1. Introduction
Acrossocheilus fasciatus (Steindachner, 1892), which belongs to Cypriniformes, Cyprinidae, Barbinae,
Acrossocheilus, is a lukewarm water fish. It is a commercial fish that is primarily found in mountain streams and rivers south of the Yangtze River in China [
1]. In recent years, the natural resources of
A. fasciatus have declined significantly due to weather, water pollution, excessive and illegal fishing, and other factors.
A. fasciatus has been bred and propagated in captivity in an effort to preserve and replenish natural resources [
2]. It has been demonstrated that the
A. fasciatus can survive in a water temperature range of approximately 12–30 °C [
3], which may be an excellent model for studying the response mechanism of stream fish to temperature.
The extreme temperature caused by global climate change constitutes a huge challenge to the
A. fasciatus breeding industry. Temperature is one of the most significant environmental elements impacting fish life activities since it affects growth, development, reproduction, behavior, and metabolism [
4,
5]. A cohabitation model suggests that variations in water temperature may affect pathogen transfer and disease development in fish farming, which likewise faces major illness issues brought on by pathogenic bacteria and stress. Because variations in water temperature may impact the spread of disease and pathogens in a cohabitation model [
6,
7], resulting in reduced temperature adaptation. Environmental stress, especially temperature, can disrupt the organism’s internal balance and adversely affect its biological functions. Fish immune systems can be impacted by low temperatures, which can impair their ability to perform certain immunological functions and reduce fish survival [
8]. Fish may experience altered protein synthesis and reduced glucose metabolism at high temperatures [
9]. Fish’s metabolic rate, energy balance, and, consequently, their energy needs are impacted by temperature changes [
10]. Fish can adjust to changes in ambient temperature by engaging in specific physiological processes. However, different fish have varying degrees of temperature adaptability [
11]. Fish are capable of adapting to environmental temperature by modifying their physiological activity within a specific temperature range, which has become a popular research topic. For instance, when exposed to temperatures that are higher than those that are suitable for fish, fish will stabilize their metabolic rate to ensure a sufficient supply of oxygen to their tissues. Exposure to higher temperatures can also cause compensatory changes in the body’s energy reserve concentration [
12], glycolysis, and enzyme activity in the mitochondria [
13]. The peripheral nervous system and temperature receptors in the hypothalamus, as well as the central nervous system, all perceive environmental information at low temperatures [
14].
Transcriptome refers to the sum of all RNAs transcribed by specific cells or tissues at a certain functional state or developmental stage, RNA sequencing (RNA-seq) is a method of transcriptome analysis using deep sequencing techniques that can be used to identify important physiological pathways in organisms that respond to a variety of conditions, and it is widely used to monitor alterations in the transcriptome of targeted tissues, including new transcripts, splice junction and gene annotations [
15,
16]. High-throughput transcriptomics has revolutionized the field of transcriptome research by providing powerful screening tools and has been widely used in molecular, genetic, and other research fields [
17]. Simon A. Wentworth analyzed the transcriptome sequences information of the
Pimephales promelas (Cyprinidae) in the low-temperature group (5 °C) and high-temperature group (22 °C) during 30 days of culture using high-throughput sequencing and obtained that the expression of 194 specific transcripts was changed. It was found that lower temperature alters the immune system of freshwater teleost fish, leading to genome-wide upregulation of innate immunity and downregulation of adaptive immunity [
18]. Wei Zhang found 2534 genes differentially expressed in the liver and 1622 genes differentially expressed in the brain by transcriptomic analysis of the liver and brain in grass carp under high-temperature stress; according to the analysis of the KEGG results, significant differences in the expression of genes involved in metabolic and immune pathways, such as the cAMP signaling pathway, apoptosis, calcium signaling pathway, lipid metabolism, protein processing in the endoplasmic reticulum and the peroxisome proliferator-activated receptor signaling pathway [
19]. Since specific responses of different tissues were different under heat stress, many heat stress-related studies used different tissues of fish, such as the liver, muscle, heart, kidney, brain, and gill. In which skeletal muscle exhibited coordinated inhibition of genes constituting sarcomere structure (e.g., actin, myosins, tropomyosin) and those involved in muscle contraction (e.g., parvalbumins and troponins), which can indicate the structural remodeling of muscle tissue in cold [
20]. Therefore, muscle tissues were used to compare and analyze the transcriptome information of acrossocheilus in response to high and low temperatures.
