
Drosophila melanogaster as a Translational Model System to Explore the Impact of Phytochemicals on Human Health
 , , ,
, , ,  , and
, and
Abstract
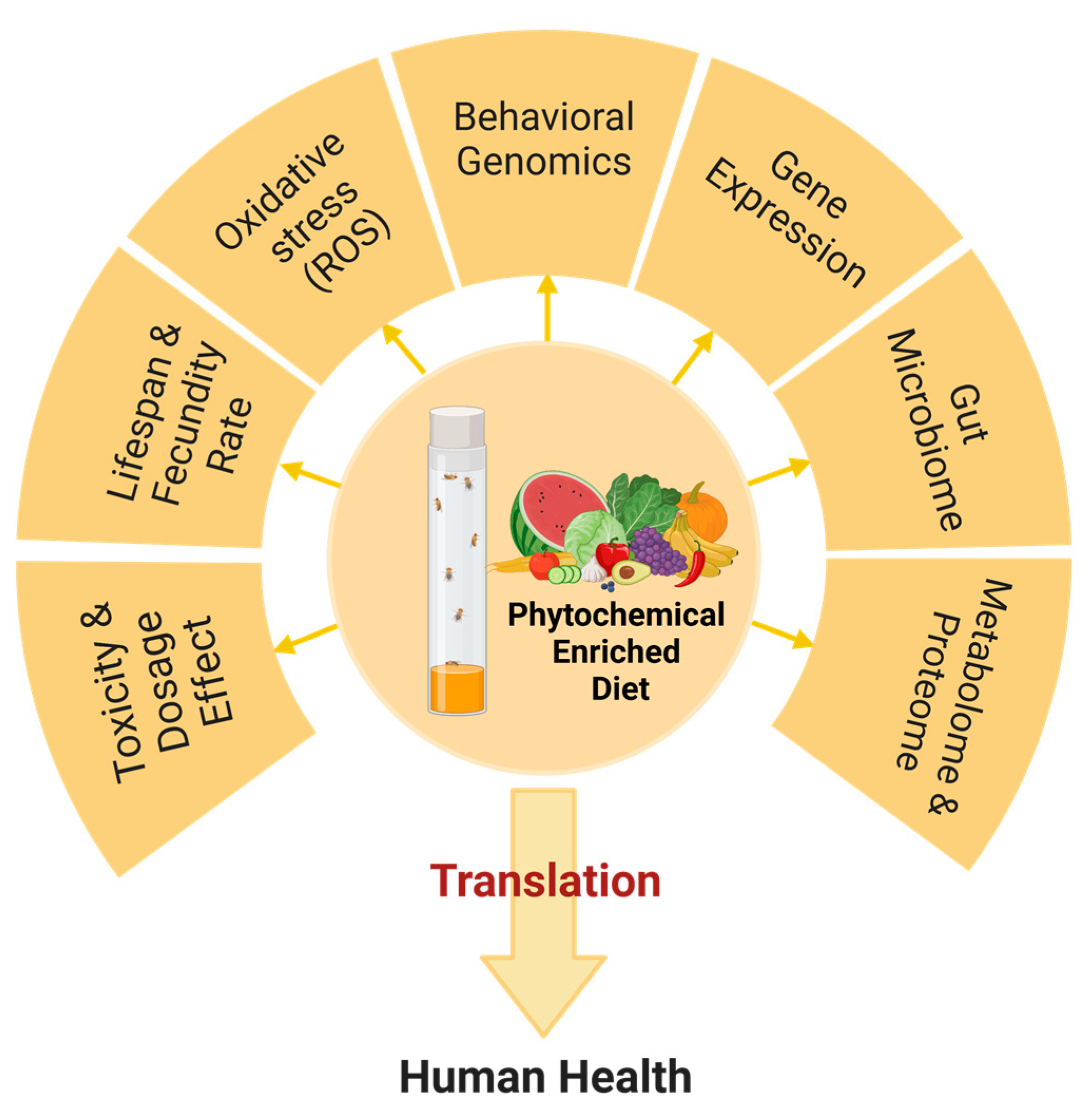
:1. Introduction
2. Phytochemicals and Their Potential Therapeutic Benefits
3. Advantages of Using Drosophila as a Translational Model for Testing Phytochemicals
4. Methods for Testing the Efficacy of Phytochemicals in Drosophila
5. Studies Evaluating the Effect of Phytochemicals by Using Drosophila
5.1. Phytochemical Effect on Aging
5.2. Development and Lifespan
5.3. Metabolism
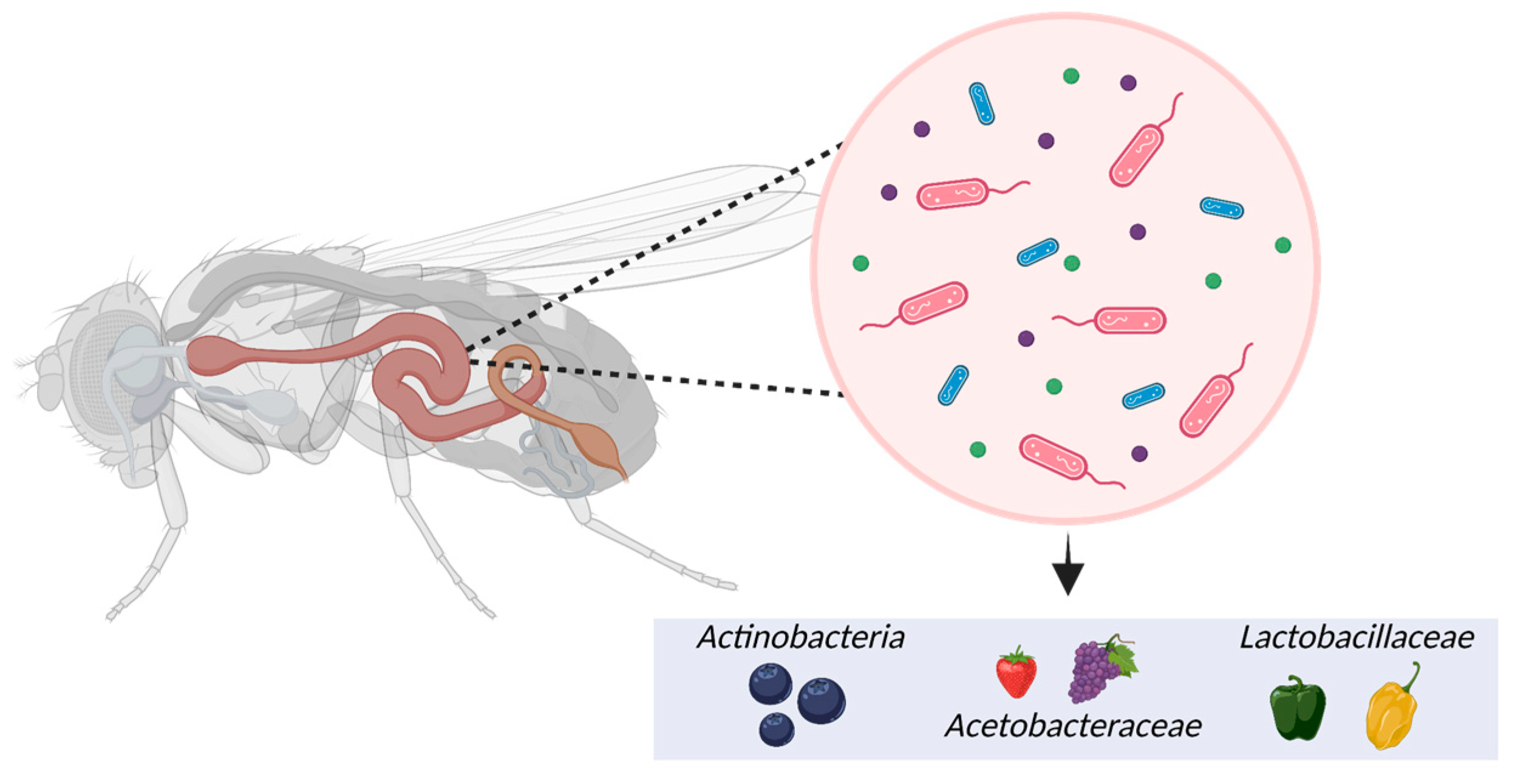
5.4. Microbiome
5.5. Neurodegenerative Diseases
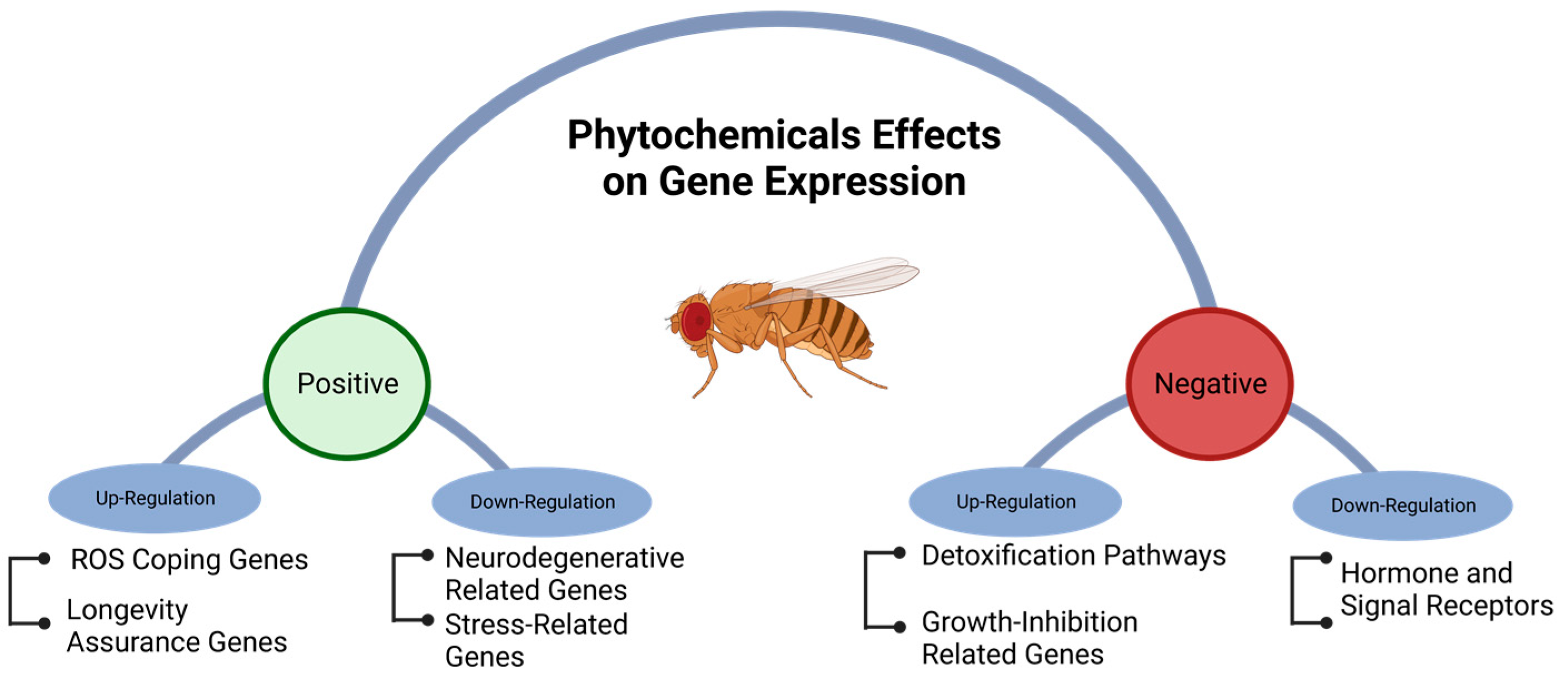
6. Gene Regulation Induced by Phytochemicals in Drosophila
7. Challenges and Limitations of Using Drosophila as a Translational Model
8. Future Directions and Opportunities for Using Drosophila to Test Phytochemicals
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Oladipo, A.; Enwemiwe, V.; Ejeromedoghene, O.; Adebayo, A.; Ogunyemi, O.; Fu, F. Production and functionalities of specialized metabolites from different organic sources. Metabolites 2022, 12, 534. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, M.; Jose, A.; Tomer, V.; Oz, E.; Proestos, C.; Zeng, M.; Elobeid, T.; Oz, F. Major Phytochemicals: Recent Advances in Health Benefits and Extraction Method. Molecules 2023, 28, 887. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Singh, G.; Sharma, A.; Aggarwal, G. Phytochemicals as candidate therapeutics: An overview. Int. J. Pharm. Sci. Rev. Res. 2010, 3, 53–55. [Google Scholar]
- Mullins, A.P.; Arjmandi, B.H. Health benefits of plant-based nutrition: Focus on beans in cardiometabolic diseases. Nutrients 2021, 13, 519. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Alam, S.; Sarker, M.M.R.; Sultana, T.N.; Chowdhury, M.N.R.; Rashid, M.A.; Chaity, N.I.; Zhao, C.; Xiao, J.; Hafez, E.E.; Khan, S.A. Antidiabetic phytochemicals from medicinal plants: Prospective candidates for new drug discovery and development. Front. Endocrinol. 2022, 13, 800714. [Google Scholar] [CrossRef]
- Pham, D.-C.; Shibu, M.; Mahalakshmi, B.; Velmurugan, B.K. Effects of phytochemicals on cellular signaling: Reviewing their recent usage approaches. Crit. Rev. Food Sci. Nutr. 2020, 60, 3522–3546. [Google Scholar] [CrossRef]
- Kim, S.-I.; Jung, J.-W.; Ahn, Y.-J.; Restifo, L.L.; Kwon, H.-W. Drosophila as a model system for studying lifespan and neuroprotective activities of plant-derived compounds. J. Asia-Pac. Entomol. 2011, 14, 509–517. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, S.; Kumar, V.; Thakur, A. Comprehensive review on nutraceutical significance of phytochemicals as functional food ingredients for human health management. J. Pharmacogn. Phytochem. 2019, 8, 385–395. [Google Scholar] [CrossRef]
- Sharma, A.; D’Souza, C.; Rai, V.; Gupta, S.C. Evaluation of Safety and Efficacy of Nutraceuticals Using Drosophila as an in vivo Tool. In Nutraceuticals in Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2019; pp. 685–692. [Google Scholar]
- Moraes, K.C.; Montagne, J. Drosophila melanogaster: A powerful tiny animal model for the study of metabolic hepatic diseases. Front. Physiol. 2021, 12, 728407. [Google Scholar] [CrossRef]
- Pratomo, A.R.; Salim, E.; Hori, A.; Kuraishi, T. Drosophila as an Animal Model for Testing Plant-Based Immunomodulators. Int. J. Mol. Sci. 2022, 23, 14801. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.L. Olfactory memory formation in Drosophila: From molecular to systems neuroscience. Annu. Rev. Neurosci. 2005, 28, 275–302. [Google Scholar] [CrossRef] [PubMed]
- Montell, C. Drosophila sensory receptors—A set of molecular Swiss Army Knives. Genetics 2021, 217, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Rudrapatna, V.A.; Cagan, R.L.; Das, T.K. Drosophila cancer models. Dev. Dyn. 2012, 241, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Srikrishna, S.; Gupta, S.C. Cancer drug development using Drosophila as an in vivo tool: From bedside to bench and back. Trends Pharmacol. Sci. 2016, 37, 789–806. [Google Scholar] [CrossRef]
