.001 compared with the respective values in vehicle-treated mice. Three-way ANOVA: group × treatment × time: F (5, 120) = 1.275, p = 0.2792, group: F (1, 120) = 17.296, p < 0.0001, treatment: F (1, 120) = 49.027, p < 0.0001, time: F (5, 120) = 73.243, p < 0.0001, (a); group × treatment × time: F (7, 160) = 2.171, p = 0.0395, (c). Two-way ANOVA: group × treatment: F (1, 20) = 0.007463, p = 0.9320, group: F (1, 20) = 28.95, p < 0.0001, treatment: F (1, 20) = 10.85, p = 0.0036, (b); group × treatment: F (1, 20) = 19.79, p = 0.0002, group: F (1, 20) = 11.82, p = 0.0026, treatment: F (1, 20) = 24.87, p < 0.0001, (d).
Figure 6.
Effect of morphine on TNCB- or capsaicin-applied hot-plate test in wild-type or IL-31RAKI mice. (a) Effect of morphine (3 mg/kg, subcutaneous) on TNCB- or capsaicin-applied hot-plate (45 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.) from 6 mice * p < 0.05 compared with the respective values in vehicle (saline)-treated mice. (b) Total antinociceptive index of morphine after 30 to 120 min (AUC30–120 min) as per the TNCB or capsaicin-treated hot-plate test. NS, not significant, * p < 0.05 compared with the respective values in vehicle-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + morphine-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + morphine-treated group. The red arrow indicates the TNCB or capsaicin application point. (c) Effect of morphine on conventional hot-plate (51 °C) test in wild-type (C57BL/6) and IL-31RAKI (C57BL/6 genetic background) mice. * p < 0.05, and *** p < 0.001 compared with the respective values in vehicle-treated mice. (d) Total antinociceptive index of morphine after 30 to 120 min (AUC30–120 min) of wild-type and IL-31RAKI mice. The blue line and column indicate the vehicle (saline)-treated group in wild-type mice; the red line and column indicates the morphine-treated group in wild-type mice; the green line and column indicates the saline-treated group in IL-31RAKI mice; the yellow line and column indicates the morphine-treated group in IL-31RAKI mice. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). *** p < 0.001 compared with the respective values in vehicle-treated mice. Three-way ANOVA: group × treatment × time: F (5, 120) = 1.275, p = 0.2792, group: F (1, 120) = 17.296, p < 0.0001, treatment: F (1, 120) = 49.027, p < 0.0001, time: F (5, 120) = 73.243, p < 0.0001, (a); group × treatment × time: F (7, 160) = 2.171, p = 0.0395, (c). Two-way ANOVA: group × treatment: F (1, 20) = 0.007463, p = 0.9320, group: F (1, 20) = 28.95, p < 0.0001, treatment: F (1, 20) = 10.85, p = 0.0036, (b); group × treatment: F (1, 20) = 19.79, p = 0.0002, group: F (1, 20) = 11.82, p = 0.0026, treatment: F (1, 20) = 24.87, p < 0.0001, (d).

