

Anti-Inflammatory Effects of Tegoprazan in Lipopolysaccharide-Stimulated Bone-Marrow-Derived Macrophages

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
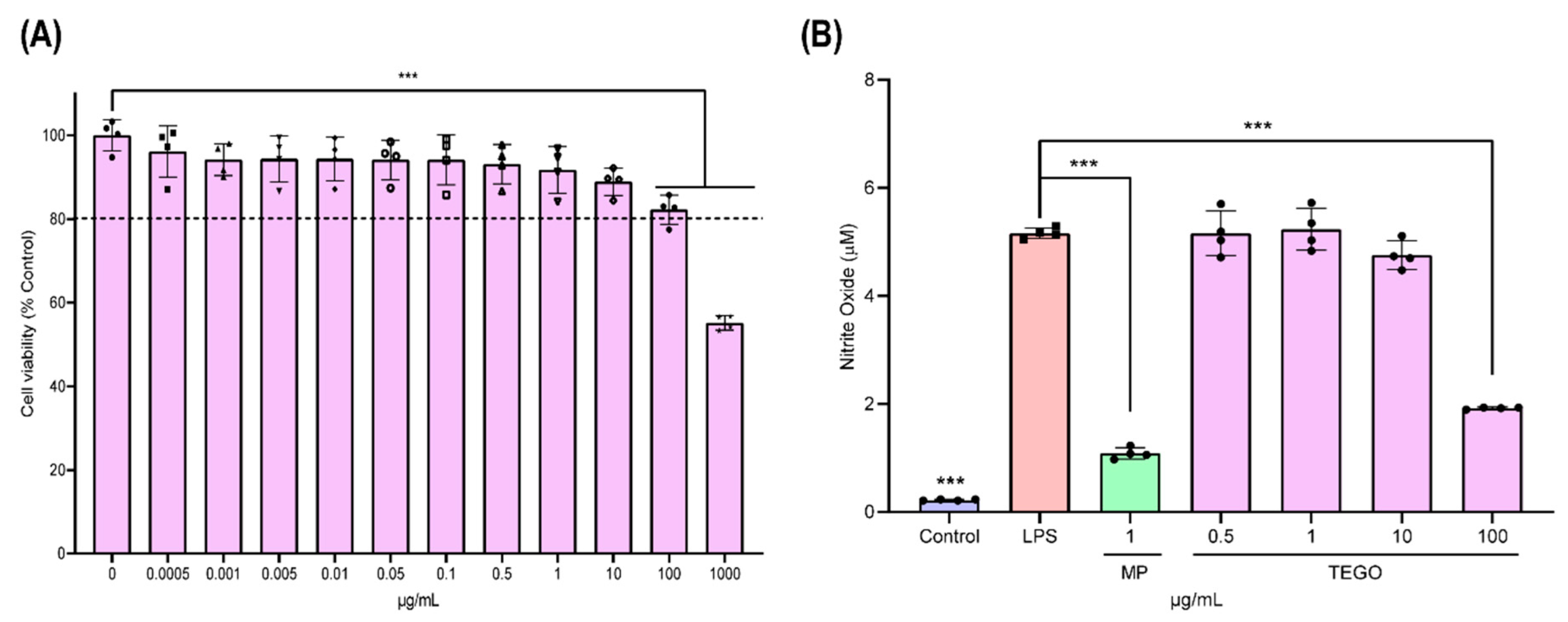
2.1. Effects of TEGO on Cytotoxicity and Nitric Oxide (NO) Production in BMMs
2.2. TEGO Treatment Increases Differentiation into M2-Phenotype Macrophages
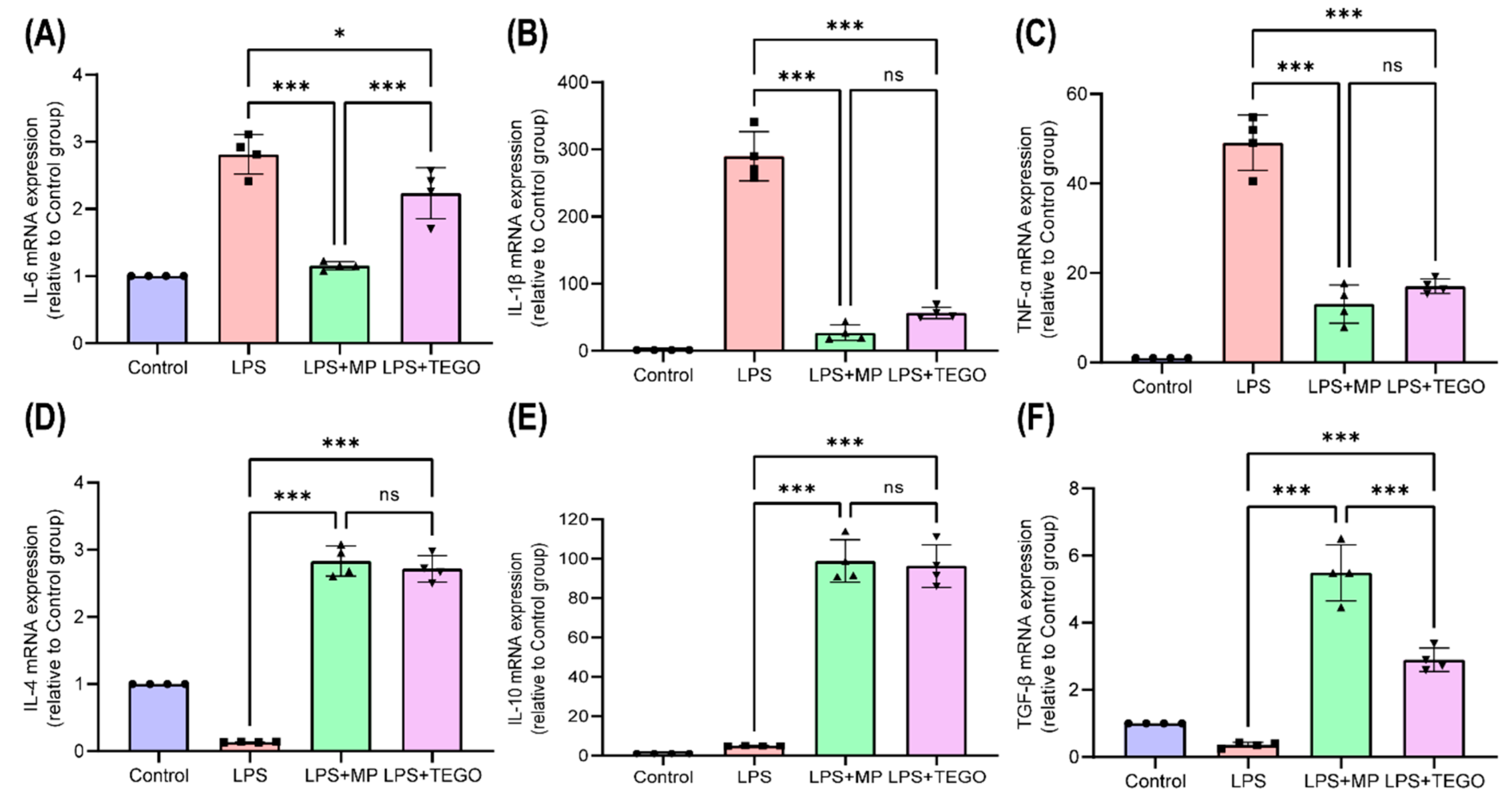
2.3. Effects of TEGO on mRNA Expression Levels of Pro-Inflammatory and Anti-Inflammatory Cytokine
2.4. Effects of TEGO on the Phosphorylation of Mitogen-Activated Protein Kinase (MAPK) Signals
3. Discussion
4. Materials and Methods
4.1. Preparation of TEGO and LPS
4.2. Isolation of BMMs and Differentiation into Macrophages
4.3. Cell Toxicity Test
4.4. NO Production
4.5. ICC Staining
4.6. FACS Analysis
4.7. qRT-PCR
4.8. Western Blotting
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Opal, S.M.; DePalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Bosca, L.; Zeini, M.; Traves, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.J.; Cobb, M.H. Mitogen-activated protein kinase pathways. Curr. Opin. Cell Biol. 1997, 9, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado Jde, D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-de-Arellano, M. Novel Markers to Delineate Murine M1 and M2 Macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Potter, V.J. Inflammation and macrophage modulation in adipose tissues. Cell. Microbiol. 2014, 16, 1484–1492. [Google Scholar] [CrossRef]
- Novak, M.L.; Koh, T.J. Macrophage phenotypes during tissue repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef]
- Ko, W.K.; Lee, S.H.; Kim, S.J.; Jo, M.J.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effects of ursodeoxycholic acid by lipopolysaccharide-stimulated inflammatory responses in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0180673. [Google Scholar] [CrossRef]
- Torres, J.; Enríquez-de-Salamanca, A.; Fernández, I.; Rodríguez-Ares, M.T.; Quadrado, M.J.; Murta, J.; del Castillo, J.M.B.; Stern, M.E.; Calonge, M. Activation of MAPK signaling pathway and NF-κB activation in pterygium and ipsilateral pterygium-free conjunctival specimens. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5842–5852. [Google Scholar] [CrossRef]
- Mussbacher, M.; Derler, M.; Basílio, J.; Schmid, J.A. NF-κB in monocytes and macrophages–an inflammatory master regulator in multitalented immune cells. Front. Immunol. 2023, 14, 1134661. [Google Scholar] [CrossRef]
