
Metalloporphyrins Reduce Proteinuria in Podocyte Immune Injury: The Role of Metal and Porphyrin Moieties
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
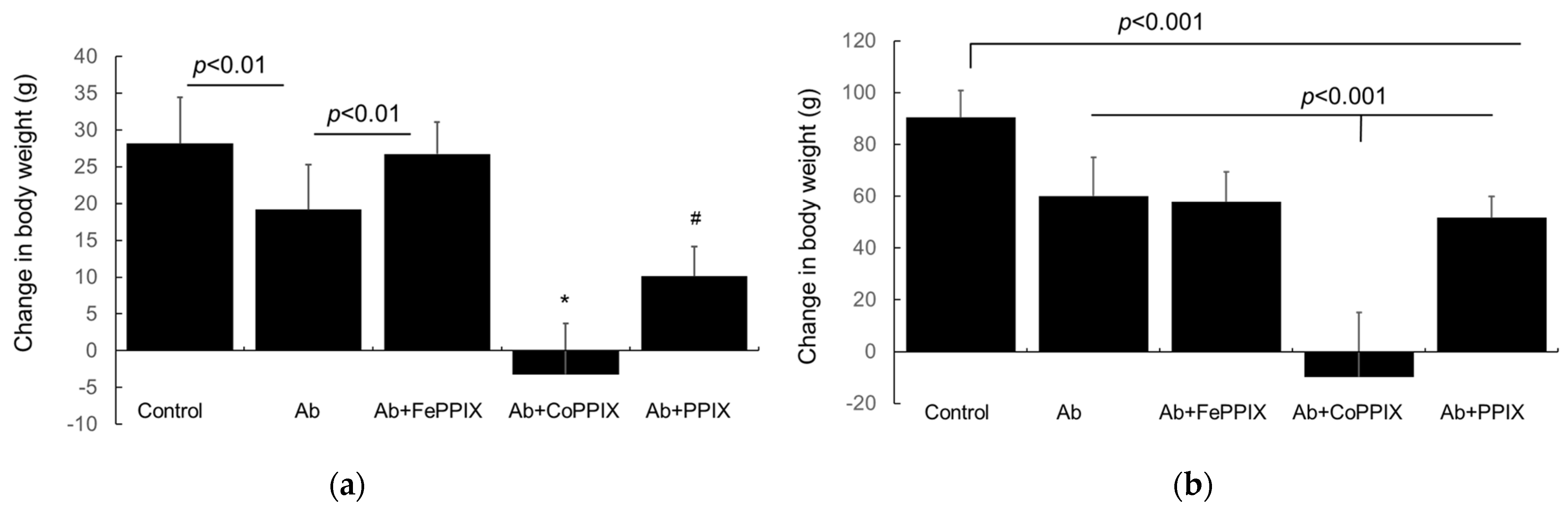
2.1. Effect of MP Treatment of Clinical Phenotype
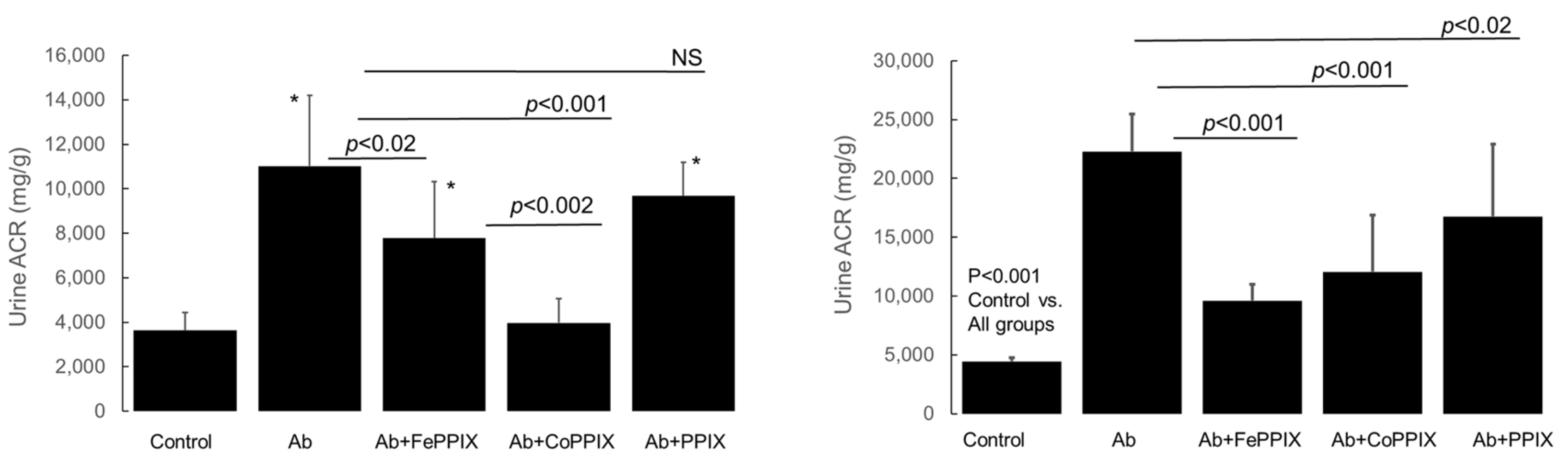
2.2. Metalloporphyrin (MP) Treatment Reduces Proteinuria in Rats with Passive Heymann Nephropathy (PHN)
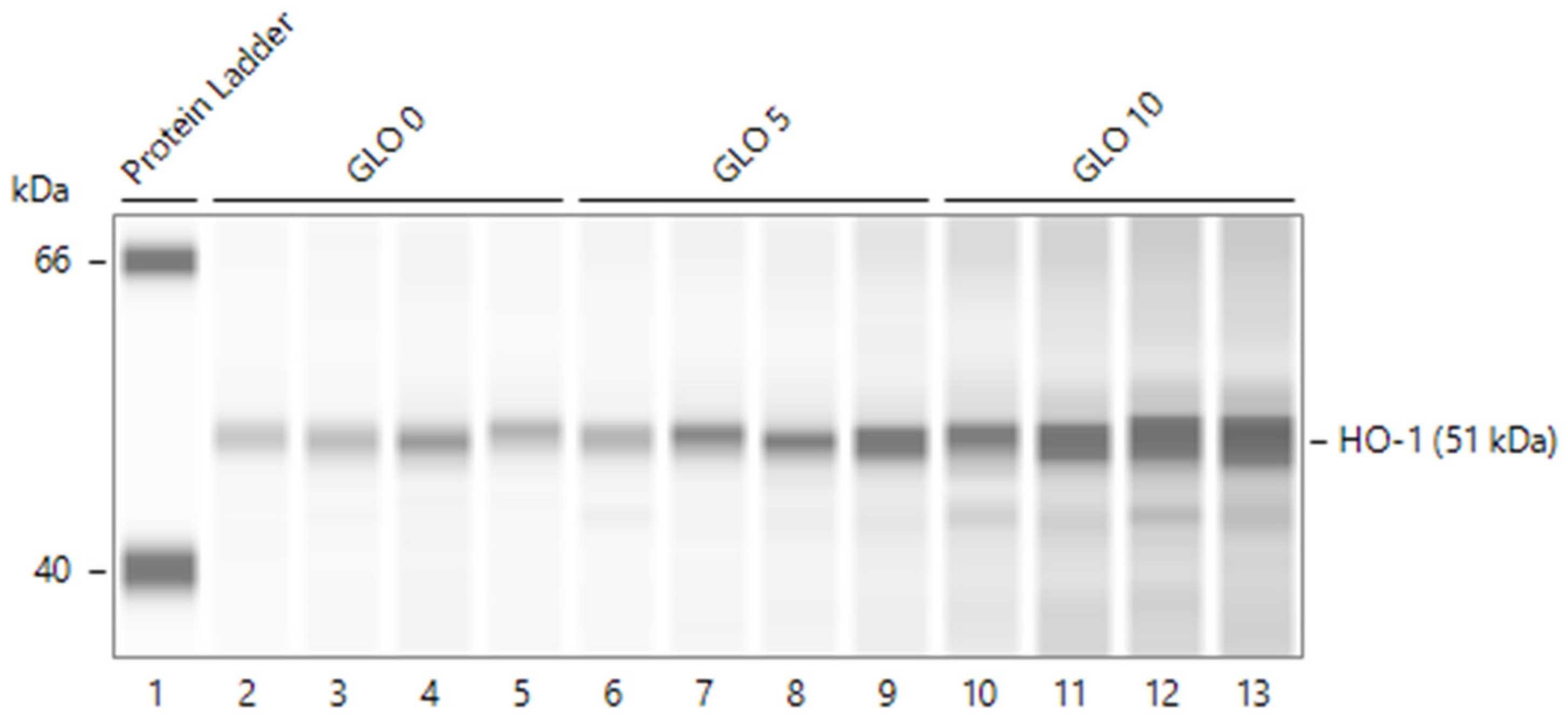
2.3. Effect of MP Treatment on Glomerular HO-1 Induction and on Determinants of Immune Injury (Complement Activation and Expression of Complement Regulatory Proteins) in PHN
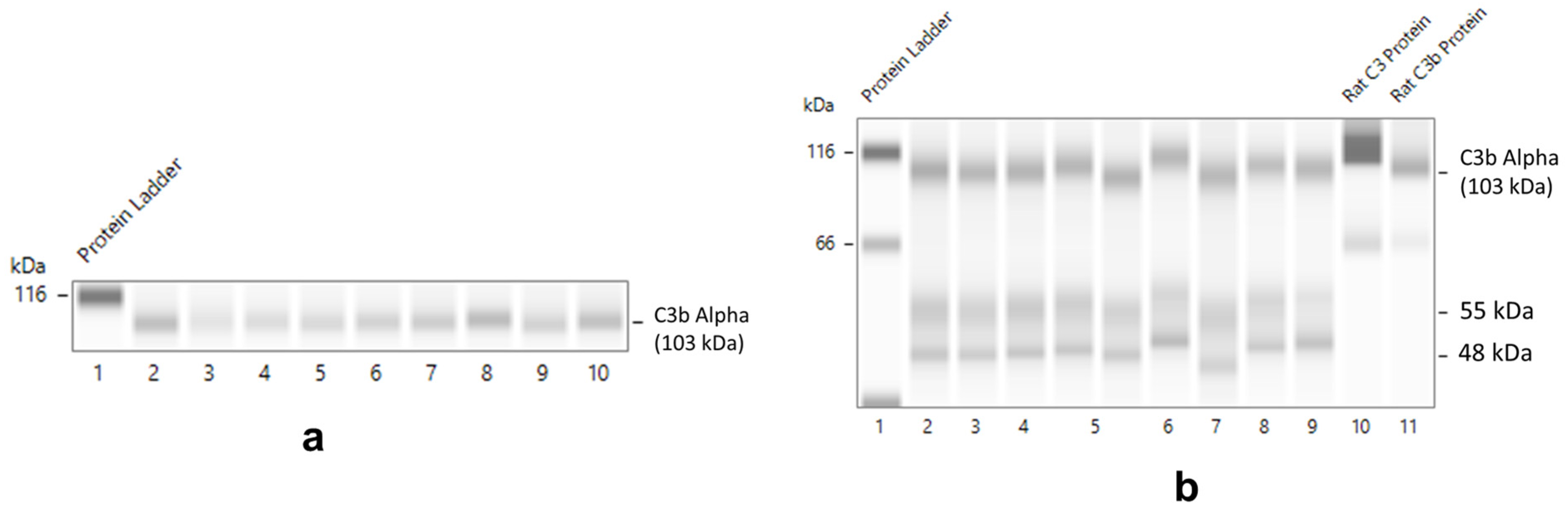
Validation of HO-1, CD55, and Complement Protein Detection in Normal Rat Glomeruli
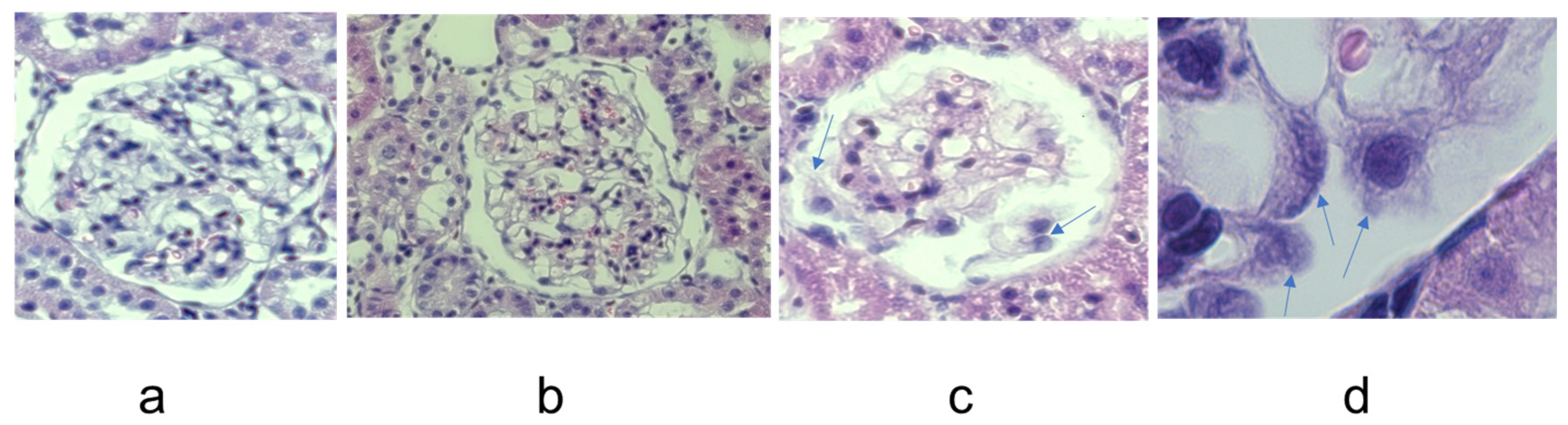
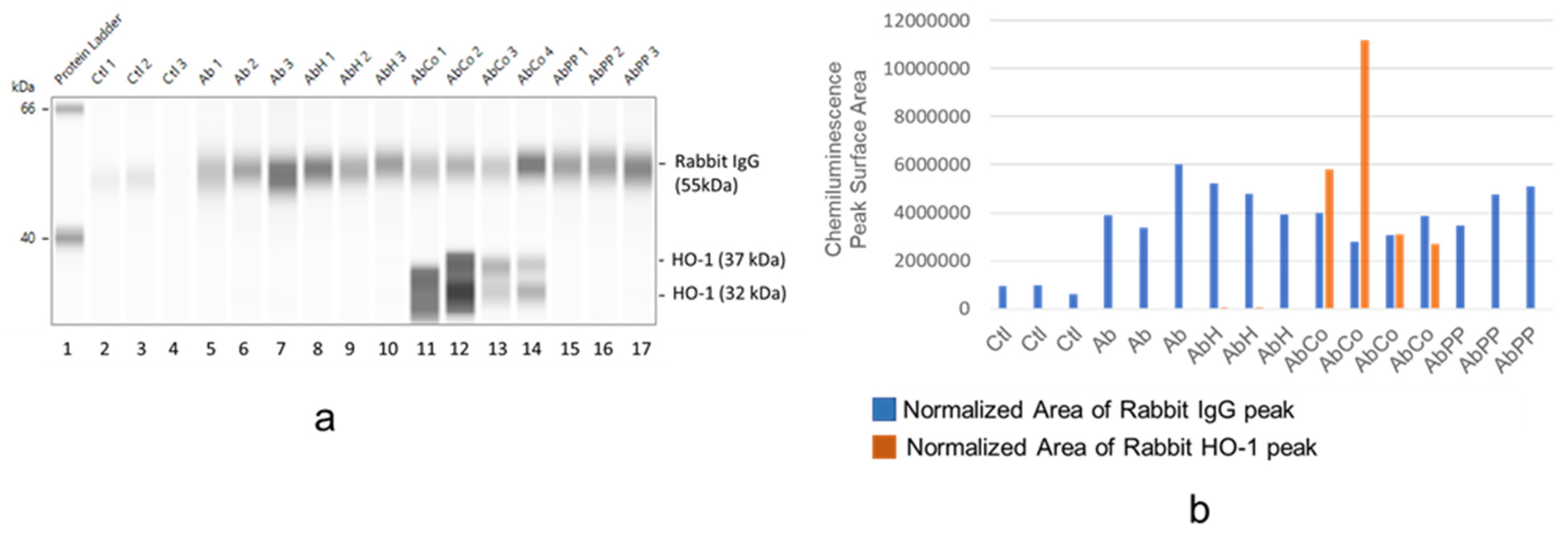
2.4. Differential HO-1 Induction in Glomeruli by MP Treatment
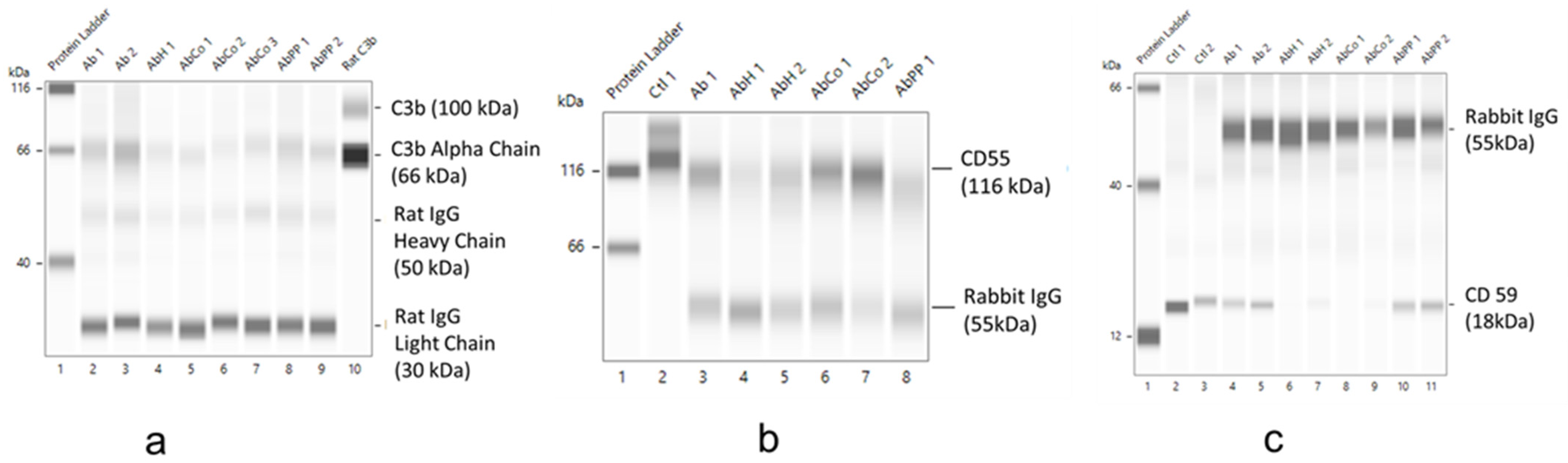
2.5. Effect of MP Treatment on Complement Deposition and CRP Expression
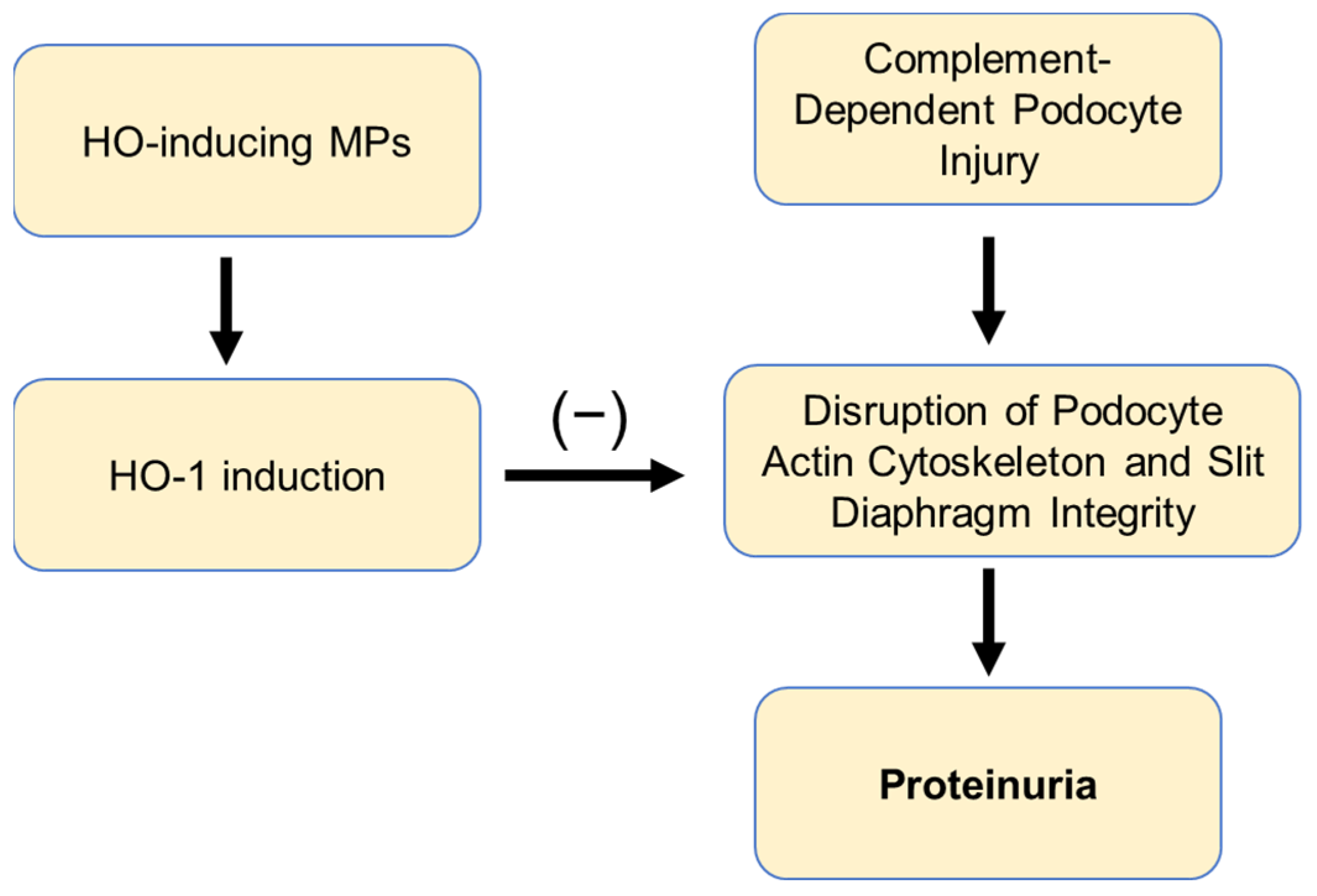
3. Discussion
4. Materials and Methods
4.1. Passive Heymann Nephropathy (PHN)
4.2. Metalloporphyrin (MP) Treatment Protocols and Tissue Retrieval
4.3. Capillary-Based Separation and Immunodetection (Western Blot Analysis) of Glomerular Proteins
4.4. Antibodies
4.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Correia, M.A.; Sinclair, P.R.; De Matteis, F. Cytochrome P450 regulation: The interplay between its heme and apoprotein moieties in synthesis, assembly, repair, and disposal. Drug Metab. Rev. 2011, 43, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.; Scott, E.E. Endogenous insertion of non-native metalloporphyrins into human membrane cytochrome P450 enzymes. J. Biol. Chem. 2018, 293, 16623–16634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pae, H.O.; Oh, G.S.; Choi, B.M.; Chae, S.C.; Kim, Y.M.; Chung, K.R.; Chung, H.T. Carbon monoxide produced by heme oxygenase-1 suppresses T cell proliferation via inhibition of IL-2 production. J. Immunol. 2004, 172, 4744–4751. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, S.; Li, C.; Cui, L.; Ma, J.; Hui, Y. The non-canonical effects of heme oxygenase-1, a classical fighter against oxidative stress. Redox Biol. 2021, 47, 102170. [Google Scholar] [CrossRef]
- Bissell, D.M.; Anderson, K.E.; Bonkovsky, H.L. Porphyria. N. Engl. J. Med. 2017, 377, 862–872. [Google Scholar] [CrossRef]
- Wilson, H.M.; Welikson, R.E.; Luo, J.; Kean, T.J.; Cao, B.; Dennis, J.E.; Allen, M.D. Can Cytoprotective Cobalt Protoporphyrin Protect Skeletal Muscle and Muscle-derived Stem Cells From Ischemic Injury. Clin. Orthop. Relat. Res. 2015, 473, 2908–2919. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Wong, R.J.; Vreman, H.J.; Stevenson, D.K. Metalloporphyrins—An update. Front. Pharmacol. 2012, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Paez-Ribes, M.; González-Gualda, E.; Doherty, G.J.; Muñoz-Espín, D. Targeting senescent cells in translational medicine. EMBO Mol. Med. 2019, 11, e10234. [Google Scholar] [CrossRef]
- Kennedy, J.C.; Pottier, R.H. Endogenous protoporphyrin IX, a clinically useful photosensitizer for photodynamic therapy. J. Photochem. Photobiol. B 1992, 14, 275–292. [Google Scholar] [CrossRef]
