
ERAP1 and ERAP2 Haplotypes Influence Suboptimal HLA-B*27:05-Restricted Anti-Viral CD8+ T Cell Responses Cross-Reactive to Self-Epitopes
 , , , ,
, , , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
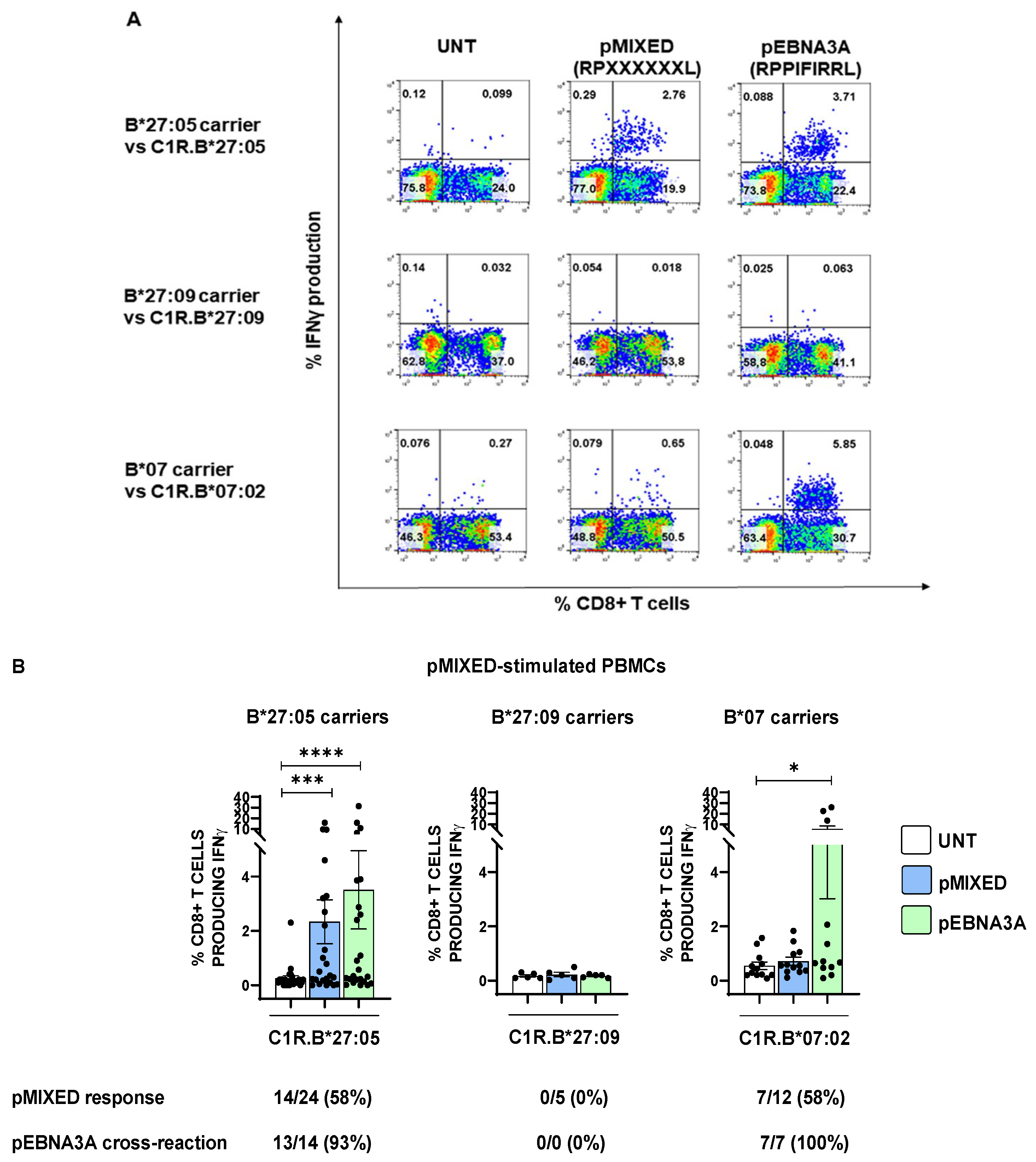
2.1. An AS-Protective ERAP1/ERAP2 Haplotype Negatively Correlates with the Ability of B*27:05 Carriers to Elicit a Specific CD8+ T Response towards the Suboptimal pEBNA3A Viral Peptide
2.2. Peptides with the RP Motif at the N-Terminus Trigger CD8+ T Cell Reactivity in the B*27:05 Context of Presentation
2.3. pEBNA3A-Responsive CD8+ T Cells Are Cross-Reactive against a Self-Peptide from Dynein Motor Protein
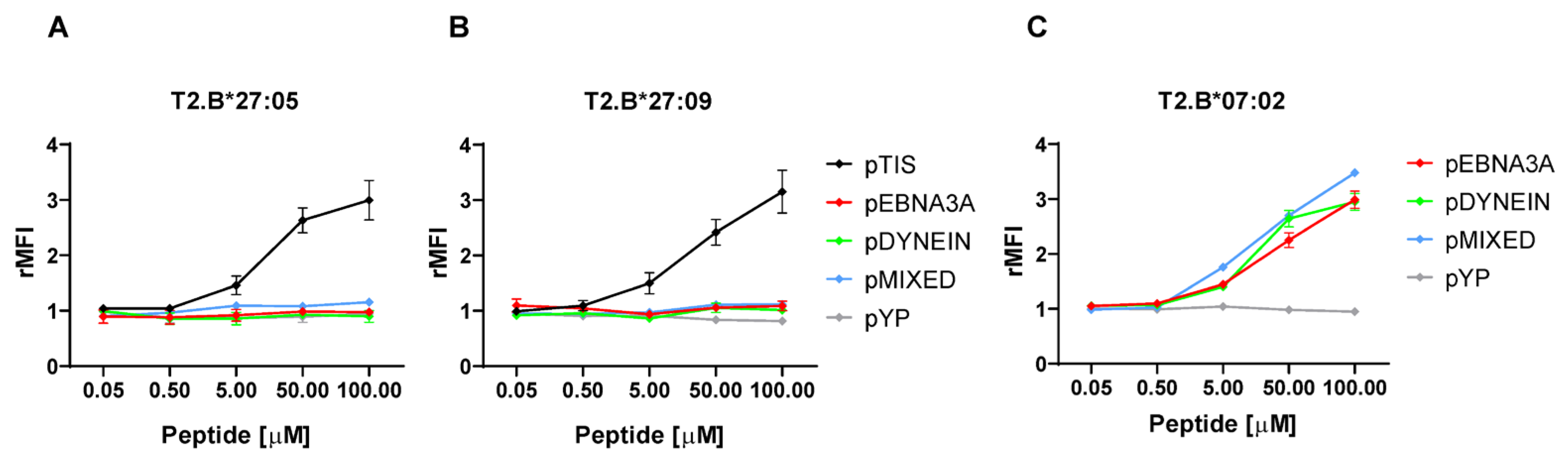
2.4. Peptides with the RP Motif at the N-Terminus Do Not Stabilize the B*27 Molecules
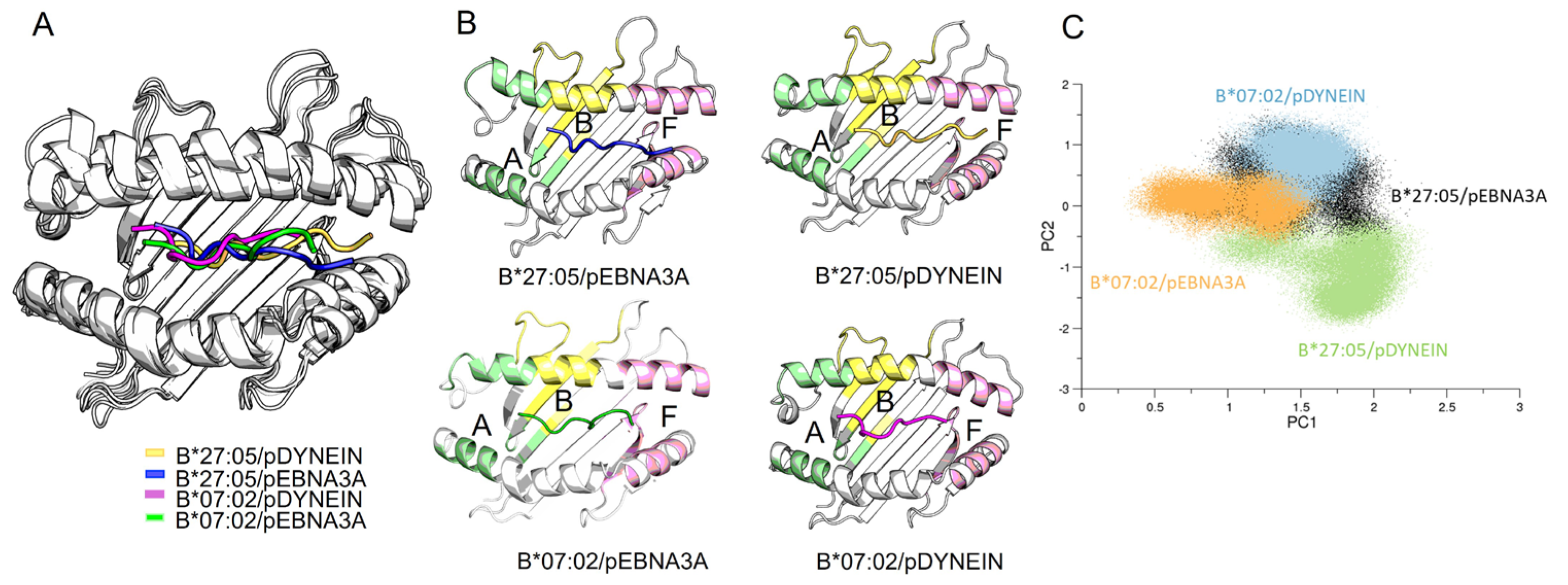
2.5. Molecular Dynamic (MD) Simulations of HLA-B Subtypes
3. Discussion
4. Materials and Methods
4.1. Study Subjects
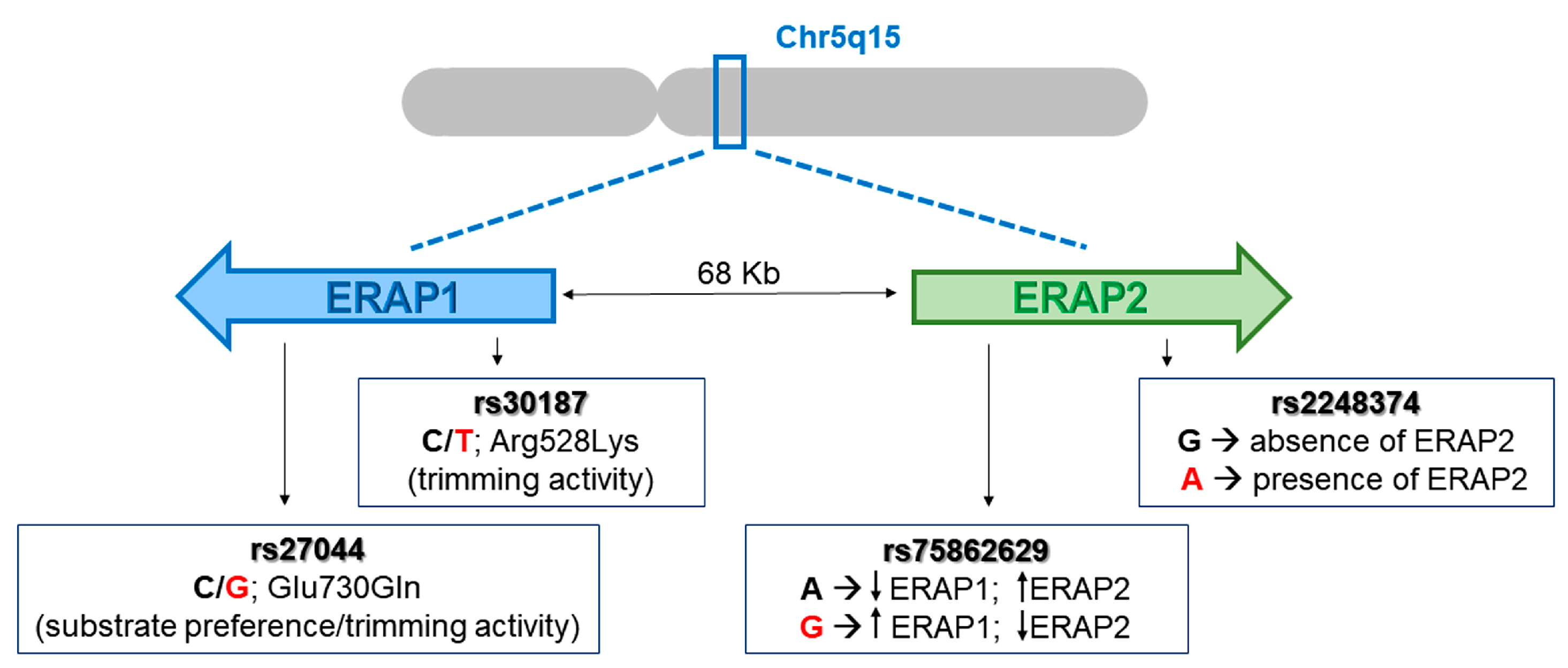
4.2. ERAPs Genotyping
4.3. EVB Seropositivity
4.4. Synthetic Peptides
4.5. Cell Lines
4.6. PBMCs Stimulation
4.7. Intracellular IFNγ Staining
4.8. T2 Stabilization Assay
4.9. Statistics
4.10. MD Simulation
4.11. Principal Component Analysis
4.12. Solvent Exposure, Hydrogen Bonds, Peptides Displacement and the Contact Matrix
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brewerton, D.A.; Hart, F.D.; Nicholls, A.; Carey, M.; James, D.C.; Sturrock, R.D. Ankylosing spondylitis and HL-A 27. Lancet 1973, 1, 904–907. [Google Scholar] [CrossRef] [PubMed]
- Schlosstein, L.; Terasaki, P.I.; Bluestone, R.; Pearson, C.M. High association of an HL-A antigen, W27, ankylosing spondylitis. N. Engl. J. Med. 1973, 288, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Mauro, D.; Thomas, R.; Guggino, G.; Lories, R.; Brown, M.A.; Ciccia, F. Ankylosing spondylitis: An autoimmune or autoinflammatory disease? Nat. Rev. Rheumatol. 2021, 17, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, R.; Parham, P. Guilt by association: HLA-B27 and ankylosing spondylitis. Immunol. Today 1990, 11, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, M.T.; Sorrentino, R. T-cell responses against viral and self-epitopes and HLA-B27 subtypes differentially associated with ankylosing spondylitis. Adv. Exp. Med. Biol. 2009, 649, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Garner, L.I.; Zvyagin, I.V.; Paley, M.A.; Komech, E.A.; Jude, K.M.; Zhao, X.; Fernandes, R.A.; Hassman, L.M.; Paley, G.L.; et al. Autoimmunity-associated T cell receptors recognize HLA-B*27-bound peptides. Nature 2022, 612, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Burton, P.R.; Clayton, D.G.; Cardon, L.R.; Craddock, N. Association scan of 14,500 nonsynonymous SNPs in four diseases identifies autoimmunity variants. Nat. Genet. 2007, 39, 1329–1337. [Google Scholar] [PubMed]