- Himalian, R.; Singh, S.K.; Singh, M.P. Ameliorative role of nutraceuticals on neurodegenerative diseases using the Drosophila melanogaster as a discovery model to define bioefficacy. J. Am. Nutr. Assoc. 2022, 41, 511–539. [Google Scholar] [CrossRef]
- Luthra, R.; Roy, A. Role of medicinal plants against neurodegenerative diseases. Curr. Pharm. Biotechnol. 2022, 23, 123–139. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T.; Moreno-Rojas, J.M. Potential health benefits of plant food-derived bioactive components: An overview. Foods 2021, 10, 839. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef]
- Rao, B.N. Bioactive phytochemicals in Indian foods and their potential in health promotion and disease prevention. Asia Pac. J. Clin. Nutr. 2003, 12, 9–22. [Google Scholar]
- AlAli, M.; Alqubaisy, M.; Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Molouki, A.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. Nutraceuticals: Transformation of conventional foods into health promoters/disease preventers and safety considerations. Molecules 2021, 26, 2540. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Saxena, J.; Nema, R.; Singh, D.; Gupta, A. Phytochemistry of medicinal plants. J. Pharmacogn. Phytochem. 2013, 1, 168–182. [Google Scholar]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Gombart, A.F.; Pierre, A.; Maggini, S. A review of micronutrients and the immune system–working in harmony to reduce the risk of infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef]
- Basith, S.; Cui, M.; Hong, S.; Choi, S. Harnessing the therapeutic potential of capsaicin and its analogues in pain and other diseases. Molecules 2016, 21, 966. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef]
- Lu, M.; Chen, C.; Lan, Y.; Xiao, J.; Li, R.; Huang, J.; Huang, Q.; Cao, Y.; Ho, C.-T. Capsaicin—The major bioactive ingredient of chili peppers: Bio-efficacy and delivery systems. Food Funct. 2020, 11, 2848–2860. [Google Scholar] [CrossRef]
- Lopez-Ortiz, C.; Edwards, M.; Natarajan, P.; Pacheco-Valenciana, A.; Nimmakayala, P.; Adjeroh, D.A.; Sirbu, C.; Reddy, U.K. Peppers in Diet: Genome-Wide Transcriptome and Metabolome Changes in Drosophila melanogaster. Int. J. Mol. Sci. 2022, 23, 9924. [Google Scholar] [CrossRef]
- Bertoncini-Silva, C.; Zingg, J.M.; Fassini, P.G.; Suen, V.M.M. Bioactive dietary components—Anti-obesity effects related to energy metabolism and inflammation. BioFactors 2022, 49, 297–321. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The benefits and risks of certain dietary carotenoids that exhibit both anti-and pro-oxidative mechanisms—A comprehensive review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Böhm, V.; Lietz, G.; Olmedilla-Alonso, B.; Phelan, D.; Reboul, E.; Bánati, D.; Borel, P.; Corte-Real, J.; De Lera, A.R.; Desmarchelier, C. From carotenoid intake to carotenoid blood and tissue concentrations–implications for dietary intake recommendations. Nutr. Rev. 2021, 79, 544–573. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Ramachandra, T. Phytochemical and pharmacological importance of turmeric (Curcuma longa): A review. Res. Rev. A J. Pharmacol. 2019, 9, 16–23. [Google Scholar]
- Micek, A.; Godos, J.; Del Rio, D.; Galvano, F.; Grosso, G. Dietary flavonoids and cardiovascular disease: A comprehensive dose–response meta-analysis. Mol. Nutr. Food Res. 2021, 65, 2001019. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Sirén, H. Bioactive phenolic compounds, metabolism and properties: A review on valuable chemical compounds in Scots pine and Norway spruce. Phytochem. Rev. 2019, 18, 623–664. [Google Scholar] [CrossRef]
- Pizarroso, N.A.; Fuciños, P.; Gonçalves, C.; Pastrana, L.; Amado, I.R. A review on the role of food-derived bioactive molecules and the microbiota–gut–brain axis in satiety regulation. Nutrients 2021, 13, 632. [Google Scholar] [CrossRef]
