

The Immune Regulatory Role of Adenosine in the Tumor Microenvironment

Abstract
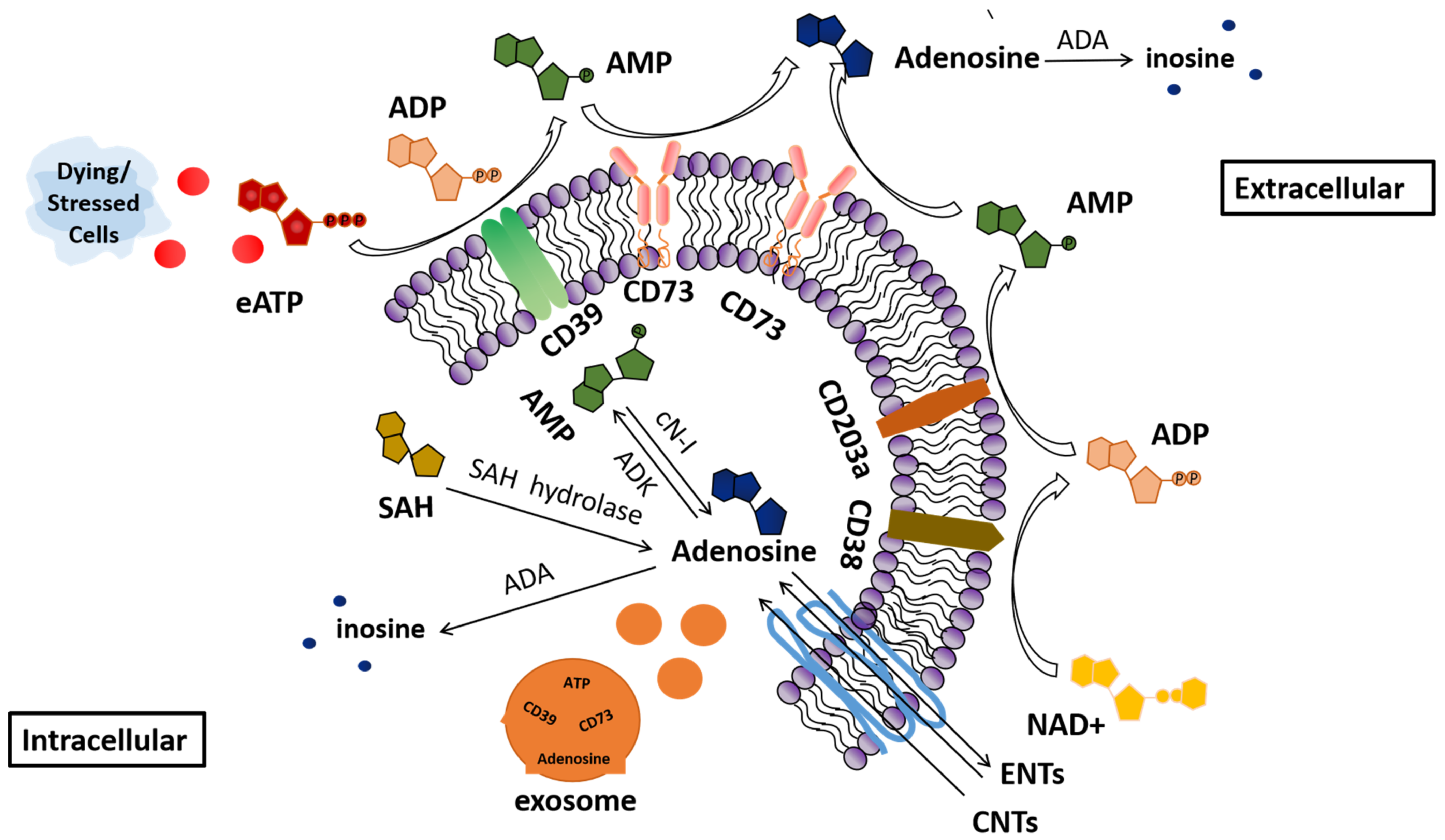
:1. Introduction
2. The Role of Adenosine on the Regulation of Various Immune Cells in the TME
2.1. Natural Killer Cells
2.2. Dendritic Cells
2.3. Effector CD4+ T Helper Cells
2.4. Cytotoxic CD8+ T Cells
2.5. Tumor-Associated Macrophages
2.6. Myeloid-Derived Suppressor Cells
2.7. CD4+ Regulatory T Cells
3. Adenosine/Adenosine Receptor Blockade
3.1. Targeting CD39 or CD73
3.2. Targeting A2AR or A2BR
3.3. Targeting CD38
3.4. Combination Therapy
4. Limitation and Prospect
Author Contributions
Funding
Conflicts of Interest
References
- Li, X.-Y.; Moesta, A.K.; Xiao, C.; Nakamura, K.; Casey, M.; Zhang, H.; Madore, J.; Lepletier, A.; Aguilera, A.R.; Sundarrajan, A.; et al. Targeting CD39 in Cancer Reveals an Extracellular ATP- and Inflammasome-Driven Tumor Immunity. Cancer Discov. 2019, 9, 1754–1773. [Google Scholar] [CrossRef] [PubMed]
- Bastid, J.; Cottalorda-Regairaz, A.; Alberici, G.; Bonnefoy, N.; Eliaou, J.F.; Bensussan, A. Entpd1/Cd39 Is a Promising Therapeutic Target in Oncology. Oncogene 2013, 32, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Chini, E.N.; Chini, C.C.; Netto, J.M.E.; de Oliveira, G.C.; van Schooten, W. The Pharmacology of Cd38/Nadase: An Emerging Target in Cancer and Diseases of Aging. Trends Pharmacol. Sci. 2018, 39, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Horenstein, A.L.; Chillemi, A.; Quarona, V.; Zito, A.; Roato, I.; Morandi, F.; Marimpietri, D.; Bolzoni, M.; Toscani, D.; Oldham, R.J.; et al. NAD+-Metabolizing Ectoenzymes in Remodeling Tumor–Host Interactions: The Human Myeloma Model. Cells 2015, 4, 520–537. [Google Scholar] [CrossRef] [PubMed]
- Vaisitti, T.; Audrito, V.; Serra, S.; Bologna, C.; Brusa, D.; Malavasi, F.; Deaglio, S. NAD+-metabolizing ecto-enzymes shape tumor-host interactions: The chronic lymphocytic leukemia model. FEBS Lett. 2011, 585, 1514–1520. [Google Scholar] [CrossRef]
- Azambuja, J.H.; Ludwig, N.; Braganhol, E.; Whiteside, T.L. Inhibition of the Adenosinergic Pathway in Cancer Rejuvenates Innate and Adaptive Immunity. Int. J. Mol. Sci. 2019, 20, 5698. [Google Scholar] [CrossRef]
- Kotulova, J.; Hajduch, M.; Dzubak, P. Current Adenosinergic Therapies: What Do Cancer Cells Stand to Gain and Lose? Int. J. Mol. Sci. 2021, 22, 12569. [Google Scholar] [CrossRef]
- Zhulai, G.; Oleinik, E.; Shibaev, M.; Ignatev, K. Adenosine-Metabolizing Enzymes, Adenosine Kinase and Adenosine Deaminase, in Cancer. Biomolecules 2022, 12, 418. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Gupta, R.S. Adenosine kinase and ribokinase—The RK family of proteins. Cell Mol. Life Sci. 2008, 65, 2875–2896. [Google Scholar] [CrossRef]
- Dutta, N.; Deb, I.; Sarzynska, J.; Lahiri, A. Inosine and Its Methyl Derivatives: Occurrence, Biogenesis, and Function in Rna. Prog. Biophys. Mol. Biol. 2022, 169–170, 21–52. [Google Scholar] [CrossRef]
- Gatsiou, A.; Vlachogiannis, N.; Lunella, F.F.; Sachse, M.; Stellos, K. Adenosine-to-Inosine RNA Editing in Health and Disease. Antioxid. Redox Signal. 2018, 29, 846–863. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Yin, S.; To, K.K.W.; Fu, L. Cd39/Cd73/A2ar Pathway and Cancer Immunotherapy. Mol. Cancer 2023, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Rubenich, D.S.; Zaręba, Ł.; Siewiera, J.; Pieper, J.; Braganhol, E.; Reichert, T.E.; Szczepański, M.J. Potential Roles of Tumor Cell- and Stroma Cell-Derived Small Extracellular Vesicles in Promoting a Pro-Angiogenic Tumor Microenvironment. Cancers 2020, 12, 3599. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Gillespie, D.G.; Menshikova, E.V.; Jackson, E.K.; Whiteside, T.L. Tumor-derived exosomes promote angiogenesis via adenosine A2B receptor signaling. Angiogenesis 2020, 23, 599–610. [Google Scholar] [CrossRef]
- Clayton, A.; Al-Taei, S.; Webber, J.; Mason, M.D.; Tabi, Z. Cancer Exosomes Express CD39 and CD73, Which Suppress T Cells through Adenosine Production. J. Immunol. 2011, 187, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G.; Boison, D. ATP and Adenosine Metabolism in Cancer: Exploitation for Therapeutic Gain. Pharmacol. Rev. 2022, 74, 799–824. [Google Scholar] [CrossRef] [PubMed]
- Allard, B.; Allard, D.; Buisseret, L.; Stagg, J. The Adenosine Pathway in Immuno-Oncology. Nat. Rev. Clin. Oncol. 2020, 17, 611–629. [Google Scholar] [CrossRef] [PubMed]
- Beavis, P.A.; Stagg, J.; Darcy, P.K.; Smyth, M.J. CD73: A potent suppressor of antitumor immune responses. Trends Immunol. 2012, 33, 231–237. [Google Scholar] [CrossRef]
- Moesta, A.K.; Li, X.-Y.; Smyth, M.J. Targeting CD39 in cancer. Nat. Rev. Immunol. 2020, 20, 739–755. [Google Scholar] [CrossRef]
- De Marchi, E.; Orioli, E.; Pegoraro, A.; Sangaletti, S.; Portararo, P.; Curti, A.; Colombo, M.P.; Di Virgilio, F.; Adinolfi, E. The P2X7 receptor modulates immune cells infiltration, ectonucleotidases expression and extracellular ATP levels in the tumor microenvironment. Oncogene 2019, 38, 3636–3650. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Sarti, A.C.; Falzoni, S.; De Marchi, E.; Adinolfi, E. Extracellular ATP and P2 purinergic signalling in the tumour microenvironment. Nat. Rev. Cancer 2018, 18, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Blay, J.; White, T.D.; Hoskin, D.W. The Extracellular Fluid of Solid Carcinomas Contains Immunosuppressive Concentrations of Adenosine. Cancer Res. 1997, 57, 2602–2605. [Google Scholar] [PubMed]
- Allard, B.; Longhi, M.S.; Robson, S.C.; Stagg, J. The ectonucleotidases CD39 and CD73: Novel checkpoint inhibitor targets. Immunol. Rev. 2017, 276, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Allard, B.; Beavis, P.A.; Darcy, P.K.; Stagg, J. Immunosuppressive activities of adenosine in cancer. Curr. Opin. Pharmacol. 2016, 29, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Samami, E.; Aleebrahim-Dehkordi, E.; Mohebalizadeh, M.; Yaribash, S.; Saghazadeh, A.; Rezaei, N. Inosine, Gut Microbiota, and Cancer Immunometabolism. Am. J. Physiol. Endocrinol. Metab. 2023, 324, E1–E8. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, F.; Zhang, Y.; Liu, J.; Zhao, B. ADAR1-Mediated RNA Editing and Its Role in Cancer. Front. Cell Dev. Biol. 2022, 10, 956649. [Google Scholar] [CrossRef]