- Robinson, P.C.; Costello, M.E.; Leo, P.; Bradbury, L.A.; Hollis, K.; Cortes, A.; Lee, S.; Joo, K.B.; Shim, S.C.; Weisman, M.; et al. ERAP2 is associated with ankylosing spondylitis in HLA-B27-positive and HLA-B27-negative patients. Ann. Rheum. Dis. 2015, 74, 1627–1629. [Google Scholar] [CrossRef]
- Reeves, E.; James, E. The role of polymorphic ERAP1 in autoinflammatory disease. Biosci. Rep. 2018, 38, BSR20171503. [Google Scholar] [CrossRef]
- Tedeschi, V.; Paldino, G.; Paladini, F.; Mattorre, B.; Tuosto, L.; Sorrentino, R.; Fiorillo, M.T. The Impact of the ‘Mis-Peptidome’ on HLA Class I-Mediated Diseases: Contribution of ERAP1 and ERAP2 and Effects on the Immune Response. Int. J. Mol. Sci. 2020, 21, 9608. [Google Scholar] [CrossRef]
- López de Castro, J.A. How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins. Front. Immunol. 2018, 9, 2463. [Google Scholar] [CrossRef]
- Ombrello, M.J.; Kastner, D.L.; Remmers, E.F. Endoplasmic reticulum-associated amino-peptidase 1 and rheumatic disease: Genetics. Curr. Opin. Rheumatol. 2015, 27, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Bravo, A.; Alvarez-Navarro, C.; Martín-Esteban, A.; Barnea, E.; Admon, A.; López de Castro, J.A. Ranking the Contribution of Ankylosing Spondylitis-associated Endoplasmic Reticulum Aminopeptidase 1 (ERAP1) Polymorphisms to Shaping the HLA-B*27 Peptidome. Mol. Cell. Proteom. 2018, 17, 1308–1323. [Google Scholar] [CrossRef] [PubMed]
- Lorente, E.; Fontela, M.G.; Barnea, E.; Martín-Galiano, A.J.; Mir, C.; Galocha, B.; Admon, A.; Lauzurica, P.; López, D. Modulation of Natural HLA-B*27:05 Ligandome by Ankylosing Spondylitis-associated Endoplasmic Reticulum Aminopeptidase 2 (ERAP2). Mol. Cell. Proteom. 2020, 19, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- García-Medel, N.; Sanz-Bravo, A.; Alvarez-Navarro, C.; Gómez-Molina, P.; Barnea, E.; Marcilla, M.; Admon, A.; de Castro, J.A. Peptide handling by HLA-B27 subtypes influences their biological behavior, association with ankylosing spondylitis and susceptibility to endoplasmic reticulum aminopeptidase 1 (ERAP1). Mol. Cell. Proteom. 2014, 13, 3367–3380. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, M.; López de Castro, J.A. Peptides: The cornerstone of HLA-B27 biology and pathogenetic role in spondyloarthritis. Tissue Antigens 2008, 71, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Infantes, S.; Lorente, E.; Barnea, E.; Beer, I.; Barriga, A.; Lasala, F.; Jiménez, M.; Admon, A.; López, D. Natural HLA-B*27:05 protein ligands with glutamine as anchor motif: Implications for HLA-B27 association with spondyloarthropathy. J. Biol. Chem. 2013, 288, 10882–10889. [Google Scholar] [CrossRef]
