
Multiple Sclerosis-Associated Gut Microbiome in the Israeli Diverse Populations: Associations with Ethnicity, Gender, Disability Status, Vitamin D Levels, and Mediterranean Diet

Abstract
:1. Introduction
2. Results
2.1. Subject Demographics
2.2. General Microbiome Composition
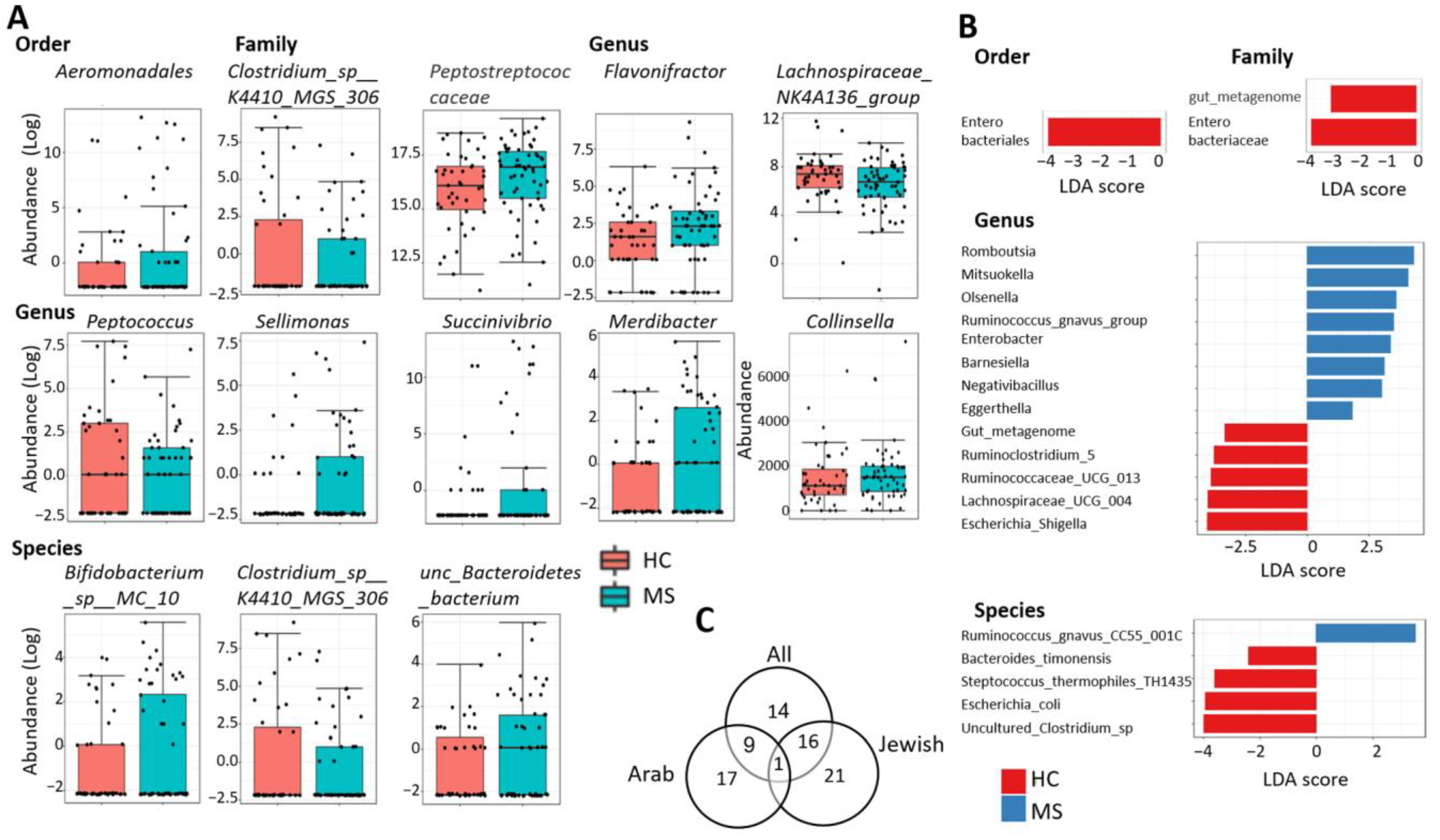
2.3. Differentially Abundant Taxa
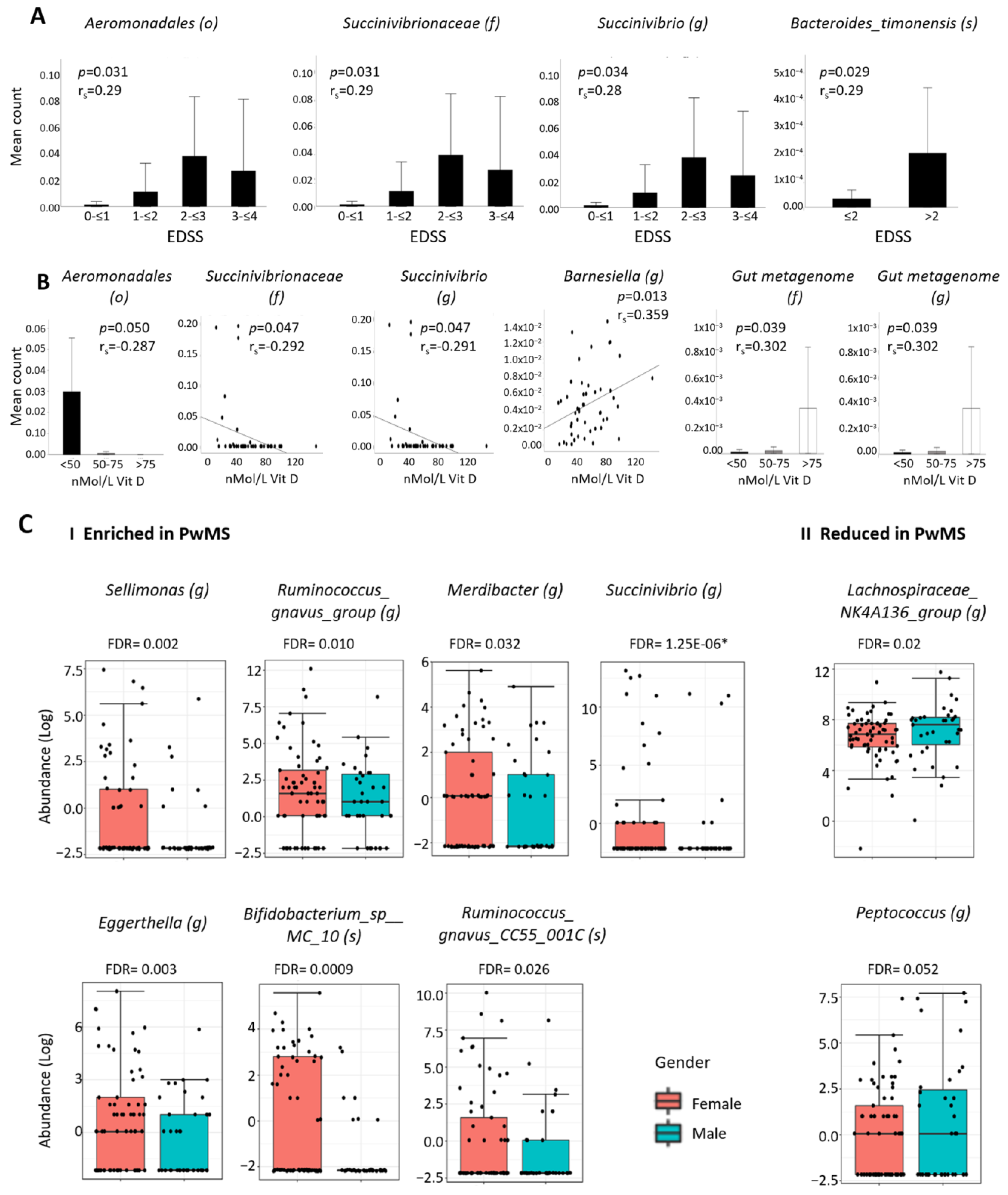
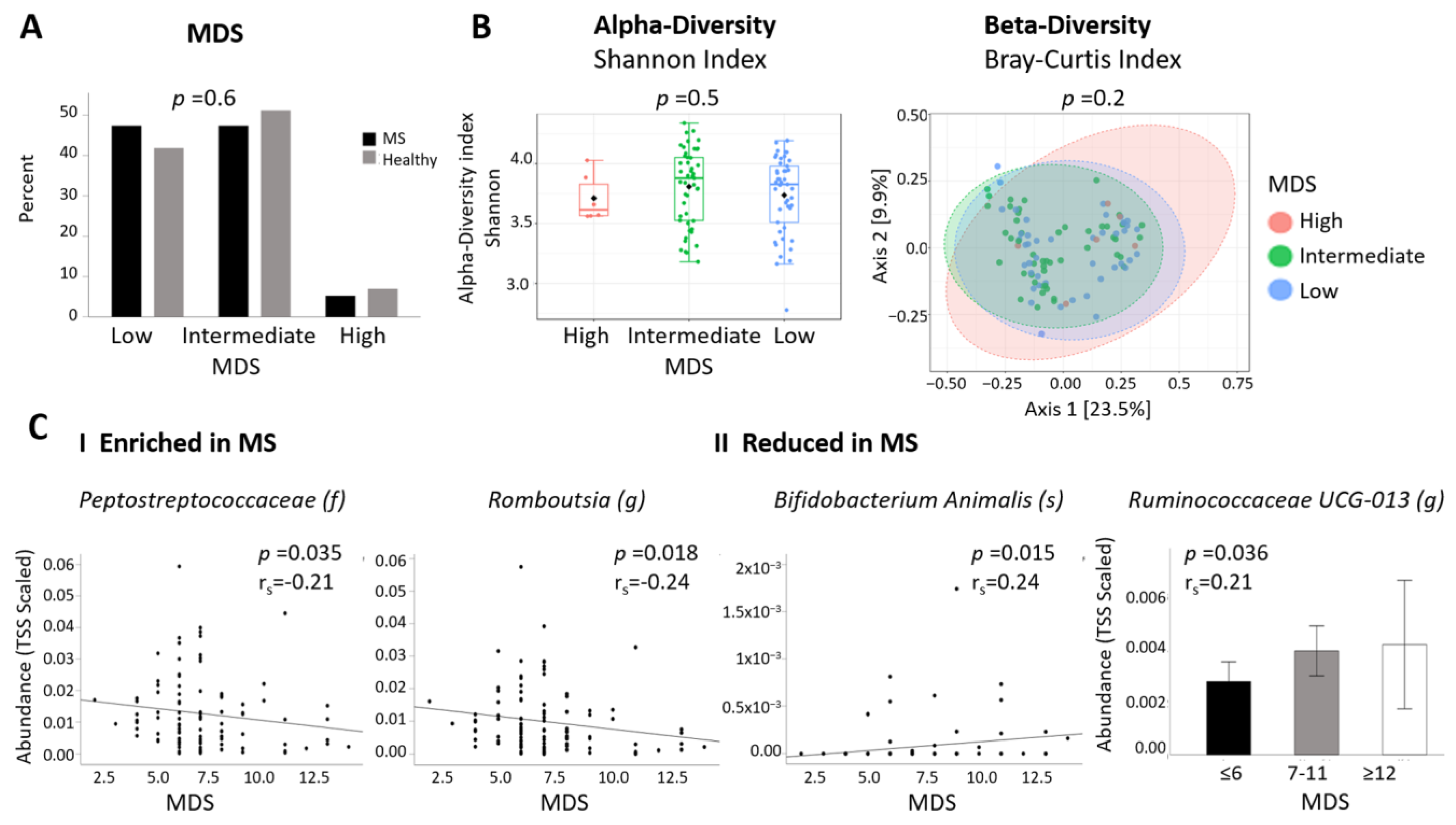
2.4. Associations between MS-Differentially Abundant Microbiota and Clinical Factors
2.5. Differential Abundant Taxa and Gender
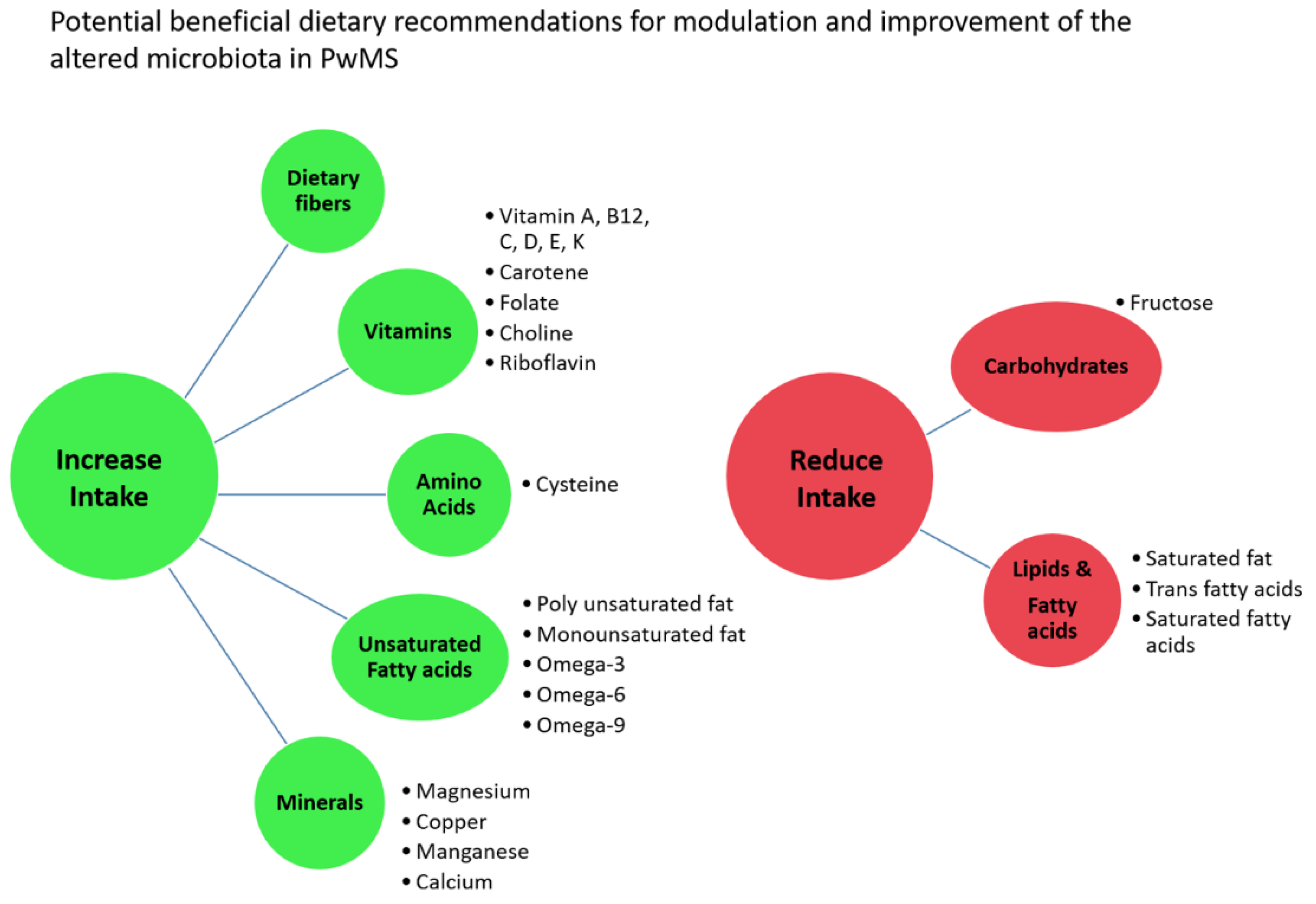
2.6. Differential Abundant Taxa and Dietary Data
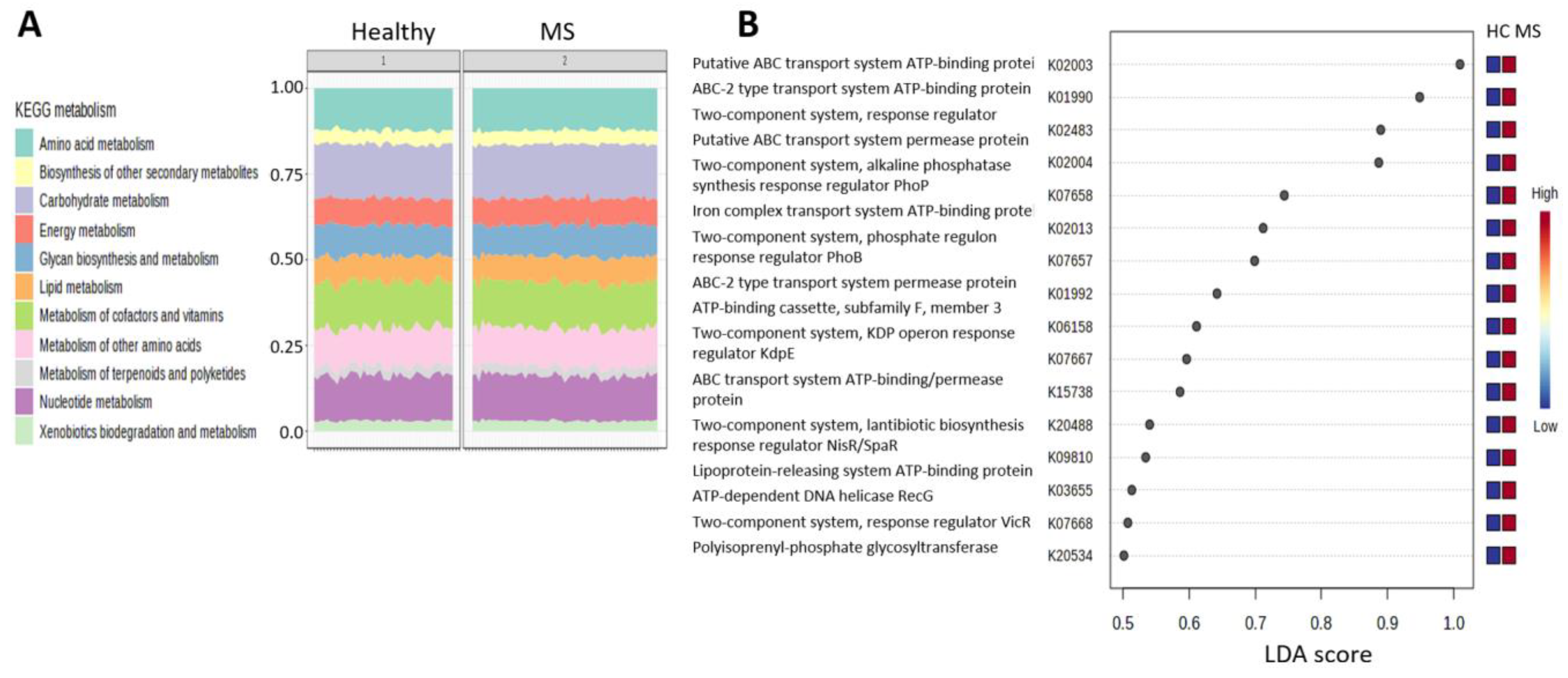
2.7. Functional Pathways
3. Discussion
4. Materials and Methods
4.1. Recruitment of Participant and Sample Collection
4.2. Microbial DNA Extration
4.3. Statistical Analysis
4.4. Functional Analysis
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olsson, T.; Barcellos, L.F.; Alfredsson, L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 2016, 13, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Dopkins, N.; Nagarkatti, P.S.; Nagarkatti, M. The role of gut microbiome and associated metabolome in the regulation of neuroinflammation in multiple sclerosis and its implications in attenuating chronic inflammation in other inflammatory and autoimmune disorders. Immunology 2018, 154, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Weiner, H.L. Microbiota Signaling Pathways that Influence Neurologic Disease. Neurotherapeutics 2018, 15, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Abou-el-hassan, H.; Maghzi, A.H.; Vincentini, J.; Weiner, H.L. The sex-specific interaction of the microbiome in neurodegenerative diseases. Brain Res. 2019, 1724, 146385. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pachecoa, Y.; Acosta-Ampudiaa, Y.; Ramírez-Santanaa, C.; Leungc, P.S.C.; Ansaric, A.A.; Gershwinc, M.E.; Anaya, J. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.; Forbes, J.D.; Zhu, F.; Bernstein, C.N.; Domselaar, G.V.; Graham, M.; Waubante, E.; Tremlett, H. The multiple sclerosis gut microbiota: A systematic review. Mult. Scler. Relat. Disord. 2020, 37, 101427. [Google Scholar] [CrossRef]
- Berer, K.; Mues, M.; Koutrolos, M.; AlRasbi, Z.; Boziki, M.; Johner, C.; Wekerle, H.; Krishnamoorthy, G.; Rasbi, Z.A.; Boziki, M.; et al. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination—With comments. Nature 2011, 479, 538–541. [Google Scholar] [CrossRef]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef]
- Berer, K.; Gerdes, L.A.; Cekanaviciute, E.; Jiac, X.; Xiaod, L.; Xiad, Z.; Liud, C.; Klotze, L.; Staufferf, U.; Baranzini, S.E.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10719–10724. [Google Scholar] [CrossRef]
- Cresci, G.A.; Bawden, E. Gut microbiome: What we do and don’t know. Nutr. Clin. Pract. 2015, 30, 734–746. [Google Scholar] [CrossRef]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut microbiome composition in obese and non-obese persons: A systematic review and meta-analysis. Nutrients 2022, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Lin, Q.; Dorsett, Y.; Ghezzi, L.; Liu, Z.; Pan, Y.; Chen, K.; Han, Y.; Li, Z.; Xiao, H.; et al. Alterations of host-gut microbiome interactions in multiple sclerosis. EBioMedicine 2022, 76, 103798. [Google Scholar] [CrossRef] [PubMed]