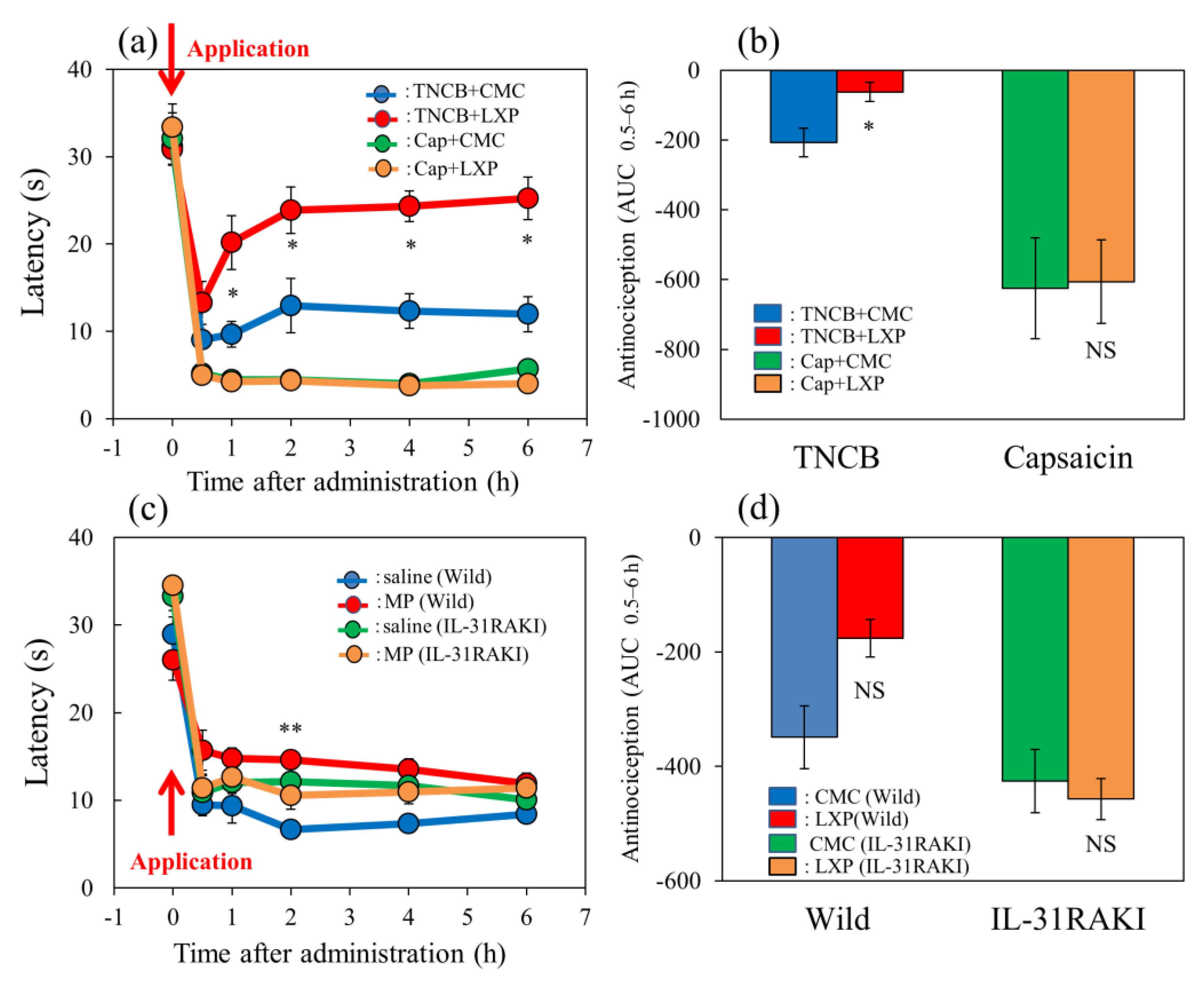
Figure 7.
Effect of loxoprofen on TNCB- or capsaicin-applied hot-plate test in wild-type and IL-31RAKI mice. (a) Effect of loxoprofen on TNCB- or capsaicin-applied hot-plate (45 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.). * p < 0.05 compared with the respective values in the vehicle (carboxymethyl cellulose sodium saline, CMC, 10 mL/kg, oral)-treated mice. (b) Total antinociceptive index of loxoprofen after 0.5 to 6 h (AUC0.5–6 h) as per the TNCB- or capsaicin-applied hot-plate test in BALB/c mice. Each value represents the mean ± standard error (S.E.) from 6 mice. NS, not significant, * p < 0.05 compared with the respective values in the CMC-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + loxoprofen-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + loxoprofen-treated group. (c) Effect of loxoprofen on TNCB-applied hot-plate (45 °C) test in wild-type (C57BL/6) and IL-31RAKI (C57BL/6 genetic background) mice. Each value represents the mean ± standard error (S.E.). ** p < 0.01 compared with the respective values in the CMC-treated mice. (d) Total antinociceptive index of loxoprofen (AUC0.5–6 h) of wild-type and IL-31RAKI mice. NS, not significant, compared with the respective values in vehicle-treated mice. The blue line indicates TNCB + CMC-and column indicate the vehicle (saline)-treated group in wild-type mice; the red line and column indicates TNCB +the loxoprofen-treated (15 mg/kg, oral) group in wild-type mice; the green line and column indicates capsaicin + CMCthe saline-treated group in IL-31RAKI mice; the yellow line and column indicates capsaicin +the loxoprofen -treated group in IL-31RAKI mice. The red arrow indicates the TNCB or capsaicin application point. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). Three-way ANOVA: group × treatment × time: F (5, 120) = 2.430, p = 0.0389, (a); group × treatment × time: F (5, 96) = 1.095, p = 0.3685, group: F (1, 96) = 6.646, p = 0.0115, treatment: F (1, 96) = 14.570, p = 0.0002, time: F (5, 96) = 102.550, p < 0.0001, (c). Two-way ANOVA: group × treatment: F (1, 20) = 0.4223, p = 0.5232, group: F (1, 20) = 24.67, p < 0.0001, treatment: F (1, 20) = 0.7201, p = 0.4062, (b); group × treatment: F (1, 16) = 4.854, p = 0.0426, group: F (1, 16) = 10.21, p = 0.0056, treatment: F (1, 16) = 3.69, p = 0.0727, (d).