- Yair-Sabag, S.; Tedeschi, V.; Vitulano, C.; Barnea, E.; Glaser, F.; Melamed Kadosh, D.; Taurog, J.D.; Fiorillo, M.T.; Sorrentino, R.; Admon, A. The Peptide Repertoire of HLA-B27 may include Ligands with Lysine at P2 Anchor Position. Proteomics 2018, 18, e1700249. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Szeto, C.; Gras, S. The pockets guide to HLA class I molecules. Biochem. Soc. Trans. 2021, 49, 2319–2331. [Google Scholar] [CrossRef]
- Schittenhelm, R.B.; Sian, T.C.; Wilmann, P.G.; Dudek, N.L.; Purcell, A.W. Revisiting the arthritogenic peptide theory: Quantitative not qualitative changes in the peptide repertoire of HLA-B27 allotypes. Arthritis Rheumatol. 2015, 67, 702–713. [Google Scholar] [CrossRef]
- Mauro, D.; Ciccia, F. Gut dysbiosis in Spondyloarthritis: Cause or effect? Best Pract. Res. Clin. Rheumatol. 2019, 33, 101493. [Google Scholar] [CrossRef] [PubMed]
- Berland, M.; Meslier, V.; Berreira Ibraim, S.; Le Chatelier, E.; Pons, N.; Maziers, N.; Thirion, F.; Gauthier, F.; Plaza Oñate, F.; Furet, J.P.; et al. Both Disease Activity and HLA-B27 Status Are Associated With Gut Microbiome Dysbiosis in Spondyloarthritis Patients. Arthritis Rheumatol. 2023, 75, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Dulphy, N.; Peyrat, M.A.; Tieng, V.; Douay, C.; Rabian, C.; Tamouza, R.; Laoussadi, S.; Berenbaum, F.; Chabot, A.; Bonneville, M.; et al. Common intra-articular T cell expansions in patients with reactive arthritis: Identical beta-chain junctional sequences and cytotoxicity toward HLA-B27. J. Immunol. 1999, 162, 3830–3839. [Google Scholar] [CrossRef] [PubMed]
- May, E.; Dulphy, N.; Frauendorf, E.; Duchmann, R.; Bowness, P.; Lopez de Castro, J.A.; Toubert, A.; Märker-Hermann, E. Conserved TCR beta chain usage in reactive arthritis; evidence for selection by a putative HLA-B27-associated autoantigen. Tissue Antigens 2002, 60, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Faham, M.; Carlton, V.; Moorhead, M.; Zheng, J.; Klinger, M.; Pepin, F.; Asbury, T.; Vignali, M.; Emerson, R.O.; Robins, H.S.; et al. Discovery of T Cell Receptor β Motifs Specific to HLA-B27-Positive Ankylosing Spondylitis by Deep Repertoire Sequence Analysis. Arthritis Rheumatol. 