- Nearing, J.T.; Douglas, G.M.; Hayes, M.G.; MacDonald, J.; Desai, D.K.; Allward, N.; Jones, C.M.A.; Wright, R.J.; Dhanani, A.S.; Comeau, A.M.; et al. Microbiome differential abundance methods produce different results across 38 datasets. Nat. Commun. 2022, 13, 342. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. Available online: https://www.wma.net/policies-post/wma-statement-on-vitamin-d-insufficiency/ (accessed on 28 January 2023).
- Filippi, M.; Bar-Or, A.; Piehl, F.; Preziosa, P.; Solari, A.; Vukusic, S.; Rocca, M.A. Multiple Sclerosis. Nat. Rev. Dis. Prim. 2018, 4, 43. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: Prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene sequences. Environ. Microbiomes 2020, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Fadrosh, D.W.; Faruqi, A.A.; Zhu, F.; Hart, J.; Roalstad, S.; Graves, J.; Lynch, S.; Waubant, E.; on behalf of the US Network of Pediatric MS Centers. Gut microbiota in early pediatric multiple sclerosis: A case-control study. Eur. J. Neurol. 2016, 23, 1308–1321. [Google Scholar] [CrossRef]
- Tremlett, H.; Zhu, F.; Arnold, D.; Bar-Or, A.; Bernstein, C.N.; Bonner, C.; Forbes, J.D.; Graham, M.; Hart, J.; Knox, N.C.; et al. The gut microbiota in pediatric multiple sclerosis and demyelinating syndromes. Ann. Clin. Transl. Neurol. 2021, 8, 2252–2269. [Google Scholar] [CrossRef]
