
Cryotherapy Attenuates Inflammation via the lncRNA SNHG1/miR-9-5p/NFKB1 Regulatory Axis in Periodontal Ligament Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cryotherapy Reduces LPS-Induced Inflammatory Response in hPDLCs
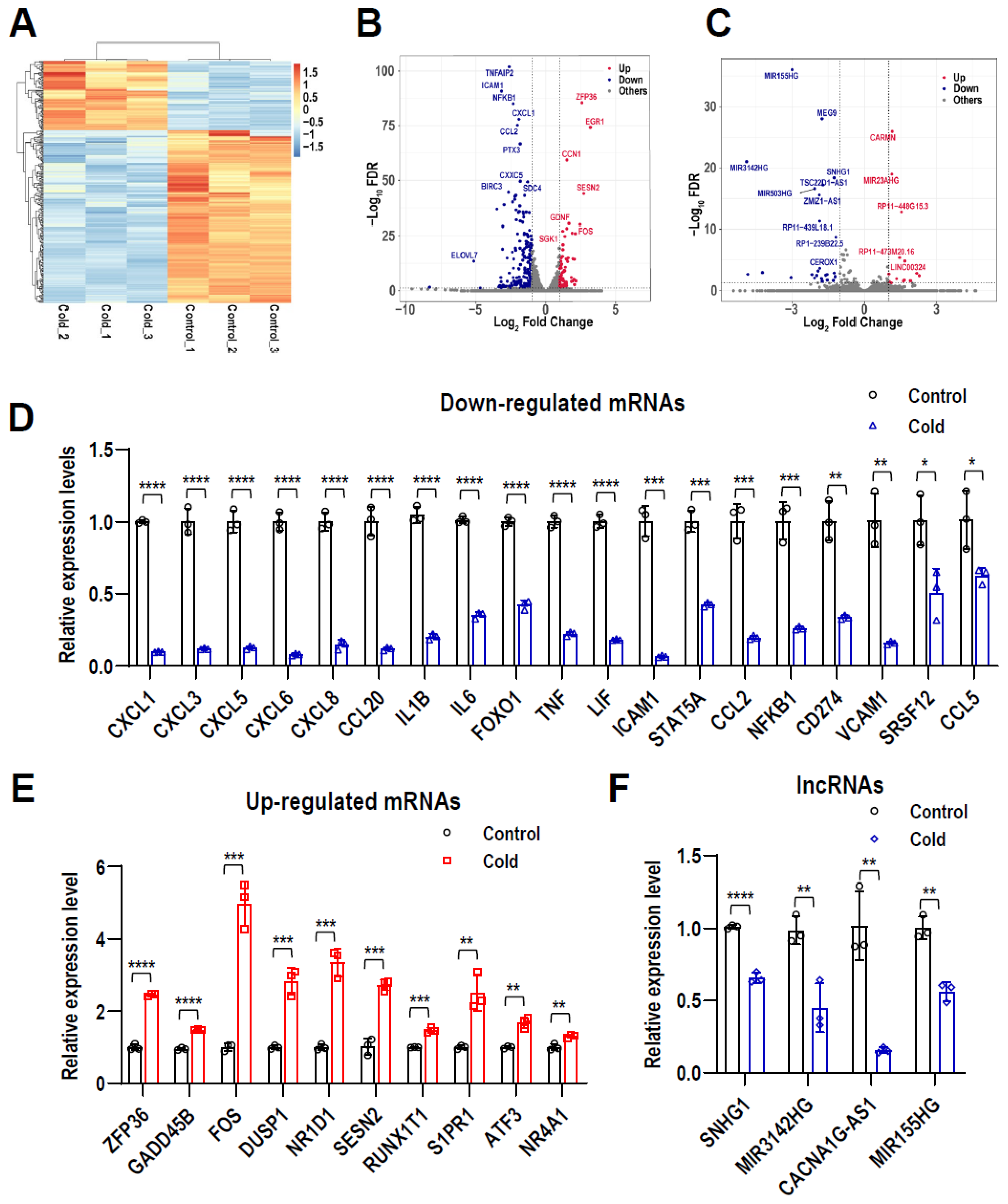
2.2. Profiles of Differentially Expressed lncRNAs and mRNAs in LPS-Stimulated hPDLCs after Cryotherapy
2.3. GO Functional Enrichment Analysis of DEGs
2.4. KEGG Enrichment Analysis and GSEA
2.5. Protein–Protein Interaction Network Analysis
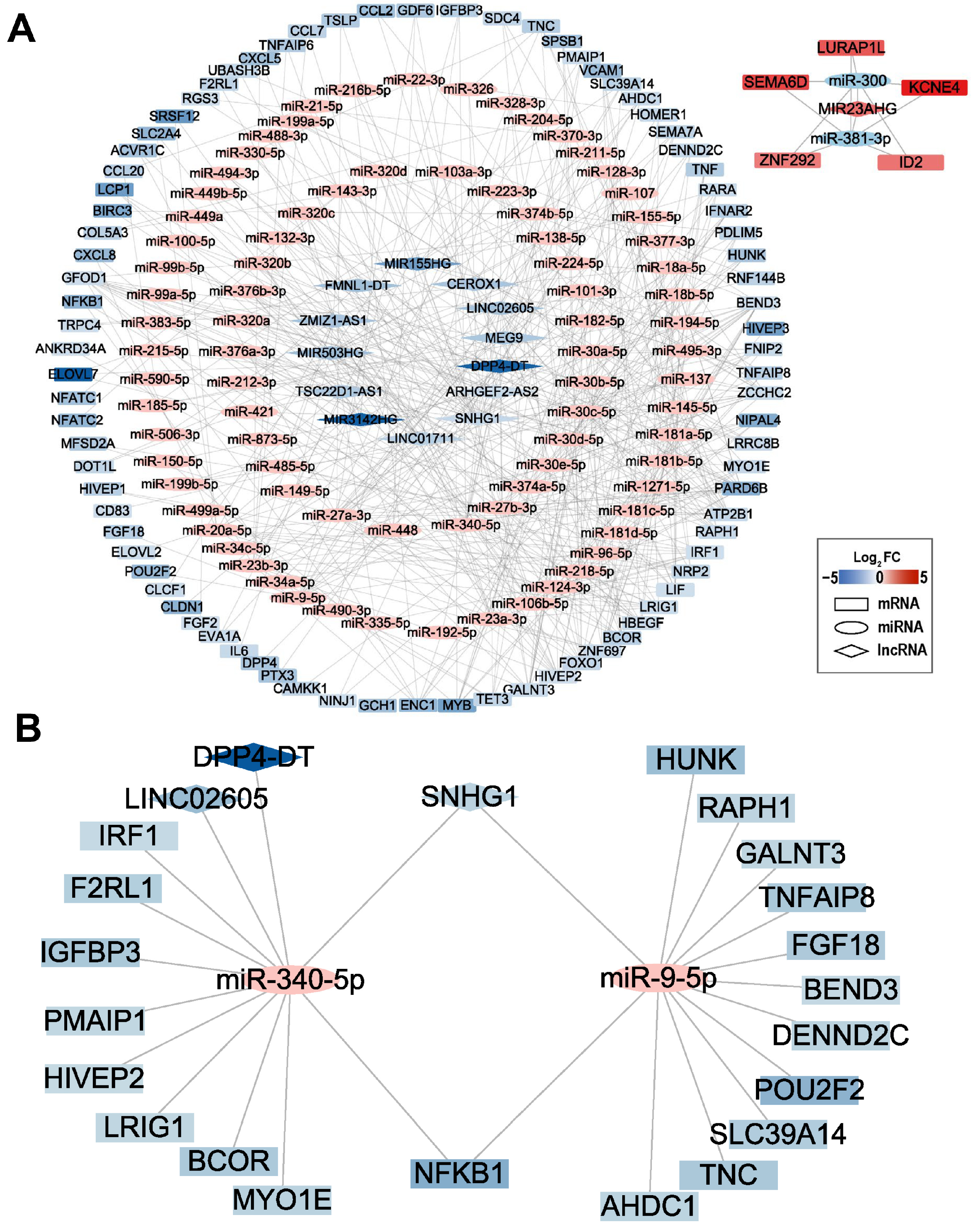
2.6. lncRNA-miRNA-mRNA Network Analysis
2.7. lncRNA SNHG1 Promotes LPS-Induced Inflammatory Response in hPDLCs
2.8. Cryotherapy Reduces Inflammation by Regulating the lncRNA SNHG1/miR-9-5p/NFKB1 Axis
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. RNA Extraction, Reverse Transcription PCR (RT-PCR), and Quantitative RT-PCR (qRT-PCR)
4.3. RNA Sequencing and DEG Identification
4.4. Functional and Pathway Enrichment Analyses of the DEGs
4.5. Protein–Protein Interaction Network Analysis
4.6. lncRNA-miRNA-mRNA Network Analysis
4.7. RNA Interference (RNAi) and Transfection
4.8. Plasmids and Transfection
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bleakley, C.M.; McDonough, S.M.; MacAuley, D.C.; Bjordal, J. Cryotherapy for acute ankle sprains: A randomised controlled study of two different icing protocols. Br. J. Sports Med. 2006, 40, 700–705; discussion 705. [Google Scholar] [CrossRef]
- Bala, M.M.; Riemsma, R.P.; Wolff, R.; Pedziwiatr, M.; Mitus, J.W.; Storman, D.; Swierz, M.J.; Kleijnen, J. Cryotherapy for liver metastases. Cochrane Database Syst. Rev. 2019, 7, Cd009058. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Risk, M.C.; Goldfarb, R.; Reddy, B.; Coles, B.; Dahm, P. Primary cryotherapy for localised or locally advanced prostate cancer. Cochrane Database Syst. Rev. 2018, 5, Cd005010. [Google Scholar] [CrossRef]
- Forouzanfar, T.; Sabelis, A.; Ausems, S.; Baart, J.A.; van der Waal, I. Effect of ice compression on pain after mandibular third molar surgery: A single-blind, randomized controlled trial. Int. J. Oral Maxillofac. Surg. 2008, 37, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Laureano Filho, J.R.; de Oliveira e Silva, E.D.; Batista, C.I.; Gouveia, F.M. The influence of cryotherapy on reduction of swelling, pain and trismus after third-molar extraction: A preliminary study. J. Am. Dent. Assoc. 