- Tolwinski, N.S. Introduction: Drosophila—A model system for developmental biology. J. Dev. Biol. 2017, 5, 9. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Jeibmann, A.; Paulus, W. Drosophila melanogaster as a model organism of brain diseases. Int. J. Mol. Sci. 2009, 10, 407–440. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. A natural solution for obesity: Bioactives for the prevention and treatment of weight gain. A review. Nutr. Neurosci. 2015, 18, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, R.S. Genetic analysis of insulin signaling in Drosophila. Trends Endocrinol. Metab. 2002, 13, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Ansah, E.; Perrimon, N. Modeling metabolic homeostasis and nutrient sensing in Drosophila: Implications for aging and metabolic diseases. Dis. Models Mech. 2014, 7, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Perrimon, N. What fuels the fly: Energy metabolism in Drosophila and its application to the study of obesity and diabetes. Sci. Adv. 2021, 7, eabg4336. [Google Scholar] [CrossRef]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Baenas, N.; Wagner, A.E. Drosophila melanogaster as an alternative model organism in nutrigenomics. Genes Nutr. 2019, 14, 14. [Google Scholar] [CrossRef]
- Maitra, U.; Stephen, C.; Ciesla, L.M. Drug discovery from natural products–Old problems and novel solutions for the treatment of neurodegenerative diseases. J. Pharm. Biomed. Anal. 2022, 210, 114553. [Google Scholar] [CrossRef]
- Deshpande, P.; Gogia, N.; Singh, A. Exploring the efficacy of natural products in alleviating Alzheimer’s disease. Neural Regen. Res. 2019, 14, 1321. [Google Scholar]
- Lee, S.-H.; Min, K.-J. Drosophila melanogaster as a model system in the study of pharmacological interventions in aging. Transl. Med. Aging 2019, 3, 98–103. [Google Scholar] [CrossRef]
- Lucanic, M.; Lithgow, G.J.; Alavez, S. Pharmacological lifespan extension of invertebrates. Ageing Res. Rev. 2013, 12, 445–458. [Google Scholar] [CrossRef]
- Li, Y.; Peng, Y.; Shen, Y.; Zhang, Y.; Liu, L.; Yang, X. Dietary polyphenols: Regulate the advanced glycation end products-RAGE axis and the microbiota-gut-brain axis to prevent neurodegenerative diseases. Crit. Rev. Food Sci. Nutr. 2022, 1–27, Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Good, T.; Tatar, M. Age-specific mortality and reproduction respond to adult dietary restriction in Drosophila melanogaster. J. Insect Physiol. 2001, 47, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Rand, M.D. Drosophotoxicology: The growing potential for Drosophila in neurotoxicology. Neurotoxicology Teratol. 2010, 32, 74–83. [Google Scholar] [CrossRef]
- Nagpal, I.; Abraham, S.K. Ameliorative effects of gallic acid, quercetin and limonene on urethane-induced genotoxicity and oxidative stress in Drosophila melanogaster. Toxicol. Mech. Methods 2017, 27, 286–292. [Google Scholar] [CrossRef]
- Tello, J.A.; Williams, H.E.; Eppler, R.M.; Steinhilb, M.L.; Khanna, M. Animal models of neurodegenerative disease: Recent advances in fly highlight innovative approaches to drug discovery. Front. Mol. Neurosci. 2022, 15, 883358. [Google Scholar] [CrossRef]
- Brunetti, A.E.; Neto, F.C.; Vera, M.C.; Taboada, C.; Pavarini, D.; Bauermeister, A.; Lopes, N.P. An integrative omics perspective for the analysis of chemical signals in ecological interactions. Chem. Soc. Rev. 2018, 47, 1574–1591. [Google Scholar] [CrossRef]
- Diegelmann, S.; Jansen, A.; Jois, S.; Kastenholz, K.; Escarcena, L.V.; Strudthoff, N.; Scholz, H. The CApillary FEeder assay measures food intake in Drosophila melanogaster. J. Vis. Exp. 2017, 121, e55024. [Google Scholar]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef]
- Forni, C.; Facchiano, F.; Bartoli, M.; Pieretti, S.; Facchiano, A.; D’Arcangelo, D.; Norelli, S.; Valle, G.; Nisini, R.; Beninati, S. Beneficial role of phytochemicals on oxidative stress and age-related diseases. BioMed Res. Int. 2019, 2019, 8748253. [Google Scholar] [CrossRef]