- Yadav, M.; Ali, S.; Shrode, R.L.; Shahi, S.K.; Jensen, S.N.; Hoang, J.; Cassidy, S.; Olalde, H.; Guseva, N.; Paullus, M.; et al. Multiple sclerosis patients have an altered gut mycobiome and increased fungal to bacterial richness. PLoS ONE 2022, 17, e0264556. [Google Scholar] [CrossRef]
- Tankou, S.K.; Regev, K.; Healy, B.C.; Tjon, E.; Laghi, L.; Cox, L.M.; Kivisäkk, P.; Pierre, I.V.; Hrishikesh, L.; Gandhi, R.; et al. A probiotic modulates the microbiome and immunity in multiple sclerosis. Ann. Neurol. 2018, 83, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar] [CrossRef]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Soldan, M.M.P.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar] [CrossRef] [PubMed]
- Miyake, S.; Kim, S.S.W.S.; Suda, W.; Oshima, K.; Nakamura, M.; Matsuoka, T.; Chihara, N.; Tomita, A.; Sato, W.; Kim, S.W.; et al. Dysbiosis in the gut microbiota of patients with multiple sclerosis, with a striking depletion of species belonging to clostridia XIVa and IV clusters. PLoS ONE 2015, 10, e0137429. [Google Scholar] [CrossRef] [PubMed]
- Ventura, R.E.; Iizumi, T.; Battaglia, T.; Liu, M.; Perez-Perez, G.I.; Herbert, I.; Blaser, M.J. Gut microbiome of treatment-naïve MS patients of different ethnicities early in disease course. Sci. Rep. 2019, 9, 16396. [Google Scholar] [CrossRef]
- Central Bureau of Statistics. Population of Israel on the Eve of 2023. Available online: https://www.cbs.gov.il/en/mediarelease/pages/2022/population-of-israel-on-the-eve-of-2023.aspx (accessed on 15 September 2023).