2005, 136, 774–778; quiz 807. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, V.K.; Fernandes, D.; Navarro, C.M.; Giro, E.M.; de Almeida, L.Y.; León, J.E.; Bufalino, A. Cryotherapy for localized juvenile spongiotic gingival hyperplasia: Preliminary findings on two cases. Int. J. Paediatr. Dent. 2017, 27, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.; Glenny, A.M.; Worthington, H.V.; Littlewood, A.; Clarkson, J.E.; McCabe, M.G. Interventions for preventing oral mucositis in patients with cancer receiving treatment: Oral cryotherapy. Cochrane Database Syst. Rev. 2015, 2015, Cd011552. [Google Scholar] [CrossRef] [Green Version]
- Kakehashi, S.; Stanley, H.R.; Fitzgerald, R.J. The effects of surgical exposures of dental pulps in germ-free and conventional laboratory rats. Oral Surg. Oral Med. Oral Pathol. 1965, 20, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.N. Apical periodontitis: A dynamic encounter between root canal infection and host response. Periodontology 2000 1997, 13, 121–148. [Google Scholar] [CrossRef]
- Rossi-Fedele, G.; Rödig, T. Effectiveness of root canal irrigation and dressing for the treatment of apical periodontitis: A systematic review and meta-analysis of clinical trials. Int. Endod. J. 2022; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.N. On the causes of persistent apical periodontitis: A review. Int. Endod. J. 2006, 39, 249–281. [Google Scholar] [CrossRef] [PubMed]
- Sathorn, C.; Parashos, P.; Messer, H. The prevalence of postoperative pain and flare-up in single- and multiple-visit endodontic treatment: A systematic review. Int. Endod. J. 2008, 41, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Vera, J.; Ochoa, J.; Romero, M.; Vazquez-Carcaño, M.; Ramos-Gregorio, C.O.; Aguilar, R.R.; Cruz, A.; Sleiman, P.; Arias, A. Intracanal Cryotherapy Reduces Postoperative Pain in Teeth with Symptomatic Apical Periodontitis: A Randomized Multicenter Clinical Trial. J. Endod. 2018, 44, 4–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadaf, D.; Ahmad, M.Z.; Onakpoya, I.J. Effectiveness of Intracanal Cryotherapy in Root Canal Therapy: A Systematic Review and Meta-analysis of Randomized Clinical Trials. J. Endod. 2020, 46, 1811–1823.e1. [Google Scholar] [CrossRef]
