
HCN2 Channels in the Ventral Hippocampal CA1 Regulate Nociceptive Hypersensitivity in Mice
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
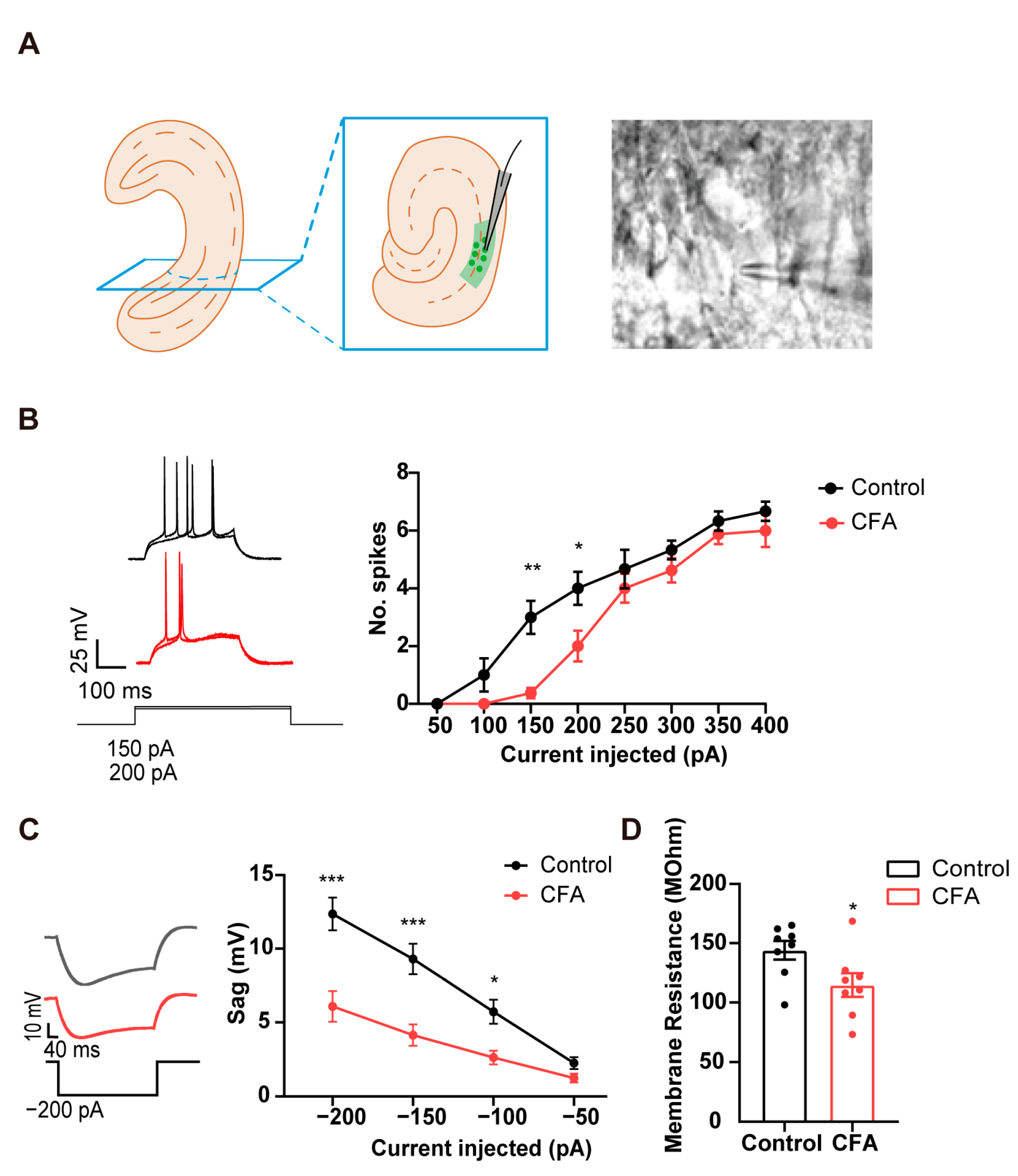
2.1. Decreased Neuronal Excitability and Sag Potential in vCA1 Pyramidal Neurons in Mice with CFA-Induced Inflammatory Pain
2.2. Decreased Expression of HCN2 in vCA1 of Mice with CFA-Induced Inflammatory Pain
2.3. Up/Down-Regulation of HCN2 in vCA1 Bidirectionally Modulates Nociceptive Hypersensitivity
2.4. Down-Regulation of HCN2 Decreased the Excitability of Pyramidal Neurons in vCA1 of Mice
2.5. Over-Expression of HCN2 in vCA1 Has Limited Effect on Anxiety-like Behaviors and Cognitive Impairment in Mice Injected with CFA
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Inflammatory Pain Model
4.3. Electrophysiological Recordings Ex Vivo
4.4. Immunostaining
4.5. Western Blot
4.6. HCN1/2 Regulation
4.7. Stereotaxic Injection
4.8. Thermal Pain Measurement
4.9. Object–Place Recognition (OPR)
4.10. Open Field Test (OFT)
4.11. The Elevated Plus Maze Test (EPM)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Svensson, C.I.; Zattoni, M.; Serhan, C.N. Lipoxins and aspirin-triggered lipoxin inhibit inflammatory pain processing. J. Exp. Med. 2007, 204, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Wang, S.; Shao, F.; Fang, J.; Xu, Y.; Wang, W.; Sun, H.; Liu, X.; Du, J.; Fang, J. Electroacupuncture stimulation alleviates CFA-induced inflammatory pain via suppressing P2X3 expression. Nat. Rev. Drug. Discov. 2019, 20, 3248. [Google Scholar] [CrossRef]
- Dougados, M.; Etcheto, A.; Molto, A.; Alonso, S.; Bouvet, S.; Daurès, J.-P.; Landais, P.; D’agostino, M.-A.; Berenbaum, F.; Breban, M.; et al. Clinical presentation of patients suffering from recent onset chronic inflammatory back pain suggestive of spondyloarthritis: The DESIR cohort. Joint. Bone. Spine. 2017, 82, 345–351. [Google Scholar] [CrossRef]
- Lee, Y.C. Effect and treatment of chronic pain in inflammatory arthritis. Curr. Rheumatol. Rep. 2013, 15, 300. [Google Scholar] [CrossRef] [PubMed]
- Siler, S.; Borneman, T.; Ferrell, B. Pain and Suffering. Semin. Oncol. Nurs. 2019, 35, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Bonezzi, C.; Fornasari, D.; Cricelli, C.; Magni, A.; Ventriglia, G. Not All Pain is Created Equal: Basic Definitions and Diagnostic Work-Up. Pain. Ther. 2020, 9, 1–15. [Google Scholar] [CrossRef]
- Anna, K.S.; Anna, J.; Natalia, H.; Konrad, R. Neuropathic pain and chronic pain as an underestimated interdisciplinary problem. Int. J. Occup. Med. Environ. Health 2022, 4, 143474. [Google Scholar] [CrossRef]
- Bedini, A.; Spampinato, S.M. Innovative opioid peptides and biased agonism: Novel avenues for more effective and safer analgesics to treat chronic pain. Curr. Med. Chem. 2018, 25, 3895–3916. [Google Scholar] [CrossRef]