- Alter, M.; Kahana, E.; Zilber, N.; Ariel Miller and for the Israeli MS Study Group. Multiple sclerosis frequency in Israel’s diverse populations. Neurology 2006, 66, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Benedek, G.; Paperna, T.; Avidan, N.; Lejbkowicz, I.; Oksenberg, J.R.; Wang, J.; Brautbar, C.; Israel, S.; Miller, A. Opposing effects of the HLA-DRB1*0301-DQB1*0201 haplotype on the risk for multiple sclerosis in diverse Arab populations in Israel. Genes. Immun. 2010, 11, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Siegel, M.; Paperna, T.; Lejbkowicz, I.; Petrou, P.; Shahien, R.; Karussis, D.; Lavi, I.; Dishon, S.; Rawashdeh, H. Miller, AMultiple sclerosis in diverse populations: Characteristics in distinct Arab ethnicities in Israel. Mult. Scler. 2012, 18, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Budhram, A.; Parvathy, S.; Kremenchutzky, M.; Silverman, M. Breaking down the gut microbiome composition in multiple sclerosis. Mult. Scler. J. 2017, 23, 628–636. [Google Scholar] [CrossRef]
- Ling, Z.; Cheng, Y.; Yan, X.; Shao, L.; Liu, X.; Zhou, D.; Zhang, L.; Yu, K.; Zhao, L. Alterations of the Fecal Microbiota in Chinese Patients With Multiple Sclerosis. Front. Immunol. 2020, 11, 590783. [Google Scholar] [CrossRef]
- Duscha, A.; Gisevius, B.; Hirschberg, S.; Yissachar, N.; Stangl, G.I.; Eilers, E.; Bader, V.; Haase, S.; Kaisler, J.; David, C.; et al. Propionic Acid Shapes the Multiple Sclerosis Disease Course by an Immunomodulatory Mechanism. Cell 2020, 180, 1067–1080.e16. [Google Scholar] [CrossRef]
- Reynders, T.; Devolder, L.; Valles-Colomer, M.; Van Remoortel, A.; Joossens, M.; De Keyser, J.; Nagels, G.; D’hooghe, M.; Raes, J. Gut microbiome variation is associated to Multiple Sclerosis phenotypic subtypes. Ann. Clin. Transl. Neurol. 2020, 7, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Maghzi, A.H.; Liu, S.; Tankou, S.K.; Dhang, F.H.; Willocq, V.; Song, A.; Wasén, C.; Tauhid, S.; Chu, R.; et al. Gut Microbiome in Progressive Multiple Sclerosis. Ann. Neurol. 2021, 89, 1195–1211. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.R.; He, P.; Lei, S.F. Novel microbiota-related gene set enrichment analysis identified osteoporosis associated gut microbiota from autoimmune diseases. J. Bone Miner. Metab. 2021, 39, 984–996. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.; Mendozzi, L.; Amico, F.D.; Saresella, M.; Rampelli, S.; Piancone, F.; La Rosa, F.; Marventano, I.; Clerici, M.; d’Arma, A.; et al. Influence of a High-Impact Multidimensional Rehabilitation Program on the Gut Microbiota of Patients with Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 7173. [Google Scholar] [CrossRef]
- Ezendam, J.; De Klerk, A.; Gremmer, E.R.; van Loveren, H. Effects of Bifidobacterium animalis administered during lactation on allergic and autoimmune responses in rodents. Clin. Exp. Immunol. 2008, 154, 424–431. [Google Scholar] [CrossRef]
- Horton, M.K.; Mccauley, K.; Fadrosh, D.; Fujimura, K.; Graves, J.; Ness, J.; Wheeler, Y.; Gorman, M.P.; Benson, L.A.; Weinstock-Guttman, B.; et al. Gut microbiome is associated with multiple sclerosis activity in children. Ann. Clin. Transl. Neurol. 2021, 8, 1867–1883. [Google Scholar] [CrossRef]
- Sanchez, J.M.S.; DePaula-Silva, A.B.; Libbey, J.E.; Fujinami, R.S. Role of diet in regulating the gut microbiota and multiple sclerosis. Clin. Immunol. 2022, 235, 108379. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek. Int. J. General. Mol. Microbiol. 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Katz Sand, I.; Zhu, Y.; Ntranos, A.; Clemente, J.C.; Cekanaviciute, E.; Brandstadter, R.; Crabtree-Hartman, E.; Singh, S.; Bencosme, Y.; Debelius, J.; et al. Disease-modifying therapies alter gut microbial composition in MS. Neurol. Neuroimmunol. NeuroInflamm. 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Levi, I.; Gurevich, M.; Perlman, G.; Magalashvili, D.; Menascu, S.; Bar, B.; Godneva, A.; Zahavi, L.; Chermon, D.; Kosower, N.; et al. Potential role of indolelactate and butyrate in multiple sclerosis revealed by integrated microbiome-metabolome analysis. Cell Reports Med. 2021, 2, 100246. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Syed, Y.A.; Khan, M.R. Understanding the Role of the Gut Microbiome in Brain Development and Its Association With Neurodevelopmental Psychiatric Disorders. Front. Cell Dev. Biol. 2022, 10, 880544. [Google Scholar] [CrossRef]
- Pinna, N.K.; Anjana, R.M.; Saxena, S.; Dutta, A.; Gnanaprakash, V.; Rameshkumar, G.; Aswath, S.; Raghavan, S.; Rani, C.S.S.; Radha, V.; et al. Trans-ethnic gut microbial signatures of prediabetic subjects from India and Denmark. Genome Med. 2021, 13, 36. [Google Scholar] [CrossRef]
- Tecer, D.; Gogus, F.; Id, A.K.; Erdogan, M.; Hasanreisoglu, M.; Ergin, A.; Karakan, T.; Kozan, R.; Coban, S.; Diker, K.S. Succinivibrionaceae is dominant family in fecal microbiota of Behcet’s Syndrome patients with uveitis. PLoS ONE 2020, 15, e0241691. [Google Scholar] [CrossRef] [PubMed]
- Cosorich, I.; Dalla-Costa, G.; Sorini, C.; Ferrarese, R.; Messina, M.J.; Dolpady, J.; Radice, E.; Mariani, A.; Testoni, P.A.; Canducci, F.; et al. High frequency of intestinal T H 17 cells correlates with microbiota alterations and disease activity in multiple sclerosis. Sci. Adv. 2017, 3, e1700492. [Google Scholar] [CrossRef] [PubMed]
- Karni, A.; Kahana, E.; Zilber, N.; Abramsky, O.; Alter, M.; Karussis, D. The frequency of multiple sclerosis in Jewish and Arab populations in greater Jerusalem. Neuroepidemiology 2003, 22, 82–86. [Google Scholar] [CrossRef]