In this study, the transcriptome information of the muscle of A. fasciatus was analyzed by Illumina sequencing technology, and the genes differentially expressed under long-term stress at different temperatures and their related metabolic pathways were screened. The results of this study can provide fundamental data to elucidate the mechanisms of temperature acclimation in A. fasciatus and can expand our understanding of the challenges of environmental adaptation in A. fasciatus.
4. Materials and Methods
4.1. Experimental Animals
The present study was conducted under the permission and supervision of the Institutional Animal Care and Use Committee of Zhejiang Ocean University. The approval number is 20210207.
A. fasciatus were obtained from the breeding base in Ta-pieh Mountains, Anhui, China. The fish were divided in the first stage of the experiment and were temporarily cultured in a tank for 2 weeks at a water temperature of 17 ± 1 °C; 450 healthy individuals with symmetrical body shapes, no obvious mechanical or artificial damage on the body surface, and similar and active individuals were randomly selected as the experimental materials. The average body length was 0.57 ± 0.16 cm, and the average body weight was 4.08 ± 0.40 g. During the experiment, fish were fed at 9:00 am and 18:00 pm (feed weighed at 2% of body weight), fish growth was checked, dead and injured fish were retrieved, the water temperature was measured with a thermometer and recorded, fish excrement and other foreign objects were removed from the tanks, and ventilation and a quiet and comfortable environment were maintained at all times.
4.2. Temperature Stress Experiment
Five temperature groups were designed for this experiment, and experimental fish were cultured for 60 days at different temperatures of 12 °C (T12), 16 °C (T16), 20 °C (control, T20), 24 °C (T24) and 28 °C (T28) in circulating water, 30 fish per tank, and each group was performed in triplicate. The experimental culture tank of each temperature group was placed in 5 temperature boxes (Shanghai Yiheng LRH-150 biochemical incubator, Shanghai, China). Each temperature group was cultured at a rising and cooling rate of 1 °C/24 h until the temperature reached the experimental water temperature adaptation for a week and maintained for 60 days. The feeding was stopped 1 d before sampling, and 10 fish were randomly selected from each tank, with a total of 30 fish in each treatment group, dissected on ice, take the back muscles of the fish and the muscle tissues were immediately snap frozen in liquid nitrogen and then stored at −80 °C in the freezer.
4.3. Total RNA Extraction and Illumina Sequencing