- Marret, E.; Kurdi, O.; Zufferey, P.; Bonnet, F. Effects of nonsteroidal anti-inflammatory drugs on patient-controlled analgesia morphine side effects: Meta-analysis of randomized controlled trials. Anesthesiology 2005, 102, 1249–1260. [Google Scholar] [CrossRef]
- Nakamoto, K.; Nishinaka, T.; Sato, N.; Mankura, M.; Koyama, Y.; Kasuya, F.; Tokuyama, S. Hypothalamic GPR40 signaling activated by free long chain fatty acids suppresses CFA-induced inflammatory chronic pain. PLoS ONE 2013, 8, e81563. [Google Scholar] [CrossRef]
- Hayes, D.J.; Chen, D.Q.; Zhong, J.; Lin, A.; Behan, B.; Walker, M.; Hodaie, M. Affective circuitry alterations in patients with trigeminal neuralgia. Front. Neuroanat. 2017, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, A.; Falfán-Melgoza, C.; Leixner, S.; Becker, R.; Singaravelu, S.K.; Sack, M.; Sartorius, A.; Spanagel, R.; Weber-Fahr, W. Longitudinal structural and functional brain network alterations in a mouse model of neuropathic pain. Neuroscience 2018, 387, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Vaculik, M.F.; Noorani, A.; Hung, P.S.; Hodaie, M. Selective hippocampal subfield volume reductions in classic trigeminal neuralgia. Neuroimage Clin. 2019, 23, 101911. [Google Scholar] [CrossRef] [PubMed]
- Manzhulo, I.; Manzhulo, O.; Tyrtyshnaia, A.; Ponomarenko, A.; Konovalova, S.; Ermolenko, E.; Milkina, E.; Starinets, A. Modulation of Hippocampal Astroglial Activity by Synaptamide in Rats with Neuropathic Pain. Brain. Sci. 2021, 11, 1561. [Google Scholar] [CrossRef]
- Zheng, J.; Jiang, Y.-Y.; Xu, L.-C.; Ma, L.-Y.; Liu, F.-Y.; Cui, S.; Cai, J.; Liao, F.-F.; Wan, Y.; Yi, M. Adult Hippocampal neurogenesis along the dorsoventral axis contributes differentially to environmental enrichment combined with voluntary exercise in alleviating chronic inflammatory pain in mice. J. Neurosci. 2017, 37, 4145–4157. [Google Scholar] [CrossRef]
- Jiang, Y.-Y.; Shao, S.; Zhang, Y.; Zheng, J.; Chen, X.; Cui, S.; Liu, F.-Y.; Wan, Y.; Yi, M. Neural pathways in medial septal cholinergic modulation of chronic pain: Distinct contribution of the anterior cingulate cortex and ventral hippocampus. Pain 2018, 159, 1550–1561. [Google Scholar] [CrossRef]
- Ma, L.; Yue, L.; Zhang, Y.; Wang, Y.; Han, B.; Cui, S.; Liu, F.-Y.; Wan, Y.; Yi, M. Spontaneous pain disrupts ventral hippocampal CA1-infralimbic cortex connectivity and modulates pain progression in rats with peripheral inflammation. Cell Rep. 2019, 29, 1579–1593. [Google Scholar] [CrossRef]
- Shao, S.; Zheng, Y.; Fu, Z.; Wang, J.; Zhang, Y.; Wang, C.; Qi, X.; Gong, T.; Ma, L.; Lin, X.; et al. Ventral hippocampal CA1 modulates pain behaviors in mice with peripheral inflammation. Cell Rep. 2023, 42, 112017. [Google Scholar] [CrossRef]
