
Gintonin-Induced Wound-Healing-Related Responses Involve Epidermal-Growth-Factor-like Effects in Keratinocytes

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Gintonin-Mediated Phosphorylation of EGF Receptor
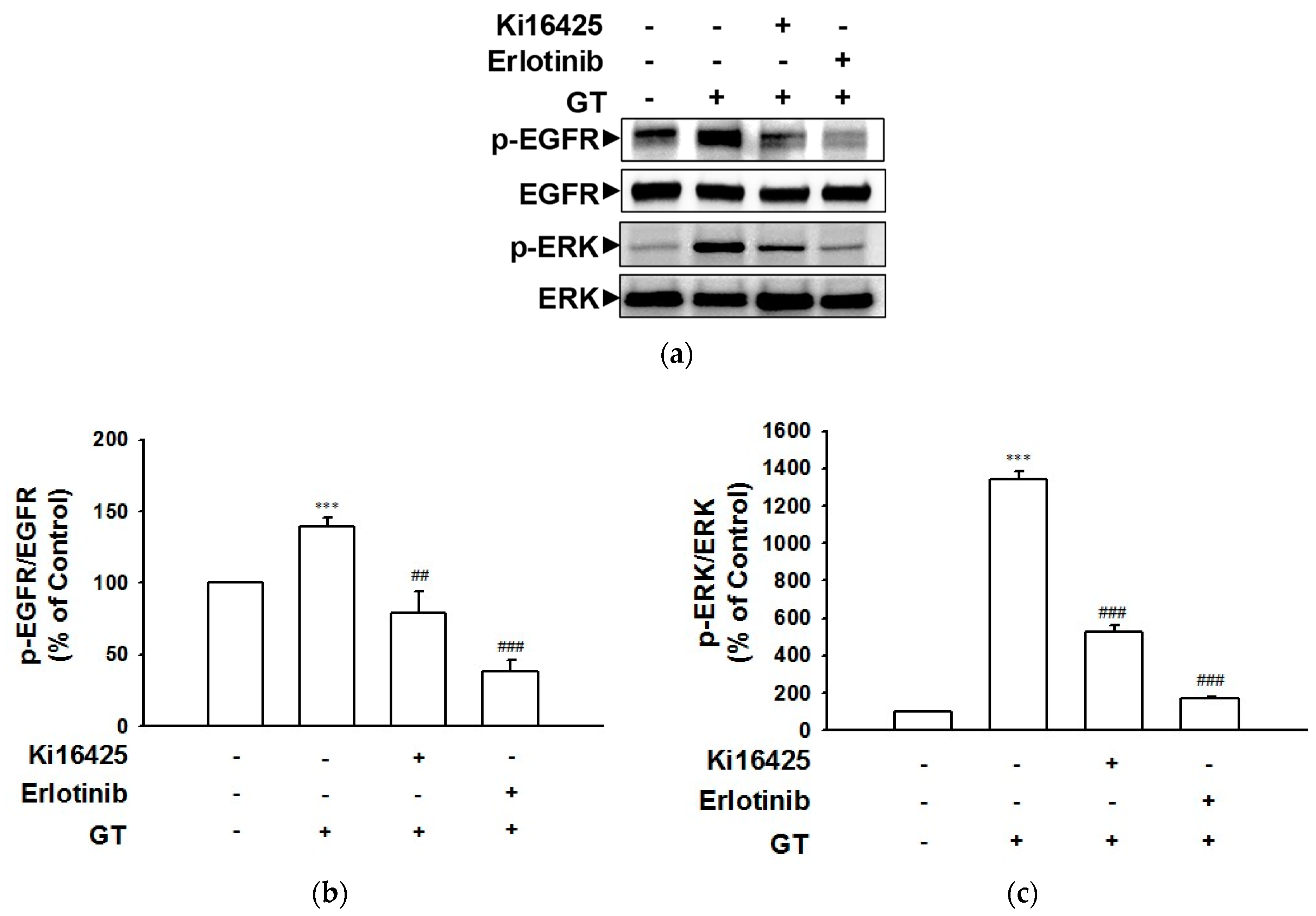
2.2. Gintonin-Induced Phosphorylation of the EGF Receptor and ERK Involves the LPA Receptor as Well as the EGF Receptor
2.3. Involvement of EGF Receptor Signaling in Gintonin-Mediated Proliferation and Migration of HaCaT Cells
2.4. Gintonin-Induced Expression and Release of HB-EGF in HaCaT Cells
2.5. The Relationship of EGF Receptor and Expression of HB-EGF in HaCaT Cells Exposed to Gintonin
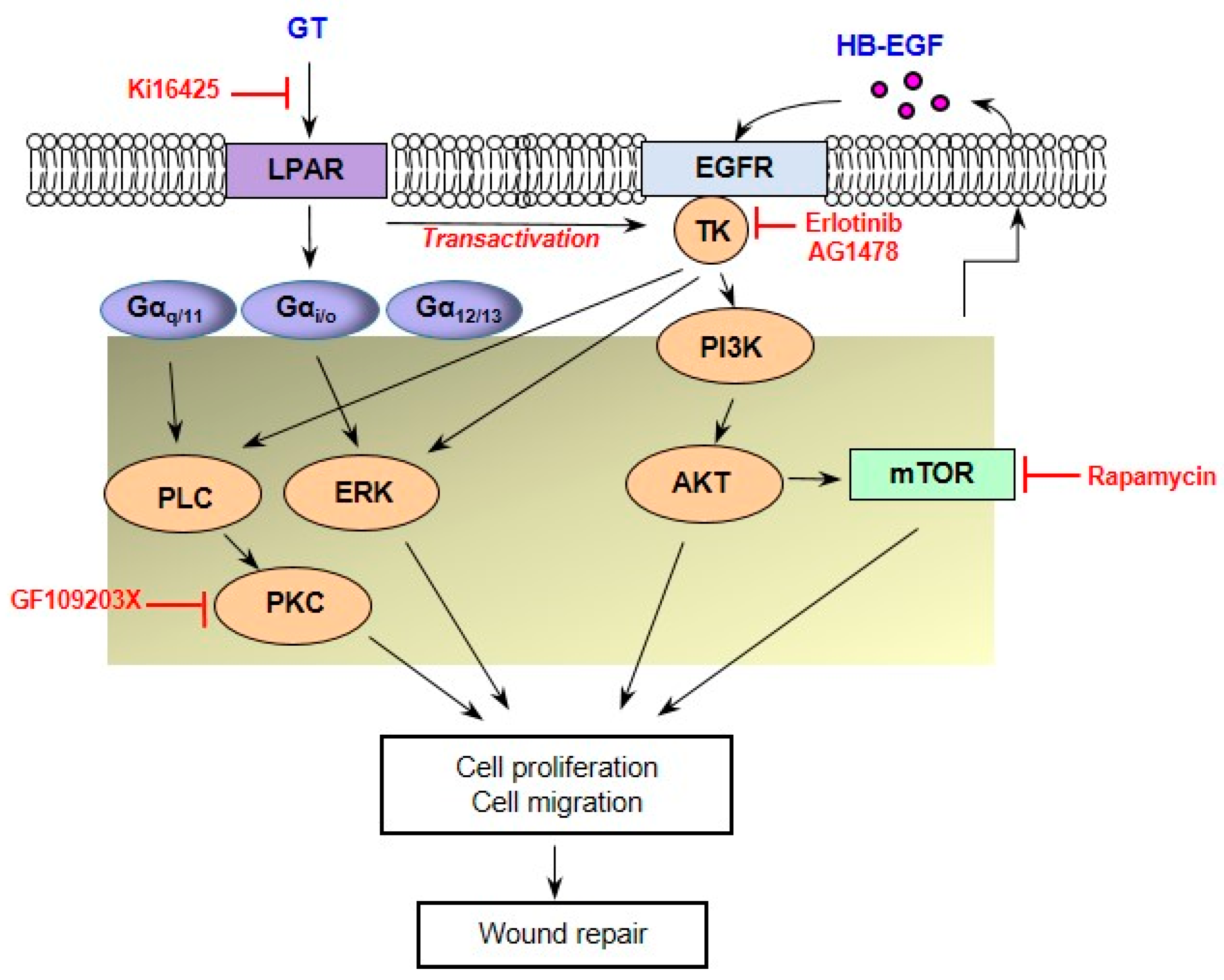
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Proliferation Assay
4.4. Migration Assay
4.5. HB-EGF ELISA
4.6. Immunoblotting
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seeger, M.A.; Paller, A.S. The roles of growth factors in keratinocyte migration. Adv. Wound Care 2015, 4, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Graves, D.T. Keratinocyte function in normal and diabetic wounds and modulation by FOXO1. J. Diabetes Res. 2020, 2020, 3714704. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Zeng, C.; Li, W.; Ke, C.; Xu, S. Concentrated growth factor promotes wound healing potential of HaCaT Cells by activating the RAS signaling pathway. Front. Biosci. Landmark Ed. 2022, 27, 319. [Google Scholar] [CrossRef] [PubMed]
- El-Serafi, A.T.; El-Serafi, I.; Steinvall, I.; Sjöberg, F.; Elmasry, M. A systematic review of keratinocyte secretions: A regenerative perspective. Int. J. Mol. Sci. 2022, 23, 7934. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Li, Y.; Fan, J.; Chen, M.; Li, W.; Woodley, D.T. Transforming growth factor-alpha: A major human serum factor that promotes human keratinocyte migration. J. Investig. Dermatol. 2006, 126, 2096–2105. [Google Scholar] [CrossRef]
- Oda, K.; Matsuoka, Y.; Funahashi, A.; Kitano, H. A comprehensive pathway map of epidermal growth factor receptor signaling. Mol. Syst. Biol. 2005, 1, 2005-0010. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, F.; Forrester, S.J.; Eguchi, S.; Zhang, M.Z.; Harris, R.C. Expression and function of the epidermal growth factor receptor in physilology and disease. Physiol. Rev. 2016, 96, 1025–1069. [Google Scholar] [CrossRef]