Figure 7.
Effect of loxoprofen on TNCB- or capsaicin-applied hot-plate test in wild-type and IL-31RAKI mice. (a) Effect of loxoprofen on TNCB- or capsaicin-applied hot-plate (45 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.). * p < 0.05 compared with the respective values in the vehicle (carboxymethyl cellulose sodium saline, CMC, 10 mL/kg, oral)-treated mice. (b) Total antinociceptive index of loxoprofen after 0.5 to 6 h (AUC0.5–6 h) as per the TNCB- or capsaicin-applied hot-plate test in BALB/c mice. Each value represents the mean ± standard error (S.E.) from 6 mice. NS, not significant, * p < 0.05 compared with the respective values in the CMC-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + loxoprofen-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + loxoprofen-treated group. (c) Effect of loxoprofen on TNCB-applied hot-plate (45 °C) test in wild-type (C57BL/6) and IL-31RAKI (C57BL/6 genetic background) mice. Each value represents the mean ± standard error (S.E.). ** p < 0.01 compared with the respective values in the CMC-treated mice. (d) Total antinociceptive index of loxoprofen (AUC0.5–6 h) of wild-type and IL-31RAKI mice. NS, not significant, compared with the respective values in vehicle-treated mice. The blue line indicates TNCB + CMC-and column indicate the vehicle (saline)-treated group in wild-type mice; the red line and column indicates TNCB +the loxoprofen-treated (15 mg/kg, oral) group in wild-type mice; the green line and column indicates capsaicin + CMCthe saline-treated group in IL-31RAKI mice; the yellow line and column indicates capsaicin +the loxoprofen -treated group in IL-31RAKI mice. The red arrow indicates the TNCB or capsaicin application point. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). Three-way ANOVA: group × treatment × time: F (5, 120) = 2.430, p = 0.0389, (a); group × treatment × time: F (5, 96) = 1.095, p = 0.3685, group: F (1, 96) = 6.646, p = 0.0115, treatment: F (1, 96) = 14.570, p = 0.0002, time: F (5, 96) = 102.550, p < 0.0001, (c). Two-way ANOVA: group × treatment: F (1, 20) = 0.4223, p = 0.5232, group: F (1, 20) = 24.67, p < 0.0001, treatment: F (1, 20) = 0.7201, p = 0.4062, (b); group × treatment: F (1, 16) = 4.854, p = 0.0426, group: F (1, 16) = 10.21, p = 0.0056, treatment: F (1, 16) = 3.69, p = 0.0727, (d).

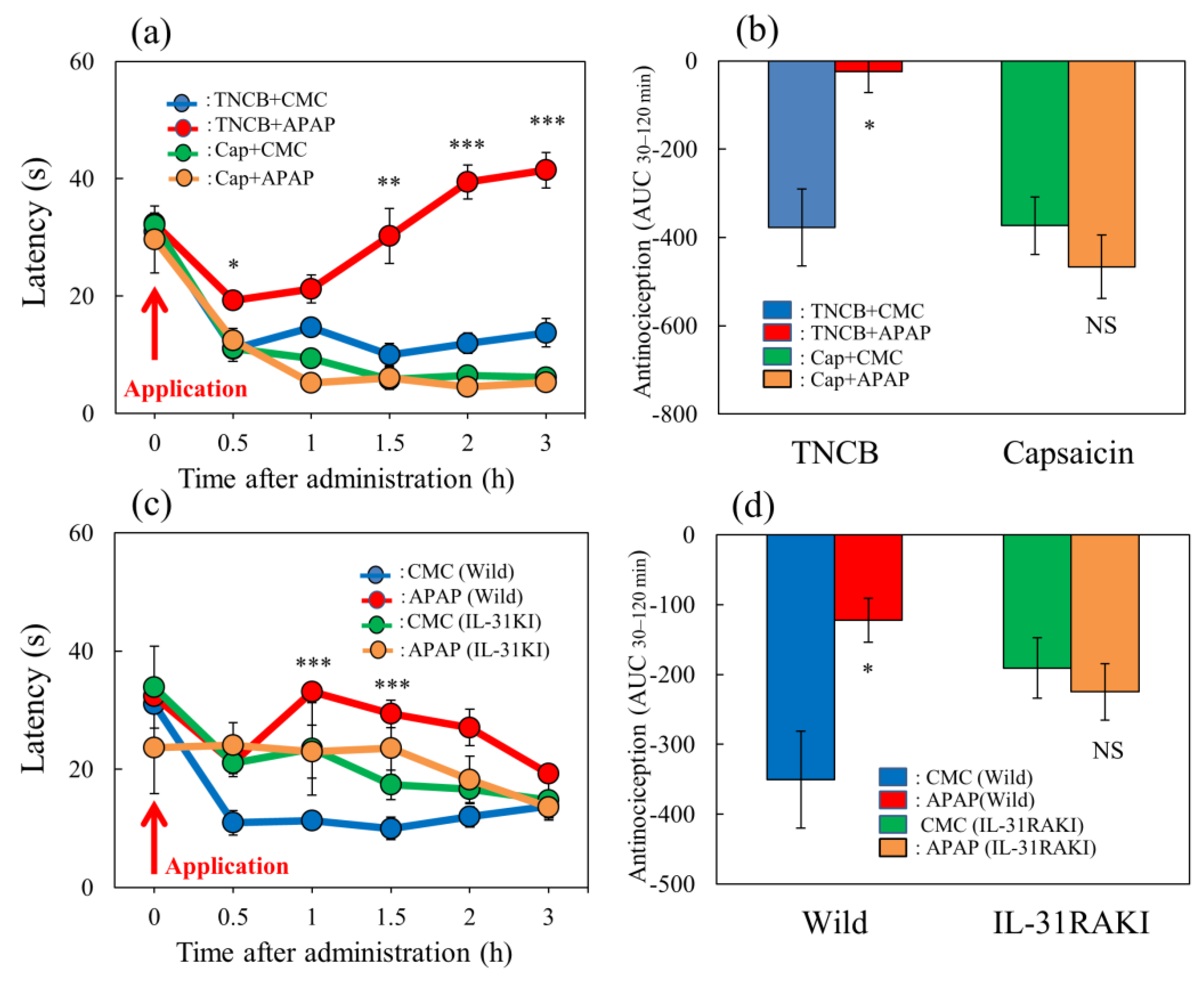
Figure 8.