- Abd Rani, N.Z.; Husain, K.; Kumolosasi, E. Moringa genus: A review of phytochemistry and pharmacology. Front. Pharmacol. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Iorjiim, W.M.; Omale, S.; Bagu, G.D.; Gyang, S.S.; Alemika, E.T. Moringa oleifera leaf extract promotes antioxidant, survival, fecundity, and locomotor activities in Drosophila melanogaster. Eur. J. Med. Plants 2020, 31, 30–42. [Google Scholar] [CrossRef]
- Ajagun-Ogunleye, O.; Adedeji, A.; Vicente-Crespo, M. Moringa oleifera ameliorates age-related memory decline and increases endogenous antioxidant response in Drosophila melanogaster exposed to stress. Afr. J. Biomed. Res. 2020, 23, 397–406. [Google Scholar]
- Singh, A.; Duggal, S.; Singh, H.; Singh, J.; Katekhaye, S. Withanolides: Phytoconstituents with significant pharmacological activities. Int. J. Green Pharm. 2010, 4, 229. [Google Scholar] [CrossRef]
- Cabey, K.; Long, D.M.; Law, A.; Gray, N.E.; McClure, C.; Caruso, M.; Lak, P.; Wright, K.M.; Stevens, J.F.; Maier, C.S. Withania somnifera and Centella asiatica Extracts Ameliorate Behavioral Deficits in an In vivo Drosophila melanogaster Model of Oxidative Stress. Antioxidants 2022, 11, 121. [Google Scholar] [CrossRef]
- Holvoet, H.; Long, D.M.; Law, A.; McClure, C.; Choi, J.; Yang, L.; Marney, L.; Poeck, B.; Strauss, R.; Stevens, J.F. Withania somnifera Extracts Promote Resilience against Age-Related and Stress-Induced Behavioral Phenotypes in Drosophila melanogaster; a Possible Role of Other Compounds besides Withanolides. Nutrients 2022, 14, 3923. [Google Scholar] [CrossRef]
- Jebb, S. Obesity: Causes and consequences. Women’s Health Med. 2004, 1, 38–41. [Google Scholar] [CrossRef]
- Chen, J.-C.; Wang, R.; Wei, C.-C. Anti-aging effects of dietary phytochemicals: From Caenorhabditis elegans, Drosophila melanogaster, rodents to clinical studies. Crit. Rev. Food Sci. Nutr. 2022, 1–26, Online ahead of print. [Google Scholar] [CrossRef]
- Sheng, X.; Zhu, Y.; Zhou, J.; Yan, L.; Du, G.; Liu, Z.; Chen, H. Antioxidant effects of caffeic acid lead to protection of drosophila intestinal stem cell aging. Front. Cell Dev. Biol. 2021, 9, 735483. [Google Scholar] [CrossRef]
- Li, Y.M.; Chan, H.Y.E.; Yao, X.Q.; Huang, Y.; Chen, Z.Y. Green tea catechins and broccoli reduce fat-induced mortality in Drosophila melanogaster. J. Nutr. Biochem. 2008, 19, 376–383. [Google Scholar] [CrossRef]
- Wagner, A.E.; Piegholdt, S.; Rabe, D.; Baenas, N.; Schloesser, A.; Eggersdorfer, M.; Stocker, A.; Rimbach, G. Epigallocatechin gallate affects glucose metabolism and increases fitness and lifespan in Drosophila melanogaster. Oncotarget 2015, 6, 30568. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Yang, T.; Korma, S.A.; Sitohy, M.; Abd El-Mageed, T.A.; Selim, S.; Al Jaouni, S.K.; Salem, H.M.; Mahmmod, Y.; Soliman, S.M. Impacts of turmeric and its principal bioactive curcumin on human health: Pharmaceutical, medicinal, and food applications: A comprehensive review. Front. Nutr. 2022, 9, 1040259. [Google Scholar] [CrossRef] [PubMed]
- Suckow, B.K.; Suckow, M.A. Lifespan extension by the antioxidant curcumin in Drosophila melanogaster. Int. J. Biomed. Sci. 2006, 2, 402. [Google Scholar] [PubMed]
- Lee, K.-S.; Lee, B.-S.; Semnani, S.; Avanesian, A.; Um, C.-Y.; Jeon, H.-J.; Seong, K.-M.; Yu, K.; Min, K.-J.; Jafari, M. Curcumin extends life span, improves health span, and modulates the expression of age-associated aging genes in Drosophila melanogaster. Rejuvenation Res. 2010, 13, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Dubiley, T.A.; Rushkevich, Y.E.; Koshel, N.M.; Voitenko, V.P.; Vaiserman, A.M. Life span extension in Drosophila melanogaster induced by morphine. Biogerontology 2011, 12, 179–184. [Google Scholar] [CrossRef]
- Buescher, J.L.; Musselman, L.P.; Wilson, C.A.; Lang, T.; Keleher, M.; Baranski, T.J.; Duncan, J.G. Evidence for transgenerational metabolic programming in Drosophila. Dis. Models Mech. 2013, 6, 1123–1132. [Google Scholar] [CrossRef]