There were 3 parallel samples per temperature group, using the RNA extraction kit (TaKaRa, Code: No. 9767) according to the manufacturer’s instructions, RNA was extracted from each group of muscle tissue, and each group of RNA was mixed in equal proportions for a total of 15 samples. The Agilent 2100 bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) was used to measure each RNA concentration; RNA 28S/18S ≥ 1.5 and RNA integrity number >7.0 were selected for further analysis. RNA availability and integrity by 1% agarose gel electrophoresis, and then high throughput sequencing via Illumina Novaseq6000 (Berry Genomics Co., Ltd., Beijing, China).
4.4. Sequencing Data Quality Control, Assembly, and Gene Function Annotation
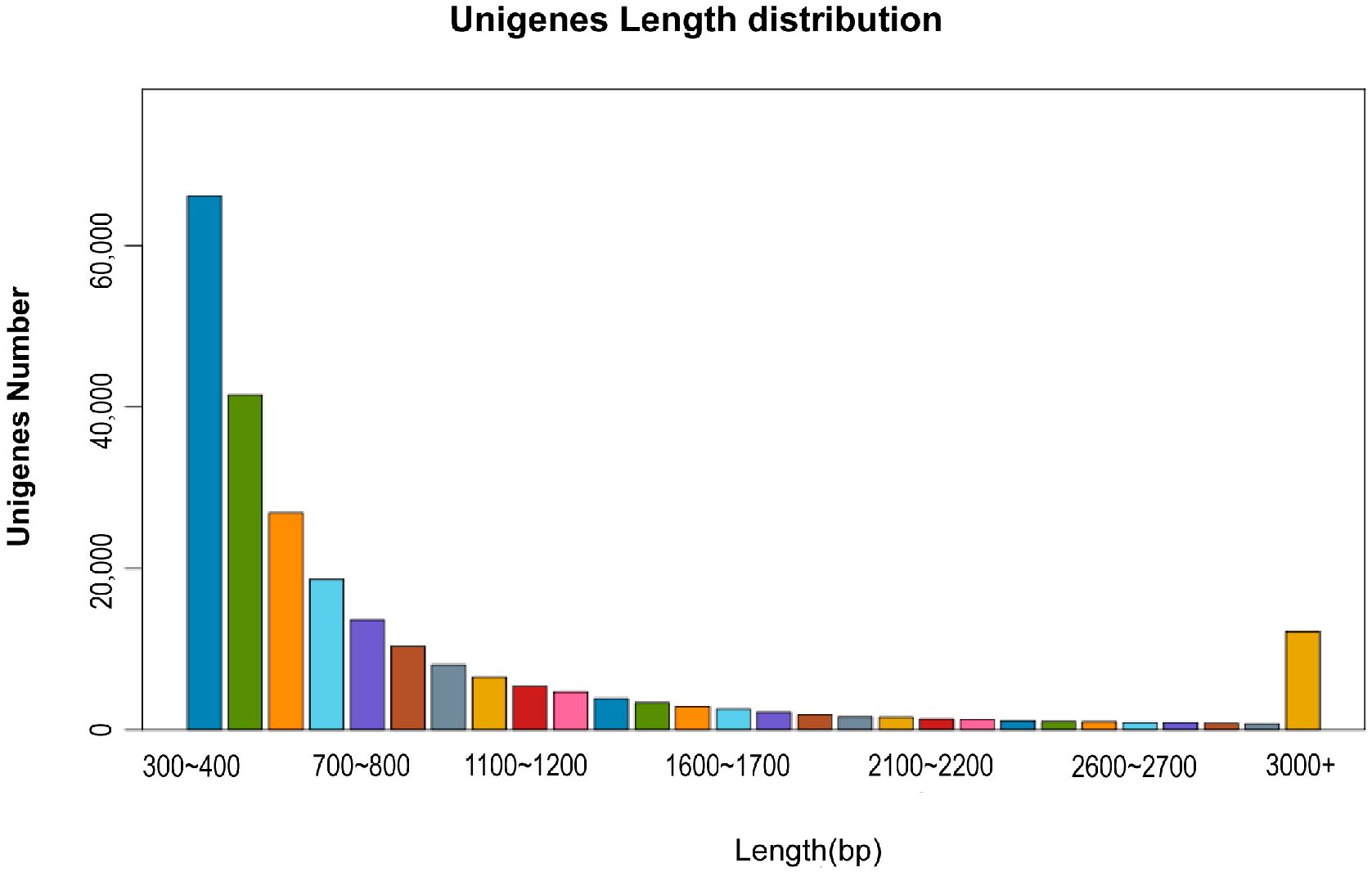
The sequenced raw reads were filtered to remove spliced, duplicated, low-quality sequences, resulting in clean reads. The sequences were assembled using Trinity 2.8.5 to obtain transcripts, and the Unigene of the corresponding species was obtained based on the component information, and the Unigene was compared with NR, NT, Swissprot, Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and Eukaryotic Ortholog Groups (KOG)/Clusters of Orthologous Groups of proteins (COG) databases respectively to obtain Unigene annotation information.
4.5. Differentially Expressed Gene Analysis, Functional and Pathway Enrichment