- Dehnavi, S.; Kiani, A.; Sadeghi, M.; Biregani, A.F.; Banach, M.; Atkin, S.L.; Jamialahmadi, T.; Sahebkar, A. Targeting AMPK by Statins: A Potential Therapeutic Approach. Drugs 2021, 81, 923–933. [Google Scholar] [CrossRef]
- Guo, S.; Han, F.; Zhu, W. CD39—A bright target for cancer immunotherapy. BioMedicine 2022, 151, 113066. [Google Scholar] [CrossRef]
- Zhang, T.; Yu-Jing, L.; Ma, T. The immunomodulatory function of adenosine in sepsis. Front. Immunol. 2022, 13, 936547. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pharmacology of Adenosine Receptors: The State of the Art. Physiol. Rev. 2018, 98, 1591–1625. [Google Scholar] [CrossRef]
- Sun, C.; Wang, B.; Hao, S. Adenosine-A2A Receptor Pathway in Cancer Immunotherapy. Front. Immunol. 2022, 13, 837230. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Shevde, L.A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Lei, Y.; Li, J.-K.; Du, W.-X.; Li, R.-G.; Yang, J.; Li, J.; Li, F.; Tan, H.-B. Immune cells within the tumor microenvironment: Biological functions and roles in cancer immunotherapy. Cancer Lett. 2020, 470, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Guerra, L.; Bonetti, L.; Brenner, D. Metabolic Modulation of Immunity: A New Concept in Cancer Immunotherapy. Cell Rep. 2020, 32, 107848. [Google Scholar] [CrossRef] [PubMed]
- Sek, K.; Mølck, C.; Stewart, G.D.; Kats, L.; Darcy, P.K.; Beavis, P.A. Targeting Adenosine Receptor Signaling in Cancer Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3837. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Gessi, S.S.; Merighi, S.; Varani, K. Adenosine as a Multi-Signalling Guardian Angel in Human Diseases: When, Where and How Does It Exert Its Protective Effects? Trends Pharmacol. Sci. 2016, 37, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, N.; Jain, A.; Campana, D. Nk Cells for Cancer Immunotherapy. Nat. Rev. Drug Discov. 2020, 19, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Brauneck, F.; Seubert, E.; Wellbrock, J.; Wiesch, J.S.Z.; Duan, Y.; Magnus, T.; Bokemeyer, C.; Koch-Nolte, F.; Menzel, S.; Fiedler, W. Combined Blockade of Tigit and Cd39 or A2ar Enhances Nk-92 Cell-Mediated Cytotoxicity in Aml. Int. J. Mol. Sci. 2021, 22, 12919. [Google Scholar] [CrossRef]
- Young, A.; Ngiow, S.F.; Gao, Y.; Patch, A.-M.; Barkauskas, D.S.; Messaoudene, M.; Lin, G.; Coudert, J.D.; Stannard, K.A.; Zitvogel, L.; et al. A2ar Adenosine Signaling Suppresses Natural Killer Cell Maturation in the Tumor Microenvironment. Cancer Res. 2018, 78, 1003–1016. [Google Scholar] [CrossRef]
- Wang, J.; Matosevic, S. Adenosinergic signaling as a target for natural killer cell immunotherapy. J. Mol. Med. 2018, 96, 903–913. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, D.; Dong, H.; Chen, Q.; Dimitrova, D.I.; Rogers, T.J.; Sitkovsky, M.; Oppenheim, J.J. Adenosine A2a Receptors Induce Heterologous Desensitization of Chemokine Receptors. Blood 2006, 108, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Raskovalova, T.; Lokshin, A.; Huang, X.; Jackson, E.K.; Gorelik, E. Adenosine-Mediated Inhibition of Cytotoxic Activity and Cytokine Production by Il-2/Nkp46-Activated Nk Cells: Involvement of Protein Kinase a Isozyme I (Pka I). Immunol. Res. 2006, 36, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Neo, S.Y.; Yang, Y.; Record, J.; Ma, R.; Chen, X.; Chen, Z.; Tobin, N.P.; Blake, E.; Seitz, C.; Thomas, R.; et al. Cd73 Immune Checkpoint Defines Regulatory Nk Cells within the Tumor Microenvironment. J. Clin. Investig. 2020, 130, 1185–1198. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Jiménez-Cortegana, C.; Tay, A.H.; Wickström, S.; Galluzzi, L.; Lundqvist, A. NK cells and solid tumors: Therapeutic potential and persisting obstacles. Mol. Cancer 2022, 21, 206. [Google Scholar] [CrossRef] [PubMed]
- Lokshin, A.; Raskovalova, T.; Huang, X.; Zacharia, L.C.; Jackson, E.K.; Gorelik, E. Adenosine-Mediated Inhibition of the Cytotoxic Activity and Cytokine Production by Activated Natural Killer Cells. Cancer Res. 2006, 66, 7758–7765. [Google Scholar] [CrossRef] [PubMed]
- Raskovalova, T.; Huang, X.; Sitkovsky, M.; Zacharia, L.C.; Jackson, E.K.; Gorelik, E. Gs Protein-Coupled Adenosine Receptor Signaling and Lytic Function of Activated NK Cells. J. Immunol. 2005, 175, 4383–4391. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.M.; Lupo, K.B.; Wang, J.; Cao, J.; Utturkar, S.; Lanman, N.; Bernal-Crespo, V.; Jalal, S.; Pine, S.R.; Torregrosa-Allen, S.; et al. Engineered Natural Killer Cells Impede the Immunometabolic Cd73-Adenosine Axis in Solid Tumors. Elife 2022, 11, e73699. [Google Scholar] [CrossRef]
- Tay, A.H.M.; Prieto-Díaz, R.; Neo, S.; Tong, L.; Chen, X.; Carannante, V.; Önfelt, B.; Hartman, J.; Haglund, F.; Majellaro, M.; et al. A2B adenosine receptor antagonists rescue lymphocyte activity in adenosine-producing patient-derived cancer models. J. Immunother. Cancer 2022, 10, e004592. [Google Scholar] [CrossRef]
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic cells in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef]
- Cekic, C.; Sag, D.; Li, Y.; Theodorescu, D.; Strieter, R.M.; Linden, J. Adenosine A2B Receptor Blockade Slows Growth of Bladder and Breast Tumors. J. Immunol. 2012, 188, 198–205. [Google Scholar] [CrossRef]
- Novitskiy, S.V.; Ryzhov, S.; Zaynagetdinov, R.; Goldstein, A.E.; Huang, Y.; Tikhomirov, O.Y.; Blackburn, M.R.; Biaggioni, I.; Carbone, D.P.; Feoktistov, I.; et al. Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 2008, 112, 1822–1831. [Google Scholar] [CrossRef] [PubMed]