- Colombani, J.; Andersen, D.S. The Drosophila gut: A gatekeeper and coordinator of organism fitness and physiology. Wiley Interdiscip. Rev. Dev. Biol. 2020, 9, e378. [Google Scholar] [CrossRef]
- Baker, K.D.; Thummel, C.S. Diabetic larvae and obese flies—Emerging studies of metabolism in Drosophila. Cell Metab. 2007, 6, 257–266. [Google Scholar] [CrossRef]
- Schlegel, A.; Stainier, D.Y.R. Lessons from “lower” organisms: What worms, flies, and zebrafish can teach us about human energy metabolism. PLoS Genet. 2007, 3, e199. [Google Scholar] [CrossRef]
- Singla, P.; Bardoloi, A.; Parkash, A.A. Metabolic effects of obesity: A review. World J. Diabetes 2010, 1, 76. [Google Scholar] [CrossRef]
- Heinrichsen, E.T.; Zhang, H.; Robinson, J.E.; Ngo, J.; Diop, S.; Bodmer, R.; Joiner, W.J.; Metallo, C.M.; Haddad, G.G. Metabolic and transcriptional response to a high-fat diet in Drosophila melanogaster. Mol. Metab. 2014, 3, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, S. Obesity in midlife: Lifestyle and dietary strategies. Climacteric 2020, 23, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Benítez, V. Phytochemicals: Dietary sources, innovative extraction, and health benefits. Foods 2021, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zheng, S.; Feng, Q.; Zhang, Q.; Xiao, X. Dietary capsaicin and its anti-obesity potency: From mechanism to clinical implications. Biosci. Rep. 2017, 37, BSR20170286. [Google Scholar] [CrossRef]
- Villa-Rivera, M.G.; Ochoa-Alejo, N. Chili pepper carotenoids: Nutraceutical properties and mechanisms of action. Molecules 2020, 25, 5573. [Google Scholar] [CrossRef]
- Chung, J.H.; Manganiello, V.; Dyck, J.R. Resveratrol as a calorie restriction mimetic: Therapeutic implications. Trends Cell Biol. 2012, 22, 546–554. [Google Scholar] [CrossRef]
- Bayliak, M.M.; Shmihel, H.V.; Lylyk, M.P.; Vytvytska, O.M.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Alpha-ketoglutarate attenuates toxic effects of sodium nitroprusside and hydrogen peroxide in Drosophila melanogaster. Environ. Toxicol. Pharmacol. 2015, 40, 650–659. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Ogunsuyi, O.B.; Oboh, G. Purple onion in combination with garlic exerts better ameliorative effects on selected biomarkers in high-sucrose diet-fed fruit fly (Drosophila melanogaster). Comp. Clin. Pathol. 2020, 29, 713–720. [Google Scholar] [CrossRef]
- Baenas, N.; Piegholdt, S.; Schloesser, A.; Moreno, D.A.; García-Viguera, C.; Rimbach, G.; Wagner, A.E. Metabolic activity of radish sprouts derived isothiocyanates in drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 251. [Google Scholar] [CrossRef]
- Rao, M.U.; Sreenivasulu, M.; Chengaiah, B.; Reddy, K.J.; Chetty, C.M. Herbal medicines for diabetes mellitus: A review. Int. J. PharmTech Res. 2010, 2, 1883–1892. [Google Scholar]
- Ugbedeojo, S.P.; Alexander, E.M.; Abigail, O.; Tochukwu, O.C. The phytochemical constituents, hypoglycemic, and antioxidant activities of Senna occidentalis (L.) ethanolic leaf extract in high sucrose diet fed drosophila melanogaster. J. Adv. Biol. Biotechnol. 2021, 24, 48–63. [Google Scholar]
- Saliu, J.A.; Olajuyin, A.M.; Akinnubi, A. Modulatory effect of Artocarpus camansi on ILP-2, InR, and Imp-L2 genes of sucrose–induced diabetes mellitus in Drosophila melanogaster. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 246, 109041. [Google Scholar] [CrossRef]
- De Filippis, F.; Paparo, L.; Nocerino, R.; Della Gatta, G.; Carucci, L.; Russo, R.; Pasolli, E.; Ercolini, D.; Berni Canani, R. Specific gut microbiome signatures and the associated pro-inflamatory functions are linked to pediatric allergy and acquisition of immune tolerance. Nat. Commun. 2021, 12, 5958. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, D.D.; Lakhawat, S.S.; Yasmeen, N.; Pandey, A.; Singla, R.K. Biogenic phytochemicals modulating obesity: From molecular mechanism to preventive and therapeutic approaches. Evid.-Based Complement. Altern. Med. 2022, 2022, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Rai, S.N. The link between obesity, microbiota dysbiosis, and neurodegenerative pathogenesis. Diseases 2021, 9, 45. [Google Scholar] [CrossRef]