2017, 69, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Komech, E.A.; Pogorelyy, M.V.; Egorov, E.S.; Britanova, O.V.; Rebrikov, D.V.; Bochkova, A.G.; Shmidt, E.I.; Shostak, N.A.; Shugay, M.; Lukyanov, S.; et al. CD8+ T cells with characteristic T cell receptor beta motif are detected in blood and expanded in synovial fluid of ankylosing spondylitis patients. Rheumatology 2018, 57, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Hanson, A.L.; Nel, H.J.; Bradbury, L.; Phipps, J.; Thomas, R.; Lê Cao, K.A.; Kenna, T.J.; Brown, M.A. Altered Repertoire Diversity and Disease-Associated Clonal Expansions Revealed by T Cell Receptor Immunosequencing in Ankylosing Spondylitis Patients. Arthritis Rheumatol. 2020, 72, 1289–1302. [Google Scholar] [CrossRef]
- Tedeschi, V.; Vitulano, C.; Cauli, A.; Paladini, F.; Piga, M.; Mathieu, A.; Sorrentino, R.; Fiorillo, M.T. The Ankylosing Spondylitis-associated HLA-B*2705 presents a B*0702-restricted EBV epitope and sustains the clonal amplification of cytotoxic T cells in patients. Mol. Med. 2016, 22, 215–223. [Google Scholar] [CrossRef]
- Tedeschi, V.; Alba, J.; Paladini, F.; Paroli, M.; Cauli, A.; Mathieu, A.; Sorrentino, R.; D’Abramo, M.; Fiorillo, M.T. Unusual Placement of an EBV Epitope into the Groove of the Ankylosing Spondylitis-Associated HLA-B27 Allele Allows CD8+ T Cell Activation. Cells 2019, 8, 572. [Google Scholar] [CrossRef]
- Hill, A.; Worth, A.; Elliott, T.; Rowland-Jones, S.; Brooks, J.; Rickinson, A.; McMichael, A. Characterization of two Epstein-Barr virus epitopes restricted by HLA-B7. Eur. J. Immunol. 1995, 25, 18–24. [Google Scholar] [CrossRef]
- Rowntree, L.C.; Nguyen, T.H.O.; Farenc, C.; Halim, H.; Hensen, L.; Rossjohn, J.; Kotsimbos, T.C.; Purcell, A.W.; Kedzierska, K.; Gras, S.; et al. A Shared TCR Bias toward an Immunogenic EBV Epitope Dominates in HLA-B*07-Expressing Individuals. J. Immunol. 2020, 205, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Dunmire, S.K.; Verghese, P.S.; Balfour, H.H., Jr. Primary Epstein-Barr virus infection. J. Clin. Virol. 2018, 102, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Vitulano, C.; Tedeschi, V.; Paladini, F.; Sorrentino, R.; Fiorillo, M.T. The interplay between HLA-B27 and ERAP1/ERAP2 aminopeptidases: From anti-viral protection to spondyloarthritis. Clin. Exp. Immunol. 2017, 190, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Paladini, F.; Fiorillo, M.T.; Tedeschi, V.; D’Otolo, V.; Piga, M.; Cauli, A.; Mathieu, A.; Sorrentino, R. The rs75862629 minor allele in the endoplasmic reticulum aminopeptidases intergenic region affects human leucocyte antigen B27 expression and protects from ankylosing spondylitis in Sardinia. Rheumatology 2019, 58, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- Hearn, A.; York, I.A.; Rock, K.L. The specificity of trimming of MHC class I-presented peptides in the endoplasmic reticulum. J. Immunol. 2009, 183, 5526–5536. [Google Scholar] [CrossRef] [PubMed]