- Vieyra, J.P.; Enriquez, E.J.J.; Acosta, F.O.; Guardado, J.A. Reduction of postendodontic pain after one-visit root canal treatment using three irrigating regimens with different temperature. Niger. J. Clin. Pract. 2019, 22, 34–40. [Google Scholar]
- Keskin, C.; Ozdemir, O.; Uzun, I.; Guler, B. Effect of intracanal cryotherapy on pain after single-visit root canal treatment. Aust. Endod. J. 2017, 43, 83–88. [Google Scholar] [CrossRef]
- Gundogdu, E.C.; Arslan, H. Effects of Various Cryotherapy Applications on Postoperative Pain in Molar Teeth with Symptomatic Apical Periodontitis: A Preliminary Randomized Prospective Clinical Trial. J. Endod. 2018, 44, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Vera, J.; Ochoa-Rivera, J.; Vazquez-Carcaño, M.; Romero, M.; Arias, A.; Sleiman, P. Effect of Intracanal Cryotherapy on Reducing Root Surface Temperature. J. Endod. 2015, 41, 1884–1887. [Google Scholar] [CrossRef]
- Zhang, R.; Niu, Z.; Liu, J.; Dang, X.; Feng, H.; Sun, J.; Pan, L.; Peng, Z. LncRNA SNHG1 promotes sepsis-induced myocardial injury by inhibiting Bcl-2 expression via DNMT1. J. Cell. Mol. Med. 2022, 26, 3648–3658. [Google Scholar] [CrossRef]
- Cao, B.; Wang, T.; Qu, Q.; Kang, T.; Yang, Q. Long Noncoding RNA SNHG1 Promotes Neuroinflammation in Parkinson’s Disease via Regulating miR-7/NLRP3 Pathway. Neuroscience 2018, 388, 118–127. [Google Scholar] [CrossRef]
- Hu, C.; Li, J.; Tan, Y.; Liu, Y.; Bai, C.; Gao, J.; Zhao, S.; Yao, M.; Lu, X.; Qiu, L.; et al. Tanreqing Injection Attenuates Macrophage Activation and the Inflammatory Response via the lncRNA-SNHG1/HMGB1 Axis in Lipopolysaccharide-Induced Acute Lung Injury. Front. Immunol. 2022, 13, 820718. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Du, J.; Liu, J.; Hou, K.; Ji, H.; Ma, S.; Ba, Y.; Chen, L.; Xie, R.; Hu, S. FtMt promotes glioma tumorigenesis and angiogenesis via lncRNA SNHG1/miR-9-5p axis. Cell. Signal. 2020, 75, 109749. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Jing, L.; Zhao, X.; Zhu, H.; Teng, J. Down-regulation of taurine-up-regulated gene 1 attenuates inflammation by sponging miR-9-5p via targeting NF-κB1/p50 in multiple sclerosis. Life Sci. 2019, 233, 116731. [Google Scholar] [CrossRef]