- Shakhakarmi, K.; Seo, J.E.; Lamichhane, S.; Thapa, C.; Lee, S. EGF, a veteran of wound healing: Highlights on its mode of action, clinical applications with focus on wound treatment, and recent drug delivery strategies. Arch. Pharm. Res. 2023, 46, 299–322. [Google Scholar] [CrossRef]
- Kobayashi, R.; Hoshikawa, E.; Saito, T.; Suebsamarn, O.; Naito, E.; Suzuki, A.; Ishihara, S.; Haga, H.; Tomihara, K.; Izumi, K. The EGF/EGFR axis and its downstream signaling pathways regulate the motility and proliferation of cultured oral keratinocytes. FEBS Open Bio 2023, 13, 1469–1484. [Google Scholar] [CrossRef]
- Değim, Z.; Çelebi, N.; Alemdaroğlu, C.; Deveci, M.; Öztürk, S.; Özoğul, C. Evaluation of chitosan gel containing liposome-loaded epidermal growth factor on burn wound healing. Int. Wound J. 2011, 8, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Alemdaroğlu, C.; Degim, Z.; Celebi, N.; Sengezer, M.; Alömeroglu, M.; Nacar, A. Investigation of epidermal growth factor containing liposome formulation effects on burn wound healing. J. Biomed. Mater. Res. A 2008, 85, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Tallapaneni, V.; Mude, L.; Pamu, D.; Palanimuthu, V.R.; Magham, S.V.; Karri, V.V.S.R.; Parvathaneni, M. Growth factor loaded thermo-responsive injectable hydrogel for enhancing diabetic wound healing. Gels 2022, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lim, J.; Woo, K.C.; Kim, K.T. Piperonylic acid stimulates keratinocyte growth and survival by activating epidermal growth factor receptor (EGFR). Sci. Rep. 2018, 8, 162. [Google Scholar] [CrossRef] [PubMed]
- Edwin, F.; Wiepz, G.J.; Singh, R.; Peet, C.R.; Chaturvedi, D.; Bertics, P.J.; Patel, T.B. A historical perspective of the EGF receptor and related systems. Methods Mol. Biol. 2006, 327, 1–24. [Google Scholar] [CrossRef]
- Daub, H.; Weiss, F.U.; Wallasch, C.; Ullrich, A. Role of transactivation of the EGF receptor in signalling by G-protein-coupled receptors. Nature 1996, 379, 557–560. [Google Scholar] [CrossRef]
- Daub, H.; Wallasch, C.; Lankenau, A.; Herrlich, A.; Ullrich, A. Signal characteristics of G protein-transactivated EGF receptor. EMBO J. 1997, 16, 7032–7044. [Google Scholar] [CrossRef]