- Borodovsky, A.; Barbon, C.M.; Wang, Y.; Ye, M.; Prickett, L.; Chandra, D.; Shaw, J.; Deng, N.; Sachsenmeier, K.; Clarke, J.D.; et al. Small Molecule Azd4635 Inhibitor of a(2a)R Signaling Rescues Immune Cell Function Including Cd103(+) Dendritic Cells Enhancing Anti-Tumor Immunity. J. Immunother. Cancer 2020, 8, e000417. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-S.; Chiang, S.-F.; Chen, C.-Y.; Hong, W.-Z.; Chen, T.-W.; Chen, W.T.-L.; Ke, T.-W.; Yang, P.-C.; Liang, J.-A.; Shiau, A.; et al. Targeting CD73 increases therapeutic response to immunogenic chemotherapy by promoting dendritic cell maturation. Cancer Immunol. Immunother. 2023, 72, 2283–2297. [Google Scholar] [CrossRef] [PubMed]
- Ben Addi, A.; Lefort, A.; Hua, X.; Libert, F.; Communi, D.; Ledent, C.; Macours, P.; Tilley, S.L.; Boeynaems, J.-M.; Robaye, B. Modulation of murine dendritic cell function by adenine nucleotides and adenosine: Involvement of the A2B receptor. Eur. J. Immunol. 2008, 38, 1610–1620. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Xu, D.; Zang, J.; Zheng, X.; Zhao, Y.; Li, Y.; He, R.; Ruan, S.; Dong, H.; et al. Targeting the Negative Feedback of Adenosine-A2AR Metabolic Pathway by a Tailored Nanoinhibitor for Photothermal Immunotherapy. Adv. Sci. 2022, 9, e2104182. [Google Scholar] [CrossRef] [PubMed]
- Borst, J.; Ahrends, T.; Bąbała, N.; Melief, C.J.M.; Kastenmüller, W. CD4+ T cell help in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2018, 18, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Speiser, D.E.; Chijioke, O.; Schaeuble, K.; Munz, C. Cd4(+) T Cells in Cancer. Nat. Cancer 2023, 4, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhang, X.; Zheng, J.; Liu, Z.; Yang, Z.; Zhang, X. Overcoming High Level Adenosine-Mediated Immunosuppression by Dzd2269, a Potent and Selective A2ar Antagonist. J. Exp. Clin. Cancer Res. 2022, 41, 302. [Google Scholar] [CrossRef]
- Kawano, M.; Takagi, R.; Tokano, M.; Matsushita, S. Adenosine Induces Il-31 Secretion by T-Helper 2 Cells: Implication for the Effect of Adenosine on Atopic Dermatitis and Its Therapeutic Strategy. Biochem. Biophys. Res. Commun. 2023, 645, 47–54. [Google Scholar] [CrossRef]
- Lee, J.; Shin, K.-O.; Kim, Y.; Cho, J.; Lim, H.W.; Yoon, S.-I.; Lee, G.-S.; Ko, H.-J.; Kim, P.-H.; Uchida, Y.; et al. Cathelicidin-Related Antimicrobial Peptide Regulates CD73 Expression in Mouse Th17 Cells via p38. Cells 2020, 9, 1561. [Google Scholar] [CrossRef]
- Chalmin, F.; Mignot, G.; Bruchard, M.; Chevriaux, A.; Végran, F.; Hichami, A.; Ladoire, S.; Derangère, V.; Vincent, J.; Masson, D.; et al. Stat3 and Gfi-1 Transcription Factors Control Th17 Cell Immunosuppressive Activity via the Regulation of Ectonucleotidase Expression. Immunity 2012, 36, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Tokano, M.; Kawano, M.; Takagi, R.; Matsushita, S. Istradefylline, an Adenosine A2a Receptor Antagonist, Inhibits the Cd4(+) T-Cell Hypersecretion of Il-17a and Il-8 in Humans. Immunol. Med. 2022, 45, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Kurtz, C.C.; Black, S.G.; Ross, W.G.; Alam, M.S.; Linden, J.; Ernst, P.B. The A2B Adenosine Receptor Promotes Th17 Differentiation via Stimulation of Dendritic Cell IL-6. J. Immunol. 2011, 186, 6746–6752. [Google Scholar] [CrossRef] [PubMed]
- Mastelic-Gavillet, B.; Rodrigo, B.N.; Décombaz, L.; Wang, H.; Ercolano, G.; Ahmed, R.; Lozano, L.E.; Ianaro, A.; Derré, L.; Valerio, M.; et al. Adenosine mediates functional and metabolic suppression of peripheral and tumor-infiltrating CD8+ T cells. J. Immunother. Cancer 2019, 7, 257. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Feng, M.; Du, S.; Wei, X.; Song, H.; Yixin, X.; Song, J.; Wenxian, G. Adenosine Generated by Regulatory T Cells Induces CD8+T Cell Exhaustion in Gastric Cancer through A2aR Pathway. BioMed Res. Int. 2019, 2019, 4093214. [Google Scholar] [CrossRef] [PubMed]
- Heng, T.S.P.; The Immunological Genome Project Consortium; Painter, M.W.; Elpek, K.; Lukacs-Kornek, V.; Mauermann, N.; Turley, S.J.; Koller, D.; Kim, F.S.; Wagers, A.J.; et al. The Immunological Genome Project: Networks of gene expression in immune cells. Nat. Immunol. 2008, 9, 1091–1094. [Google Scholar] [CrossRef]
- Canale, F.P.; Ramello, M.C.; Núñez, N.; Furlan, C.L.A.; Bossio, S.N.; Serrán, M.G.; Boari, J.T.; del Castillo, A.; Ledesma, M.; Sedlik, C.; et al. CD39 Expression Defines Cell Exhaustion in Tumor-Infiltrating CD8+ T Cells. Cancer Res 2018, 78, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Zeng, D.-N.; Li, J.-Z.; Hua, Q.-M.; Xiao, Z.; He, C.; Mao, K.; Zhu, L.-Y.; Chu, Y.; Wen, W.-P.; et al. Targeting adenosinergic pathway enhances the anti-tumor efficacy of sorafenib in hepatocellular carcinoma. Hepatol. Int. 2020, 14, 80–95. [Google Scholar] [CrossRef]
- Briceño, P.; Rivas-Yañez, E.; Rosemblatt, M.V.; Parra-Tello, B.; Farías, P.; Vargas, L.; Simon, V.; Cárdenas, C.; Lladser, A.; Salazar-Onfray, F.; et al. CD73 Ectonucleotidase Restrains CD8+ T Cell Metabolic Fitness and Anti-tumoral Activity. Front. Cell Dev. Biol. 2021, 9, 638037. [Google Scholar] [CrossRef]
- Lai, Y.-P.; Kuo, L.-C.; Lin, B.-R.; Lin, H.-J.; Lin, C.-Y.; Chen, Y.-T.; Hsiao, P.-W.; Chang, H.-T.; Ko, P.C.-I.; Chen, H.-C.; et al. CD28 engagement inhibits CD73-mediated regulatory activity of CD8+ T cells. Commun. Biol. 2021, 4, 595. [Google Scholar] [CrossRef]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef] [PubMed]
- Stout, R.D.; Suttles, J. Functional plasticity of macrophages: Reversible adaptation to changing microenvironments. J. Leukoc. Biol. 2004, 76, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Rao, L.; Yao, H.; Wang, Z.; Ning, P.; Chen, X. Engineering Macrophages for Cancer Immunotherapy and Drug Delivery. Adv. Mater. 2020, 32, e2002054. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Chu, Y.; Li, Z.; Yu, X.; Huang, Z.; Xu, J.; Zheng, L. Tumor-derived adenosine promotes macrophage proliferation in human hepatocellular carcinoma. J. Hepatol. 2021, 74, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.; Ryan, N.; Alkhimovitch, A.; Siddiqui, A.; Oghumu, S. Inhibition of Pi3k Isoform P110gamma Increases Both Anti-Tumor and Immunosuppressive Responses to Aggressive Murine Head and Neck Squamous Cell Carcinoma with Low Immunogenicity. Cancers 2021, 13, 953. [Google Scholar] [CrossRef]