- Grennan, D.; Wang, S. Steroid side effects. Jama 2019, 322, 282. [Google Scholar] [CrossRef] [PubMed]
- Levitte, S.; Yarani, R.; Ganguly, A.; Martin, L.; Gubatan, J.; Nadel, H.R.; Franc, B.; Gugig, R.; Syed, A.; Goyal, A.; et al. Case Series of Precision Delivery of Methylprednisolone in Pediatric Inflammatory Bowel Disease: Feasibility, Clinical Outcomes, and Identification of a Vasculitic Transcriptional Program. J. Clin. Med. 2023, 12, 2386. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Levi, A.D.; Vanni, S.; Shebert, R.T.; Sipski, M.L. High-dose methylprednisolone may cause myopathy in acute spinal cord injury patients. Spinal Cord. 2005, 43, 199–203. [Google Scholar] [CrossRef]
- Henry, D. Side-effects of non-steroidal anti-inflammatory drugs. Bailliere’s Clin. Rheumatol. 1988, 2, 425–454. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.; Carlsson, E. Potassium-competitive acid blockade: A new therapeutic strategy in acid-related diseases. Pharmacol. Ther. 2005, 108, 294–307. [Google Scholar] [CrossRef]
- Kim, D.K.; Lee, K.-H.; Kim, S.-j.; Kim, S.-J.; Lee, S.J.; Park, C.H.; Kim, B.-T.; Song, G.-S.; Moon, B.-S.; Ryu, S.-Y. Effects of tegoprazan, a novel potassium-competitive acid blocker, on rat models of gastric acid–related disease. J. Pharmacol. Exp. Ther. 2019, 369, 318–327. [Google Scholar] [CrossRef]
- Lee, K.J.; Son, B.K.; Kim, G.H.; Jung, H.K.; Jung, H.Y.; Chung, I.K.; Sung, I.K.; Kim, J.I.; Kim, J.H.; Lee, J.S. Randomised phase 3 trial: Tegoprazan, a novel potassium-competitive acid blocker, vs. esomeprazole in patients with erosive oesophagitis. Aliment. Pharmacol. Ther. 2019, 49, 864–872. [Google Scholar] [CrossRef]
- Han, S.; Choi, H.Y.; Kim, Y.H.; Nam, J.Y.; Kim, B.; Song, G.S.; Lim, H.S.; Bae, K.S. Randomised clinical trial: Safety, tolerability, pharmacokinetics, and pharmacodynamics of single and multiple oral doses of tegoprazan (CJ-12420), a novel potassium-competitive acid blocker, in healthy male subjects. Aliment. Pharmacol. Ther. 2019, 50, 751–759. [Google Scholar] [CrossRef]
- Takahashi, N.; Take, Y. Tegoprazan, a novel potassium-competitive acid blocker to control gastric acid secretion and motility. J. Pharmacol. Exp. Ther. 2018, 364, 275–286. [Google Scholar] [CrossRef]