- Marikovsky, M.; Breuing, K.; Liu, P.Y.; Eriksson, E.; Higashiyama, S.; Farber, P.; Abraham, J.; Klagsbrun, M. Appearance of heparin-binding EGF-like growth factor in wound fluid as a response to injury. Proc. Natl. Acad. Sci. USA 1993, 90, 3889–3893. [Google Scholar] [CrossRef]
- Marikovsky, M.; Vogt, P.; Eriksson, E.; Rubin, J.S.; Taylor, W.G.; Joachim, S.; Klagsbrun, M. Wound fluid-derived heparin-binding EGF-like growth factor (HB-EGF) is synergistic with insulin-like growth factor-I for Balb/MK keratinocyte proliferation. J. Investig. Dermatol. 1996, 106, 616–621. [Google Scholar] [CrossRef]
- Hashimoto, K.; Higashiyama, S.; Asada, H.; Hashimura, E.; Kobayashi, T.; Sudo, K.; Nakagawa, T.; Damm, D.; Yoshikawa, K.; Taniguchi, N. Heparin-binding epidermal growth factor-like growth factor is an autocrine growth factor for human keratinocytes. J. Biol. Chem. 1994, 269, 20060–20066. [Google Scholar] [CrossRef]
- Shirakata, Y.; Kimura, R.; Nanba, D.; Iwamoto, R.; Tokumaru, S.; Morimoto, C.; Yokota, K.; Nakamura, M.; Sayama, K.; Mekada, E.; et al. Heparin-binding EGF-like growth factor accelerates keratinocyte migration and skin wound healing. J. Cell Sci. 2005, 118, 2363–2370. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, L.; Jiang, G.; Häkkinen, L.; Chan, B.; Larjava, H. HaCaT keratinocyte migration is dependent on epidermal growth factor receptor signaling and glycogen synthase kinase-3alpha. Exp. Cell. Res. 2006, 312, 2791–2805. [Google Scholar] [CrossRef]
- Thönes, S.; Rother, S.; Wippold, T.; Blaszkiewicz, J.; Balamurugan, K.; Moeller, S.; Ruiz-Gómez, G.; Schnabelrauch, M.; Scharnweber, D.; Saalbach, A.; et al. Hyaluronan/collagen hydrogels containing sulfated hyaluronan improve wound healing by sustained release of heparin-binding EGF-like growth factor. Acta Biomater. 2019, 86, 135–147. [Google Scholar] [CrossRef]
- Giltaire, S.; Lambert, S.; Poumay, Y. HB-EGF synthesis and release induced by cholesterol depletion of human epidermal keratinocytes is controlled by extracellular ATP and involves both p38 and ERK1/2 signaling pathways. J. Cell. Physiol. 2011, 226, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Shin, T.J.; Choi, S.H.; Cho, H.J.; Lee, B.H.; Pyo, M.K.; Lee, J.H.; Kang, J.; Kim, H.J.; Park, C.W.; et al. Gintonin, newly identified compounds from ginseng, is novel lysophosphatidic acids-protein complexes and activates G protein-coupled lysophosphatidic acid receptors with high affinity. Mol. Cells 2012, 33, 151–162. [Google Scholar] [CrossRef]