- Cybulsky, A.V.; Quigg, R.J.; Salant, D.J. Experimental membranous nephropathy redux. Am. J. Physiol. Renal. Physiol. 2005, 289, F660–F671. [Google Scholar] [CrossRef]
- Morgan, B.P.; Meri, S. Membrane proteins that protect against complement lysis. Springer Semin. Immunopathol. 1994, 15, 369–396. [Google Scholar] [CrossRef]
- Detsika, M.G.; Duann, P.; Atsaves, V.; Papalois, A.; Lianos, E.A. Heme Oxygenase 1 Up-Regulates Glomerular Decay Accelerating Factor Expression and Minimizes Complement Deposition and Injury. Am. J. Pathol. 2016, 186, 2833–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detsika, M.G.; Duann, P.; Lianos, E.A. HO-1 expression control in the rat glomerulus. Biochem. Biophys. Res. Commun. 2015, 460, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Spiller, O.B.; St John, P.L.; Haas, M.; Hack, B.K.; Ren, G.; Cunningham, P.N.; Doshi, M.; Abrahamson, D.R.; Morgan, B.P.; et al. Decay-accelerating factor expression in the rat kidney is restricted to the apical surface of podocytes. Kidney Int. 2002, 62, 2010–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, O.; Zhou, W.; Miyazaki, M.; Abe, K.; Koji, T.; Verroust, P.; Tsukasaki, S.; Ozono, Y.; Harada, T.; Nakane, P.K.; et al. Intraglomerular C3 synthesis in rats with passive Heymann nephritis. Am. J. Pathol. 1997, 151, 1249–1256. [Google Scholar] [PubMed]