Effect of acetaminophen on TNCB– or capsaicin–applied hot-plate (35 °C) test in wild-type and IL-31RAKI mice. (a) Effect of acetaminophen (APAP, 300 mg/kg, oral) on TNCB- or capsaicin-applied hot-plate (35 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.).* p < 0.05, ** p < 0.01, and *** p < 0.001 compared with the respective values in vehicle (carboxymethyl cellulose sodium, CMC, 10 mL/kg, oral)-treated mice. (b) Total antinociceptive index of acetaminophen after 30–120 min (AUC30–120 min) as per the TNCB- or capsaicin-applied hot-plate test in BALB/c mice. Each value represents the mean ± standard error (S.E.). NS, not significant, * p < 0.05 compared with the respective values in vehicle-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + acetaminophen-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + acetaminophen-treated group. (c) Effect of acetaminophen on TNCB-applied hot-plate (35 °C) test in wild-type (C57BL/6) mice and IL-31RAKI (C57BL/6 genetic background) mice. Each value represents the mean ± standard error (S.E.). *** p < 0.001 compared with the respective values in the CMC-treated mice. (d) Total antinociceptive index of acetaminophen after 30–120 min (AUC30–120 min) of wild-type and IL-31RAKI mice. The blue line indicatesand column indicate the TNCB + CMC-vehicle (saline)-treated group in wild-type mice; the red line and column indicates the TNCB + acetaminophenAPAP-treated group in wild-type mice; the green line and column indicates the capsaicin + CMCsaline-treated group in IL-31RAKI mice; the yellow line and column indicates the capsaicin + acetaminophenAPAP-treated group in IL-31RAKI mice. The red arrow indicates the TNCB or capsaicin application point. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). NS, not significant, * p < 0.05 compared with the respective values in the CMC-treated mice. Three-way ANOVA: group × treatment × time: F (5, 126) = 1.200, p = 0.3128, group: F (1, 126) = 381.730, p < 0.0001, treatment: F (1, 126) = 6.316, p = 0.0132, time: F (5, 126) = 44.926, p < 0.0001, (a); group × treatment × time: F (5, 120) = 0.248, p = 0.9400, group: F (1, 120) = 2.609, p = 0.1089, treatment: F (1, 120) = 21.475, p < 0.0001, time: F (5, 120) = 4.594, p = 0.0007, (c). Two-way ANOVA: group × treatment: F (1, 20) = 10.33, p = 0.0043, group: F (1, 20) = 9.935, p = 0.0050, treatment: F (1, 20) = 3.516, p = 0.0754, (b); group × treatment: F (1, 20) = 10.33, p = 0.0043, group: F (1, 20) = 9.935, p = 0.0050, treatment: F (1, 20) = 3.516, p = 0.0754, (d).