- Saveanu, L.; Carroll, O.; Lindo, V.; Del Val, M.; Lopez, D.; Lepelletier, Y.; Greer, F.; Schomburg, L.; Fruci, D.; Niedermann, G.; et al. Concerted peptide trimming by human ERAP1 and ERAP2 aminopeptidase complexes in the endoplasmic reticulum. Nat. Immunol. 2005, 6, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Zervoudi, E.; Papakyriakou, A.; Georgiadou, D.; Evnouchidou, I.; Gajda, A.; Poreba, M.; Salvesen, G.S.; Drag, M.; Hattori, A.; Swevers, L.; et al. Probing the S1 specificity pocket of the aminopeptidases that generate antigenic peptides. Biochem. J. 2011, 435, 411–420. [Google Scholar] [CrossRef]
- Birtley, J.R.; Saridakis, E.; Stratikos, E.; Mavridis, I.M. The crystal structure of human endoplasmic reticulum aminopeptidase 2 reveals the atomic basis for distinct roles in antigen processing. Biochemistry 2012, 51, 286–295. [Google Scholar] [CrossRef]
- Mukherjee, S.; Sanchez-Bernabeu, A.; Demmers, L.C.; Wu, W.; Heck, A.J.R. The HLA Ligandome Comprises a Limited Repertoire of O-GlcNAcylated Antigens Preferentially Associated With HLA-B*07. Front. Immunol. 2021, 12, 796584. [Google Scholar] [CrossRef]
- Lee, K.H.; Chang, Y.C.; Chen, T.F.; Juan, H.F.; Tsai, H.K.; Chen, C.Y. Connecting MHC-I-binding motifs with HLA alleles via deep learning. Commun. Biol. 2021, 4, 1194. [Google Scholar] [CrossRef]
- Ranganathan, V.; Gracey, E.; Brown, M.A.; Inman, R.D.; Haroon, N. Pathogenesis of ankylosing spondylitis—Recent advances and future directions. Nat. Rev. Rheumatol. 2017, 13, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Guerini, F.R.; Bolognesi, E.; Lax, A.; Bianchi, L.N.C.; Caronni, A.; Zanzottera, M.; Agliardi, C.; Albergoni, M.P.; Banfi, P.I.; Navarro, J.; et al. HLA Allele Frequencies and Association with Severity of COVID-19 Infection in Northern Italian Patients. Cells 2022, 11, 1792. [Google Scholar] [CrossRef] [PubMed]
- Rowntree, L.C.; Nguyen, T.H.O.; Halim, H.; Purcell, A.W.; Rossjohn, J.; Gras, S.; Kotsimbos, T.C.; Mifsud, N.A. Inability To Detect Cross-Reactive Memory T Cells Challenges the Frequency of Heterologous Immunity among Common Viruses. J. Immunol. 2018, 200, 3993–4003. [Google Scholar] [CrossRef] [PubMed]
- Mattorre, B.; Tedeschi, V.; Paldino, G.; Fiorillo, M.T.; Paladini, F.; Sorrentino, R. The emerging multifunctional roles of ERAP1, ERAP2 and IRAP between antigen processing and renin-angiotensin system modulation. Front. Immunol. 2022, 13, 1002375. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.M.; Spencer, C.C.; Pointon, J.J.; Su, Z.; Harvey, D.; Kochan, G.; Oppermann, U.; Dilthey, A.; Pirinen, M.; Stone, M.A.; et al. Wellcome Trust Case Control Consortium 2 (WTCCC2). Interaction between ERAP1 and HLA-B27 in ankylosing spondylitis implicates peptide handling in the mechanism for HLA-B27 in disease susceptibility. Nat. Genet. 2011, 43, 761–767, Erratum in Nat Genet. 2011, 43, 919. [Google Scholar] [CrossRef]