- Troldborg, A.; Jensen, L.; Deleuran, B.; Stengaard-Pedersen, K.; Thiel, S.; Jensenius, J.C. The C3dg Fragment of Complement Is Superior to Conventional C3 as a Diagnostic Biomarker in Systemic Lupus Erythematosus. Front. Immunol. 2018, 9, 581. [Google Scholar] [CrossRef] [Green Version]
- Kasinath, B.S.; Maaba, M.R.; Schwartz, M.M.; Lewis, E.J. Demonstration and characterization of C3 receptors on rat glomerular epithelial cells. Kidney Int. 1986, 30, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Tomaro, M.L.; Frydman, J.; Frydman, R.B. Heme oxygenase induction by CoCl2, Co-protoporphyrin IX, phenylhydrazine, and diamide: Evidence for oxidative stress involvement. Arch. Biochem. Biophys. 1991, 286, 610–617. [Google Scholar] [CrossRef]
- Datta, P.K.; Duann, P.; Lianos, E.A. Long-term effect of heme oxygenase (HO)-1 induction in glomerular immune injury. J. Lab. Clin. Med. 2006, 147, 150–155. [Google Scholar] [CrossRef]
- Wu, C.C.; Lu, K.C.; Chen, J.S.; Hsieh, H.Y.; Lin, S.H.; Chu, P.; Wang, J.Y.; Sytwu, H.K.; Lin, Y.F. HO-1 induction ameliorates experimental murine membranous nephropathy: Anti-oxidative, anti-apoptotic and immunomodulatory effects. Nephrol. Dial. Transplant. 2008, 23, 3082–3090. [Google Scholar] [CrossRef] [Green Version]
- Heymann, W.; Hackel, D.B.; Harwood, S.; Wilson, S.G.; Hunter, J.L. Production of nephrotic syndrome in rats by Freund’s adjuvants and rat kidney suspensions. Proc. Soc. Exp. Biol. Med. 1959, 100, 660–664. [Google Scholar] [CrossRef]