Figure 8.
Effect of acetaminophen on TNCB– or capsaicin–applied hot-plate (35 °C) test in wild-type and IL-31RAKI mice. (a) Effect of acetaminophen (APAP, 300 mg/kg, oral) on TNCB- or capsaicin-applied hot-plate (35 °C) test in BALB/c mice. Each value represents the mean ± standard error (S.E.).* p < 0.05, ** p < 0.01, and *** p < 0.001 compared with the respective values in vehicle (carboxymethyl cellulose sodium, CMC, 10 mL/kg, oral)-treated mice. (b) Total antinociceptive index of acetaminophen after 30–120 min (AUC30–120 min) as per the TNCB- or capsaicin-applied hot-plate test in BALB/c mice. Each value represents the mean ± standard error (S.E.). NS, not significant, * p < 0.05 compared with the respective values in vehicle-treated mice. The blue line and column indicate the TNCB + vehicle (saline)-treated group; the red line and column indicates the TNCB + acetaminophen-treated group; the green line and column indicates the capsaicin + saline-treated group; the yellow line and column indicates the capsaicin + acetaminophen-treated group. (c) Effect of acetaminophen on TNCB-applied hot-plate (35 °C) test in wild-type (C57BL/6) mice and IL-31RAKI (C57BL/6 genetic background) mice. Each value represents the mean ± standard error (S.E.). *** p < 0.001 compared with the respective values in the CMC-treated mice. (d) Total antinociceptive index of acetaminophen after 30–120 min (AUC30–120 min) of wild-type and IL-31RAKI mice. The blue line indicatesand column indicate the TNCB + CMC-vehicle (saline)-treated group in wild-type mice; the red line and column indicates the TNCB + acetaminophenAPAP-treated group in wild-type mice; the green line and column indicates the capsaicin + CMCsaline-treated group in IL-31RAKI mice; the yellow line and column indicates the capsaicin + acetaminophenAPAP-treated group in IL-31RAKI mice. The red arrow indicates the TNCB or capsaicin application point. Wild, C57BL/6 mice; IL-31RAKI, IL-31 receptor A-deficient mice (C57BL/6 genetic background). Each value represents the mean ± standard error (S.E.) from 6 mice (total 48 mice). NS, not significant, * p < 0.05 compared with the respective values in the CMC-treated mice. Three-way ANOVA: group × treatment × time: F (5, 126) = 1.200, p = 0.3128, group: F (1, 126) = 381.730, p < 0.0001, treatment: F (1, 126) = 6.316, p = 0.0132, time: F (5, 126) = 44.926, p < 0.0001, (a); group × treatment × time: F (5, 120) = 0.248, p = 0.9400, group: F (1, 120) = 2.609, p = 0.1089, treatment: F (1, 120) = 21.475, p < 0.0001, time: F (5, 120) = 4.594, p = 0.0007, (c). Two-way ANOVA: group × treatment: F (1, 20) = 10.33, p = 0.0043, group: F (1, 20) = 9.935, p = 0.0050, treatment: F (1, 20) = 3.516, p = 0.0754, (b); group × treatment: F (1, 20) = 10.33, p = 0.0043, group: F (1, 20) = 9.935, p = 0.0050, treatment: F (1, 20) = 3.516, p = 0.0754, (d).