- Fruci, D.; Romania, P.; D’Alicandro, V.; Locatelli, F. Endoplasmic reticulum aminopeptidase 1 function and its pathogenic role in regulating innate and adaptive immunity in cancer and major histocompatibility complex class I-associated autoimmune diseases. Tissue Antigens 2014, 84, 177–186. [Google Scholar] [CrossRef]
- Kuiper, J.J.; Prinz, J.C.; Stratikos, E.; Kuśnierczyk, P.; Arakawa, A.; Springer, S.; Mintoff, D.; Padjen, I.; Shumnalieva, R.; Vural, S.; et al. Identifying disease-overarching mechanisms across disciplines and borders. Ann. Rheum. Dis. 2023, 82, 887–896. [Google Scholar] [CrossRef]
- Arakawa, A.; Reeves, E.; Vollmer, S.; Arakawa, Y.; He, M.; Galinski, A.; Stöhr, J.; Dornmair, K.; James, E.; Prinz, J.C. ERAP1 Controls the Autoimmune Response against Melanocytes in Psoriasis by Generating the Melanocyte Autoantigen and Regulating Its Amount for HLA-C*06:02 Presentation. J. Immunol. 2021, 207, 2235–2244. [Google Scholar] [CrossRef]
- Garrido-Mesa, J.; Brown, M.A. T cell Repertoire Profiling and the Mechanism by which HLA-B27 Causes Ankylosing Spondylitis. Curr. Rheumatol. Rep. 2022, 24, 398–410. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Kroemer, G. Hallmarks of T cell aging. Nat. Immunol. 2021, 22, 687–698. [Google Scholar] [CrossRef]
- van der Linden, S.; Valkenburg, H.A.; Cats, A. Evaluation of diagnostic criteria for ankylosing spondylitis. A proposal for modification of the New York criteria. Arthritis Rheum. 1984, 27, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.; Gladman, D.; Helliwell, P.; Marchesoni, A.; Mease, P.; Mielants, H.; CASPAR Study Group. Classification criteria for psoriatic arthritis: Development of new criteria from a large international study. Arthritis Rheum. 2006, 54, 2665–2673. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., III; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Pekka, M.; Lennart, N. Structure and Dynamics of the TIP3P, SPC, and SPC/E Water Models at 298 K. J. Phys. Chem. A 2001, 105, 9954–9960. [Google Scholar]
- Abraham, M.J.; van der Spoel, D.; Lindahl, E.; Hess, B.; Team, D. GROMACS User Manual Version 2021.5. Zenodo 2021. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [PubMed]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Huang, J.; MacKerell, A.D., Jr. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef]