- Cho, Y.K.; Kim, J.H.; Kim, H.S.; Kim, T.O.; Oh, J.H.; Choi, S.C.; Moon, J.S.; Lee, S.K.; Jung, S.W.; Kim, S.S. Randomised clinical trial: Comparison of tegoprazan and lansoprazole as maintenance therapy for healed mild erosive oesophagitis. Aliment. Pharmacol. Ther. 2023, 57, 72–80. [Google Scholar] [CrossRef]
- Qiu, M.; Campbell, T.; Breit, S. A potassium ion channel is involved in cytokine production by activated human macrophages. Clin. Exp. Immunol. 2002, 130, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Iwasawa, A.; Ayaki, M.; Niwano, Y. Cell viability score (CVS) as a good indicator of critical concentration of benzalkonium chloride for toxicity in cultured ocular surface cell lines. Regul. Toxicol. Pharmacol. 2013, 66, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, S.; Yang, F.; Guo, Y.; Chen, Z.; Yu, N.; Yao, L.; Huang, J.; Fan, W.; Xu, Z.; et al. The role of macrophage polarization and associated mechanisms in regulating the anti-inflammatory action of acupuncture: A literature review and perspectives. Chin. Med. 2021, 16, 56. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef]
- Groeneveld, P.H.; Kwappenberg, K.M.; Langermans, J.A.; Nibbering, P.H.; Curtis, L. Relation between pro-and anti-inflammatory cytokines and the production of nitric oxide (NO) in severe sepsis. Cytokine 1997, 9, 138–142. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef]
- Son, M.; Park, I.S.; Kim, S.; Ma, H.W.; Kim, J.H.; Kim, T.I.; Kim, W.H.; Han, J.; Kim, S.W.; Cheon, J.H. Novel Potassium-Competitive Acid Blocker, Tegoprazan, Protects Against Colitis by Improving Gut Barrier Function. Front. Immunol. 2022, 13, 870817. [Google Scholar] [CrossRef]
- Shibli, F.; Kitayama, Y.; Fass, R. Novel Therapies for Gastroesophageal Reflux Disease: Beyond Proton Pump Inhibitors. Curr. Gastroenterol. Rep. 2020, 22, 16. [Google Scholar] [CrossRef]
- Yeo, M.; Kim, D.K.; Han, S.U.; Lee, J.E.; Kim, Y.B.; Cho, Y.K.; Kim, J.H.; Cho, S.W.; Hahm, K.B. Novel action of gastric proton pump inhibitor on suppression of Helicobacter pylori induced angiogenesis. Gut 2006, 55, 26–33. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjornsson, B. The Role of Mitogen-Activated Protein Kinase-Activated Protein Kinases (MAPKAPKs) in Inflammation. Genes 2013, 4, 101–133. [Google Scholar] [CrossRef]
- Innes, J.F.; Clayton, J.; Lascelles, B.D. Review of the safety and efficacy of long-term NSAID use in the treatment of canine osteoarthritis. Vet. Rec. 2010, 166, 226–230. [Google Scholar] [CrossRef]