- Hwang, S.H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Won, K.J.; Lee, H.M.; Rhim, H.; Kim, H.C.; Nah, S.Y. Effects of gintonin on the proliferation, migration, and tube formation of human umbilical-vein endothelial cells: Involvement of lysophosphatidic-acid receptors and vascular-endothelial-growth-factor signaling. J. Ginseng Res. 2016, 40, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.E.; Park, S.D.; Hwang, H.; Choi, S.H.; Lee, R.M.; Nam, S.M.; Choi, J.H.; Rhim, H.; Cho, I.H.; Kim, H.C.; et al. Effects of a gintonin-enriched fraction on hair growth: An in vitro and in vivo study. J. Ginseng Res. 2020, 44, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Lee, N.E.; Hwang, H.; Rhim, H.; Cho, I.H.; Nah, S.Y. Ginseng gintonin enhances hyaluronic acid and collagen release from human dermal fibroblasts through lysophosphatidic acid receptor interaction. Molecules 2019, 24, 4438. [Google Scholar] [CrossRef]
- Choi, S.H.; Won, K.J.; Lee, R.; Cho, H.S.; Hwang, S.H.; Nah, S.Y. Wound healing effect of gintonin involves lysophosphatidic acid receptor/vascular endothelial growth factor signaling pathway in keratinocytes. Int. J. Mol. Sci. 2021, 22, 10155. [Google Scholar] [CrossRef]
- Kim, D.; Kim, H.J.; Baek, J.O.; Roh, J.Y.; Jun, H.S. Lysophosphatidic acid mediates imiquimod-induced psoriasis-like symptoms by promoting keratinocyte proliferation through LPAR1/ROCK2/PI3K/AKT signaling pathway. Int. J. Mol. Sci. 2021, 22, 10777. [Google Scholar] [CrossRef]
- Ockenga, W.; Kühne, S.; Bocksberger, S.; Banning, A.; Tikkanen, R. Epidermal growth factor receptor transactivation is required for mitogen-activated protein kinase activation by muscarinic acetylcholine receptors in HaCaT keratinocytes. Int. J. Mol. Sci. 2014, 15, 21433–21454. [Google Scholar] [CrossRef] [PubMed]
- Chernyavsky, A.I.; Arredondo, J.; Karlsson, E.; Wessler, I.; Grando, S.A. The Ras/Raf-1/MEK1/ERK signaling pathway coupled to integrin expression mediates cholinergic regulation of keratinocyte directional migration. J. Biol. Chem. 2005, 280, 39220–39228. [Google Scholar] [CrossRef] [PubMed]