- Edgington, T.S.; Glassock, R.J.; Dixon, F.J. Autologous immune complex nephritis induced with renal tubular antigen. I. Identification and isolation of the pathogenetic antigen. J. Exp. Med. 1968, 127, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, M.G.; Saito, A.; Kerjaschki, D.; Orlando, R.A. The Heymann nephritis antigenic complex: Megalin (gp330) and RAP. J. Am. Soc. Nephrol. 1995, 6, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Kerjaschki, D.; Farquhar, M.G. Immunocytochemical localization of the Heymann nephritis antigen (GP330) in glomerular epithelial cells of normal Lewis rats. J. Exp. Med. 1983, 157, 667–686. [Google Scholar] [CrossRef] [PubMed]
- Cybulsky, A.V.; Rennke, H.G.; Feintzeig, I.D.; Salant, D.J. Complement-induced glomerular epithelial cell injury. Role of the membrane attack complex in rat membranous nephropathy. J. Clin. Investig. 1986, 77, 1096–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerjaschki, D.; Schulze, M.; Binder, S.; Kain, R.; Ojha, P.P.; Susani, M.; Horvat, R.; Baker, P.J.; Couser, W.G. Transcellular transport and membrane insertion of the C5b-9 membrane attack complex of complement by glomerular epithelial cells in experimental membranous nephropathy. J. Immunol. 1989, 143, 546–552. [Google Scholar] [CrossRef]
- Spicer, S.T.; Tran, G.T.; Killingsworth, M.C.; Carter, N.; Power, D.A.; Paizis, K.; Boyd, R.; Hodgkinson, S.J.; Hall, B.M. Induction of passive Heymann nephritis in complement component 6-deficient PVG rats. J. Immunol. 2007, 179, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Saran, A.M.; Yuan, H.; Takeuchi, E.; McLaughlin, M.; Salant, D.J. Complement mediates nephrin redistribution and actin dissociation in experimental membranous nephropathy. Kidney Int. 2003, 64, 2072–2078. [Google Scholar] [CrossRef] [Green Version]