Using the transcriptome assembled by Trinity as the reference sequence. The clean reads of each sample were compared back to the reference sequence using bowtie2 2.3.2 software, the RSEM counted the results of the bowtie2 comparison and further obtained the number of read counts of each sample compared to each Unigene and converted them to the number of bases per thousand (FPKM) compared to the transcript per million compared fragments. The edgeR 3.3.3 software was used to analyze the differential expression of genes in each sample and calculate the
p-value and padj values of DEGs; padj is the corrected
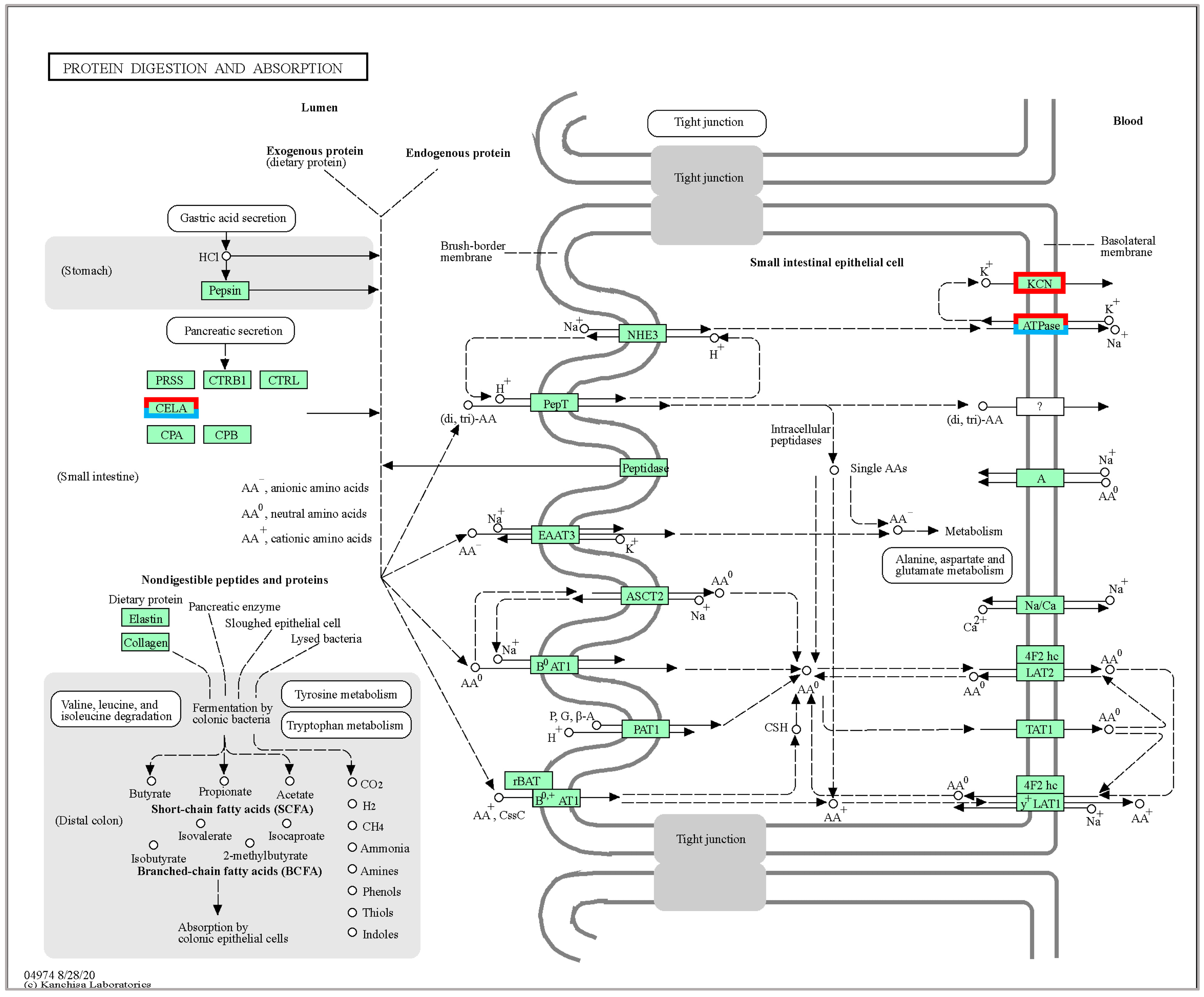
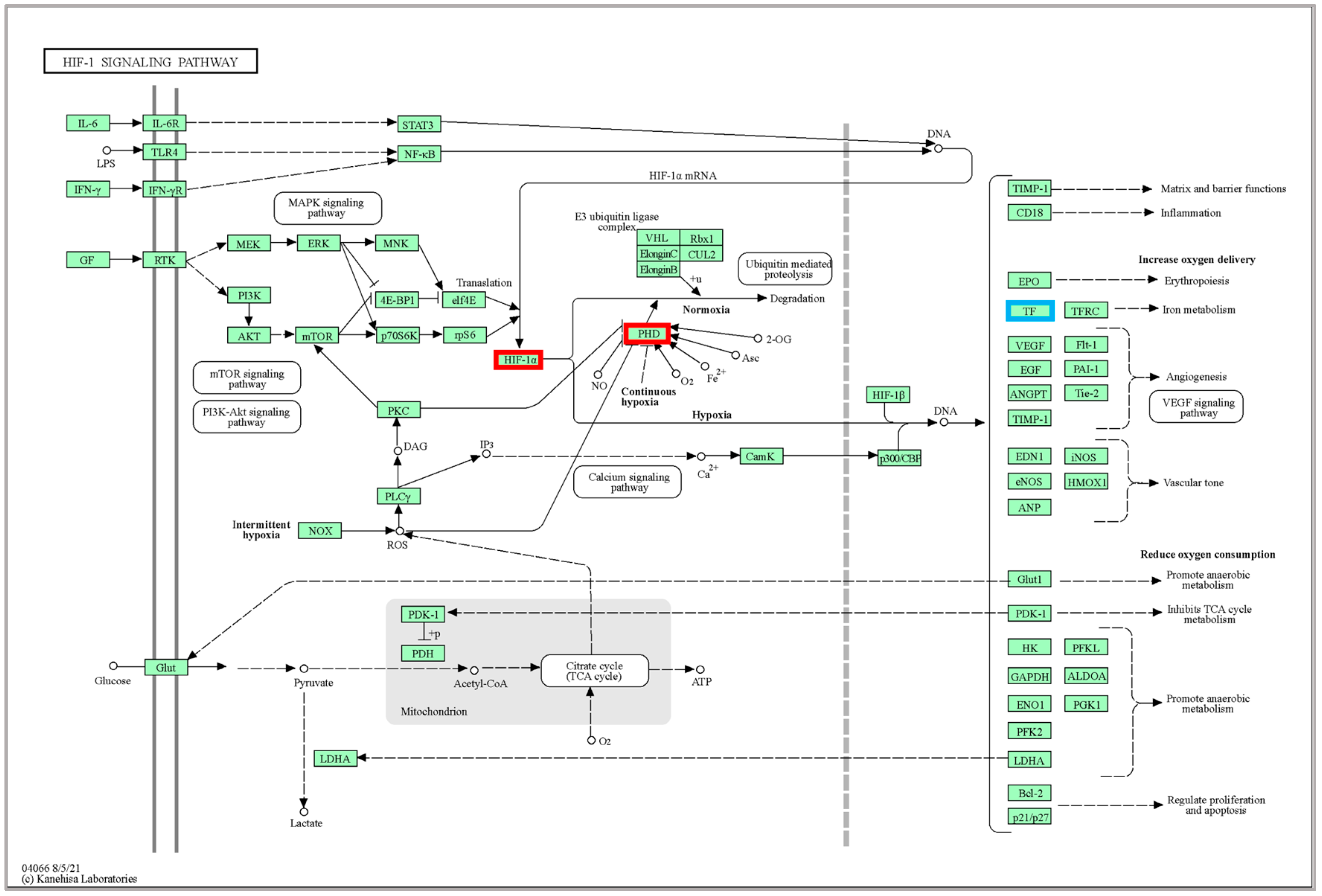
p-value. The smaller the padj value, the more significant the gene expression difference. To control the false positive rate, the padj value combined with Fold Change was required to screen for differential genes, and the screening conditions are as follows: padj < 0.05 and |log2FoldChange| > 1, and cluster analysis was performed on the DEGs obtained from different temperature culture. To further understand the function of DEGs at different temperatures, KEGG enrichment analysis using KOBAS 3.0 software [
46] was used to classify DEGs in different temperature groups, and the significance of differential gene enrichment in each pathway entry was calculated by a hypergeometric distribution test. Each functional module had a
p-value, and the false discovery rate (FDR) correction for the
p-value, the function of FDR ≤ 0.05, was set as a significant enrichment.
