
Excessive Pregestational Weight and Maternal Obstetric Complications: The Role of Adipokines
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Search Strategies
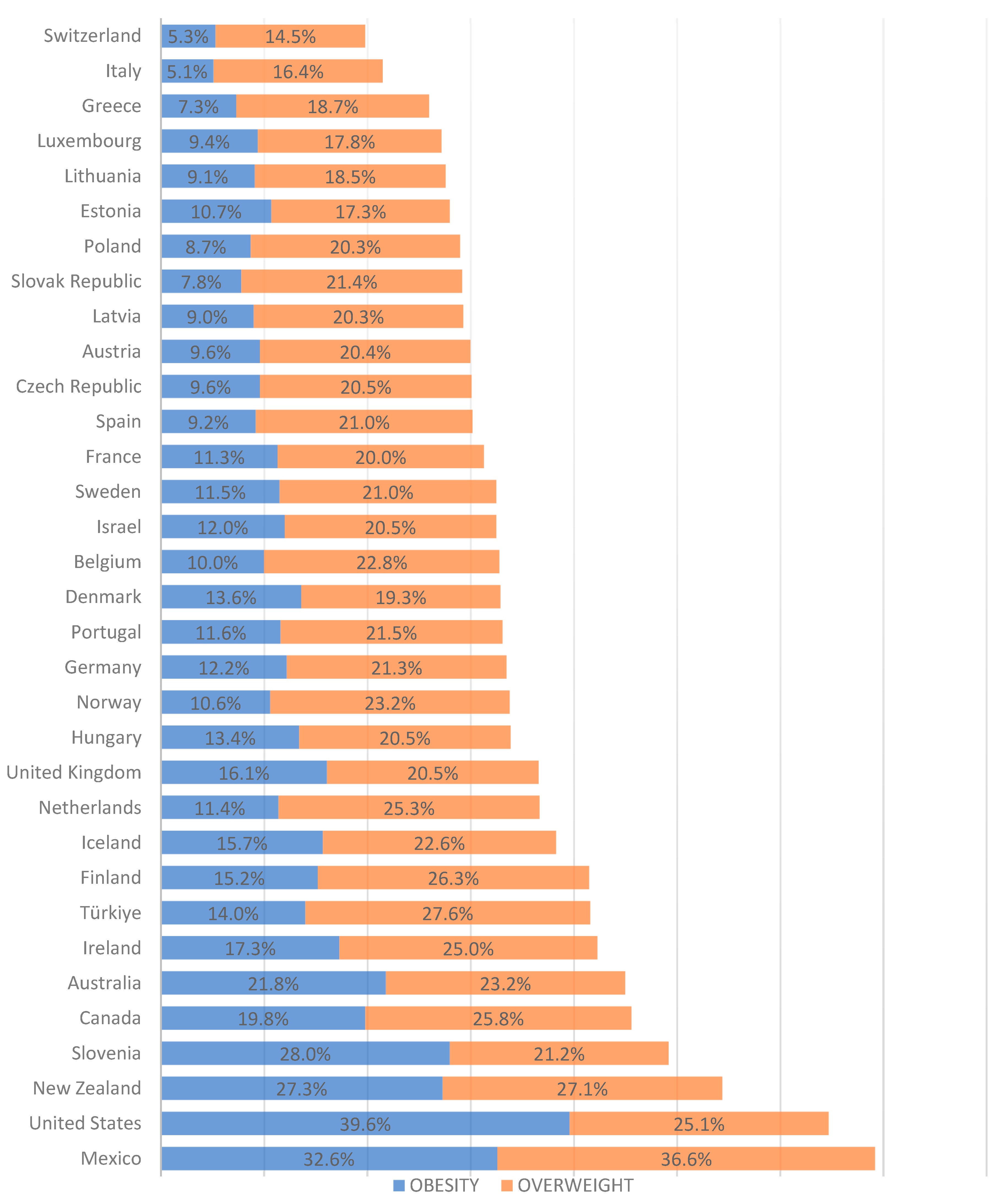
3. Risk of Maternal Obstetric Complications in Pregnant Women with Excess Weight
4. Adipose Tissue, Adipokines, and Excessive Weight in Pregnancy
5. Potential Pathophysiological Mechanisms of Leptin and Adiponectin in Maternal Obstetric Complications
5.1. Leptin
5.2. Adiponectin
6. Perspectives on the Study of Adipokines in Excess-Weight Pregnant Women
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- A Healthy Lifestyle—WHO Recommendations. Available online: https://www.euro.who.int/en/health-topics/disease-prevention/nutrition/a-healthy-lifestyle/body-mass-index-bmi (accessed on 15 May 2023).
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and obesity: Prevalence, consequences, and causes of a growing public health problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef]
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128.9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef]
- World Obesity. Global Obesity Observatory. Available online: https://data.worldobesity.org/ (accessed on 20 May 2023).
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Arima, Y.; Fukuoka, H. Developmental origins of health and disease theory in cardiology. J. Cardiol. 2020, 76, 14–17. [Google Scholar] [CrossRef]
- Lewandowska, M.; Więckowska, B.; Sajdak, S. Pre-Pregnancy Obesity, Excessive Gestational Weight Gain, and the Risk of Pregnancy-Induced Hypertension and Gestational Diabetes Mellitus. J. Clin. Med. 2020, 9, 1980. [Google Scholar] [CrossRef]
- Shen, M.; Smith, G.N.; Rodger, M.; White, R.R.; Walker, M.C.; Wen, S.W. Comparison of risk factors and outcomes of gestational hypertension and pre-eclampsia. PLoS ONE 2017, 12, e0175914. [Google Scholar] [CrossRef]
- Shin, D.; Song, W.O. Prepregnancy body mass index is an independent risk factor for gestational hypertension, gestational diabetes, preterm labor, and small- and large-for-gestational-age infants. J. Matern.-Fetal Neonatal Med. 2015, 28, 1679–1686. [Google Scholar] [CrossRef]
- He, X.J.; Dai, R.X.; Hu, C.L. Maternal prepregnancy overweight and obesity and the risk of preeclampsia: A meta-analysis of cohort studies. Obes. Res. Clin. Pract. 2020, 14, 27–33. [Google Scholar] [CrossRef]
- Yang, Y.; Le Ray, I.; Zhu, J.; Zhang, J.; Hua, J.; Reilly, M. Preeclampsia Prevalence, Risk Factors, and Pregnancy Outcomes in Sweden and China. JAMA Netw. Open 2021, 4, e218401. [Google Scholar] [CrossRef]
- Li, G.; Wei, T.; Ni, W.; Zhang, A.; Zhang, J.; Xing, Y.; Xing, Q. Incidence and Risk Factors of Gestational Diabetes Mellitus: A Prospective Cohort Study in Qingdao, China. Front. Endocrinol. 2020, 11, 636. [Google Scholar] [CrossRef]
- Alwash, S.M.; McIntyre, H.D.; Mamun, A. The association of general obesity, central obesity and visceral body fat with the risk of gestational diabetes mellitus: Evidence from a systematic review and meta-analysis. Obes. Res. Clin. Pract. 2021, 15, 425–430. [Google Scholar] [CrossRef]
- Butwick, A.J.; Abreo, A.; Bateman, B.T.; Lee, H.C.; El-Sayed, Y.Y.; Stephansson, O.; Flood, P. Effect of Maternal Body Mass Index on Postpartum Hemorrhage. Anesthesiology 2018, 128, 774–783. [Google Scholar] [CrossRef]
- Ende, H.B.; Lozada, M.J.D.; Chestnut, D.H.; Osmundson, S.S.; Walden, R.L.M.; Shotwell, M.S.; Bauchat, J.R. Risk Factors for Atonic Postpartum Hemorrhage: A Systematic Review and Meta-analysis. Obstet. Gynecol. 2021, 137, 305–323. [Google Scholar] [CrossRef]
- Li, S.; Gao, J.; Liu, J.; Hu, J.; Chen, X.; He, J.; Tang, Y.; Liu, X.; Cao, Y.; Liu, X.; et al. Incidence and Risk Factors of Postpartum Hemorrhage in China: A Multicenter Retrospective Study. Front. Med. 2021, 8, 673500. [Google Scholar] [CrossRef]
- Nkoka, O.; Ntenda, P.A.M.; Senghore, T.; Bass, P. Maternal overweight and obesity and the risk of caesarean birth in Malawi. Reprod. Health 2019, 16, 40. [Google Scholar] [CrossRef]
- Pettersen-Dahl, A.; Murzakanova, G.; Sandvik, L.; Laine, K. Maternal body mass index as a predictor for delivery method. Acta Obstet. Gynecol. Scand. 2018, 97, 212–218. [Google Scholar] [CrossRef]
- Kvalvik, S.A.; Rasmussen, S.; Thornhill, H.F.; Baghestan, E. Risk factors for surgical site infection following cesarean delivery: A hospital-based case-control study. Acta Obstet. Gynecol. Scand. 2021, 100, 2167–2175. [Google Scholar] [CrossRef]
- Leth, R.A.; Uldbjerg, N.; Nørgaard, M.; Møller, J.K.; Thomsen, R.W. Obesity, diabetes, and the risk of infections diagnosed in hospital and post-discharge infections after cesarean section: A prospective cohort study. Acta Obstet. Gynecol. Scand. 2011, 90, 501–509. [Google Scholar] [CrossRef]
- Trivett, C.; Lees, Z.J.; Freeman, D.J. Adipose tissue function in healthy pregnancy, gestational diabetes mellitus and pre-eclampsia. Eur. J. Clin. Nutr. 2021, 75, 1745–1756. [Google Scholar] [CrossRef]