- Kaneda, M.M.; Messer, K.S.; Ralainirina, N.; Li, H.; Leem, C.J.; Gorjestani, S.; Woo, G.; Nguyen, A.V.; Figueiredo, C.C.; Foubert, P.; et al. Pi3kgamma Is a Molecular Switch That Controls Immune Suppression. Nature 2016, 539, 437–442. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, X.; Zhou, J.; Guo, J.; Liu, Y.; Liang, C.; Wang, W.; Xing, Y.; Wu, J.; Hu, D. A2aR on lung adenocarcinoma cells: A novel target for cancer therapy via recruiting and regulating tumor-associated macrophages. Chem. Interact. 2023, 382, 110543. [Google Scholar] [CrossRef]
- Cekic, C.; Day, Y.-J.; Sag, D.; Linden, J. Myeloid Expression of Adenosine A2A Receptor Suppresses T and NK Cell Responses in the Solid Tumor Microenvironment. Cancer Res. 2014, 74, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- Montalban Del Barrio, I.; Penski, C.; Schlahsa, L.; Stein, R.G.; Diessner, J.; Wockel, A.; Dietl, J.; Lutz, M.B.; Mittelbronn, M.; Wischhusen, J.; et al. Adenosine-Generating Ovarian Cancer Cells Attract Myeloid Cells Which Differentiate into Adenosine-Generating Tumor Associated Macrophages—A Self-Amplifying, Cd39- and Cd73-Dependent Mechanism for Tumor Immune Escape. J. Immunother. Cancer 2016, 4, 49. [Google Scholar] [CrossRef]
- Jin, R.; Liu, L.; Xing, Y.; Meng, T.; Ma, L.; Pei, J.; Cong, Y.; Zhang, X.; Ren, Z.; Wang, X.; et al. Dual Mechanisms of Novel Cd73-Targeted Antibody and Antibody-Drug Conjugate in Inhibiting Lung Tumor Growth and Promoting Antitumor Immune-Effector Function. Mol. Cancer Ther. 2020, 19, 2340–2352. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Ryzhov, S.; Novitskiy, S.V.; Goldstein, A.E.; Biktasova, A.; Blackburn, M.R.; Biaggioni, I.; Dikov, M.M.; Feoktistov, I. Adenosinergic Regulation of the Expansion and Immunosuppressive Activity of CD11b+Gr1+ Cells. J. Immunol. 2011, 187, 6120–6129. [Google Scholar] [CrossRef]
- Groth, C.; Hu, X.; Weber, R.; Fleming, V.; Altevogt, P.; Utikal, J.; Umansky, V. Immunosuppression mediated by myeloid-derived suppressor cells (MDSCs) during tumour progression. Br. J. Cancer 2019, 120, 16–25. [Google Scholar] [CrossRef]
- King, R.J.; Shukla, S.K.; He, C.; Vernucci, E.; Thakur, R.; Attri, K.S.; Dasgupta, A.; Chaika, N.V.; Mulder, S.E.; Abrego, J.; et al. CD73 induces GM-CSF/MDSC-mediated suppression of T cells to accelerate pancreatic cancer pathogenesis. Oncogene 2022, 41, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Chiu, D.K.; Tse, A.P.; Xu, I.M.; Di Cui, J.; Lai, R.K.; Li, L.L.; Koh, H.Y.; Tsang, F.H.; Wei, L.L.; Wong, C.M.; et al. Hypoxia Inducible Factor Hif-1 Promotes Myeloid-Derived Suppressor Cells Accumulation through Entpd2/Cd39l1 in Hepatocellular Carcinoma. Nat. Commun. 2017, 8, 517. [Google Scholar] [CrossRef]
- Sarkar, O.S.; Donninger, H.; Al Rayyan, N.; Chew, L.C.; Stamp, B.; Zhang, X.; Whitt, A.; Li, C.; Hall, M.; Mitchell, R.A.; et al. Monocytic MDSCs exhibit superior immune suppression via adenosine and depletion of adenosine improves efficacy of immunotherapy. Sci. Adv. 2023, 9, eadg3736. [Google Scholar] [CrossRef]
- Iannone, R.; Miele, L.; Maiolino, P.; Pinto, A.; Morello, S. Blockade of A2b Adenosine Receptor Reduces Tumor Growth and Immune Suppression Mediated by Myeloid-Derived Suppressor Cells in a Mouse Model of Melanoma. Neoplasia 2013, 15, 1400–1409. [Google Scholar] [CrossRef]
- Sorrentino, C.; Miele, L.; Porta, A.; Pinto, A.; Morello, S. Myeloid-derived suppressor cells contribute to A2B adenosine receptor-induced VEGF production and angiogenesis in a mouse melanoma model. Oncotarget 2015, 6, 27478–27489. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. What Are Regulatory T Cells (Treg) Regulating in Cancer and Why? In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2012; Volume 22, pp. 327–334. [Google Scholar]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) Cells in Cancer: Can Treg Cells Be a New Therapeutic Target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Park, H.J.; Son, J.; Lee, J.G.; Chung, K.Y.; Cho, N.H.; Shim, H.S.; Park, S.; Kim, G.; Yoon, H.I.; et al. Tumor microenvironment dictates regulatory T cell phenotype: Upregulated immune checkpoints reinforce suppressive function. J. Immunother. Cancer 2019, 7, 339. [Google Scholar] [CrossRef] [PubMed]
- Beavis, P.A.; Henderson, M.A.; Giuffrida, L.; Mills, J.K.; Sek, K.; Cross, R.S.; Davenport, A.J.; John, L.B.; Mardiana, S.; Slaney, C.Y.; et al. Targeting the adenosine 2A receptor enhances chimeric antigen receptor T cell efficacy. J. Clin. Investig. 2017, 127, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Maj, T.; Wang, W.; Crespo, J.; Zhang, H.; Wang, W.; Wei, S.; Zhao, L.; Vatan, L.; Shao, I.; Szeliga, W.; et al. Oxidative stress controls regulatory T cell apoptosis and suppressor activity and PD-L1-blockade resistance in tumor. Nat. Immunol. 2017, 18, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Huang, X.; Lu, M.; Zhang, G.; Han, X.; Liang, T. Transcriptional control of pancreatic cancer immunosuppression by metabolic enzyme CD73 in a tumor-autonomous and -autocrine manner. Nat. Commun. 2023, 14, 3364. [Google Scholar] [CrossRef]
- Su, Y.; Jackson, E.K.; Gorelik, E. Receptor desensitization and blockade of the suppressive effects of prostaglandin E2 and adenosine on the cytotoxic activity of human melanoma-infiltrating T lymphocytes. Cancer Immunol. Immunother. 2011, 60, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L.; Jackson, E.K. Adenosine and Prostaglandin E2 Production by Human Inducible Regulatory T Cells in Health and Disease. Front. Immunol. 2013, 4, 212. [Google Scholar] [CrossRef]
- Apetoh, L.; Quintana, F.J.; Pot, C.; Joller, N.; Xiao, S.; Kumar, D.; Burns, E.J.; Sherr, D.H.; Weiner, H.L.; Kuchroo, V.K. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nat. Immunol. 2010, 11, 854–861. [Google Scholar] [CrossRef]
- Chihara, N.; Madi, A.; Karwacz, K.; Awasthi, A.; Kuchroo, V.K. Differentiation and Characterization of Tr1 Cells. Curr. Protoc. Immunol. 2016, 113, 3.27.1–3.27.10. [Google Scholar] [CrossRef]