- Amadei, A.; Linssen, A.B.; Berendsen, H.J. Essential dynamics of proteins. Proteins 1993, 17, 412–425. [Google Scholar] [CrossRef]
- Michaud-Agrawal, N.; Denning, E.J.; Woolf, T.B.; Beckstein, O. MDAnalysis: A toolkit for the analysis of molecular dynamics simulations. J. Comput. Chem. 2011, 32, 2319–2327. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pEBNA3A Responders (N = 73) | pEBNA3A Non-Responders (N = 29) | |||||
|---|---|---|---|---|---|---|
| Genotype | N | (%) | N | (%) | ||
| ERAP1 rs27044 C/G Glu730Gln | CC | 20 | 27.4 | 12 | 41.4 | |
| GC | 42 | 57.5 | 15 | 51.7 | ||
| GG | 11 | 15.1 | 2 | 6.9 | ||
| Allele | ||||||
| C | 82 | 56.2 | 39 | 67.2 | ||
| G | 64 | 43.8 | 19 | 32.8 | ||
| Genotype | N | (%) | N | (%) | ||
| ERAP1 rs30187 C/T Arg528Lys | CC | 7 | 9.6 | 11 | 37.9 | *** p value 0.0007 (CC vs. TT + CT) |
| CT | 48 | 65.7 | 12 | 41.4 | ||
| TT | 18 | 24.7 | 6 | 20.7 | ||
| Allele | ||||||
| C | 62 | 42.5 | 34 | 58.6 | * p value 0.037 | |
| T | 84 | 57.5 | 24 | 41.4 | ||
| Genotype | N | (%) | N | (%) | ||
| ERAP2 rs75862629intergenic region A/G | AA | 58 | 79.5 | 25 | 86.2 | |
| AG | 13 | 17.8 | 4 | 13.8 | ||
| GG | 2 | 2.7 | 0 | 0.0 | ||
| Allele | ||||||
| A | 129 | 88.4 | 54 | 93.1 | ||
| G | 17 | 11.6 | 4 | 6.9 | ||
| Genotype | N | (%) | N | (%) | ||
| ERAP2 rs2248374 G-> absence A-> presence | GG | 17 | 23.3 | 7 | 24.1 | |
| AG | 43 | 58.9 | 20 | 69.0 | ||
| AA | 13 | 17.8 | 2 | 6.9 | ||
| Allele | ||||||
| G | 77 | 52.7 | 34 | 58.6 | ||
| A | 69 | 47.3 | 24 | 41.4 | ||
| (rs27044-rs30187-rs75862629-rs2248374) Haplotypes | Frequency | pEBNA3A- Responders | pEBNA3A-Non-Responders | χ2 | p Value |
|---|---|---|---|---|---|
| G-T-A-G | 0.261 | 0.271 | 0.235 | 0.267 | 0.6052 |
| C-C-A-A | 0.238 | 0.221 | 0.281 | 0.844 | 0.3582 |
| C-C-A-G | 0.182 | 0.141 | 0.283 | 5.604 | * 0.0179 |
| C-T-A-A | 0.078 | 0.092 | 0.042 | 1.458 | 0.2272 |
| G-T-G-A | 0.065 | 0.071 | 0.050 | 0.309 | 0.5782 |
| C-T-A-G | 0.058 | 0.063 | 0.047 | 0.177 | 0.6736 |
| G-T-A-A | 0.057 | 0.064 | 0.038 | 0.554 | 0.4567 |
| C-C-G-G | 0.027 | 0.031 | 0.017 | 0.283 | 0.5947 |
| G-C-A-G | 0.017 | 0.022 | 0.003 | 0.910 | 0.3401 |
| C-T-G-A | 0.011 | 0.015 | 0.002 | 0.624 | 0.4295 |
| Groove B*27:05/B*07:02 | pDYNEIN/ HLA-B*27:05 | pEBNA3A/ HLA-B*27:05 | pDYNEIN/ HLA-B*07:02 | pEBNA3A/ HLA-B*07:02 |
|---|---|---|---|---|
| Glu 45/Glu 45 | P1 | P1 | ||
| Tyr 59/Tyr 59 | P1 | |||
| Glu 63/Asn 63 | P1 | P1 | P1 | |
| Lys 70/Gln 70 | P7 | |||
| Thr 73/Thr 73 | P8 | |||
| Asp 77/Ser 77 | ||||
| Thr 80/Asn 80 | P9 | |||
| Asn 97/Ser 97 | P7 | |||