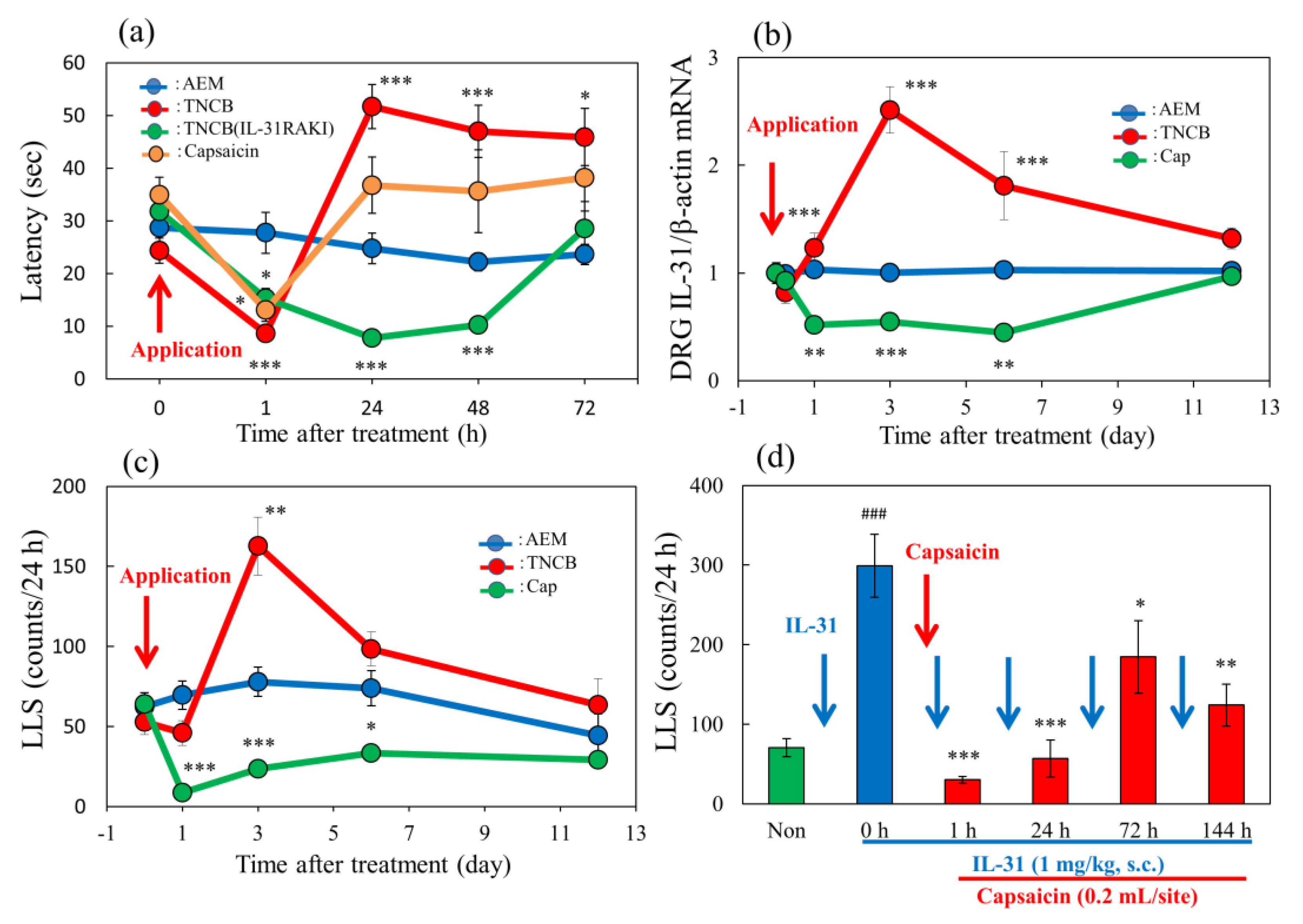
Figure 9.
Long–term changes after TNCB or capsaicin application on the sense of itch and pain. (a) Effects of topical application of TNCB or capsaicin on modified hot-plate (45 °C) test in BALB/c mice and IL-31RAKI mice. The blue line indicates the vehicle (acetone-ethanol mixed liquor, AEM)-treated mice; the red line indicates the TNCB-treated mice; the green line indicates the capsaicin-treated mice; the yellow line indicates the TNCB-treated IL-31RAKI mice. (b) Effects of topical application of TNCB or capsaicin on IL-31RA expression in the DRG. The blue line indicates AEM-treated mice; the red line indicates TNCB-treated mice; the green line indicates capsaicin-treated mice. (c) Effects of topical application of TNCB or capsaicin on itch-associated behavior counts (LLS counts/24 h). The blue line indicates AEM-treated mice; the red line indicates TNCB-treated mice; the green line indicates capsaicin-treated mice. The red arrow indicates the AEM or TNCB or capsaicin application point. Each value represents the mean ± standard error (S.E.) from 6 mice (total 54 mice). * p < 0.05, ** p < 0.01, and *** p < 0.001 compared with the respective values of vehicle (AEM)-treated group. (d) Time-course change of topical application of capsaicin on IL-31-induced LLS counts. The blue arrows indicate the IL-31 (1 mg/kg, intravenous) administration point. The red arrow indicates the capsaicin (1%, 0.2 mL/site) application point. Each value represents the mean ± standard error (S.E.) from 6 mice. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared with IL-31-induced LLS counts before capsaicin application. Two-way ANOVA: group × time: F (12, 100) = 7.899, p < 0.0001, group: F (3, 100) = 18.81, p < 0.0001, time: F (4, 100) = 12.45, p < 0.0001, (a); group × time: F (10, 90) = 15.2, p < 0.0001, group: F (2, 90) = 63.57, p < 0.0001, time: F (5, 90) = 6.516, p < 0.0001, (b); group × time: F (8, 75) = 8.204, p < 0.0001, group: F (2, 75) = 31.92, p < 0.0001, time: F (4, 75) = 9.405, p < 0.0001, (c). One-way ANOVA: F (5, 30) = 12.11, p < 0.0001, (d).