- Cypess, A.M. Reassessing Human Adipose Tissue. N. Engl. J. Med. 2022, 386, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Selovic, A.; Sarac, J.; Missoni, S. Changes in adipose tissue distribution during pregnancy estimated by ultrasonography. J. Matern. Fetal Neonatal Med. 2016, 29, 2131–2137. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, H.M.; Huston-Presley, L.; Catalano, P.M. The influence of obesity and gestational diabetes mellitus on accretion and the distribution of adipose tissue in pregnancy. Am. J. Obstet. Gynecol. 2003, 189, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Neeland, I.J.; Ross, R.; Després, J.-P.; Matsuzawa, Y.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; et al. Visceral and ectopic fat, atherosclerosis, and cardiometabolic disease: A position statement. Lancet Diabetes Endocrinol. 2019, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Mazurek, D.; Bronkowska, M. Maternal Anthropometric Factors and Circulating Adipokines as Predictors of Birth Weight and Length. Int. J. Environ. Res. Public Health 2020, 17, 4799. [Google Scholar] [CrossRef]
- Solis-Paredes, M.; Espino y Sosa, S.; Estrada-Gutierrez, G.; Nava-Salazar, S.; Ortega-Castillo, V.; Rodriguez-Bosch, M.; Bravo-Flores, E.; Espejel-Nuñez, A.; Tolentino-Dolores, M.; Gaona-Estudillo, R.; et al. Maternal and Fetal Lipid and Adipokine Profiles and Their Association with Obesity. Int. J. Endocrinol. 2016, 2016, 7015626. [Google Scholar] [CrossRef]
- Franco-Sena, A.B.; de Oliveira, L.C.; Pinto, T.d.J.P.; Farias, D.R.; Vaz, J.d.S.; Kac, G. Factors associated with prospective leptin concentrations throughout pregnancy in pregestational normal weight, overweight and obese women. Clin. Endocrinol. 2015, 82, 127–135. [Google Scholar] [CrossRef]
- Haghiac, M.; Basu, S.; Presley, L.; Serre, D.; Catalano, P.M.; Hauguel-de Mouzon, S. Patterns of adiponectin expression in term pregnancy: Impact of obesity. J. Clin. Endocrinol. Metab. 2014, 99, 3427–3434. [Google Scholar] [CrossRef]
- Ozias, M.K.; Li, S.; Hull, H.R.; Brooks, W.M.; Carlson, S.E. Relationship of circulating adipokines to body composition in pregnant women. Adipocyte 2014, 4, 44–49. [Google Scholar] [CrossRef]
- Straughen, J.K.; Misra, D.P.; Kumar, P.; Misra, V.K. The influence of overweight and obesity on maternal soluble fms-like tyrosine kinase 1 and its relationship with leptin during pregnancy. Reprod. Sci. 2013, 20, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Savard, C.; Lemieux, S.; Plante, A.-S.; Gagnon, M.; Leblanc, N.; Veilleux, A.; Tchernof, A.; Morisset, A.-S. Longitudinal changes in circulating concentrations of inflammatory markers throughout pregnancy: Are there associations with diet and weight status? Appl. Physiol. Nutr. Metab. 2022, 47, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Andersson-Hall, U.; Svedin, P.; Svensson, H.; Lönn, M.; Mallard, C.; Holmäng, A. Longitudinal changes in adipokines and free leptin index during and after pregnancy in women with obesity. Int. J. Obes. 2020, 44, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, S.N.; Rawal, S.; Liu, D.; Chen, J.; Tsai, M.Y.; Zhang, C. Maternal adipokines longitudinally measured across pregnancy and their associations with neonatal size, length, and adiposity. Int. J. Obes. 2019, 43, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Vernini, J.M.; Moreli, J.B.; Costa, R.A.; Negrato, C.A.; Rudge, M.V.; Calderon, I.M. Maternal adipokines and insulin as biomarkers of pregnancies complicated by overweight and obesity. Diabetol. Metab. Syndr. 2016, 13, 8–68. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Thomas, A.; Reisetter, A.C.; Scholtens, D.M.; Wolever, T.M.; Josefson, J.; Layden, B.T. Maternal short-chain fatty acids are associated with metabolic parameters in mothers and newborns. Transl. Res. 2014, 164, 153–157. [Google Scholar] [CrossRef]
- Sen, S.; Iyer, C.; Klebenov, D.; Histed, A.; Aviles, J.A.; Meydani, S.N. Obesity impairs cell-mediated immunity during the second trimester of pregnancy. Am. J. Obstet. Gynecol. 2013, 208, 139.e1–139.e8. [Google Scholar] [CrossRef]
- Maguire, R.L.; House, J.S.; Lloyd, D.T.; Skinner, H.G.; Allen, T.K.; Raffi, A.M.; Skaar, D.A.; Park, S.S.; McCullough, L.E.; Kollins, S.H.; et al. Associations between maternal obesity, gestational cytokine levels and child obesity in the NEST cohort. Pediatr. Obes. 2021, 16, e12763. [Google Scholar] [CrossRef]
- Hanssens, S.; Marousez, L.; Pécheux, O.; Besengez, C.; Storme, L.; Deruelle, P.; Eberlé, D.; Lesage, J. Maternal obesity reduces apelin level in cord blood without altering the placental apelin/elabela-APJ system. Placenta 2022, 128, 112–115. [Google Scholar] [CrossRef]
- Pendeloski, K.P.T.; Ono, E.; Torloni, M.R.; Mattar, R.; Daher, S. Maternal obesity and inflammatory mediators: A controversial association. Am. J. Reprod. Immunol. 2017, 77, e12674. [Google Scholar] [CrossRef]
- Frühbeck, G. Intracellular signalling pathways activated by leptin. Biochem. J. 2006, 393, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, C.J.; Hoggard, N.; Thomas, L.; Mercer, J.G.; Wallace, J.M.; Lea, R.G. Placental leptin. Rev. Reprod. 2000, 5, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Kelesidis, I.; Chou, S.; Mantzoros, C.S. Narrative review: The role of leptin in human physiology: Emerging clinical applications. Ann. Intern. Med. 2010, 152, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Naylor, C.; Petri, W.A. Leptin Regulation of Immune Responses. Trends. Mol. Med. 2016, 22, 88–98. [Google Scholar] [CrossRef] [PubMed]
- González, R.R.; Simón, C.; Caballero-Campo, P.; Norman, R.; Chardonnens, D.; Devoto, L.; Bischof, P. Leptin and reproduction. Hum. Reprod. Update 2000, 6, 290–300. [Google Scholar] [CrossRef]
- van den Heuvel, J.K.; Eggels, L.; van Rozen, A.J.; Luijendijk, M.C.M.; Fliers, E.; Kalsbeek, A.; Adan, R.A.H.; la Fleur, S.E. Neuropeptide Y and leptin sensitivity is dependent on diet composition. J. Neuroendocrinol. 2014, 26, 377–385. [Google Scholar] [CrossRef]