- Grünwald, V.; Hidalgo, M. Developing inhibitors of the epidermal growth factor receptor for cancer treatment. J. Natl. Cancer Inst. 2003, 95, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Parmar, S.; Peng, X.X.; Shen, T.; Robey, R.W.; Bates, S.E.; Fu, L.W.; Shao, Y.; Chen, Y.M.; Zang, F.; et al. The epidermal growth factor tyrosine kinase inhibitor AG1478 and erlotinib reverse ABCG2-mediated drug resistance. Oncol. Rep. 2009, 21, 483–489. [Google Scholar]
- Kim, H.; Lee, B.H.; Choi, S.H.; Kim, H.J.; Jung, S.W.; Hwang, S.H.; Rhim, H.; Kim, H.C.; Cho, I.H.; Nah, S.Y. Gintonin stimulates gliotransmitter release in cortical primary astrocytes. Neurosci. Lett. 2015, 603, 19–24. [Google Scholar] [CrossRef]
- Nakai, K.; Yoneda, K.; Moriue, T.; Igarashi, J.; Kosaka, H.; Kubota, Y. HB-EGF-induced VEGF production and eNOS activation depend on both PI3 kinase and MAP kinase in HaCaT cells. J. Dermatol. Sci. 2009, 55, 170–178. [Google Scholar] [CrossRef]
- Choi, S.H.; Shin, T.J.; Lee, B.H.; Hwang, S.H.; Kang, J.; Kim, H.J.; Park, C.W.; Nah, S.Y. An edible gintonin preparation from ginseng. J. Ginseng Res. 2011, 35, 471–478. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, K.-J.; Lee, R.; Choi, S.-H.; Kim, J.-H.; Hwang, S.-H.; Nah, S.-Y. Gintonin-Induced Wound-Healing-Related Responses Involve Epidermal-Growth-Factor-like Effects in Keratinocytes. Int. J. Mol. Sci. 2023, 24, 14094. https://doi.org/10.3390/ijms241814094
Won K-J, Lee R, Choi S-H, Kim J-H, Hwang S-H, Nah S-Y. Gintonin-Induced Wound-Healing-Related Responses Involve Epidermal-Growth-Factor-like Effects in Keratinocytes. International Journal of Molecular Sciences. 2023; 24(18):14094. https://doi.org/10.3390/ijms241814094
Chicago/Turabian StyleWon, Kyung-Jong, Rami Lee, Sun-Hye Choi, Ji-Hun Kim, Sung-Hee Hwang, and Seung-Yeol Nah. 2023. "Gintonin-Induced Wound-Healing-Related Responses Involve Epidermal-Growth-Factor-like Effects in Keratinocytes" International Journal of Molecular Sciences 24, no. 18: 14094. https://doi.org/10.3390/ijms241814094
APA StyleWon, K.-J., Lee, R., Choi, S.-H., Kim, J.-H., Hwang, S.-H., & Nah, S.-Y. (2023). Gintonin-Induced Wound-Healing-Related Responses Involve Epidermal-Growth-Factor-like Effects in Keratinocytes. International Journal of Molecular Sciences, 24(18), 14094. https://doi.org/10.3390/ijms241814094