- Schulze, M.; Donadio, J.V.; Pruchno, C.J.; Baker, P.J.; Johnson, R.J.; Stahl, R.A.; Watkins, S.; Martin, D.C.; Wurzner, R.; Gotze, O. Elevated urinary excretion of the C5b-9 complex in membranous nephropathy. Kidney Int. 1991, 40, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Ryter, S.W. Heme Oxygenase-1: An Anti-Inflammatory Effector in Cardiovascular, Lung, and Related Metabolic Disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef]
- Maines, M.D.; Kappas, A. Enzymatic oxidation of cobalt protoporphyrin IX: Observations on the mechanism of heme oxygenase action. Biochemistry 1977, 16, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D.; Cohn, J. Bile pigment formation by skin heme oxygenase: Studies on the response of the enzyme to heme compounds and tissue injury. J. Exp. Med. 1977, 145, 1054–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lianos, E.A.; Detsika, M.G. Metalloporphyrins as Tools for Deciphering the Role of Heme Oxygenase in Renal Immune Injury. Int. J. Mol. Sci. 2023, 24, 6815. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wu, Y.; Fan, C.; Feng, C.; Wang, H.; Bai, F.; Zuo, J.; Tang, W. Heme supplementation ameliorates lupus nephritis through rectifying the disorder of splenocytes and alleviating renal inflammation and oxidative damage. Int. Immunopharmacol. 2021, 94, 107482. [Google Scholar] [CrossRef] [PubMed]
- Mackern-Oberti, J.P.; Obreque, J.; Méndez, G.P.; Llanos, C.; Kalergis, A.M. Carbon monoxide inhibits T cell activation in target organs during systemic lupus erythematosus. Clin. Exp. Immunol. 2015, 182, 1–13. [Google Scholar] [CrossRef]