- Harris, R.B.; Apolzan, J.W. Changes in glucose tolerance and leptin responsiveness of rats offered a choice of lard, sucrose, and chow. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1327–R1339. [Google Scholar] [CrossRef]
- Hâkansson, M.L.; Brown, H.; Ghilardi, N.; Skoda, R.C.; Meister, B. Leptin receptor immunoreactivity in chemically defined target neurons of the hypothalamus. J. Neurosci. 1998, 18, 559–572. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A Chronic Low-Grade Inflammation and Its Markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef]
- Wunderlich, C.M.; Hövelmeyer, N.; Wunderlich, F.T. Mechanisms of chronic JAK-STAT3-SOCS3 signaling in obesity. JAKSTAT 2013, 2, e23878. [Google Scholar] [CrossRef]
- Reed, A.S.; Unger, E.K.; Olofsson, L.E.; Piper, M.L.; Myers, M.G.; Xu, A.W. Functional role of suppressor of cytokine signaling 3 upregulation in hypothalamic leptin resistance and long-term energy homeostasis. Diabetes 2010, 59, 894–906. [Google Scholar] [CrossRef]
- Trujillo, M.L.; Spuch, C.; Carro, E.; Señarís, R. Hyperphagia and Central Mechanisms for Leptin Resistance during Pregnancy. Endocrinology 2011, 152, 1355–1365. [Google Scholar] [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and Obesity: Role and Clinical Implication. Front. Endocrinol. 2021, 18, 585887. [Google Scholar] [CrossRef]
- Rugină, C.; Mărginean, C.O.; Meliţ, L.E.; Huţanu, A.; Ghiga, D.V.; Modi, V.; Mărginean, C. Systemic inflammatory status—A bridge between gestational weight gain and neonatal outcomes (STROBE-compliant article). Medicine 2021, 100, e24511. [Google Scholar] [CrossRef]
- Li, K.; Yang, C.; Fan, J.; Li, X.; Gu, C.; Liu, H. Prepregnancy body mass index, gestational weight gain, and maternal prepartum inflammation in normal pregnancies: Findings from a Chinese cohort. BMC Pregnancy Childbirth 2022, 22, 531. [Google Scholar] [CrossRef]
- Xiao, W.-Q.; He, J.-R.; Shen, S.-Y.; Lu, J.-H.; Kuang, Y.-S.; Wei, X.-L.; Qiu, X. Maternal circulating leptin profile during pregnancy and gestational diabetes mellitus. Diabetes Res. Clin. Pract. 2020, 161, 108041. [Google Scholar] [CrossRef]
- Hoggard, N.; Haggarty, P.; Thomas, L.; Lea, R.G. Leptin expression in placental and fetal tissues: Does leptin have a functional role? Biochem. Soc. Trans. 2001, 29, 57–63. [Google Scholar] [CrossRef]
- Faulkner, J.L.; Wright, D.; Antonova, G.; Jaffe, I.Z.; Kennard, S.; Belin de Chantemèle, E.J. Midgestation Leptin Infusion Induces Characteristics of Clinical Preeclampsia in Mice, Which Is Ablated by Endothelial Mineralocorticoid Receptor Deletion. Hypertension 2022, 79, 1536–1547. [Google Scholar] [CrossRef]
- Faulkner, J.L.; Lluch, E.; Kennard, S.; Antonova, G.; Jaffe, I.Z.; Belin de Chantemèle, E.J. Selective deletion of endothelial mineralocorticoid receptor protects from vascular dysfunction in sodium-restricted female mice. Biol. Sex Differ. 2020, 11, 64. [Google Scholar] [CrossRef]
- Huby, A.C.; Otvos, L.; Belin de Chantemèle, E.J. Leptin Induces Hypertension and Endothelial Dysfunction via Aldosterone-Dependent Mechanisms in Obese Female Mice. Hypertension 2016, 67, 1020–1028. [Google Scholar] [CrossRef]
- Mise, H.; Sagawa, N.; Matsumoto, T.; Yura, S.; Nanno, H.; Itoh, H.; Mori, T.; Masuzaki, H.; Hosoda, K.; Ogawa, Y.; et al. Augmented placental production of leptin in preeclampsia: Possible involvement of placental hypoxia. J. Clin. Endocrinol. Metab. 1998, 83, 3225–3229. [Google Scholar] [CrossRef]
- Iwagaki, S.; Yokoyama, Y.; Tang, L.; Takahashi, Y.; Nakagawa, Y.; Tamaya, T. Augmentation of leptin and hypoxia-inducible factor 1alpha mRNAs in the pre-eclamptic placenta. Gynecol. Endocrinol. 2004, 18, 263–268. [Google Scholar] [CrossRef]
- Taylor, B.D.; Ness, R.B.; Olsen, J.; Hougaard, D.M.; Skogstrand, K.; Roberts, J.M.; Haggerty, C.L. Serum leptin measured in early pregnancy is higher in women with preeclampsia compared with normotensive pregnant women. Hypertension 2015, 65, 594–599. [Google Scholar] [CrossRef]
- Shahid, R.; Bari, M.F.; Hussain, M. Serum biomarkers for the prediction and diagnosis of preeclampsia: A meta-analysis. J. Taibah Univ. Med. Sci. 2021, 17, 14–27. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.H.; Chen, Y.P.; Yuan, X.L.; Wang, J.; Zhu, H.; Lu, C.M. Maternal Circulating Concentrations of Tumor Necrosis Factor-Alpha, Leptin, and Adiponectin in Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. Sci. World J. 2014, 2014, 926932. [Google Scholar] [CrossRef]
- Roca, M.D.; Ramos, P.; López, C.; Aguilar, M. Significance of Serum-Plasma Leptin Profile during Pregnancy in Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. J. Clin. Med. 2022, 11, 2433. [Google Scholar] [CrossRef]
- Hennige, A.M.; Stefan, N.; Kapp, K.; Lehmann, R.; Weigert, C.; Beck, A.; Moeschel, K.; Mushack, J.; Schleicher, E.; Häring, H.-U. Leptin down-regulates insulin action through phosphorylation of serine-318 in insulin receptor substrate 1. FASEB J. 2006, 20, 1206–1208. [Google Scholar] [CrossRef]
- Kieffer, T.J.; Habener, J.F. The adipoinsular axis: Effects of leptin on pancreatic beta-cells. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E1–E14. [Google Scholar] [CrossRef]
- Qiu, C.; Williams, M.A.; Vadachkoria, S.; Frederick, I.O.; Luthy, D.A. Increased maternal plasma leptin in early pregnancy and risk of gestational diabetes mellitus. Obstet. Gynecol. 2004, 103, 519–525. [Google Scholar] [CrossRef]