- Douglas, A.E. The Drosophila model for microbiome research. Lab Anim. 2018, 47, 157–164. [Google Scholar] [CrossRef]
- Jehrke, L.; Stewart, F.A.; Droste, A.; Beller, M. The impact of genome variation and diet on the metabolic phenotype and microbiome composition of Drosophila melanogaster. Sci. Rep. 2018, 8, 6215. [Google Scholar] [CrossRef]
- Wong, A.C.-N.; Dobson, A.J.; Douglas, A.E. Gut microbiota dictates the metabolic response of Drosophila to diet. J. Exp. Biol. 2014, 217, 1894–1901. [Google Scholar]
- Xu, Z.; Knight, R. Dietary effects on human gut microbiome diversity. Br. J. Nutr. 2015, 113, S1–S5. [Google Scholar] [CrossRef]
- Garcia-Lozano, M.; Haynes, J.; Lopez-Ortiz, C.; Natarajan, P.; Peña-Garcia, Y.; Nimmakayala, P.; Stommel, J.; Alaparthi, S.B.; Sirbu, C.; Balagurusamy, N. Effect of pepper-containing diets on the diversity and composition of gut microbiome of drosophila melanogaster. Int. J. Mol. Sci. 2020, 21, 945. [Google Scholar] [CrossRef]
- Olekhnovich, E.I.; Batotsyrenova, E.G.; Yunes, R.A.; Kashuro, V.A.; Poluektova, E.U.; Veselovsky, V.A.; Ilina, E.N.; Danilenko, V.N.; Klimina, K.M. The effects of Levilactobacillus brevis on the physiological parameters and gut microbiota composition of rats subjected to desynchronosis. Microb. Cell Factories 2021, 20, 226. [Google Scholar] [CrossRef]
- Jiménez-Padilla, Y.; Esan, E.O.; Floate, K.D.; Sinclair, B.J. Persistence of diet effects on the microbiota of Drosophila suzukii (Diptera: Drosophilidae). Can. Entomol. 2020, 152, 516–531. [Google Scholar] [CrossRef]
- Newell, P.D.; Douglas, A.E. Interspecies interactions determine the impact of the gut microbiota on nutrient allocation in Drosophila melanogaster. Appl. Environ. Microbiol. 2014, 80, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Jayarathne, S.; Stull, A.J.; Park, O.H.; Kim, J.H.; Thompson, L.; Moustaid-Moussa, N. Protective effects of anthocyanins in obesity-associated inflammation and changes in gut microbiome. Mol. Nutr. Food Res. 2019, 63, 1900149. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.; Lomis, N.; Prakash, S. A novel polyphenolic prebiotic and probiotic formulation have synergistic effects on the gut microbiota influencing Drosophila melanogaster physiology. Artif. Cells Nanomed. Biotechnol. 2018, 46, 441–455. [Google Scholar] [CrossRef]
- Islam, M.S.; Akhtar, M.M.; Ciavattini, A.; Giannubilo, S.R.; Protic, O.; Janjusevic, M.; Procopio, A.D.; Segars, J.H.; Castellucci, M.; Ciarmela, P. Use of dietary phytochemicals to target inflammation, fibrosis, proliferation, and angiogenesis in uterine tissues: Promising options for prevention and treatment of uterine fibroids? Mol. Nutr. Food Res. 2014, 58, 1667–1684. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Hernández, D.; Lugo-Cervantes, E.D.C.; Escobedo-Reyes, A.; Mojica, L. Black bean (Phaseolus vulgaris L.) polyphenolic extract exerts antioxidant and antiaging potential. Molecules 2021, 26, 6716. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Ji, X.; Xue, J.; Gao, Y.; Zhu, X.; Xiao, G. Cyanidin-3-O-glucoside represses tumor growth and invasion in vivo by suppressing autophagy via inhibition of the JNK signaling pathways. Food Funct. 2021, 12, 387–396. [Google Scholar] [CrossRef]
- Shin, W.S.; Di, J.; Cao, Q.; Li, B.; Seidler, P.M.; Murray, K.A.; Bitan, G.; Jiang, L. Amyloid β-protein oligomers promote the uptake of tau fibril seeds potentiating intracellular tau aggregation. Alzheimer’s Res. Ther. 2019, 11, 86. [Google Scholar] [CrossRef]