- Churov, A.; Zhulai, G. Targeting adenosine and regulatory T cells in cancer immunotherapy. Hum. Immunol. 2021, 82, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Mandapathil, M.; Hilldorfer, B.; Szczepanski, M.J.; Czystowska, M.; Szajnik, M.; Ren, J.; Lang, S.; Jackson, E.K.; Gorelik, E.; Whiteside, T.L. Generation and Accumulation of Immunosuppressive Adenosine by Human CD4+CD25highFOXP3+ Regulatory T Cells. J. Biol. Chem. 2010, 285, 7176–7186. [Google Scholar] [CrossRef] [PubMed]
- Aroua, N.; Boet, E.; Ghisi, M.; Nicolau-Travers, M.L.; Saland, E.; Gwilliam, R.; de Toni, F.; Hosseini, M.; Mouchel, P.L.; Farge, T.; et al. Extracellular Atp and Cd39 Activate Camp-Mediated Mitochondrial Stress Response to Promote Cytarabine Resistance in Acute Myeloid Leukemia. Cancer Discov. 2020, 10, 1544–1565. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-L.; Cai, Y.-Q.; Zhu, M.-C.; Xing, L.-J.; Wang, X. The yin and yang functions of extracellular ATP and adenosine in tumor immunity. Cancer Cell Int. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Schakel, L.; Schmies, C.C.; Idris, R.M.; Luo, X.; Lee, S.Y.; Lopez, V.; Mirza, S.; Vu, T.H.; Pelletier, J.; Sevigny, J.; et al. Nucleotide Analog Arl67156 as a Lead Structure for the Development of Cd39 and Dual Cd39/Cd73 Ectonucleotidase Inhibitors. Front. Pharmacol. 2020, 11, 1294. [Google Scholar] [CrossRef] [PubMed]
- Savio, L.E.B.; de Andrade Mello, P.; Figliuolo, V.R.; de Avelar Almeida, T.F.; Santana, P.T.; Oliveira, S.D.S.; Silva, C.L.M.; Feldbrugge, L.; Csizmadia, E.; Minshall, R.D.; et al. Cd39 Limits P2x7 Receptor Inflammatory Signaling and Attenuates Sepsis-Induced Liver Injury. J. Hepatol. 2017, 67, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.C.; Gabriely, G.; Rothhammer, V.; Mascanfroni, I.D.; Wheeler, M.A.; Chao, C.-C.; Gutiérrez-Vázquez, C.; Kenison, J.; Tjon, E.C.; Barroso, A.; et al. Control of tumor-associated macrophages and T cells in glioblastoma via AHR and CD39. Nat. Neurosci. 2019, 22, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Feng, L.; Mello, P.d.A.; Mao, C.; Near, R.; Csizmadia, E.; Chan, L.L.-Y.; Enjyoji, K.; Gao, W.; Zhao, H.; et al. Glycoengineered anti-CD39 promotes anticancer responses by depleting suppressive cells and inhibiting angiogenesis in tumor models. J. Clin. Investig. 2022, 132, e157431. [Google Scholar] [CrossRef]
- Kashyap, A.S.; Thelemann, T.; Klar, R.; Kallert, S.M.; Festag, J.; Buchi, M.; Hinterwimmer, L.; Schell, M.; Michel, S.; Jaschinski, F.; et al. Antisense oligonucleotide targeting CD39 improves anti-tumor T cell immunity. J. Immunother. Cancer 2019, 7, 67. [Google Scholar] [CrossRef]
- Da Silva, H.B.; Beura, L.K.; Wang, H.; Hanse, E.A.; Gore, R.; Scott, M.C.; Walsh, D.A.; Block, K.E.; Fonseca, R.; Yan, Y.; et al. The purinergic receptor P2RX7 directs metabolic fitness of long-lived memory CD8+ T cells. Nature 2018, 559, 264–268. [Google Scholar] [CrossRef]
- Casey, M.; Segawa, K.; Law, S.C.; Sabdia, M.B.; Nowlan, B.; Salik, B.; Lee, C.; Winterford, C.; Pearson, S.; Madore, J.; et al. Inhibition of CD39 unleashes macrophage antibody-dependent cellular phagocytosis against B-cell lymphoma. Leukemia 2023, 37, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, J.H.; Schuh, R.S.; Michels, L.R.; Iser, I.C.; Beckenkamp, L.R.; Roliano, G.G.; Lenz, G.S.; Scholl, J.N.; Sévigny, J.; Wink, M.R.; et al. Blockade of CD73 delays glioblastoma growth by modulating the immune environment. Cancer Immunol. Immunother. 2020, 69, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Lin, E.; Xie, W.; Flaifel, A.; Steinharter, J.A.; Gatof, E.N.S.; Bouchard, G.; Fleischer, J.H.; Martinez-Chanza, N.; Gray, C.; et al. Prognostic significance and immune correlates of CD73 expression in renal cell carcinoma. J. Immunother. Cancer 2020, 8, e001467. [Google Scholar] [CrossRef] [PubMed]
- Hammami, A.; Allard, D.; Allard, B.; Stagg, J. Targeting the adenosine pathway for cancer immunotherapy. Semin. Immunol. 2019, 42, 101304. [Google Scholar] [CrossRef] [PubMed]
- Battastini, A.M.O.; Figueiro, F.; Leal, D.B.R.; Doleski, P.H.; Schetinger, M.R.C. Cd39 and Cd73 as Promising Therapeutic Targets: What Could Be the Limitations? Front. Pharmacol. 2021, 12, 633603. [Google Scholar] [CrossRef] [PubMed]
- Jacoberger-Foissac, C.; Cousineau, I.; Bareche, Y.; Allard, D.; Chrobak, P.; Allard, B.; Pommey, S.; Messaoudi, N.; McNicoll, Y.; Soucy, G.; et al. Cd73 Inhibits Cgas-Sting and Cooperates with Cd39 to Promote Pancreatic Cancer. Cancer Immunol. Res. 2023, 22, 56–71. [Google Scholar] [CrossRef]
- Ferris, R.L.; Moskovitz, J.; Kunning, S.; Ruffin, A.T.; Reeder, C.; Ohr, J.; Gooding, W.E.; Kim, S.; Karlovits, B.J.; Vignali, D.A.; et al. Phase I Trial of Cetuximab, Radiotherapy, and Ipilimumab in Locally Advanced Head and Neck Cancer. Clin. Cancer Res. 2022, 28, 1335–1344. [Google Scholar] [CrossRef]
- Allard, D.; Allard, B.; Stagg, J. On the mechanism of anti-CD39 immune checkpoint therapy. J. Immunother. Cancer 2020, 8, e000186. [Google Scholar] [CrossRef]
- Keerthana, C.K.; Rayginia, T.P.; Shifana, S.C.; Anto, N.P.; Kalimuthu, K.; Isakov, N.; Anto, R.J. The role of AMPK in cancer metabolism and its impact on the immunomodulation of the tumor microenvironment. Front. Immunol. 2023, 14, 1114582. [Google Scholar] [CrossRef]
- Fullerton, M.D.; Galic, S.; Marcinko, K.; Sikkema, S.; Pulinilkunnil, T.; Chen, Z.-P.; O’Neill, H.M.; Ford, R.J.; Palanivel, R.; O’Brien, M.; et al. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nat. Med. 2013, 19, 1649–1654. [Google Scholar] [CrossRef]
- Marsin, A.-S.; Bertrand, L.; Rider, M.H.; Deprez, J.; Beauloye, C.; Vincent, M.F.; Van den Berghe, G.; Carling, D.; Hue, L. Phosphorylation and activation of heart PFK-2 by AMPK has a role in the stimulation of glycolysis during ischaemia. Curr. Biol. 2000, 10, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Li, C.-F.; Han, F.; Liu, C.; Zhang, A.; Hsu, C.-C.; Peng, D.; Zhang, X.; Jin, G.; Rezaeian, A.-H.; et al. Phosphorylation of PDHA by AMPK Drives TCA Cycle to Promote Cancer Metastasis. Mol. Cell 2020, 80, 263–278.e7. [Google Scholar] [CrossRef] [PubMed]