- Carvajal, J.A.; Oporto, J.I. The Myometrium in Pregnant Women with Obesity. Curr. Vasc. Pharmacol. 2021, 19, 193–200. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Rice, G.E. Leptin and adiponectin stimulate the release of proinflammatory cytokines and prostaglandins from human placenta and maternal adipose tissue via nuclear factor-kappaB, peroxisomal proliferator-activated receptor-gamma and extracellularly regulated kinase 1/2. Endocrinology 2005, 146, 3334–3342. [Google Scholar] [CrossRef]
- Carlson, N.S.; Hernandez, T.L.; Hurt, K.J. Parturition dysfunction in obesity: Time to target the pathobiology. Reprod. Biol. Endocrinol. 2015, 13, 135. [Google Scholar] [CrossRef]
- Moynihan, A.T.; Hehir, M.P.; Glavey, S.V.; Smith, T.J.; Morrison, J.J. Inhibitory effect of leptin on human uterine contractility in vitro. Am. J. Obstet. Gynecol. 2006, 195, 504–509. [Google Scholar] [CrossRef]
- Cowman, W.; Scroggins, S.M.; Hamilton, W.S.; Karras, A.E.; Bowdler, N.C.; Devor, E.J.; Santillan, M.K.; Santillan, D.A. Association between plasma leptin and cesarean section after induction of labor: A case control study. BMC Pregnancy Childbirth 2022, 22, 29. [Google Scholar] [CrossRef]
- Koutras, A.; Fasoulakis, Z.; Syllaios, A.; Garmpis, N.; Diakosavvas, M.; Pagkalos, A.; Ntounis, T.; Kontomanolis, E.N. Physiology and Pathology of Contractility of the Myometrium. Vivo 2021, 35, 1401–1408. [Google Scholar] [CrossRef]
- Olsen, M.A.; Higham-Kessler, J.; Yokoe, D.S.; Butler, A.M.; Vostok, J.; Stevenson, K.B.; Khan, Y.; Fraser, V.J. Developing a risk stratification model for surgical site infection after abdominal hysterectomy. Infect. Control Hosp. Epidemiol. 2009, 30, 1077–1083. [Google Scholar] [CrossRef]
- Waisbren, E.; Rosen, H.; Bader, A.M.; Lipsitz, S.R.; Rogers, S.O., Jr.; Eriksson, E. Percent body fat and prediction of surgical site infection. J. Am. Coll. Surg. 2010, 210, 381–389. [Google Scholar] [CrossRef]
- Di Leo, A.; Piffer, S.; Ricci, F.; Manzi, A.; Poggi, E.; Porretto, V.; Fambri, P.; Piccini, G.; Patrizia, T.; Fabbri, L.; et al. Surgical site infections in an Italian surgical ward: A prospective study. Surg. Infect. 2009, 10, 533–538. [Google Scholar] [CrossRef]
- Beldi, G.; Bisch-Knaden, S.; Banz, V.; Muhlemann, K.; Candinas, D. Impact of intraoperative behavior on surgical site infections. Am. J. Surg. 2009, 198, 157–162. [Google Scholar] [CrossRef]
- Pugliese, G.; Liccardi, A.; Graziadio, C.; Barrea, L.; Muscogiuri, G.; Colao, A. Obesity and infectious diseases: Pathophysiology and epidemiology of a double pandemic condition. Int. J. Obes. 2022, 46, 449–465. [Google Scholar] [CrossRef]
- Madhu, S.V.; Aslam, M.; Mishra, B.K.; Gupta, A.; Jhamb, R. Association of 25 (OH) Vitamin D and Leptin in Individuals with Insulin Resistance. Indian J. Endocrinol. Metab. 2022, 26, 435–438. [Google Scholar] [CrossRef]
- Khademi, Z.; Hamedi-Shahraki, S.; Amirkhizi, F. Vitamin D insufficiency is associated with inflammation and deregulation of adipokines in patients with metabolic syndrome. BMC Endocr. Disord. 2022, 22, 223. [Google Scholar] [CrossRef]
- Menendez, C.; Lage, M.; Peino, R.; Baldelli, R.; Concheiro, P.; Dieguez, C.; Casanueva, F. Retinoic acid and vitamin D3 powerfully inhibit in vitro leptin secretion by human adipose tissue. J. Endocrinol. 2001, 170, 425–432. [Google Scholar] [CrossRef]
- Chen, L.W.; Chien, C.H.; Kuo, S.F.; Yu, C.Y.; Lin, C.L.; Chien, R.N. Low vitamin D level was associated with metabolic syndrome and high leptin level in subjects with nonalcoholic fatty liver disease: A community-based study. BMC Gastroenterol. 2019, 19, 126. [Google Scholar] [CrossRef]
- Maetani, M.; Maskarinec, G.; Franke, A.A.; Cooney, R.V. Association of leptin, 25-hydroxyvitamin D, and parathyroid hormone in women. Nutr. Cancer 2009, 61, 225–231. [Google Scholar] [CrossRef]
- Barrea, L.; Frias-Toral, E.; Pugliese, G.; Garcia-Velasquez, E.; De Los Angeles Carignano, M.; Savastano, S.; Colao, A.; Muscogiuri, G. Vitamin D in obesity and obesity-related diseases: An overview. Minerva Endocrinol. 2021, 46, 177–192. [Google Scholar] [CrossRef]
- Trochoutsou, A.I.; Kloukina, V.; Samitas, K.; Xanthou, G. Vitamin-D in the Immune System: Genomic and Non-Genomic Actions. Mini Rev. Med. Chem. 2015, 15, 953–963. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, X.; Chen, D.; Li, Z. The Controversial Role of Adiponectin in Appetite Regulation of Animals. Nutrients 2021, 13, 3387. [Google Scholar] [CrossRef]
- Lee, H.; Tu, T.H.; Park, B.S.; Yang, S.; Kim, J.G. Adiponectin Reverses the Hypothalamic Microglial Inflammation during Short-Term Exposure to Fat-Rich Diet. Int. J. Mol. Sci. 2019, 20, 5738. [Google Scholar] [CrossRef]
- Mazaki-Tovi, S.; Kanety, H.; Pariente, C.; Hemi, R.; Wiser, A.; Schiff, E.; Sivan, E. Maternal serum adiponectin levels during human pregnancy. J. Perinatol. 2007, 27, 77–81. [Google Scholar] [CrossRef]
- D’Anna, R.; Baviera, G.; Corrado, F.; Giordano, D.; Di Benedetto, A.; Jasonni, V.M. Plasma Adiponectin Concentration in Early Pregnancy and Subsequent Risk of Hypertensive Disorders. Obstet. Gynecol. 2005, 106, 340–344. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I.; Sata, M.; Arita, Y.; Nishida, M.; Maeda, N.; Kumada, M.; Okamoto, Y.; Nagaretani, H.; Nishizawa, H.; et al. Role of adiponectin in preventing vascular stenosis. The missing link of adipo-vascular axis. J. Biol. Chem. 2002, 277, 37487–37491. [Google Scholar] [CrossRef]
- Ouchi, N.; Kobayashi, H.; Kihara, S.; Kumada, M.; Sato, K.; Inoue, T.; Funahashi, T.; Walsh, K. Adiponectin stimulates angiogenesis by promoting cross-talk between AMP-activated protein kinase and Akt signaling in endothelial cells. J. Biol. Chem. 2004, 279, 1304–1309. [Google Scholar] [CrossRef]