- Ahuja, A.; Tyagi, P.K.; Tyagi, S.; Kumar, A.; Kumar, M.; Sharifi-Rad, J. Potential of Pueraria tuberosa (Willd.) DC. to rescue cognitive decline associated with BACE1 protein of Alzheimer’s disease on Drosophila model: An integrated molecular modeling and in vivo approach. Int. J. Biol. Macromol. 2021, 179, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-X.; Zhang, H.; Peng, C. Effects of puerarin on the prevention and treatment of cardiovascular diseases. Front. Pharmacol. 2021, 12, 771793. [Google Scholar] [CrossRef] [PubMed]
- Modi, P.; Mohamad, A.; Phom, L.; Koza, Z.; Das, A.; Chaurasia, R.; Samadder, S.; Achumi, B.; Muralidhara, R.S.P.; Yenisetti, S.C. Understanding Pathophysiology of Sporadic Parkinson’s Disease in Drosophila Model: Potential Opportunities and Notable Limitations; IntechOpen: London, UK, 2016. [Google Scholar]
- Maitra, U.; Harding, T.; Liang, Q.; Ciesla, L. GardeninA confers neuroprotection against environmental toxin in a Drosophila model of Parkinson’s disease. Commun. Biol. 2021, 4, 162. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Y.H.; Naz, F.; Mantasha, I.; Shahid, M. Lemongrass Extract Alleviates Oxidative Stress and Delayed the Loss of Climbing Ability in Transgenic Drosophila Model of Parkinson’s Disease. Lett. Drug Des. Discov. 2021, 18, 987–997. [Google Scholar] [CrossRef]
- Ssempijja, F.; Dare, S.S.; Bukenya, E.E.; Kasozi, K.I.; Kenganzi, R.; Fernandez, E.M.; Vicente-Crespo, M. Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models. J. Evid. -Based Integr. Med. 2023, 28, 2515690X231160191. [Google Scholar] [CrossRef]
- Ing-Simmons, E.; Vaid, R.; Bing, X.Y.; Levine, M.; Mannervik, M.; Vaquerizas, J.M. Independence of chromatin conformation and gene regulation during Drosophila dorsoventral patterning. Nat. Genet. 2021, 53, 487–499. [Google Scholar] [CrossRef]
- Rivera, J.; Keränen, S.V.E.; Gallo, S.M.; Halfon, M.S. REDfly: The transcriptional regulatory element database for Drosophila. Nucleic Acids Res. 2019, 47, D828–D834. [Google Scholar] [CrossRef]
- Fear, J.M.; León-Novelo, L.G.; Morse, A.M.; Gerken, A.R.; Van Lehmann, K.; Tower, J.; Nuzhdin, S.V.; McIntyre, L.M. Buffering of genetic regulatory networks in Drosophila melanogaster. Genetics 2016, 203, 1177–1190. [Google Scholar] [CrossRef]
- Zhang, Z.-G.; Niu, X.-Y.; Lu, A.-P.; Xiao, G.G. Effect of curcumin on aged Drosophila Melanogaster: A pathway prediction analysis. Chin. J. Integr. Med. 2015, 21, 115–122. [Google Scholar] [CrossRef]
- Park, J.H.; Jung, J.W.; Ahn, Y.-J.; Kwon, H.W. Neuroprotective properties of phytochemicals against paraquat-induced oxidative stress and neurotoxicity in Drosophila melanogaster. Pestic. Biochem. Physiol. 2012, 104, 118–125. [Google Scholar] [CrossRef]
- Jensen, H.; Scott, I.; Sims, S.; Trudeau, V.; Arnason, J. The effect of a synergistic concentration of a Piper nigrum extract used in conjunction with pyrethrum upon gene expression in Drosophila melanogaster. Insect Mol. Biol. 2006, 15, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Adedara, A.O.; Babalola, A.D.; Stephano, F.; Awogbindin, I.O.; Olopade, J.O.; Rocha, J.B.; Whitworth, A.J.; Abolaji, A.O. An assessment of the rescue action of resveratrol in parkin loss of function-induced oxidative stress in Drosophila melanogaster. Sci. Rep. 2022, 12, 3922. [Google Scholar] [CrossRef] [PubMed]
- Staats, S.; Wagner, A.E.; Kowalewski, B.; Rieck, F.T.; Soukup, S.T.; Kulling, S.E.; Rimbach, G. Dietary resveratrol does not affect life span, body composition, stress response, and longevity-related gene expression in Drosophila melanogaster. Int. J. Mol. Sci. 2018, 19, 223. [Google Scholar] [CrossRef] [PubMed]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in intestinal mucosal defense and inflammation: Learning from clinical and experimental studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Shin, J.H.; Seeley, R.J. Reg3 proteins as gut hormones? Endocrinology 2019, 160, 1506–1514. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Li, L.; Chen, C.; Xiang, Q.; Fan, S.; Xiao, T.; Chen, Y.; Zheng, D. Transient Receptor Potential Cation Channel Subfamily V Member 1 Expression Promotes Chemoresistance in Non-Small-Cell Lung Cancer. Front. Oncol. 2022, 12, 773654. [Google Scholar] [CrossRef]