- Braz-Silva, P.H.; Bergamini, M.L.; Mardegan, A.P.; De Rosa, C.S.; Hasseus, B.; Jonasson, P. Inflammatory profile of chronic apical periodontitis: A literature review. Acta Odontol. Scand. 2019, 77, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Wan, Q.; Cheng, L.; Xu, R. Mechanisms of bone remodeling and therapeutic strategies in chronic apical periodontitis. Front. Cell. Infect. Microbiol. 2022, 12, 908859. [Google Scholar] [CrossRef]
- Silva, T.A.; Garlet, G.P.; Fukada, S.Y.; Silva, J.S.; Cunha, F.Q. Chemokines in oral inflammatory diseases: Apical periodontitis and periodontal disease. J. Dent. Res. 2007, 86, 306–319. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Guan, X.; He, Y.; Wei, Z.; Shi, C.; Li, Y.; Zhao, R.; Pan, L.; Han, Y.; Hou, T.; Yang, J. Crosstalk between Wnt/β-catenin signaling and NF-κB signaling contributes to apical periodontitis. Int. Immunopharmacol. 2021, 98, 107843. [Google Scholar] [CrossRef]
- Shen, Z.; Silva, R.M. MicroRNAs: Emerging players in apical periodontitis. J. Appl. Oral Sci. Rev. FOB 2021, 29, e20201058. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Wang, W.C.; Lin, P.Y.; Huang, C.P.; Chen, C.Y.; Chen, Y.K. The roles of autophagy and hypoxia in human inflammatory periapical lesions. Int. Endod. J. 2018, 51 (Suppl. 2), e125–e145. [Google Scholar] [CrossRef] [Green Version]
- Georgiou, A.C.; van der Waal, S.V.; Buijs, M.J.; Crielaard, W.; Zaura, E.; Brandt, B.W. Host-microbiome interactions in apical periodontitis: The endodontic microbiome in relation to circulatory immunologic markers. Int. Endod. J. 2023, 56, 748–764. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.; Liu, M.; Guo, J.; Jia, R. Cryotherapy Attenuates Inflammation via the lncRNA SNHG1/miR-9-5p/NFKB1 Regulatory Axis in Periodontal Ligament Cells. Int. J. Mol. Sci. 2023, 24, 12097. https://doi.org/10.3390/ijms241512097
Lin C, Liu M, Guo J, Jia R. Cryotherapy Attenuates Inflammation via the lncRNA SNHG1/miR-9-5p/NFKB1 Regulatory Axis in Periodontal Ligament Cells. International Journal of Molecular Sciences. 2023; 24(15):12097. https://doi.org/10.3390/ijms241512097
Chicago/Turabian StyleLin, Can, Miaomiao Liu, Jihua Guo, and Rong Jia. 2023. "Cryotherapy Attenuates Inflammation via the lncRNA SNHG1/miR-9-5p/NFKB1 Regulatory Axis in Periodontal Ligament Cells" International Journal of Molecular Sciences 24, no. 15: 12097. https://doi.org/10.3390/ijms241512097