- Kobayashi, H.; Ouchi, N.; Kihara, S.; Walsh, K.; Kumada, M.; Abe, Y.; Funahashi, T.; Matsuzawa, Y. Selective suppression of endothelial cell apoptosis by the high molecular weight form of adiponectin. Circ. Res. 2004, 94, e27–e31. [Google Scholar] [CrossRef]
- Kobashi, C.; Urakaze, M.; Kishida, M.; Kibayashi, E.; Kobayashi, H.; Kihara, S.; Funahashi, T.; Takata, M.; Temaru, R.; Sato, A.; et al. Adiponectin inhibits endothelial synthesis of interleukin-8. Circ. Res. 2005, 97, 1245–1252. [Google Scholar] [CrossRef]
- Ouchi, N.; Kihara, S.; Arita, Y.; Okamoto, Y.; Maeda, K.; Kuriyama, H.; Hotta, K.; Nishida, M.; Takahashi, M.; Muraguchi, M.; et al. Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-kappaB signaling through a cAMP-dependent pathway. Circulation 2000, 102, 1296–1301. [Google Scholar] [CrossRef]
- Valencia-Ortega, J.; González-Reynoso, R.; Ramos-Martínez, E.G.; Ferreira-Hermosillo, A.; Peña-Cano, M.I.; Morales-Ávila, E.; Saucedo, R. New Insights into Adipokines in Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2022, 23, 6279. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.-S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.S.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2005–2010. [Google Scholar] [CrossRef] [PubMed]
- Ceddia, R.B.; Somwar, R.; Maida, A.; Fang, X.; Bikopoulos, G.; Sweeney, G. Globular adiponectin increases GLUT4 translocation and glucose uptake but reduces glycogen synthesis in rat skeletal muscle cells. Diabetologia 2005, 48, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Combs, T.P.; Berg, A.H.; Obici, S.; Scherer, P.E.; Rossetti, L. Endogenous glucose production is inhibited by the adipose-derived protein Acrp30. J. Clin. Investig. 2001, 108, 1875–1881. [Google Scholar] [CrossRef]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.; Guerra, D.D.; Bok, R.; Powell, T.; Jansson, T.; Hurt, K.J. Adiponectin links maternal metabolism to uterine contractility. FASEB J. 2019, 33, 14588–14601. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.; Ducza, E.; Bódi, N.; Bagyánszki, M.; Szalai, Z.; Mirdamadi, M.; Barna, T.; Szűcs, K.F.; Gáspár, R. The ontogenies of endometrial and myometrial leptin and adiponectin receptors in pregnant rats: Their putative impact on uterine contractility. Life Sci. 2022, 297, 120465. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Ghosh, A.; Al-Khairi, I.; Madiraju, S.R.M.; Abubaker, J.; Prentki, M. The multi-faces of Angptl8 in health and disease: Novel functions beyond lipoprotein lipase modulation. Prog. Lipid Res. 2020, 80, 101067. [Google Scholar] [CrossRef]
- Ruszała, M.; Niebrzydowska, M.; Pilszyk, A.; Kimber-Trojnar, Ż.; Trojnar, M.; Leszczyńska-Gorzelak, B. Novel Biomolecules in the Pathogenesis of Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 11578. [Google Scholar] [CrossRef]
- Luo, M.; Peng, D. ANGPTL8: An Important Regulator in Metabolic Disorders. Front. Endocrinol. 2018, 9, 169. [Google Scholar] [CrossRef]
- Kong, F.J.; Ma, L.L.; Li, G.; Chen, Y.X.; Zhou, J.Q. Circulating Betatrophin Levels and Gestational Diabetes Mellitus: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169941. [Google Scholar] [CrossRef]
- Akdemir, E.; Sahin, E.; Madendag, Y.; Sahin, M.E.; Madendag, I.C.; Acmaz, G.; Ozdemir, F.; Muhtaroglu, S.; Muderris, I.I. Effect of preeclampsia and preeclampsia severity on insulin, HOMA-IR, and betatrophin levels in non-diabetic pregnant women. Placenta 2020, 100, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Simsek, E.E.; Kasikci, H.O.; Sakin, O.; Korkut, S. Can Betatrophin predict the risk of preeclampsia? J. Clin. Diagn. Res. 2018, 12, QC12–QC15. [Google Scholar] [CrossRef]
- Adamis, A.P. Pathological Retinal Angiogenesis. In Encyclopedia of the Eye; Academic Press: Cambridge, MA, USA, 2010; pp. 280–289. [Google Scholar] [CrossRef]
- Famulla, S.; Lamers, D.; Hartwig, S.; Passlack, W.; Horrighs, A.; Cramer, A. Pigment epithelium-derived factor (PEDF) is one of the most abundant proteins secreted by human adipocytes and induces insulin resistance and inflammatory signaling in muscle and fat cells. Int. J. Obes. 2011, 35, 762–772. [Google Scholar] [CrossRef]
- He, X.; Cheng, R.; Benyajati, S.; Ma, J. PEDF and its roles in physiological and pathological conditions: Implication in diabetic and hypoxia-induced angiogenic diseases. Clin. Sci. 2015, 128, 805–823. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Yang, H.L.; Zhang, Y.P.; Ma, Q.L.; Long, Y.; Zheng, Z.X. Pigment epithelium-derived factor/vascular endothelial growth factor ratio for early prediction of preeclampsia: A prospective multicenter study in China. Pregnancy Hypertens. 2018, 14, 43–48. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, X.; Jiao, S.; Wang, Y. Upregulated pigment epithelium-derived factor (PEDF) promotes trophoblast apoptosis and inhibits invasion in preeclampsia. Reprod. Biol. 2021, 21, 100576. [Google Scholar] [CrossRef]
- Li, T.H.; Qiu, C.J.; Yu, X.J.; Liu, D.D.; Zhou, P.F.; Wu, L. Increased serum pigment epithelium-derived factor in women with gestational diabetes is associated with type 2 diabetes. Int. J. Endocrinol. 2015, 2015, 346938. [Google Scholar] [CrossRef]
- Gęca, T.; Kwiatek, M.; Krzyżanowski, A.; Kwaśniewska, A. C1q/TNF-Related Protein-3 (CTRP-3) and Pigment Epithelium-Derived Factor (PEDF) Concentrations in Patients with Gestational Diabetes Mellitus: A Case-Control Study. J. Clin. Med. 2020, 9, 2587. [Google Scholar] [CrossRef]