- Kim, S.J.; Ko, W.K.; Jo, M.J.; Arai, Y.; Choi, H.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effect of Tauroursodeoxycholic acid in RAW 264.7 macrophages, Bone marrow-derived macrophages, BV2 microglial cells, and spinal cord injury. Sci. Rep. 2018, 8, 3176. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Ko, W.K.; Kim, S.J.; Han, I.B.; Hong, J.B.; Sheen, S.H.; Sohn, S. Inhibitory Effects of Gold and Silver Nanoparticles on the Differentiation into Osteoclasts In Vitro. Pharmaceutics 2021, 13, 462. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.; Ko, W.K.; Kim, S.J.; Han, G.H.; Lee, D.; Sheen, S.H.; Sohn, S. Lobeglitazone Exerts Anti-Inflammatory Effect in Lipopolysaccharide-Induced Bone-Marrow Derived Macrophages. Biomedicines 2021, 9, 1432. [Google Scholar] [CrossRef] [PubMed]
- Han, G.H.; Kim, S.J.; Ko, W.K.; Lee, D.; Han, I.B.; Sheen, S.H.; Hong, J.B.; Sohn, S. Transplantation of tauroursodeoxycholic acid-inducing M2-phenotype macrophages promotes an anti-neuroinflammatory effect and functional recovery after spinal cord injury in rats. Cell. Prolif. 2021, 54, e13050. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Ko, W.K.; Heo, D.N.; Lee, S.J.; Lee, D.; Heo, M.; Han, I.B.; Kwon, I.; Sohn, S. Anti-neuroinflammatory gold nanocomplex loading ursodeoxycholic acid following spinal cord injury. Chem. Eng. J. 2019, 375, 122088. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, G.-H.; Kim, S.-J.; Ko, W.-K.; Hong, J.-B.; Sheen, S.-H.; Cho, M.-J.; Sohn, S. Anti-Inflammatory Effects of Tegoprazan in Lipopolysaccharide-Stimulated Bone-Marrow-Derived Macrophages. Int. J. Mol. Sci. 2023, 24, 14589. https://doi.org/10.3390/ijms241914589
Han G-H, Kim S-J, Ko W-K, Hong J-B, Sheen S-H, Cho M-J, Sohn S. Anti-Inflammatory Effects of Tegoprazan in Lipopolysaccharide-Stimulated Bone-Marrow-Derived Macrophages. International Journal of Molecular Sciences. 2023; 24(19):14589. https://doi.org/10.3390/ijms241914589
Chicago/Turabian StyleHan, Gong-Ho, Seong-Jun Kim, Wan-Kyu Ko, Je-Beom Hong, Seung-Hun Sheen, Min-Jai Cho, and Seil Sohn. 2023. "Anti-Inflammatory Effects of Tegoprazan in Lipopolysaccharide-Stimulated Bone-Marrow-Derived Macrophages" International Journal of Molecular Sciences 24, no. 19: 14589. https://doi.org/10.3390/ijms241914589