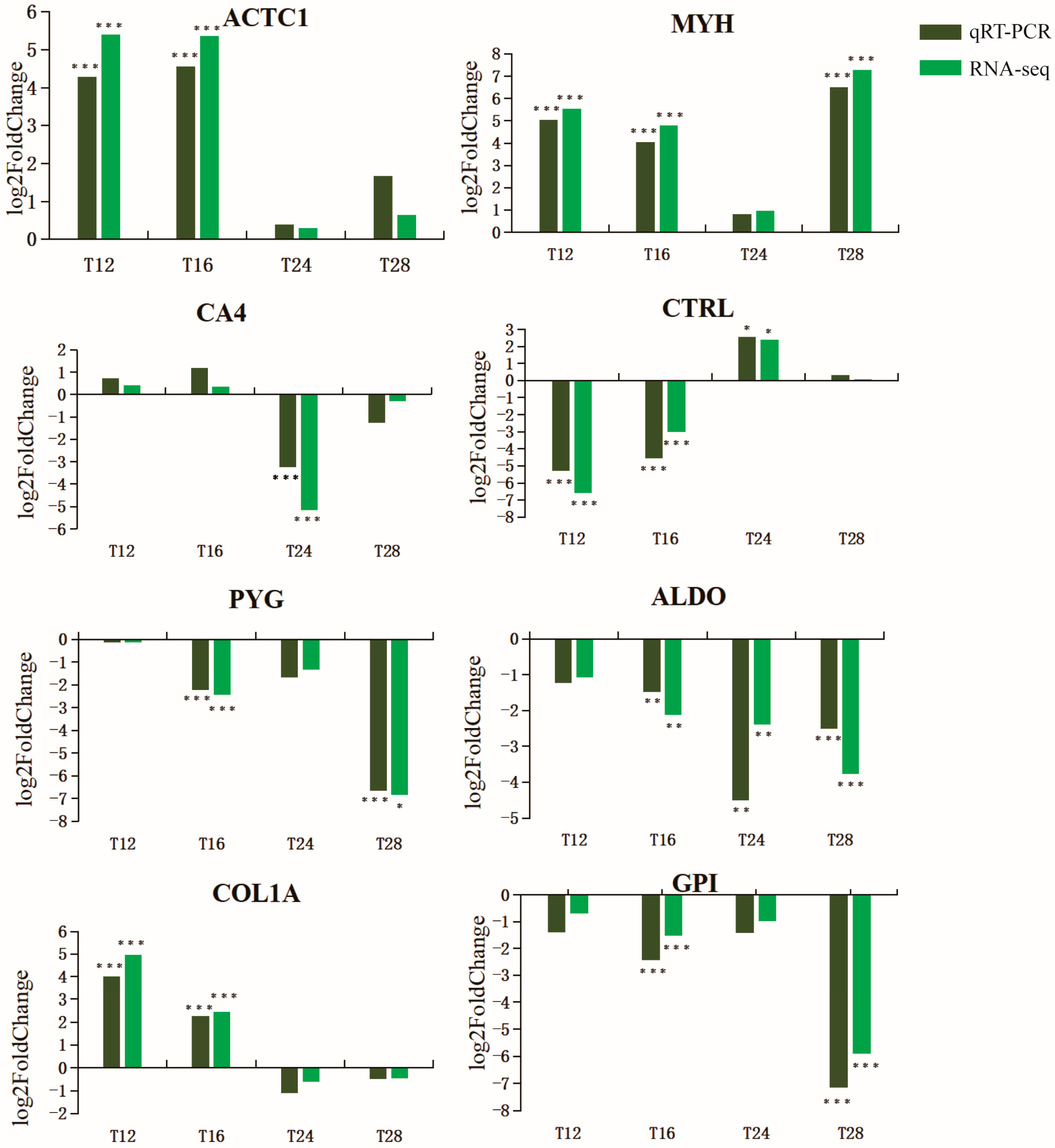
4.6. Quantitative Real-Time PCR
We used PrimeScript
TM RT Master Mix kit (TaKaRa, Kusatsu, Japan, Code: RR036A) to reverse transcribe the total RNA to obtain the cDNA template used in the qRT-PCR experiment. According to the results of the standard curve, samples were diluted 10-fold in nuclease-free water as templates for qRT-PCR. According to the manufacturer’s instructions, qRT-PCR experiments using TB Green
® Premix Ex Taq
TM II kit (TaKaRa, Code: RR820A). A 20 µL reaction system was amplified, including 10 µL of TB Green Premix Ex TaqII (2×), 2 µL of cDNA template, 0.4 µL of ROX Reference Dye (50×), 0.8µL of PCR Forward Primer (10 µM), 0.8µL of PCR Reverse Primer (10 µM) and 6 µL of RNase-free water. The amplification processes consisted of a holding stage of 30 s at 95 °C, followed by 40 cycles of 5 s at 95 °C and 30 s at 60 °C. Three parallel experiments on each cDNA template were performed to reduce the error of the experimental results. Primer Premier 6 [
47] software was used to design gene-specific primers with β-actin as an internal reference gene (
Table 6) and sent to Sunya Biotechnology Co., Ltd., Nanjing, China for synthesis. The relative expression of each of the 8 genes was analyzed using the comparative cycle threshold (2
−ΔΔCT) method (ΔCT = CTtarget gene − CTreference gene, ΔΔCT = ΔCTtreatment −ΔCTcontrol). SPSS 19.0 [
48] was used for statistical analysis, and the one-way ANOVA and
t-test were used to determine the significance of differences.
p-values < 0.05 were considered statistically significant.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}