- Yin, X.; Huo, Y.; Liu, L.; Pan, Y.; Liu, S.; Wang, R. Serum Levels and Placental Expression of NGAL in Gestational Diabetes Mellitus. Int. J. Endocrinol. 2020, 2020, 8760563. [Google Scholar] [CrossRef]
- Jaberi, S.A.; Cohen, A.; D’Souza, C.; Abdulrazzaq, Y.M.; Ojha, S.; Bastaki, S. Lipocalin-2: Structure, function, distribution and role in metabolic disorders. Biomed. Pharmacother. 2021, 142, 112002. [Google Scholar] [CrossRef]
- Lu, L.; Li, C.; Deng, J.; Luo, J.; Huang, C. Maternal serum NGAL in the first trimester of pregnancy is a potential biomarker for the prediction of gestational diabetes mellitus. Front. Endocrinol. 2022, 13, 977254. [Google Scholar] [CrossRef] [PubMed]
- Sisti, G.; Rubin, G.; Zhou, C.; Orth, T.; Schiattarella, A. Neutrophil gelatinase-associated lipocalin as a predictor of pre-eclampsia: A systematic review and meta-analysis. Int. J. Gynaecol. Obstet. 2023, 163, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Khadir, A.; Kavalakatt, S.; Madhu, D.; Hammad, M.; Devarajan, S.; Tuomilehto, J.; Tiss, A. Fetuin-A levels are increased in the adipose tissue of diabetic obese humans but not in circulation. Lipids Health Dis. 2018, 17, 291. [Google Scholar] [CrossRef] [PubMed]
- Lee-Ødegård, S.; Ueland, T.; Thorsby, P.M.; Aukrust, P.; Michelsen, A.E.; Halvorsen, B.; Drevon, C.A.; Birkeland, K.I. Fetuin-A mediates the difference in adipose tissue insulin resistance between young adult pakistani and norwegian patients with type 2 diabetes. BMC Endocr. Disord. 2022, 22, 208. [Google Scholar] [CrossRef]
- Pal, D.; Dasgupta, S.; Kundu, R.; Maitra, S.; Das, G.; Mukhopadhyay, S.; Ray, S.; Majumdar, S.S.; Bhattacharya, S. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat. Med. 2012, 18, 1279–1285. [Google Scholar] [CrossRef]
- Kalabay, L.; Cseh, K.; Pajor, A.; Baranyi, E.; Csakany, G.; Melczer, Z.; Speer, G.; Kovacs, M.; Siller, G.; Karadi, I.; et al. Correlation of maternal serum fetuin/alpha2-HS-glycoprotein concentration with maternal insulin resistance and anthropometric parameters of neonates in normal pregnancy and gestational diabetes. Eur. J. Endocrinol. 2002, 147, 243–248. [Google Scholar] [CrossRef]
- Kralisch, S.; Hoffmann, A.; Lössner, U.; Kratzsch, J.; Blüher, M.; Stumvoll, M.; Fasshauer, M.; Ebert, T. Regulation of the novel adipokines/hepatokines fetuin A and fetuin B in gestational diabetes mellitus. Metabolism 2017, 68, 88–94. [Google Scholar] [CrossRef]
- Ren, Y.; Zhao, H.; Yin, C.; Lan, X.; Wu, L.; Du, X.; Griffiths, H.R.; Gao, D. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Front. Endocrinol. 2022, 13, 873699. [Google Scholar] [CrossRef]
- Rhinn, H.; Tatton, N.; McCaughey, S.; Kurnellas, M.; Rosenthal, A. Progranulin as a therapeutic target in neurodegenerative diseases. Trends Pharmacol. Sci. 2022, 43, 641–652. [Google Scholar] [CrossRef]
- Tang, W.; Lu, Y.; Tian, Q.-Y.; Zhang, Y.; Guo, F.-J.; Liu, G.-Y.; Syed, N.M.; Lai, Y.; Lin, E.A.; Kong, L.; et al. The Growth Factor Progranulin Binds to TNF Receptors and is Therapeutic Against Inflammatory Arthritis in Mice. Science 2011, 332, 478–484. [Google Scholar] [CrossRef]
- Matsubara, T.; Mita, A.; Minami, K.; Hosooka, T.; Kitazawa, S.; Takahashi, K.; Tamori, Y.; Yokoi, N.; Watanabe, M.; Matsuo, E.-I.; et al. PGRN is a Key Adipokine Mediating High Fat Diet-Induced Insulin Resistance and Obesity Through IL-6 in Adipose Tissue. Cell Metab. 2012, 15, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Osaka, M.; Takemori, S.; Watanabe, M.; Tanigaki, S.; Kobayashi, Y. Contraction-associated proteins expression by human uterine smooth muscle cells depends on maternal serum and progranulin associated with gestational weight gain. Endocr. J. 2020, 67, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Serdar Açıkgöz, A.; Tüten, A.; Öncül, M.; Eskalen, Ş.; Çakmak Dinçgez, B.; Şimşek, A.; Aytaç Yüksel, M.; Guralp, O. Evaluation of maternal serum progranulin levels in normotensive pregnancies, and pregnancies with early- and late-onset preeclampsia. J. Matern.-Fetal Neonatal Med. 2016, 29, 2658–2664. [Google Scholar] [CrossRef] [PubMed]
- Stubert, J.; Schattenberg, F.; Richter, D.U.; Dieterich, M.; Briese, V. Trophoblastic progranulin expression is upregulated in cases of fetal growth restriction and preeclampsia. J. Perinat. Med. 2012, 40, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Todoric, J.; Handisurya, A.; Perkmann, T.; Knapp, B.; Wagner, O.; Tura, A.; Pacini, G.; Esterbauer, H.; Kautzky-Willer, A. Circulating progranulin levels in women with gestational diabetes mellitus and healthy controls during and after pregnancy. Eur. J. Endocrinol. 2012, 167, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, Z.; Wang, C.; Li, K.; Li, S.; Boden, G.; Li, L.; Yang, G. Nesfatin-1 action in the brain increases insulin sensitivity through Akt/AMPK/TORC2 pathway in diet-induced insulin resistance. Diabetes 2012, 61, 1959–1968. [Google Scholar] [CrossRef]
- Wu, D.; Yang, M.; Chen, Y.; Jia, Y.; Ma, Z.A.; Boden, G.; Li, L.; Yang, G. Hypothalamic nesfatin-1/NUCB2 knockdown augments hepatic gluconeogenesis that is correlated with inhibition of mTOR-STAT3 signaling pathway in rats. Diabetes 2014, 63, 1234–1247. [Google Scholar] [CrossRef]