- Wang, Y.; Kojetin, D.; Burris, T.P. Anti-proliferative actions of a synthetic REV-ERBα/β agonist in breast cancer cells. Biochem. Pharmacol. 2015, 96, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Fakih, T.M.; Kurniawan, F.; Yusuf, M.; Mudasir, M.; Tjahjono, D.H. Molecular Dynamics of Cobalt Protoporphyrin Antagonism of the Cancer Suppressor REV-ERBβ. Molecules 2021, 26, 3251. [Google Scholar] [CrossRef]
- Galbraith, R.A.; Hodgdon, I.; Grimm, M.S.; Vizzard, M.A. Prolonged retention of the anorectic cobalt protoporphyrin in the hypothalamus and the resulting expression of Fos. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R465–R471. [Google Scholar] [CrossRef]
- Rosenberg, D.W. Pharmacokinetics of cobalt chloride and cobalt-protoporphyrin. Drug Metab. Dispos. 1993, 21, 846–849. [Google Scholar]
- Siegert, S.W.; Holt, R.J. Physicochemical properties, pharmacokinetics, and pharmacodynamics of intravenous hematin: A literature review. Adv. Ther. 2008, 25, 842–857. [Google Scholar] [CrossRef]
- Zhou, W.; Andrews, P.A.; Wang, Y.; Wolff, J.; Pratt, J.; Hartley, B.R.; Verroust, P.; Sacks, S.H. Evidence for increased synthesis of complement C4 in the renal epithelium of rats with passive Heymann nephritis. J. Am. Soc. Nephrol. 1997, 8, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Detsika, M.G.; Goudevenou, K.; Geurts, A.M.; Gakiopoulou, H.; Grapsa, E.; Lianos, E.A. Generation of a novel decay accelerating factor (DAF) knock-out rat model using clustered