Figure 9.
Long–term changes after TNCB or capsaicin application on the sense of itch and pain. (a) Effects of topical application of TNCB or capsaicin on modified hot-plate (45 °C) test in BALB/c mice and IL-31RAKI mice. The blue line indicates the vehicle (acetone-ethanol mixed liquor, AEM)-treated mice; the red line indicates the TNCB-treated mice; the green line indicates the capsaicin-treated mice; the yellow line indicates the TNCB-treated IL-31RAKI mice. (b) Effects of topical application of TNCB or capsaicin on IL-31RA expression in the DRG. The blue line indicates AEM-treated mice; the red line indicates TNCB-treated mice; the green line indicates capsaicin-treated mice. (c) Effects of topical application of TNCB or capsaicin on itch-associated behavior counts (LLS counts/24 h). The blue line indicates AEM-treated mice; the red line indicates TNCB-treated mice; the green line indicates capsaicin-treated mice. The red arrow indicates the AEM or TNCB or capsaicin application point. Each value represents the mean ± standard error (S.E.) from 6 mice (total 54 mice). * p < 0.05, ** p < 0.01, and *** p < 0.001 compared with the respective values of vehicle (AEM)-treated group. (d) Time-course change of topical application of capsaicin on IL-31-induced LLS counts. The blue arrows indicate the IL-31 (1 mg/kg, intravenous) administration point. The red arrow indicates the capsaicin (1%, 0.2 mL/site) application point. Each value represents the mean ± standard error (S.E.) from 6 mice. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared with IL-31-induced LLS counts before capsaicin application. Two-way ANOVA: group × time: F (12, 100) = 7.899, p < 0.0001, group: F (3, 100) = 18.81, p < 0.0001, time: F (4, 100) = 12.45, p < 0.0001, (a); group × time: F (10, 90) = 15.2, p < 0.0001, group: F (2, 90) = 63.57, p < 0.0001, time: F (5, 90) = 6.516, p < 0.0001, (b); group × time: F (8, 75) = 8.204, p < 0.0001, group: F (2, 75) = 31.92, p < 0.0001, time: F (4, 75) = 9.405, p < 0.0001, (c). One-way ANOVA: F (5, 30) = 12.11, p < 0.0001, (d).

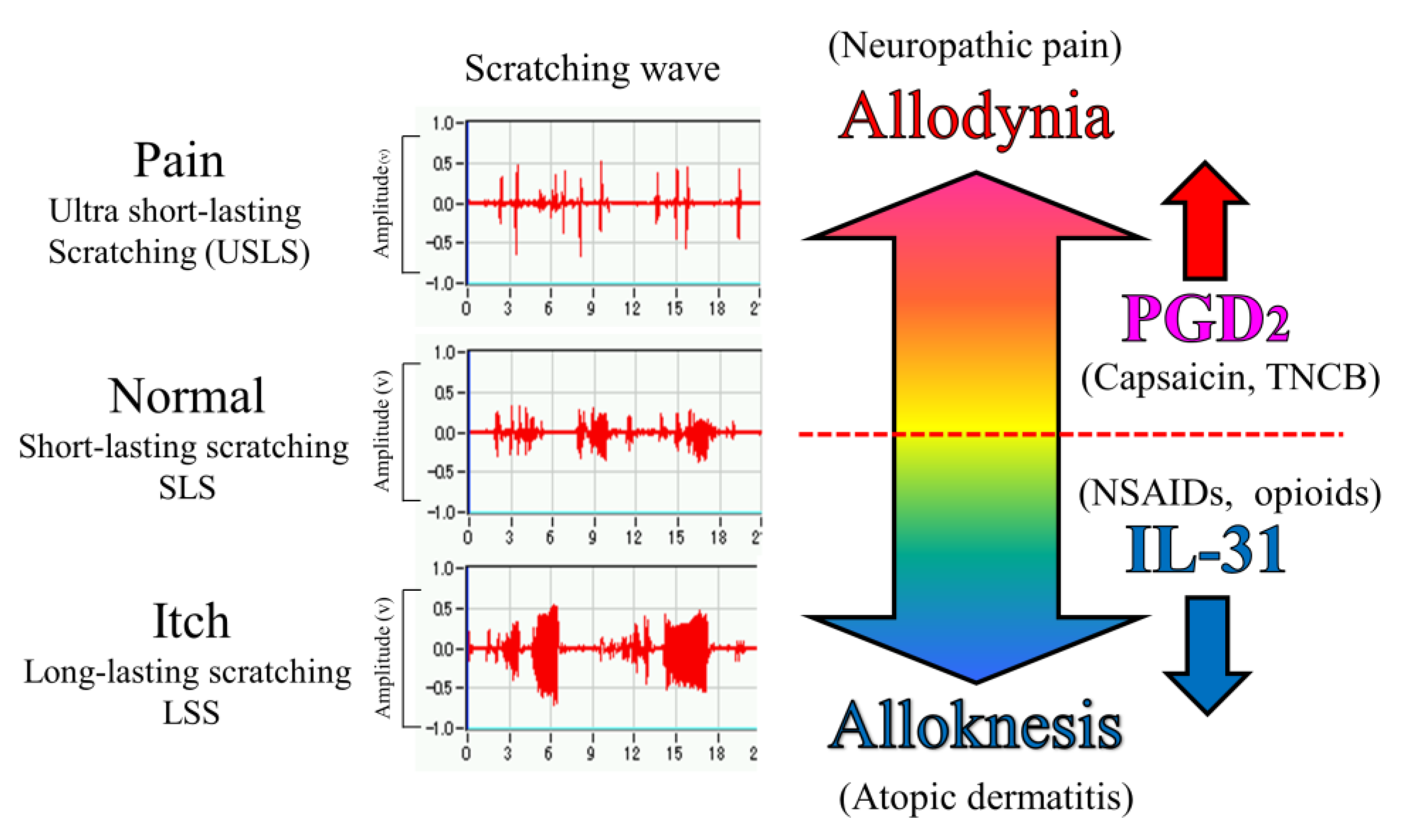
Figure 10.