- Saaroni, H.; Sigal, A.; Lejbkowicz, I.; Miller, A. Mediterranean weather conditions and exacerbations of multiple sclerosis. Neuroepidemiology 2010, 35, 142–151. [Google Scholar] [CrossRef]
- Tremlett, H.; Fadrosh, D.W.; Faruqi, A.A.; Hart, J.; Roalstad, S.; Graves, J.; Lynch, S.; Waubant, E.; Aaen, G.; Belman, A.; et al. Gut microbiota composition and relapse risk in pediatric MS: A pilot study. J. Neurol. Sci. 2016, 363, 153–157. [Google Scholar] [CrossRef]
- Sintzel, M.B.; Rametta, M.; Reder, A.T. Vitamin D and Multiple Sclerosis: A Comprehensive Review. Neurol. Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef]
- Pierrot-deseilligny, C.; Souberbielle, J. Is hypovitaminosis D one of the environmental risk factors for multiple sclerosis? Brain 2010, 133, 1869–1888. [Google Scholar] [CrossRef]
- Golan, D.; Staun-Ram, E.; Glass-Marmor, L.; Lavi, I.; Rozenberg, O.; Dishon, S.; Barak, M.; Ish-Shalom, S.; Miller, A. The influence of vitamin D supplementation on melatonin status in patients with multiple sclerosis. Brain Behav. Immun. 2013, 32, 180–185. [Google Scholar] [CrossRef]
- Golan, D.; Halhal, B.; Glass-Marmor, L.; Staun-Ram, E.; Rozenberg, O.; Lavi, I.; Dishon, S.; Barak, M.; Ish-Shalom, S.; Miller, A. Vitamin D supplementation for patients with multiple sclerosis treated with interferon-beta: A randomized controlled trial assessing the effect on flu-like symptoms and immunomodulatory properties. BMC Neurol. 2013, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Ghareghani, M.; Reiter, R.J.; Zibara, K.; Farhadi, N. Latitude, Vitamin D, Melatonin, and Gut Microbiota Act in Concert to Initiate Multiple Sclerosis: A New Mechanistic Pathway. Front. Immunol. 2018, 9, 2484. [Google Scholar] [CrossRef] [PubMed]
- Riccio, P.; Rossano, R. Diet, Gut Microbiota, and Vitamins D + A in Multiple Sclerosis. Neurotherapeutics 2018, 15, 75–91. [Google Scholar] [CrossRef]
- Petrak, F.; Herpertz, S.; Hirsch, J.; Röhrig, B.; Donati-Hirsch, I.; Juckel, G.; Meier, J.J.; Gatermann, S. Gut microbiota differs in composition between adults with type 1 diabetes with or without depression and healthy control participants: A case-control study. BMC Microbiol. 2022, 22, 169. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Sanchez, N.; Dunn, S.E. Potential biological contributers to the sex difference in multiple sclerosis progression. Front. Immunol. 2023, 14, 1175874. [Google Scholar] [CrossRef] [PubMed]
- Harbo, H.F.; Gold, R.; Tintora, M. Sex and gender issues in multiple sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar] [CrossRef]
- Fish, E. The X-files in immunity: Sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J. Autoimmun. 2018, 92, 12–34. [Google Scholar] [CrossRef]
- Gold, S.M.; Voskuhl, R.R. Estrogen and testosterone therapies in multiple sclerosis. Prog. Brain Res. 2009, 75, 239–251. [Google Scholar]
- Bouman, A.; Heineman, M.J.; Faas, M.M. Sex hormones and the immune response in humans. Hum. Reprod. Update 2005, 11, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Selmi, C. The X in sex: How autoimmune diseases revolve around sex chromosomes. Best. Pract. Res. Clin. Rheumatol. 2008, 22, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Borgo, F.; Garbossa, S.; Riva, A.; Severgnini, M.; Luigiano, C.; Benetti, A.; Pontiroli, A.E.; Morace, G.; Borghi, E. Body Mass Index and Sex Affect Diverse Microbial Niches within the Gut. Front. Microbiol. 2018, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Naito, Y.; Inoue, R.; Kashiwagi, S.; Uchiyama, K.; Mizushima, K.; Tsuchiya, S.; Dohi, O.; Yoshida, N.; Kamada, K.; et al. Differences in gut microbiota associated with age, sex, and stool consistency in healthy Japanese subjects. J. Gastroenterol. 2019, 54, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, M.; Xue, J.; Huang, J.; Zhuang, R.; Zhou, X.; Zhang, H.; Fu, Q.; Hao, Y. Body Mass Index Differences in the Gut Microbiota Are Gender Specific. Front. Microbiol. 2018, 9, 1250. [Google Scholar] [CrossRef]
- Ortana, E.; Pierdominici, M.; Maselli, A.; Veroni, C.; Aloisi, F.; Shoenfeld, Y. Sex-based differences in autoimmune diseases. Ann. Ist. Super. Sanità 2016, 52, 205–212. [Google Scholar]
- Benedek, G.; Zhang, J.; Nguyen, H.; Kent, G.; Seifert, H.A.; Davin, S.; Stauffer, P.; Vandenbark, A.A.; Karstens, L.; Asquith, M.; et al. Estrogen protection against EAE modulates the microbiota and mucosal-associated regulatory cells. J. Neuroimmunol. 2017, 310, 51–59. [Google Scholar] [CrossRef]
- Markle, J.G.M.; Frank, D.N.; Mortin-Toth, S.; Robertson, C.E.; Feazel, L.M.; Rolle-Kampczyk, U.; Von Bergen, M.; McCoy, K.D.; Macpherson, A.J.; Danska, J.S. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science (80-) 2013, 339, 1084–1088. [Google Scholar] [CrossRef]
- Stoiloudis, P.; Kesidou, E.; Bakirtzis, C.; Sintila, S.A.; Konstantinidou, N.; Boziki, M.; Grigoriadis, N. The Role of Diet and Interventions on Multiple Sclerosis: A Review. Nutrients 2022, 14, 1150. [Google Scholar] [CrossRef]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Sánchez, B.; Margolles, A.; González, S. Mediterranean diet and faecal microbiota: A transversal study. Food Funct. 2016, 7, 2347–2356. [Google Scholar] [CrossRef]
- Bilenko, N.; Fraser, D.; Vardi, H.; Shai, I.; Shahar, D.R. Mediterranean diet and cardiovascular diseases in an Israeli population. Prev. Med. 2005, 40, 299–305. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2015, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.H.; May, C.; Wilck, N.; et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef]