- Narayanan, R.; Johnston, D. The Ih Channel Mediates Location Dependence and Plasticity of Intrinsic Phase Response in Rat Hippocampal Neurons. J. Neurosci. 2008, 28, 5846–5860. [Google Scholar] [CrossRef]
- Magee, J.C. Dendritic hyperpolarization-activated currents modify the integrative properties of hippocampal CA1 pyramidal neurons. J. Neurosci. 1998, 18, 7613–7624. [Google Scholar] [CrossRef]
- Tsay, D.; Dudman, J.T.; Siegelbaum, S.A. HCN1 channels constrain synaptically evoked Ca2+ spikes in distal dendrites of CA1 pyramidal neurons. Neuron 2007, 56, 1076–1089. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Q.; Sun, Q.; Tu, H.Y.; Wan, Y. Characteristics of HCN Channels and Their Participation in Neuropathic Pain. Neurochem. Res. 2008, 33, 1979–1989. [Google Scholar] [CrossRef]
- Robinson, R.B.; Siegelbaum, S.A. Hyperpolarization-activated cation currents: From molecules to physiological function. Annu. Rev. Physiol. 2003, 65, 453–480. [Google Scholar] [CrossRef] [PubMed]
- Biel, M.; Wahl-Schott, C.; Michalakis, S.; Zong, X. Hyperpolarization-activated cation channels: From genes to function. Physiol. Rev. 2009, 89, 847–885. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; Johnston, D. Long-Term Potentiation in Rat Hippocampal Neurons Is Accompanied by Spatially Widespread Changes in Intrinsic Oscillatory Dynamics and Excitability. Neuron 2007, 56, 1061–1075. [Google Scholar] [CrossRef]
- Kozák, G. Insights on the Role of Thalamocortical HCN Channels in Absence Epilepsy. J. Neurosci. 2019, 39, 578–580. [Google Scholar] [CrossRef]
- Lin, W.; Qin, J.; Ni, G.; Li, Y.; Xie, H.; Yu, J.; Li, H.; Sui, L.; Guo, Q.; Fang, Z.; et al. Downregulation of hyperpolarization-activated cyclic nucleotide-gated channels (HCN) in the hippocampus of patients with medial temporal lobe epilepsy and hippocampal sclerosis (MTLE-HS). Hippocampus 2020, 30, 1112–1126. [Google Scholar] [CrossRef]
- Tibbs, G.R.; Rowley, T.J.; Sanford, R.L.; Herold, K.F.; Proekt, A.; Hemmings, H.C.; Andersen, O.S.; Goldstein, P.A.; Flood, P.D. HCN1 channels as targets for anesthetic and nonanesthetic propofol analogs in the amelioration of mechanical and thermal hyperalgesia in a mouse model of neuropathic pain. J. Pharmacol. Exp. Ther. 2013, 345, 363–373. [Google Scholar] [CrossRef]
- Zhang, S.; You, Z.; Wang, S.; Yang, J.; Yang, L.; Sun, Y.; Mi, W.; Yang, L.; McCabe, M.F.; Shen, S.; et al. Neuropeptide S modulates the amygdaloidal HCN activities (I) in rats: Implication in chronic pain. Neuropharmacology 2016, 105, 420–433. [Google Scholar] [CrossRef]
- Young, G.T.; Emery, E.C.; Mooney, E.R.; Tsantoulas, C.; McNaughton, P.A. Inflammatory and neuropathic pain are rapidly suppressed by peripheral block of hyperpolarisation-activated cyclic nucleotide-gated ion channels. Pain 2014, 155, 1708–1719. [Google Scholar] [CrossRef]
- Daniel, C.C.; Peter, A.G. Non-canonical molecular targets for novel analgesics: Intracellular calcium and HCN channels. Curr. Neuropharmacol. 2021, 19, 1937–1951. [Google Scholar] [CrossRef]