- Nakata, M.; Manaka, K.; Yamamoto, S.; Mori, M.; Yada, T. Nesfatin-1 enhances glucose-induced insulin secretion by promoting Ca2+ influx through L-type channels in mouse islet β-cells. Endocr. J. 2011, 58, 305–313. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, D.; Xu, J.; Chen, C.; Deng, D.; Pan, F.; Dong, L.; Li, S.; Ye, S. Circulating FABP4, nesfatin-1, and osteocalcin concentrations in women with gestational diabetes mellitus: A meta-analysis. Lipids Health Dis. 2020, 19, 199. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, Y.; Wang, Y.; Li, J.; Liu, R.; Liu, H. Decreased levels of serum nesfatin-1 in patients with preeclampsia. Biomarkers 2014, 19, 402–406. [Google Scholar] [CrossRef]
- Skheel, H.D.; Abdalkareem, Z.M.; Jasim, A.M. Correlation between serum nesfatin-1 level and severity of preeclampsia—A case control study. J. Pak. Med. Assoc. 2021, 71, S49–S52. [Google Scholar]
- Yang, R.-Z.; Lee, M.-J.; Hu, H.; Pray, J.; Wu, H.-B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.-W. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1253–E1261. [Google Scholar] [CrossRef] [PubMed]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem. Biophys. Res. Commun. 2011, 408, 339–343. [Google Scholar] [CrossRef]
- Kazama, K.; Usui, T.; Okada, M.; Hara, Y.; Yamawaki, H. Omentin plays an anti-inflammatory role through inhibition of TNF-α-induced superoxide production in vascular smooth muscle cells. Eur. J. Pharmacol. 2012, 686, 116–123. [Google Scholar] [CrossRef]
- Sun, J.; Ren, J.; Zuo, C.; Deng, D.; Pan, F.; Chen, R.; Zhu, J.; Chen, C.; Ye, S. Circulating apelin, chemerin and omentin levels in patients with gestational diabetes mellitus: A systematic review and meta-analysis. Lipids Health Dis. 2020, 19, 26. [Google Scholar] [CrossRef]
- Liu, H.; Wu, J.; Wang, H.; Sheng, L.; Tang, N.; Li, Y.; Hao, T. Association of serum omentin-1 concentrations with the presence and severity of preeclampsia. Ann. Clin. Biochem. 2015, 52, 245–250. [Google Scholar] [CrossRef]
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, H.; Ju, H.; Sun, M. Circulating chemerin levels and gestational diabetes mellitus: A systematic review and meta-analysis. Lipids Health Dis. 2018, 17, 169. [Google Scholar] [CrossRef]
- Bartho, L.A.; Kandel, M.; Walker, S.P.; Cluver, C.A.; Hastie, R.; Bergman, L. Circulating Chemerin Is Elevated in Women with Preeclampsia. Endocrinology 2023, 164, bqad041. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, L.; Liu, H.; Li, Z.; Li, L.; Wu, X.; Lei, Q.; Yin, A.; Tong, J.; Liu, K.; et al. Third-Trimester Maternal Serum Chemerin and Hypertension After Preeclampsia: A Prospective Cohort Study. J. Am. Heart. Assoc. 2023, 12, e027930. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Groups Compared | Gestational Age at Measurement | Concentrations in Women with Excessive Weight | Comments |
|---|---|---|---|---|
| Leptin | ||||
| Mazurek and Bronkowska, 2020 [28] | Overweight (n = 36) vs. normal weight women (n = 100) | After 39 weeks | ↑ | Leptin concentrations were directly proportional to the pregestational BMI and gestational weight gain. |
| Solis-Paredes et al., 2016 [29] | Overweight (n = 25) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| Franco-Sena et al., 2015 [30] | Overweight (n = 64) vs. normal weight women (n = 135) | 5–13 weeks 20–26 weeks 30–36 weeks | ↑ ↔ ↔ | In overweight or obese women, leptin concentrations tended to be higher than in women of normal weight, but a significant difference was only observed in the first trimester of pregnancy. |
| Haghiac et al., 2013 [31] | Overweight (n = 37) vs. normal weight women (n = 32) | 38.5–40 weeks | ↑ | Leptin concentrations increased according to the degree of excess body weight. |
| Ozias et al., 2015 [32] | Overweight/Obese (n = 21) vs. normal weight women (n = 17) | 35–39 weeks | ↑ | Leptin concentration correlated positively with BMI and total body fat mass in all women. |
| Straughen et al., 2013 [33] | Overweight/Obese (n = 143) vs. normal weight women (n = 143) | 6–10 weeks 10–14 weeks 16–20 weeks 22–26 weeks 32–36 weeks | ↑ ↑ ↑ ↑ ↑ | ~ |
| Savard et al., 2022 [34] | Obese (n = 15) vs. non obese women (BMI < 25 kg/m2, n = 45) | 9.9–14.7 weeks 20.7–25.3 weeks 31.6–34.6 weeks | ↑ | Only one arrow is shown because mixed models for repeated measurements were used to analyze leptin concentrations considering gestational trimester and BMI category. Pregestational BMI correlated positively with leptin concentrations. |
| Mazurek and Bronkowska, 2020 [28] | Obese (n = 14) vs. normal weight women (n = 100) | After 39 weeks | ↑ | Leptin concentrations were directly proportional to the pregestational BMI and gestational weight gain. |
| Andersson-Hall et al., 2019 [35] | Obese (n = 19) vs. normal weight women (n = 30) | 8–12 weeks 24–26 weeks 35–37 weeks | ↑ ↑ ↑ | The concentration of the soluble leptin receptor was also measured, and the free leptin index was calculated, which was significantly higher in obese women.In all women, changes in the free leptin index correlated positively with changes in fat mass across the pregnancy. |
| Hinkle et al., 2019 [36] | Obese (n = 66) vs. non obese women (BMI < 30 kg/m2, n = 255) | 10–14 weeks 15–26 weeks 23–31 weeks 33–39 weeks | ↑ ↑ ↑ ↑ | The concentration of the soluble leptin receptor was also measured, and the free leptin index was calculated, which was significantly higher in obese women. |
| Solis-Paredes et al., 2016 [29] | Obese (n = 22) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| Vernini et al., 2016 [37] | Obese (n = 31) vs. normal weight women (n = 23) | Weeks 37–38 | ↑ | ~ |