- Bodmer, D.; Levano-Huaman, S. Sesn2/Ampk/Mtor Signaling Mediates Balance between Survival and Apoptosis in Sensory Hair Cells under Stress. Cell Death Dis. 2017, 8, e3068. [Google Scholar] [CrossRef] [PubMed]
- Toyama, E.Q.; Herzig, S.; Courchet, J.; Lewis, T.L., Jr.; Loson, O.C.; Hellberg, K.; Young, N.P.; Chen, H.; Polleux, F.; Chan, D.C.; et al. Metabolism. Amp-Activated Protein Kinase Mediates Mitochondrial Fission in Response to Energy Stress. Science 2016, 351, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Hardie, D.G. New insights into activation and function of the AMPK. Nat. Rev. Mol. Cell Biol. 2023, 24, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhu, S.; Chen, P.; Hou, W.; Wen, Q.; Liu, J.; Xie, Y.; Liu, J.; Klionsky, D.J.; Kroemer, G.; et al. Ampk-Mediated Becn1 Phosphorylation Promotes Ferroptosis by Directly Blocking System X(C)(-) Activity. Curr. Biol. 2018, 28, 2388–2399.e5. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, R.H.; Acharya, S.; Ahn, J.-H.; Gu, Y.; Pandit, M.; Kim, J.-O.; Park, Y.-Y.; Kang, B.; Ko, H.-J.; Chang, J.-H. AMPK promotes antitumor immunity by downregulating PD-1 in regulatory T cells via the HMGCR/p38 signaling pathway. Mol. Cancer 2021, 20, 133. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Peng, D.; Cai, Z.; Lin, H.-K. AMPK signaling and its targeting in cancer progression and treatment. Semin. Cancer Biol. 2022, 85, 52–68. [Google Scholar] [CrossRef]
- Arruga, F.; Serra, S.; Vitale, N.; Guerra, G.; Papait, A.; Gyau, B.B.; Tito, F.; Efremov, D.; Vaisitti, T.; Deaglio, S. Targeting of the A2A adenosine receptor counteracts immunosuppression in vivo in a mouse model of chronic lymphocytic leukemia. Haematologica 2021, 106, 1343–1353. [Google Scholar] [CrossRef]
- Sidders, B.; Zhang, P.; Goodwin, K.; O’Connor, G.; Russell, D.L.; Borodovsky, A.; Armenia, J.; McEwen, R.; Linghu, B.; Bendell, J.C.; et al. Adenosine Signaling Is Prognostic for Cancer Outcome and Has Predictive Utility for Immunotherapeutic Response. Clin. Cancer Res. 2020, 26, 2176–2187. [Google Scholar] [CrossRef]
- Yang, R.; Elsaadi, S.; Misund, K.; Abdollahi, P.; Vandsemb, E.N.; Moen, S.H.; Kusnierczyk, A.; Slupphaug, G.; Standal, T.; Waage, A.; et al. Conversion of ATP to adenosine by CD39 and CD73 in multiple myeloma can be successfully targeted together with adenosine receptor A2A blockade. J. Immunother. Cancer 2020, 8, e000610. [Google Scholar] [CrossRef] [PubMed]
- Willingham, S.B.; Ho, P.Y.; Hotson, A.; Hill, C.M.; Piccione, E.C.; Hsieh, J.; Liu, L.; Buggy, J.J.; McCaffery, I.; Miller, R.A. A2AR Antagonism with CPI-444 Induces Antitumor Responses and Augments Efficacy to Anti–PD-(L)1 and Anti–CTLA-4 in Preclinical Models. Cancer Immunol. Res. 2018, 6, 1136–1149. [Google Scholar] [CrossRef] [PubMed]
- Festag, J.; Thelemann, T.; Schell, M.; Raith, S.; Michel, S.; Jaschinski, F.; Klar, R. Preventing Atp Degradation by Aso-Mediated Knockdown of Cd39 and Cd73 Results in A2ar-Independent Rescue of T Cell Proliferation. Mol. Ther. Nucleic Acids 2020, 21, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Bareche, Y.; Pommey, S.; Carneiro, M.; Buisseret, L.; Cousineau, I.; Thebault, P.; Chrobak, P.; Communal, L.; Allard, D.; Robson, S.C.; et al. High-dimensional analysis of the adenosine pathway in high-grade serous ovarian cancer. J. Immunother. Cancer 2021, 9, e001965. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zou, Y.; Du, B.; Li, W.; Yu, G.; Li, L.; Zhou, L.; Gu, X.; Song, S.; Liu, Y.; et al. Snp-Mediated Lncrna-Entpd3-As1 Upregulation Suppresses Renal Cell Carcinoma Via Mir-155/Hif-1alpha Signaling. Cell Death Dis. 2021, 12, 672. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.A.; Powell, J.D. Inhibition of the Adenosine Pathway to Potentiate Cancer Immunotherapy: Potential for Combinatorial Approaches. Annu. Rev. Med. 2021, 72, 331–348. [Google Scholar] [CrossRef]
- Guan, S.; Suman, S.; Amann, J.M.; Wu, R.; Carbone, D.P.; Wang, J.; Dikov, M.M. Metabolic reprogramming by adenosine antagonism and implications in non-small cell lung cancer therapy. Neoplasia 2022, 32, 100824. [Google Scholar] [CrossRef] [PubMed]
- Morello, S.; Miele, L. Targeting the adenosine A2b receptor in the tumor microenvironment overcomes local immunosuppression by myeloid-derived suppressor cells. OncoImmunology 2014, 3, e27989. [Google Scholar] [CrossRef]
- Lokhorst, H.M.; Plesner, T.; Laubach, J.P.; Nahi, H.; Gimsing, P.; Hansson, M.; Minnema, M.C.; Lassen, U.; Krejcik, J.; Palumbo, A.; et al. Targeting CD38 with Daratumumab Monotherapy in Multiple Myeloma. N. Engl. J. Med. 2015, 373, 1207–1219. [Google Scholar] [CrossRef]
- Krejcik, J.; Frerichs, K.A.; Nijhof, I.S.; van Kessel, B.; van Velzen, J.F.; Bloem, A.C.; Broekmans, M.E.; Zweegman, S.; van Meerloo, J.; Musters, R.J.; et al. Monocytes and Granulocytes Reduce CD38 Expression Levels on Myeloma Cells in Patients Treated with Daratumumab. Clin. Cancer Res. 2017, 23, 7498–7511. [Google Scholar] [CrossRef]
- Van de Donk, N.W.; Richardson, P.G.; Malavasi, F. Cd38 Antibodies in Multiple Myeloma: Back to the Future. Blood 2018, 131, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, O.; Belisario, D.C.; Compagno, M.; Giovinazzo, F.; Bracci, C.; Pastorino, U.; Horenstein, A.; Malavasi, F.; Ferracini, R.; Scala, S.; et al. CXCR4 Inhibition Counteracts Immunosuppressive Properties of Metastatic NSCLC Stem Cells. Front. Immunol. 2020, 11, 02168. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.H.M.; Lee, R.Y.; Goh, S.; Tay, I.S.Y.; Lim, X.; Lee, B.; Chew, V.; Li, H.; Tan, B.; Lim, S.; et al. Immunohistochemical Scoring of Cd38 in the Tumor Microenvironment Predicts Responsiveness to Anti-Pd-1/Pd-L1 Immunotherapy in Hepatocellular Carcinoma. J. Immunother Cancer 2020, 8, 02168. [Google Scholar] [CrossRef] [PubMed]