- Elsayed, N.S.; Aston, P.; Bayanagari, V.R.; Shukla, S.K. The gut microbiome molecular mimicry piece in the multiple sclerosis puzzle. Front. Immunol. 2022, 13, 972160. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wang, Q.; Wu, Q.; Mao-Draayer, Y.; Kim, C.H. Bidirectional regulatory potentials of short-chain fatty acids and their G-protein-coupled receptors in autoimmune neuroinflammation. Sci. Rep. 2019, 9, 8837. [Google Scholar] [CrossRef] [PubMed]
- Noto, D.; Miyake, S. Gut dysbiosis and multiple sclerosis. Clin. Immunol. 2022, 235, 108380. [Google Scholar] [CrossRef]
- Esposito, S.; Sparaco, M.; Maniscalco, G.T.; Signoriello, E.; Signoriello, E.; Lanzillo, R.; Russo, C.; Carmisciano, L.; Cepparulo, S.; Lavorgna, L.; et al. Lifestyle and Mediterranean diet adherence in a cohort of Southern Italian patients with Multiple Sclerosis. Mult. Scler. Relat. Disord. 2021, 47, 102636. [Google Scholar] [CrossRef]
- Öztürk, Y.E.; Helvaci, E.M.; Kaya, P.S.; Terzi, M. Is Mediterranean diet associated with multiple sclerosis related symptoms and fatigue severity? Nutr. Neurosci. 2023, 26, 228–234. [Google Scholar] [CrossRef]
- Katz Sand, I.; Benn, E.K.T.; Fabian, M.; Fitzgerald, K.C.; Digga, E.; Deshpande, R.; Miller, A.; Gallo, S.; Arab, L. Randomized-controlled trial of a modified Mediterranean dietary program for multiple sclerosis: A pilot study. Mult. Scler. Relat. Disord. 2019, 36, 101403. [Google Scholar] [CrossRef]
- Bates, D.E.; Cartlidge, N.E.F.; French, J.M.; Jackson, M.J.; Nightingale, S.; Shaw, D.A.; Smith, S.; Woo, E.; Hawkins, S.A.; Millar, J.H.D. A double-blind controlled trial of long chain n-3 polyunsaturated fatty acids in the treatment of multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 1989, 52, 18–22. [Google Scholar] [CrossRef]
- Saresella, M.; Mendozzi, L.; Rossi, V.; Mazzali, F.; Piancone, F.; LaRosa, F.; Marventano, I.; Caputo, D.; Felis, G.E.; Clerici, M. Immunological and clinical effect of diet modulation of the gut microbiome in multiple sclerosis patients: A pilot study. Front. Immunol. 2017, 8, 1391. [Google Scholar] [CrossRef] [PubMed]
- Løken-Amsrud, K.I.; Myhr, K.M.; Bakke, S.J.; Beiske, A.G.; Bjerve, K.S.; Bjørnarå, B.T.; Hovdal, H.; Lilleås, F.; Midgard, R.; Pedersen, T.; et al. Retinol levels are associated with magnetic resonance imaging outcomes in multiple sclerosis. Mult. Scler. J. 2013, 19, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.; Ritzmann, F.; Eckstein, M.; Huch, M.; Briviba, K.; Behsnilian, D.; Neve, H.; Franz, C.M.A.P. Quantification of Slackia and Eggerthella spp. in Human Feces and Adhesion of Representatives Strains to Caco-2 Cells. Front. Microbiol. 2016, 7, 658. [Google Scholar] [CrossRef]
- Braccia, D.J.; Jiang, X.; Pop, M.; Hall, A.B. The Capacity to Produce Hydrogen Sulfide (H2S) via Cysteine Degradation Is Ubiquitous in the Human Gut Microbiome. Front. Microbiol. 2021, 12, 705583. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; Wilkinson, S.A.; Callaway, L.K.; McIntyre, H.D.; Morrison, M.; Dekker Nitert, M. Low dietary fiber intake increases Collinsella abundance in the gut microbiota of overweight and obese pregnant women. Gut Microbes 2018, 9, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Mena-Vázquez, N.; Ruiz-Limón, P.; Moreno-Indias, I.; Manrique-Arija, S.; Tinahones, F.J.; Fernández-Nebro, A. Expansion of rare and harmful lineages is associated with established rheumatoid arthritis. J. Clin. Med. 2020, 9, 1044. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef]
- Fitzgerald, K.C.; Smith, M.D.; Kim, S.; Sotirchos, E.S.; Kornberg, M.D.; Douglas, M.; Nourbakhsh, B.; Graves, J.; Rattan, R.; Poisson, L.; et al. Multi-omic evaluation of metabolic alterations in multiple sclerosis identifies shifts in aromatic amino acid metabolism. Cell Rep. Med. 2021, 2, 100424. [Google Scholar] [CrossRef]
- Quintana, F.J.; Basso, A.S.; Iglesias, A.H.; Korn, T.; Farez, M.F.; Bettelli, E.; Caccamo, M.; Oukka, M.; Weiner, H.L. Control of T reg and T H 17 cell differentiation by the aryl hydrocarbon receptor. Nature 2008, 453, 65–72. [Google Scholar] [CrossRef]
- Rothhammer, V.; Mascanfroni, I.D.; Lukas, B.; Takenaka, M.C.; Kenison, J.E.; Mayo, L.; Chao, C.C.; Patel, B.; Yan, R.; Blain, M.; et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef]
- Schrijver, I.A.; Van Meurs, M.; Melief, M.J.; Ang, C.W.; Buljevac, D.; Ravid, R.; Hazenberg, M.P.; Laman, J.D. Bacterial peptidoglycan and immune reactivity in the central nervous system in multiple sclerosis. Brain 2001, 124, 1544–1554. [Google Scholar] [CrossRef] [PubMed]
- Kodikara, S.; Ellul, S.; Le Cao, K.A. Statistical challenges in longitudinal microbiome data analysis. Brief. Bioinform. 2022, 23, bbac273. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Ben-Avraham, S.; Kohn, E.; Tepper, S.; Lubetzky, R.; Mandel, D.; Berkovitch, M.; Shahar, D.R. Ultra-processed food (UPF) intake in pregnancy and maternal and neonatal outcomes. Eur. J. Nutr. 2023, 62, 1403–1413. [Google Scholar] [CrossRef]
- Abu-Saad, K.; Endevelt, R.; Goldsmith, R.; Freedman, L.S.; Ziv, A.; Alpert, G.; Atamna, A.; Kalter-Leibovici, O. Adaptation and predictive utility of a Mediterranean diet screener score. Clin. Nutr. 2019, 38, 2928–2935. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, G.; Ewald, J.; Pang, Z.; Shiri, T.; Xia, J. MicrobiomeAnalyst 2.0: Comprehensive statistical, functional and integrative analysis of microbiome data. Nucleic Acids Res. 2023, 51, W310–W318. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MS (N = 57) | Healthy Controls (N = 43) | p-Value | |