- Emery, E.C.; Young, G.T.; Berrocoso, E.M.; Chen, L.; McNaughton, P.A. HCN2 ion channels play a central role in inflammatory and neuropathic pain. Science 2011, 333, 1462–1466. [Google Scholar] [CrossRef]
- Momin, A.; Cadiou, H.; Mason, A. Role of the hyperpolarization-activated current Ih in somatosensory neurons. J. Physiol. 2008, 586, 5911–5929. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, M.; Stegmayr, C.; Balfanz, S.; Baumann, A. Loss of HCN2 in Dorsal Hippocampus of Young Adult Mice Induces Specific Apoptosis of the CA1 Pyramidal Neuron Layer. Int. J. Mol. Sci. 2021, 22, 6699. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Chang, P.Y.; Johnston, D. Enhancement of dorsal hippocampal activity by knockdown of HCN1 channels leads to anxiolytic- and antidepressant-like behaviors. Neuron 2021, 75, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-Q.; Xing, G.-G.; Wang, S.-L.; Tu, H.-Y.; Chi, Y.-N.; Li, J.; Liu, F.-Y.; Han, J.-S.; Wan, Y. Axonal accumulation of hyperpolarization-activated cyclic nucleotide-gated cation channels contributes to mechanical allodynia after peripheral nerve injury in rat. Pain 2008, 137, 495–506. [Google Scholar] [CrossRef]
- Price, D.D. Psychological and neural mechanisms of the affective dimension of pain. Science 2000, 288, 1769–1772. [Google Scholar] [CrossRef]
- Bair, M.J.; Robinson, R.L.; Katon, W.; Kroenke, K. Depression and pain comorbidity: A literature review. Arch. Intern. Med. 2003, 163, 2433–2445. [Google Scholar] [CrossRef]
- Fiore, N.T.; Austin, P.J. Peripheral Nerve Injury Triggers Neuroinflammation in the Medial Prefrontal Cortex and Ventral Hippocampus in a Subgroup of Rats with Coincident Affective Behavioural Changes. Neuroscience 2019, 416, 147–167. [Google Scholar] [CrossRef]
- Maroso, M.; Szabo, G.G.; Kim, H.K.; Alexander, A.; Bui, A.D.; Lee, S.-H.; Lutz, B.; Soltesz, I. Cannabinoid control of learning and memory through HCN channels. Neuron 2016, 89, 1059–1073. [Google Scholar] [CrossRef]
- Vasnik, S.; Sikdar, S.K. Cholinergic receptor-independent modulation of intrinsic resonance in the rat subiculum neurons through inhibition of hyperpolarization-activated cyclic nucleotide-gated channels. Acta Physiol. 2021, 231, e13603. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.S.; Shigemoto, R.; Mercer, J.N.; Surmeier, D.J. HCN2 and HCN1 channels govern the regularity of autonomous pacemaking and synaptic resetting in globus pallidus neurons. J. Neurosci. 2004, 24, 9921–9932. [Google Scholar] [CrossRef] [PubMed]
- Roth, F.C.; Hu, H. An axon-specific expression of HCN channels catalyzes fast action potential signaling in GABAergic interneurons. Nat. Commun. 2020, 11, 2248. [Google Scholar] [CrossRef] [PubMed]