- Tewari, K.S.; Monk, B.J.; Vergote, I.; Miller, A.; de Melo, A.C.; Kim, H.-S.; Kim, Y.M.; Lisyanskaya, A.; Samouëlian, V.; Lorusso, D.; et al. Protocol Investigators for, and Engot Protocol En-Cx. Survival with Cemiplimab in Recurrent Cervical Cancer. N. Engl. J. Med. 2022, 386, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Boydell, E.; Sandoval, J.L.; Michielin, O.; Obeid, M.; Addeo, A.; Friedlaender, A. Neoadjuvant Immunotherapy: A Promising New Standard of Care. Int. J. Mol. Sci. 2023, 24, 11849. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Taube, J.M.; Pardoll, D.M. Neoadjuvant checkpoint blockade for cancer immunotherapy. Science 2020, 367, eaax0182. [Google Scholar] [CrossRef] [PubMed]
- Kamai, T.; Kijima, T.; Tsuzuki, T.; Nukui, A.; Abe, H.; Arai, K.; Yoshida, K.I. Increased Expression of Adenosine 2a Receptors in Metastatic Renal Cell Carcinoma Is Associated with Poorer Response to Anti-Vascular Endothelial Growth Factor Agents and Anti-Pd-1/Anti-Ctla4 Antibodies and Shorter Survival. Cancer Immunol. Immunother. 2021, 70, 2009–2021. [Google Scholar] [CrossRef] [PubMed]
- Perrot, I.; Michaud, H.A.; Giraudon-Paoli, M.; Augier, S.; Docquier, A.; Gros, L.; Courtois, R.; Dejou, C.; Jecko, D.; Becquart, O.; et al. Blocking Antibodies Targeting the Cd39/Cd73 Immunosuppressive Pathway Unleash Immune Responses in Combination Cancer Therapies. Cell Rep. 2019, 27, 2411–2425.e9. [Google Scholar] [CrossRef]
- Kim, M.; Min, Y.K.; Jang, J.; Park, H.; Lee, S.; Lee, C.H. Single-cell RNA sequencing reveals distinct cellular factors for response to immunotherapy targeting CD73 and PD-1 in colorectal cancer. J. Immunother. Cancer 2021, 9, e002503. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Zhao, X.; Xiao, J.; Bi, J.; Li, X.-Y.; Chen, G.; Lu, L. Review immune response of targeting CD39 in cancer. Biomark. Res. 2023, 11, 63. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Study ID | Conditions | Interventions | Phase | Start Date |
|---|---|---|---|---|
| NCT04306900 | Solid Tumor, Adult | COMBINATION_PRODUCT: TTX-030, budigalimab and mFOLFOX6|COMBINATION_PRODUCT: TTX-030, budigalimab and docetaxel|COMBINATION_PRODUCT: TTX-030 and mFOLFOX6|COMBINATION_PRODUCT: TTX-030 and budigalimab|COMBINATION_PRODUCT: TTX-030, budigalimab, nab-paclitaxel and gemcitabine|COMBINATION_PRODUCT: TTX-030 and pembrolizumab|COMBINATION_PRODUCT: TTX-030, nab-paclitaxel and gemcitabine|COMBINATION_PRODUCT: Budigalimab and mFOLFOX6 | PHASE1 | 30 March 2020 |
| NCT05374226 | Advanced Solid Tumors or Lymphomas | BIOLOGICAL: JS019 | PHASE1 | 31 March 2022 |
| NCT05508373 | Advanced Solid Tumors | BIOLOGICAL: JS019 | PHASE1 | 29 March 2022 |
| NCT05075564 | Advanced Solid Tumor | DRUG: Part 1 ES002023|DRUG: Part 2 ES002023 | PHASE1 | 23 December 2021 |
| NCT05381935 | Advanced Solid Tumor | DRUG: ES014|DRUG: ES014 | PHASE1 | 21 April 2023 |
| NCT04668300 | Metastatic Angiosarcoma|Metastatic Dedifferentiated Liposarcoma|Metastatic Osteosarcoma|Recurrent Angiosarcoma|Recurrent Dedifferentiated Liposarcoma|Recurrent Osteosarcoma|Refractory Dedifferentiated Liposarcoma|Refractory Osteosarcoma | BIOLOGICAL: Durvalumab|BIOLOGICAL: Oleclumab | PHASE2 | 26 November 2020 |
| NCT04148937 | Advanced Cancer | DRUG: LY3475070|DRUG: Pembrolizumab | PHASE1 | 16 January 2020 |
| NCT05174585 | Solid Tumor | BIOLOGICAL: JAB-BX102 (anti-CD73 monoclonal antibody)|BIOLOGICAL: pembrolizumab (anti-PD-1 monoclonal antibody) | PHASE1|PHASE2 | 18 August 2022 |
| NCT05329766 | Gastrointestinal Tract Malignancies | DRUG: Domvanalimab|DRUG: Quemliclustat|DRUG: Zimberelimab|DRUG: Fluorouracil|DRUG: Leucovorin|DRUG: Oxaliplatin | PHASE2 | 10 June 2022 |
| NCT04797468 | Advanced Solid Tumor | DRUG: HLX23 | PHASE1 | 18 July 2022 |
| NCT05688215 | Borderline Resectable Pancreatic Adenocarcinoma|Locally Advanced Pancreatic Ductal Adenocarcinoma | PROCEDURE: Biospecimen Collection|PROCEDURE: Computed Tomography|PROCEDURE: Core Biopsy|DRUG: Fluorouracil|DRUG: Irinotecan|DRUG: Leucovorin|DRUG: Leucovorin Calcium|DRUG: Oxaliplatin|DRUG: Quemliclustat|DRUG: Zimberelimab | PHASE1|PHASE2 | 7 March 2023 |
| NCT04572152 | Advanced or Metastatic Solid Tumors | BIOLOGICAL: AK119|BIOLOGICAL: AK104 | PHASE1 | 18 January 2021 |
| NCT03954704 | Advanced Solid Tumors | DRUG: Dalutrafusp alfa|DRUG: mFOLFOX6 Regimen|DRUG: dalutrafusp alfa | PHASE1 | 3 June 2019 |
| NCT04672434 | Metastatic Cancer|Solid Tumor | DRUG: Sym021|DRUG: Sym024 | PHASE1 | 19 November 2020 |
| NCT03875573 | Luminal B | DRUG: Durvalumab|RADIATION: Stereotactic Body Radiotherapy|DRUG: Oleclumab | PHASE2 | 6 November 2019 |
| NCT03454451 | Non-Small-Cell Lung Cancer|Renal Cell Cancer|Colorectal Cancer|Triple Negative Breast Cancer|Cervical Cancer|Ovarian Cancer|Pancreatic Cancer|Endometrial Cancer|Sarcoma|Squamous Cell Carcinoma of the Head and Neck|Bladder Cancer|Metastatic Castration Resistant Prostate Cancer|Non-hodgkin Lymphoma | DRUG: CPI-006|DRUG: CPI-006 + ciforadenant|DRUG: CPI-006 + pembrolizumab|DRUG: CPI-006|DRUG: CPI-006 + ciforadenant|DRUG: CPI-006 + pembrolizumab | PHASE1 | 25 April 2018 |
| NCT05632328 | Advanced Pancreatic Ductal Adenocarcinoma|Pancreatic Ductal Adenocarcinoma|Pancreatic Cancer | DRUG: AGEN1423|DRUG: Balstilimab|DRUG: Gemcitabine|DRUG: Nab-paclitaxel | PHASE2 | 23 April 2023 |
| NCT05559541 | Solid Tumor, Adult | DRUG: AK119|DRUG: AK104 | PHASE1|PHASE2 | 15 December 2022 |
| NCT05689853 | Solid Tumor, Adult | DRUG: AK119|DRUG: AK112 | PHASE1|PHASE2 | 14 April 2023 |
| NCT04940286 | Borderline Resectable Pancreatic Adenocarcinoma|Resectable Pancreatic Adenocarcinoma|Stage IA Pancreatic Cancer AJCC v8|Stage IB Pancreatic Cancer AJCC v8|Stage IIA Pancreatic Cancer AJCC v8|Stage IIB Pancreatic Cancer AJCC v8 | BIOLOGICAL: Durvalumab|DRUG: Gemcitabine|DRUG: Nab-paclitaxel|BIOLOGICAL: Oleclumab | PHASE2 | 28 September 2021 |
| NCT04660812 | Metastatic Colorectal Cancer | DRUG: AB680|DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Bevacizumab|DRUG: m-FOLFOX-6 regimen|DRUG: Regorafenib | PHASE1|PHASE2 | 10 May 2021 |
| NCT05143970 | Metastatic Cancer|Metastatic Breast Cancer|Metastatic Pancreatic Cancer|Metastatic Gastric Cancer|Metastatic Lung Cancer|Metastatic Ovary Cancer|Oesophageal Cancer|Endometrial Cancer|Advanced Solid Tumor | DRUG: IPH5301 ALONE OR IN COMBINATION WITH CHEMOTHERAPY AND TRASTUZUMAB | PHASE1 | 21 January 2022 |