regularly-interspaced short palindromic repeats, (CRISPR)/associated protein 9 (Cas9), genome editing. Transgenic Res. 2021, 30, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.H.; Heikkila, J.J. Effect of hemin, baicalein and heme oxygenase-1 (HO-1) enzyme activity inhibitors on Cd-induced accumulation of HO-1, HSPs and aggresome-like structures in Xenopus kidney epithelial cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 210, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; McCarthy, E.T.; Sharma, R.; Fish, B.L.; Savin, V.J.; Cohen, E.P.; Moulder, J.E. Arachidonic acid metabolites mediate the radiation-induced increase in glomerular albumin permeability. Exp. Biol. Med. 2006, 231, 99–106. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lianos, E.A.; Phung, G.N.; Foster, M.; Zhou, J.; Sharma, M. Metalloporphyrins Reduce Proteinuria in Podocyte Immune Injury: The Role of Metal and Porphyrin Moieties. Int. J. Mol. Sci. 2023, 24, 12777. https://doi.org/10.3390/ijms241612777
Lianos EA, Phung GN, Foster M, Zhou J, Sharma M. Metalloporphyrins Reduce Proteinuria in Podocyte Immune Injury: The Role of Metal and Porphyrin Moieties. International Journal of Molecular Sciences. 2023; 24(16):12777. https://doi.org/10.3390/ijms241612777
Chicago/Turabian StyleLianos, Elias A., Gia Nghi Phung, Michelle Foster, Jianping Zhou, and Mukut Sharma. 2023. "Metalloporphyrins Reduce Proteinuria in Podocyte Immune Injury: The Role of Metal and Porphyrin Moieties" International Journal of Molecular Sciences 24, no. 16: 12777. https://doi.org/10.3390/ijms241612777