|---|---|---|---|
| Age | 33.6 ± 1.4 | 38.1 ± 1.8 | 0.06 |
| Female (%) | 70.2 | 58.1 | 0.2 |
| BMI (kg/m2) | 25.4 ± 0.6 | 24.7 ± 0.5 | 0.6 |
| Smoking (%) | 35.1 | 25.6 | 0.3 |
| Vegetarian N (%) | 1 (1.8) | 1 (2.3) | 0.8 |
| Ethnicity N (%) | 0.06 | ||
| Jewish | 31 (54.4) | 32 (74.4) | |
| Arab | 26 (45.6) | 11 (25.6) | |
| MDS | 6.95 ± 0.3 | 7.33 ± 0.4 | 0.4 |
| MDS N (%) | 0.6 | ||
| Low (≤6) | 27 (47.4) | 18 (41.9) | |
| Intermediate (7–11) | 27 (47.4) | 22 (51.2) | |
| High (≥12) | 3 (5.3) | 3 (7.0) | |
| Vitamin D (ng/mL) | 44.5 ± 3.7 | ||
| Disease Duration (months) | 2.6 ± 0.6 | ||
| EDSS mean ± SE | 1.56 ± 0.14 | ||
| Annual relapse rate mean ± SE | 0.85 ± 0.07 |
| Differentially Abundant Taxa | EdgeR | DeSeq2 | Metag.Seq | LefSe | Highest | |||
|---|---|---|---|---|---|---|---|---|
| FDR | FC | FDR | FC | FDR | p-Value | LDA | in | |
| Order | ||||||||
| Aeromonadales | 0.059 | 4.7 | 0.002 | 26.1 | ns | ns | - | MS |
| Enterobacteriales | ns | - | ns | - | ns | 0.007 | −4.1 | HC |
| Family | ||||||||
| Clostridium_sp__K4410_MGS_306 | 0.019 | 0.31 | ns | - | 0.022 | ns | - | HC |
| Enterobacteriaceae | ns | - | ns | - | ns | 0.007 | −4.1 | HC |
| gut_metagenome | 1.10−7 | 0.18 | ns | - | 1.10−5 | 0.018 | −3.4 | HC |
| Peptostreptococcaceae | ns | - | ns | - | ns | 0.048 * | 4.3 * | MS |
| Succinivibrionaceae | 0.055 | 4.5 | 0.002 | 42.8 | ns | ns | - | MS |
| Genus | ||||||||
| Azospirillum_sp__47_25 | 3.39−7 | 0.16 | ns | - | 0.021 | ns | - | HC |
| Barnesiella | ns | - | ns | - | ns | 0.034 | 3.1 | MS |
| Collinsella | ns | - | ns | - | 0.096 | 0.045 * | 4.5 * | MS |
| Eggerthella | ns | - | ns | - | ns | 0.014 | 1.8 | MS |
| Enterobacter | 0.013 | 3.5 | ns | - | ns | 0.020 | 3.4 | MS |
| Escherichia_Shigella | 0.003 Ϯ | 0.18 Ϯ | ns | - | ns | 0.018 | −4.1 | HC |
| Flavonifractor | 0.0006 | 4.0 | 0.077 | 2.5 | ns | ns | - | MS |
| gut_metagenome | 1.59−6 | 0.20 | ns | - | 6.28−5 | 0.018 | −3.4 | HC |
| Lachnospiraceae_UCG_003 | 0.0007 | 3.6 | ns | - | 0.023 | ns | ns | MS |
| Lachnospiraceae_UCG_004 | ns | - | ns | - | ns | 0.007 | −4.0 | HC |
| Lachnospiraceae_NK4A136_group | 0.016 | 0.53 | 0.085 * | 0.58 * | ns | ns | - | HC |
| Merdibacter | 0.0014 | 2.5 | 0.036 | 3.7 | 0.0005 | ns | - | MS |
| Mitsuokella | 6.88−7 | 13.6 | 5.02−6 | 56.3 | 1.25−5 | 0.048 | 4.1 | MS |
| Negativibacillus | 0.013 | 2.2 | 0.099 | 2.3 | 0.049 | 0.042 | 3.0 | MS |
| Olsenella | 0.050 | 2.5 | ns | - | 0.023 | 0.031 | 3.6 | MS |
| Peptococcus | 0.054 | 0.47 | 0.032 * | 0.22 * | ns | ns | - | HC |
| Romboutsia | ns | - | ns | - | ns | 0.017 | 4.3 | MS |
| Ruminiclostridium_5 | ns | - | ns | - | ns | 0.049 | −3.8 | HC |
| Ruminococcaceae_UCG_013 | 0.055 | 0.60 | 0.099 | 0.60 | ns | 0.011 | −3.9 | HC |
| Ruminococcus_gnavus_group | ns | - | 4.77−7 | 7.5 | ns | 0.019 | 3.5 | MS |
| Sellimonas | 0.006 | 3.3 | ns | - | 0.050 | ns | - | MS |
| Succinivibrio | 0.008 | 6.8 | 0.009 | 57.0 | 0.060 | ns | - | MS |
| Tyzzerella_4 | 1.59−7 | 0.12 | ns | - | 0.088 | ns | - | HC |
| Species | ||||||||
| Bacteroides_timonensis | ns | - | ns | - | ns | 0.019 | −2.4 | HC |
| Bifidobacterium_animalis | 0.011 | 0.44 | ns | - | 0.002 | ns | - | HC |
| Bifidobacterium_sp__MC_10 | 0.092 | 1.8 | ns | - | 0.003 | ns | - | MS |
| Clostridium_sp__K4410_MGS_306 | 0.001 | 0.24 | ns | - | 0.037 | ns | - | HC |
| Escherichia_coli | ns | - | 0.043 | 2.9 | ns | 0.024 | −3.9 | HC |
| Negativibacillus_massiliensis | 0.0003 | 3.6 | ns | - | 8.01−5 | ns | - | MS |
| Ruminococcus_gnavus_CC55_001C | 0.099 | 2.6 | 0.0003 | 18.4 | 0.071 | 0.009 | 3.5 | MS |
| Streptococcus_thermophilus_TH1435 | ns | - | ns | - | ns | 0.028 | −3.6 | HC |
| uncultured_Bacteroidetes_bacterium | 0.001 | 2.5 | 0.032 | 3.6 | 0.016 | ns | - | MS |
| uncultured_Clostridium_sp_1 | 0.085 | 0.6 | 0.053 Ϯ | 0.6 | ns | 0.013 | −3.9 | HC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nitzan, Z.; Staun-Ram, E.; Volkowich, A.; Miller, A. Multiple Sclerosis-Associated Gut Microbiome in the Israeli Diverse Populations: Associations with Ethnicity, Gender, Disability Status, Vitamin D Levels, and Mediterranean Diet. Int. J. Mol. Sci. 2023, 24, 15024. https://doi.org/10.3390/ijms241915024
Nitzan Z, Staun-Ram E, Volkowich A, Miller A. Multiple Sclerosis-Associated Gut Microbiome in the Israeli Diverse Populations: Associations with Ethnicity, Gender, Disability Status, Vitamin D Levels, and Mediterranean Diet. International Journal of Molecular Sciences. 2023; 24(19):15024. https://doi.org/10.3390/ijms241915024
Chicago/Turabian StyleNitzan, Zehavit, Elsebeth Staun-Ram, Anat Volkowich, and Ariel Miller. 2023. "Multiple Sclerosis-Associated Gut Microbiome in the Israeli Diverse Populations: Associations with Ethnicity, Gender, Disability Status, Vitamin D Levels, and Mediterranean Diet" International Journal of Molecular Sciences 24, no. 19: 15024. https://doi.org/10.3390/ijms241915024
APA StyleNitzan, Z., Staun-Ram, E., Volkowich, A., & Miller, A. (2023). Multiple Sclerosis-Associated Gut Microbiome in the Israeli Diverse Populations: Associations with Ethnicity, Gender, Disability Status, Vitamin D Levels, and Mediterranean Diet. International Journal of Molecular Sciences, 24(19), 15024. https://doi.org/10.3390/ijms241915024