| NCT05431270 | Metastatic Cancer|Refractory Cancer|Non-Small-Cell Lung Cancer|Pancreatic Adenocarcinoma|Pancreatic Neoplasms|Lung Cancer | DRUG: PT199|DRUG: Anti-PD-1 monoclonal antibody | PHASE1 | 23 June 2022 |
| NCT05119998 | Solid Tumor | DRUG: IBI325 + sintilimab|DRUG: IBI325 | PHASE1 | 8 February 2022 |
| NCT05246995 | Solid Tumor | DRUG: IBI325 + Sintilimab | PHASE1 | 23 March 2022 |
| Study ID | Conditions | Interventions | Phase | Start Date |
|---|---|---|---|---|
| NCT04969315 | Renal Cell Cancer|Castrate Resistant Prostate Cancer|Non-Small-Cell Lung Cancer | DRUG: TT-10 | PHASE1|PHASE2 | 1 May 2023 |
| NCT04976660 | Colorectal Cancer|Gastric Cancer|Hepatocellular Carcinoma|Pancreatic Cancer | DRUG: TT-4 | PHASE1|PHASE2 | 15 December 2022 |
| Study ID | Conditions | Interventions | Phase | Start Date |
|---|---|---|---|---|
| NCT03319667 | Plasma Cell Myeloma | DRUG: Isatuximab SAR650984|DRUG: Bortezomib|DRUG: Lenalidomide|DRUG: Dexamethasone | PHASE3 | 7 December 2017 |
| NCT03275285 | Plasma Cell Myeloma | DRUG: isatuximab SAR650984|DRUG: carfilzomib|DRUG: dexamethasone | PHASE3 | 25 October 2017 |
| NCT05461209 | Relapsed/Refractory Multiple Myeloma | DRUG: Talquetamab|DRUG: Belantamab Mafodotin | PHASE3 | 20 October 2022 |
| NCT05572515 | Relapsed or Refractory Multiple Myeloma | DRUG: Teclistamab|DRUG: Pomalidomide|DRUG: Bortezomib|DRUG: Dexamethasone|DRUG: Carfilzomib | PHASE3 | 29 March 2023 |
| NCT04270409 | Plasma Cell Myeloma | DRUG: Isatuximab SAR650984|DRUG: Lenalidomide|DRUG: Dexamethasone | PHASE3 | 16 June 2020 |
| NCT02419118 | Multiple Myeloma | DRUG: Daratumumab|DRUG: Lenalidomide|DRUG: Dexamethasone | PHASE2|PHASE3 | 15 January 2023 |
| NCT02990338 | Plasma Cell Myeloma | DRUG: Isatuximab|DRUG: Pomalidomide|DRUG: Dexamethasone | PHASE3 | 22 December 2016 |
| NCT03937635 | Smoldering Plasma Cell Myeloma | BIOLOGICAL: Daratumumab|DRUG: Dexamethasone|DRUG: Lenalidomide|OTHER: Quality-of-Life Assessment|OTHER: Questionnaire Administration | PHASE3 | 16 September 2019 |
| Study ID | Conditions | Interventions | Phase | Start Date |
|---|---|---|---|---|
| NCT04306900 | Solid Tumor, Adult | COMBINATION_PRODUCT: TTX-030, budigalimab and mFOLFOX6|COMBINATION_PRODUCT: TTX-030, budigalimab and docetaxel|COMBINATION_PRODUCT: TTX-030 and mFOLFOX6|COMBINATION_PRODUCT: TTX-030 and budigalimab|COMBINATION_PRODUCT: TTX-030, budigalimab, nab-paclitaxel and gemcitabine|COMBINATION_PRODUCT: TTX-030 and pembrolizumab|COMBINATION_PRODUCT: TTX-030, nab-paclitaxel and gemcitabine|COMBINATION_PRODUCT: Budigalimab and mFOLFOX6 | PHASE1 | 30 March 2020 |
| NCT03884556 | Solid Tumor|Lymphoma | DRUG: TTX-030|DRUG: Pembrolizumab|DRUG: Gemcitabine|DRUG: nab paclitaxel | PHASE1 | 10 April 2019 |
| NCT05177770 | Metastatic Castration-resistant Prostate Cancer|Prostate Cancer | DRUG: SRF617|DRUG: etrumadenant|DRUG: zimberelimab | PHASE2 | 17 January 2022 |
| NCT04381832 | Prostatic Neoplasms, Castration-Resistant|Androgen-Resistant Prostatic Neoplasms|Castration Resistant Prostatic Neoplasms|Prostatic Cancer, Castration-Resistant | DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Quemliclustat|DRUG: Enzalutamide|DRUG: Docetaxel|DRUG: SG | PHASE1|PHASE2 | 7 July 2020 |
| NCT05329766 | Gastrointestinal Tract Malignancies | DRUG: Domvanalimab|DRUG: Quemliclustat|DRUG: Zimberelimab|DRUG: Fluorouracil|DRUG: Leucovorin|DRUG: Oxaliplatin | PHASE2 | 10 June 2022 |
| NCT05688215 | Borderline Resectable Pancreatic Adenocarcinoma|Locally Advanced Pancreatic Ductal Adenocarcinoma | PROCEDURE: Biospecimen Collection|PROCEDURE: Computed Tomography|PROCEDURE: Core Biopsy|DRUG: Fluorouracil|DRUG: Irinotecan|DRUG: Leucovorin|DRUG: Leucovorin Calcium|DRUG: Oxaliplatin|DRUG: Quemliclustat|DRUG: Zimberelimab | PHASE1|PHASE2 | 7 March 2023 |
| NCT04660812 | Metastatic Colorectal Cancer | DRUG: AB680|DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Bevacizumab|DRUG: m-FOLFOX-6 regimen|DRUG: Regorafenib | PHASE1|PHASE2 | 10 May 2021 |
| NCT04381832 | Prostatic Neoplasms, Castration-Resistant|Androgen-Resistant Prostatic Neoplasms|Castration Resistant Prostatic Neoplasms|Prostatic Cancer, Castration-Resistant | DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Quemliclustat|DRUG: Enzalutamide|DRUG: Docetaxel|DRUG: SG | PHASE1|PHASE2 | 7 July 2020 |
| NCT04660812 | Metastatic Colorectal Cancer | DRUG: AB680|DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Bevacizumab|DRUG: m-FOLFOX-6 regimen|DRUG: Regorafenib | PHASE1|PHASE2 | 10 May 2021 |
| NCT05177770 | Metastatic Castration-resistant Prostate Cancer|Prostate Cancer | DRUG: SRF617|DRUG: etrumadenant|DRUG: zimberelimab | PHASE2 | 17 January 2022 |
| NCT04381832 | Prostatic Neoplasms, Castration-Resistant|Androgen-Resistant Prostatic Neoplasms|Castration Resistant Prostatic Neoplasms|Prostatic Cancer, Castration-Resistant | DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Quemliclustat|DRUG: Enzalutamide|DRUG: Docetaxel|DRUG: SG | PHASE1|PHASE2 | 7 July 2020 |
| NCT04660812 | Metastatic Colorectal Cancer | DRUG: AB680|DRUG: Etrumadenant|DRUG: Zimberelimab|DRUG: Bevacizumab|DRUG: m-FOLFOX-6 regimen|DRUG: Regorafenib | PHASE1|PHASE2 | 10 May 2021 |
| NCT05177770 | Metastatic Castration-resistant Prostate Cancer|Prostate Cancer | DRUG: SRF617|DRUG: etrumadenant|DRUG: zimberelimab | PHASE2 | 17 January 2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, J.; Zhang, J.; Wang, J. The Immune Regulatory Role of Adenosine in the Tumor Microenvironment. Int. J. Mol. Sci. 2023, 24, 14928. https://doi.org/10.3390/ijms241914928
Xing J, Zhang J, Wang J. The Immune Regulatory Role of Adenosine in the Tumor Microenvironment. International Journal of Molecular Sciences. 2023; 24(19):14928. https://doi.org/10.3390/ijms241914928
Chicago/Turabian StyleXing, Jianlei, Jinhua Zhang, and Jinyan Wang. 2023. "The Immune Regulatory Role of Adenosine in the Tumor Microenvironment" International Journal of Molecular Sciences 24, no. 19: 14928. https://doi.org/10.3390/ijms241914928