| Tyr 99/Tyr 99 | P1 | |||
| His114/Asp 114 | P7 | |||
| Ile 142/Ile 142 | ||||
| Lys 146/Lys 146 | P7–P9 | P8 | P8–P9 | |
| Trp 147/Trp 147 | ||||
| Leu 156/Arg 156 | P7 | |||
| Tyr 159/Tyr 159 | P1 | P1 | P1 | |
| Glu 163/Glu 163 | P1 | P4 |
| HLA/Peptide Complex | P1—A Pocket Distance (nm) | P1—B Pocket Distance (nm) | P9—F Pocket Distance (nm) |
|---|---|---|---|
| B*27:05/pDYNEIN | 0.36 ± 0.04 | 0.46 ± 0.03 | 1.23 ± 0.14 |
| B*27:05/pEBNA3A | 0.42 ± 0.05 | 0.39 ± 0.04 | 0.8 ± 0.03 |
| B*07:02/pDYNEIN | 0.35 ± 0.04 | 0.63 ± 0.07 | 1.024 ± 0.04 |
| B*07:02/pEBNA3A | 0.45 ± 0.04 | 0.45 ± 0.04 | 0.34 ± 0.04 |
| HLA/Peptide Complex | Solvent Exposure (nm2) |
|---|---|
| B*27:05/pDYNEIN | 14.5 ± 0.3 |
| B*27:05/pEBNA3A | 16.4 ± 0.1 |
| B*07:02/pDYNEIN | 14.4 ± 0.7 |
| B*07:02/pEBNA3A | 16.6 ± 0.5 |
| Complexes | TΔS (KJ/mol) |
|---|---|
| B*27:05/pEBNA3A-pDYNEIN | −11.16 ± 5 |
| B*07:02/pEBNA3A-pDYNEIN | −50.5 ± 6.2 |
| B*07:02-B*27:05/pDYNEIN | −5.8 ± 17.9 |
| B*07:02-B*27:05/pEBNA3A | −44.98 ± 18.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedeschi, V.; Paldino, G.; Alba, J.; Molteni, E.; Paladini, F.; Scrivo, R.; Congia, M.; Cauli, A.; Caccavale, R.; Paroli, M.; et al. ERAP1 and ERAP2 Haplotypes Influence Suboptimal HLA-B*27:05-Restricted Anti-Viral CD8+ T Cell Responses Cross-Reactive to Self-Epitopes. Int. J. Mol. Sci. 2023, 24, 13335. https://doi.org/10.3390/ijms241713335
Tedeschi V, Paldino G, Alba J, Molteni E, Paladini F, Scrivo R, Congia M, Cauli A, Caccavale R, Paroli M, et al. ERAP1 and ERAP2 Haplotypes Influence Suboptimal HLA-B*27:05-Restricted Anti-Viral CD8+ T Cell Responses Cross-Reactive to Self-Epitopes. International Journal of Molecular Sciences. 2023; 24(17):13335. https://doi.org/10.3390/ijms241713335
Chicago/Turabian StyleTedeschi, Valentina, Giorgia Paldino, Josephine Alba, Emanuele Molteni, Fabiana Paladini, Rossana Scrivo, Mattia Congia, Alberto Cauli, Rosalba Caccavale, Marino Paroli, and et al. 2023. "ERAP1 and ERAP2 Haplotypes Influence Suboptimal HLA-B*27:05-Restricted Anti-Viral CD8+ T Cell Responses Cross-Reactive to Self-Epitopes" International Journal of Molecular Sciences 24, no. 17: 13335. https://doi.org/10.3390/ijms241713335
APA StyleTedeschi, V., Paldino, G., Alba, J., Molteni, E., Paladini, F., Scrivo, R., Congia, M., Cauli, A., Caccavale, R., Paroli, M., Di Franco, M., Tuosto, L., Sorrentino, R., D’Abramo, M., & Fiorillo, M. T. (2023). ERAP1 and ERAP2 Haplotypes Influence Suboptimal HLA-B*27:05-Restricted Anti-Viral CD8+ T Cell Responses Cross-Reactive to Self-Epitopes. International Journal of Molecular Sciences, 24(17), 13335. https://doi.org/10.3390/ijms241713335