- Hammelmann, V.; Stieglitz, M.S.; Hülle, H.; Le Meur, K.; Kass, J.; Brümmer, M.; Gruner, C.; Rötzer, R.D.; Fenske, S.; Hartmann, J.; et al. Abolishing cAMP sensitivity in HCN2 pacemaker channels induces generalized seizures. JCI Insight. 2019, 4, e126418. [Google Scholar] [CrossRef]
- Balducci, V.; Credi, C.; Sacconi, L.; Romanelli, M.N.; Sartiani, L.; Cerbai, E. The HCN channel as a pharmacological target: Why, where, and how to block it. Prog. Biophys. Mol. Biol. 2021, 166, 173–181. [Google Scholar] [CrossRef]
- Tsantoulas, C.; Mooney, E.R.; McNaughton, P.A. HCN2 ion channels: Basic science opens up possibilities for therapeutic intervention in neuropathic pain. Biochem. J. 2016, 473, 2717–2736. [Google Scholar] [CrossRef]
- Jansen, L.-A.R.; Forster, L.A.; Smith, X.L.; Rubaharan, M.; Murphy, A.Z.; Baro, D.J. Changes in peripheral HCN2 channels during persistent inflammation. Channels 2021, 15, 165–179. [Google Scholar] [CrossRef]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Szok, D.; Tajti, J.; Nyári, A.; Vécsei, L. Therapeutic Approaches for Peripheral and Central Neuropathic Pain. Behav. Neurol. 2019, 2019, 8685954. [Google Scholar] [CrossRef]
- Schnorr, S.; Eberhardt, M.; Kistner, K. HCN2 channels account for mechanical (but not heat) hyperalgesia during long-standing inflammation. Pain 2014, 155, 1079–1090. [Google Scholar] [CrossRef]
- Byczkowicz, N.; Eshra, A.; Montanaro, J.; Trevisiol, A.; Hirrlinger, J.; Kole, M.H.; Shigemoto, R.; Hallermann, S. HCN channel-mediated neuromodulation can control action potential velocity and fidelity in central axons. Elife 2019, 8, e42766. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, X.; Yao, K.; Sun, Y.; Jiang, F.; Yan, H.; Mao, P.; Fan, S.; Wei, X.; Liu, Y.; et al. HCN channel antagonist ZD7288 ameliorates neuropathic pain and associated depression. Brain. Res. 2019, 1717, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Omrani, A.; van der Vaart, T.; Mientjes, E.; van Woerden, G.M.; Hojjati, M.R.; Li, K.W.; Gutmann, D.H.; Levelt, C.N.; Smit, A.B.; Silva, A.J.; et al. HCN channels are a novel therapeutic target for cognitive dysfunction in Neurofibromatosis type 1. Mol. Psychiatry 2015, 20, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Gergues, M.M.; Han, K.J.; Choi, H.S.; Brown, B.; Clausing, K.J.; Turner, V.S.; Vainchtein, I.D.; Molofsky, A.V.; Kheirbek, M.A. Circuit and molecular architecture of a ventral hippocampal network. Nat. Neurosci. 2020, 23, 1444–1452. [Google Scholar] [CrossRef]
- Aleksic, M.; Brkic, Z.; Petrovic, Z.; Francija, E.; Lukic, I.; Adzic, M. Sex-specific contribution of glucocorticoid receptor alpha isoforms to anxiety and depressive-like behavior in mice. J. Neurosci. Res. 2022, 100, 1239–1253. [Google Scholar] [CrossRef]