- Chang, A.E.; Vaughan, A.G.; Wilson, R.I. A mechanosensory circuit that mixes opponent channels to produce selectivity for complex stimulus features. Neuron 2016, 92, 888–901. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant/Fruit | Phytochemicals | Description | Beneficial Activity in Human Related Disease |
|---|---|---|---|
| Pepper | Capsaicinoids Carotenoids | Promote fatty acid oxidation and antioxidant activity | Obesity |
| Curcuma domestica | Curcumin | Antioxidant, inhibition of lipid peroxide-induce DNA damage | Cancer Lifespan |
| Moringa oleifera | Polyphenols Flavonoids Tannins | Antioxidant response by glutathione-S-transferase and catalase | Aging Lifespan |
| Withania somnifera | Withanolides | Reduces oxidative stress | Aging |
| Coffee | Phenolic caffeic | Prevent dysregulation of regeneration and differentiation cells | Lifespan |
| Camellia sinensis | Catechins | Upregulates superoxide dismutase and catalase | Lifespan |
| Papaver somniferum | Morphine | Stimulate growth and nervous fibers | Lifespan |
| Red grapes | Resveratrol | Antioxidant activity reduces lipid accumulation | Obesity |
| Radish sprouts | Glucosinolates | Influence energy metabolism and the expression of insulin signaling gene | Diabetes |
| Phaseolus vulgaris | Cyanidin-3-O-glucoside | Reduces tumor proliferation and blocks autophagy | Cancer |
| Pueraria tuberosa | Puerarin | Inhibit the development of beta-amyloid plaques | Alzheimer |
| Lemongrass | Flavonoids Tannins Alkaloids Glycosides | Decreases ROS levels | Parkinson |
| Imperata cylindrica | Alkaloids Polyphenolic compounds | Inhibition of voltage-gated sodium channels and reduce inflammation | Epilepsy |
| Gene ID | Gene Name | Annotation | Human Orthologue | Disease | Plant Extract |
|---|---|---|---|---|---|
| FBgn0003079 | Raf | Encodes a serine-threonine protein kinase; it activates the MEK/ERK pathway to regulate cell proliferation. | Raf-1 | Cancer | Phaseolus vulgaris |
| FBgn0032049 | Bace | Beta-site APP-cleaving enzyme encodes an aspartic protease that cleaves amyloid precursor proteins. | BACE1 | Alzheimer | Pueraria tuberosa |
| FBgn0026420 | SNCA | Engineered foreign gene involves several processes, including negative and positive transport regulation and protein metabolic process. | SNCA | Parkinson | Lemongrass |
| FBgn0285944 | para | A gene is required for locomotor activity. It encodes an α-subunit of voltage-gated sodium channels. | SCN | Epilepsy | Imperatacylindrica |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Ortiz, C.; Gracia-Rodriguez, C.; Belcher, S.; Flores-Iga, G.; Das, A.; Nimmakayala, P.; Balagurusamy, N.; Reddy, U.K. Drosophila melanogaster as a Translational Model System to Explore the Impact of Phytochemicals on Human Health. Int. J. Mol. Sci. 2023, 24, 13365. https://doi.org/10.3390/ijms241713365
Lopez-Ortiz C, Gracia-Rodriguez C, Belcher S, Flores-Iga G, Das A, Nimmakayala P, Balagurusamy N, Reddy UK. Drosophila melanogaster as a Translational Model System to Explore the Impact of Phytochemicals on Human Health. International Journal of Molecular Sciences. 2023; 24(17):13365. https://doi.org/10.3390/ijms241713365
Chicago/Turabian StyleLopez-Ortiz, Carlos, Celeste Gracia-Rodriguez, Samantha Belcher, Gerardo Flores-Iga, Amartya Das, Padma Nimmakayala, Nagamani Balagurusamy, and Umesh K. Reddy. 2023. "Drosophila melanogaster as a Translational Model System to Explore the Impact of Phytochemicals on Human Health" International Journal of Molecular Sciences 24, no. 17: 13365. https://doi.org/10.3390/ijms241713365
APA StyleLopez-Ortiz, C., Gracia-Rodriguez, C., Belcher, S., Flores-Iga, G., Das, A., Nimmakayala, P., Balagurusamy, N., & Reddy, U. K. (2023). Drosophila melanogaster as a Translational Model System to Explore the Impact of Phytochemicals on Human Health. International Journal of Molecular Sciences, 24(17), 13365. https://doi.org/10.3390/ijms241713365