Schematic diagram of the putative roles of IL-31 and PGD2 in the regulation of the sensation of cutaneous itch and pain in mice. We evaluated a wave pattern of scratching behavior from the movement of the hind leg of the mouse, objectively. In normal conditions, the mouse elicits 0.3–1.0 s lasting scratching behavior (SLS) caused by several stimulants. Under itchy conditions, the mouse elicits over 1.0 s lasting scratching behavior (LLS) caused by IL-31. If this stimulation continues for a long time, it may become atopic dermatitis. Under painful conditions, the mouse elicits below 0.3 s lasting scratching behavior (USLS) caused by chronic inflammatory stimulants. If this stimulation continues for a long time, it may become chronic neuropathic pain. IL-31 can change non-selective stimulation into itch stimulation. In contrast, PGD2 can change non-selective stimulation into pain stimulation transmitted by the primary nerves of C-fibers and by second-order nerves and spinothalamic tract neurons in the spinal cord. This suggests that IL-31 and PGD2 regulate the perception of sense (pain or itch) through their mutual functional antagonism.
Figure 10.
Schematic diagram of the putative roles of IL-31 and PGD2 in the regulation of the sensation of cutaneous itch and pain in mice. We evaluated a wave pattern of scratching behavior from the movement of the hind leg of the mouse, objectively. In normal conditions, the mouse elicits 0.3–1.0 s lasting scratching behavior (SLS) caused by several stimulants. Under itchy conditions, the mouse elicits over 1.0 s lasting scratching behavior (LLS) caused by IL-31. If this stimulation continues for a long time, it may become atopic dermatitis. Under painful conditions, the mouse elicits below 0.3 s lasting scratching behavior (USLS) caused by chronic inflammatory stimulants. If this stimulation continues for a long time, it may become chronic neuropathic pain. IL-31 can change non-selective stimulation into itch stimulation. In contrast, PGD2 can change non-selective stimulation into pain stimulation transmitted by the primary nerves of C-fibers and by second-order nerves and spinothalamic tract neurons in the spinal cord. This suggests that IL-31 and PGD2 regulate the perception of sense (pain or itch) through their mutual functional antagonism.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