| Franco-Sena et al., 2015 [30] | Obese (n = 33) vs. normal weight women (n = 135) | 5–13 weeks 20–26 weeks 30–36 weeks | ↑ ↔ ↔ | In overweight or obese women, leptin concentrations tended to be higher than in women of normal weight, but a significant difference was only observed in the first trimester of pregnancy. |
| Priyadarshini et al., 2014 [38] | Obese (n = 10) vs. normal weight women (n = 10) | 36–38 weeks | ↑ | ~ |
| Haghiac et al., 2013 [31] | Women with BMI 30–40 (n = 36) vs. normal weight women (n = 32) | 38.5–40 weeks | ↑ | Leptin concentrations increased according to the degree of excess body weight. |
| Haghiac et al., 2013 [31] | Women with BMI > 40 (n = 28) vs. normal weight women (n = 32) | 38.5–40 weeks | ↑ | See the previous comment for this same reference. |
| Sen et al., 2013 [39] | Obese (n = 15) vs. normal weight women (n = 15) | 24–28 weeks | ↑ | ~ |
| Adiponectin | ||||
| Savard et al., 2022 [34] | Overweight (n = 19) vs. non obese women (BMI < 25 kg/m2, n = 45) | 9.9–14.7 weeks 20.7–25.3 weeks 31.6–34.6 weeks | ↓ | Only one arrow is shown because mixed models for repeated measurements were used to analyze adiponectin concentrations considering gestational trimester and BMI category. Pregestational BMI correlated negatively with adiponectin concentrations in the first and second trimesters. |
| Solis-Paredes et al., 2016 [29] | Overweight (n = 25) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| Savard et al., 2022 [34] | Obese (n = 15) vs. non obese women (BMI < 25 kg/m2, n = 45) | 9.9–14.7 weeks 20.7–25.3 weeks 31.6–34.6 weeks | ↓ | See the previous comment for this same reference. |
| Andersson-Hall et al., 2019 [35] | Obese (n = 19) vs. normal weight women (n = 30) | 8–12 weeks 24–26 weeks 35–37 weeks | ↓ ↓ ↓ | ~ |
| Hinkle et al., 2019 [36] | Obese (n = 66) vs. non obese women (BMI < 30 kg/m2, n = 255) | 10–14 weeks 15–26 weeks 23–31 weeks 33–39 weeks | ↓ ↓ ↓ ↓ | Arrows indicate total and high molecular weight adiponectin concentrations, although the difference was greater in the latter at weeks 23–31 and 33–39. |
| Priyadarshini et al., 2014 [38] | Obese (n = 10) vs. normal weight women (n = 10) | 36–38 weeks | ↔ | ~ |
| Vernini et al., 2016 [37] | Obese (n = 31) vs. normal weight women (n = 23) | 37–38 weeks | ↔ | ~ |
| Solis-Paredes et al., 2016 [29] | Obese (n = 22) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| Ozias et al., 2015 [32] | Overweight/Obese (n = 21) vs. normal weight women (n = 17) | 35–39 weeks | ↔ | Total and high molecular weight adiponectin concentrations were measured, and none were different between groups. |
| Haghiac et al., 2013 [31] | Women with BMI > 40 (n = 28) vs. normal weight women (n = 32) | 38.5–40 weeks | ↓ | Adiponectin concentrations of women with BMI 25–30 and 30–40 were also compared with women with normal weight, but no significant difference was observed. |
| Sen et al., 2013 [39] | Obese (n = 15) vs. normal weight women (n = 15) | 24–28 weeks | ↓ | ~ |
| Resistin | ||||
| Solis-Paredes et al., 2016 [29] | Overweight (n = 25) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| Vernini et al., 2016 [37] | Obese (n = 31) vs. normal weight women (n = 23) | 37–38 weeks | ↔ | ~ |
| Ozias et al., 2015 [32] | Overweight/Obese (n = 21) vs. normal weight women (n = 17) | 35–39 weeks | ↔ | In all women, resistin concentration correlated positively with abdominal visceral fat mass relative to total body fat mass. |
| Solis-Paredes et al., 2016 [29] | Obese (n = 22) vs. normal weight women (n = 20) | 37–40 weeks | ↔ | ~ |
| TNF-α | ||||
| Maguire et al., 2021 [40] | Obese (n = 124) vs. non obese women (BMI < 30 kg/m2, n = 237) | Median = 11.6 weeks (IQR = 9.3–18.7) | ↑ | Maternal concentrations of IL-8, IL-1β, IL-4, IFN-γ, IL-12 p70 subunit, and IL-17A were also analyzed, but no significant differences were observed. |
| IL-6 | ||||
| Savard et al., 2022 [34] | Overweight (n = 19) and obese (n = 15) vs. non obese women (BMI < 25 kg/m2, n = 45) | 9.9–14.7 weeks 20.7–25.3 weeks 31.6–34.6 weeks | ↔ | Only one arrow is shown because mixed models for repeated measurements were used to analyze IL-6 concentrations considering gestational trimester and BMI category. Pregestational BMI correlated positively with IL-6 concentrations in the first and second trimesters. |
| Maguire et al., 2021 [40] | Obese (n = 124) vs. non obese women (BMI < 30 kg/m2, n = 237) | Median = 11.6 weeks (IQR = 9.3–18.7) | ↔ | ~ |
| Visfatin | ||||
| Ozias et al., 2015 [32] | Overweight/Obese (n = 21) vs. normal weight women (n = 17) | 35–39 weeks | ↔ | ~ |
| Apelin | ||||
| Hanssens et al., 2022 [41] | Obese (n = 30) vs. normal weight women (n = 36) | 35–40 weeks | ↔ | ~ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valencia-Ortega, J.; Solis-Paredes, J.M.; Saucedo, R.; Estrada-Gutierrez, G.; Camacho-Arroyo, I. Excessive Pregestational Weight and Maternal Obstetric Complications: The Role of Adipokines. Int. J. Mol. Sci. 2023, 24, 14678. https://doi.org/10.3390/ijms241914678
Valencia-Ortega J, Solis-Paredes JM, Saucedo R, Estrada-Gutierrez G, Camacho-Arroyo I. Excessive Pregestational Weight and Maternal Obstetric Complications: The Role of Adipokines. International Journal of Molecular Sciences. 2023; 24(19):14678. https://doi.org/10.3390/ijms241914678
Chicago/Turabian StyleValencia-Ortega, Jorge, Juan Mario Solis-Paredes, Renata Saucedo, Guadalupe Estrada-Gutierrez, and Ignacio Camacho-Arroyo. 2023. "Excessive Pregestational Weight and Maternal Obstetric Complications: The Role of Adipokines" International Journal of Molecular Sciences 24, no. 19: 14678. https://doi.org/10.3390/ijms241914678