- Fukutoku, T.; Kumagai, G.; Fujita, T.; Sasaki, A.; Wada, K.; Liu, X.; Tanaka, T.; Kudo, H.; Asari, T.; Nikaido, Y.; et al. Sex-Related Differences in Anxiety and Functional Recovery after Spinal Cord Injury in Mice. J. Neurotrauma. 2020, 37, 2235–2243. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef]
- Krause, W.C.; Rodriguez, R.; Gegenhuber, B.; Matharu, N.; Rodriguez, A.N.; Padilla-Roger, A.M.; Toma, K.; Herber, C.B.; Correa, S.M.; Duan, X.; et al. Oestrogen engages brain MC4R signalling to drive physical activity in female mice. Nature 2021, 599, 131–135. [Google Scholar] [CrossRef]
- Grégoire, S.; Jang, S.H.; Szyf, M.; Stone, L.S. Prenatal maternal stress is associated with increased sensitivity to neuropathic pain and sex-specific changes in supraspinal mRNA expression of epigenetic- and stress-related genes in adulthood. Behav. Brain. Res. 2020, 380, 112396. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Li, X.; Liu, X.; Qin, C. Early-Life Neglect Alters Emotional and Cognitive Behavior in a Sex-Dependent Manner and Reduces Glutamatergic Neuronal Excitability in the Prefrontal Cortex. Front. Psychiatry 2021, 11, 572224. [Google Scholar] [CrossRef]
- Whissell, P.D.; Rosenzweig, S.; Lecker, I.; Wang, D.-S.; Wojtowicz, J.M.; Orser, B.A. Gamma aminobutyric acid type A receptors that contain the delta subunit promote memory and neurogenesis in the dentate gyrus. Ann. Neurol. 2013, 74, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Pennington, Z.T.; Dong, Z.; Feng, Y.; Vetere, L.M.; Page-Harley, L.; Shuman, T.; Cai, D.J. ezTrack: An open-source video analysis pipeline for the investigation of animal behavior. Sci. Rep. 2019, 9, 19979. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Shao, S.; Zhang, Y.; Yuan, S.; Xing, Y.; Wang, J.; Qi, X.; Cui, K.; Tong, J.; Liu, F.; et al. HCN2 Channels in the Ventral Hippocampal CA1 Regulate Nociceptive Hypersensitivity in Mice. Int. J. Mol. Sci. 2023, 24, 13823. https://doi.org/10.3390/ijms241813823
Zheng Y, Shao S, Zhang Y, Yuan S, Xing Y, Wang J, Qi X, Cui K, Tong J, Liu F, et al. HCN2 Channels in the Ventral Hippocampal CA1 Regulate Nociceptive Hypersensitivity in Mice. International Journal of Molecular Sciences. 2023; 24(18):13823. https://doi.org/10.3390/ijms241813823
Chicago/Turabian StyleZheng, Yawen, Shan Shao, Yu Zhang, Shulu Yuan, Yuanwei Xing, Jiaxin Wang, Xuetao Qi, Kun Cui, Jifu Tong, Fengyu Liu, and et al. 2023. "HCN2 Channels in the Ventral Hippocampal CA1 Regulate Nociceptive Hypersensitivity in Mice" International Journal of Molecular Sciences 24, no. 18: 13823. https://doi.org/10.3390/ijms241813823
APA StyleZheng, Y., Shao, S., Zhang, Y., Yuan, S., Xing, Y., Wang, J., Qi, X., Cui, K., Tong, J., Liu, F., Cui, S., Wan, Y., & Yi, M. (2023). HCN2 Channels in the Ventral Hippocampal CA1 Regulate Nociceptive Hypersensitivity in Mice. International Journal of Molecular Sciences, 24(18), 13823. https://doi.org/10.3390/ijms241813823