
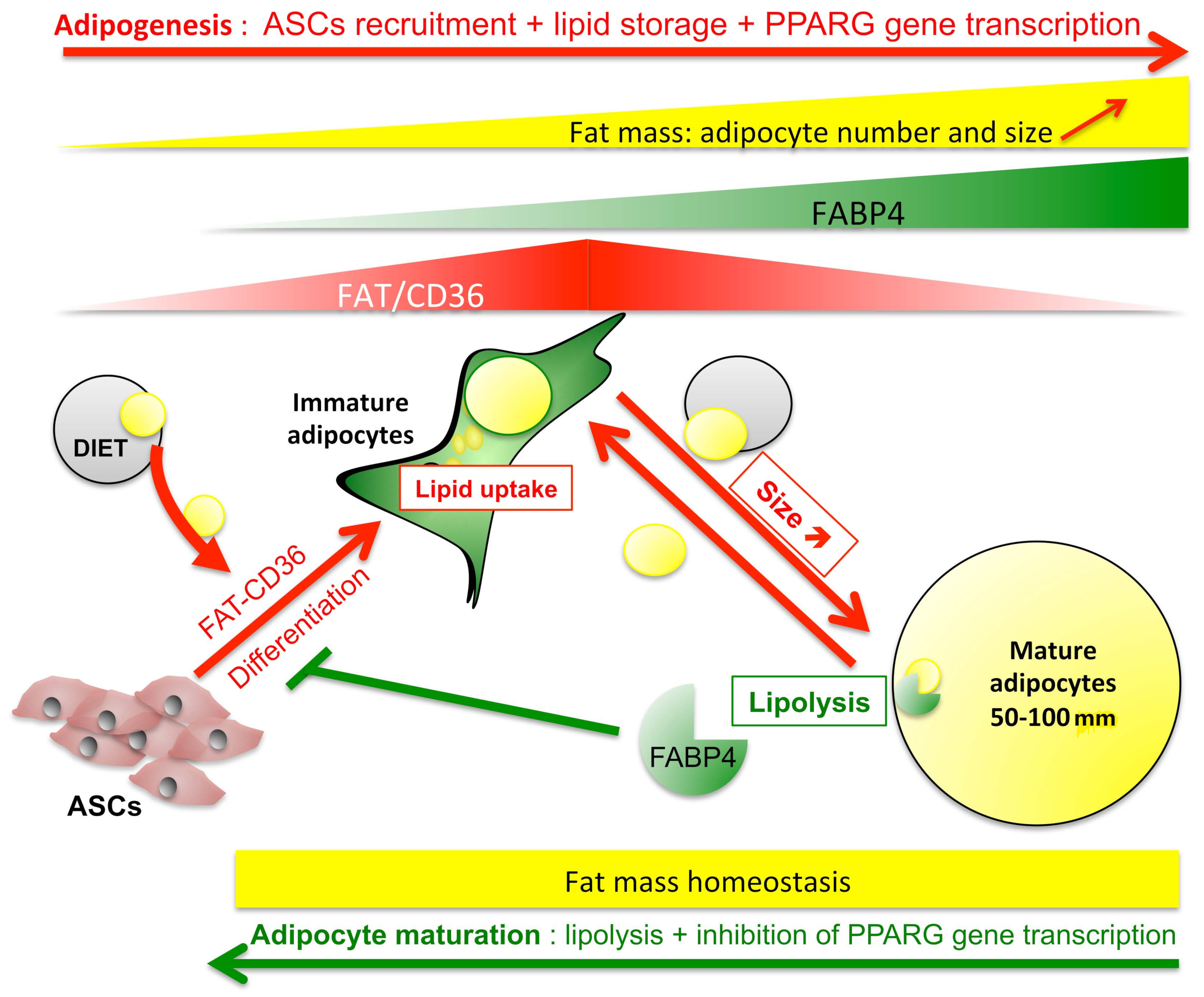
FABP4 Controls Fat Mass Expandability (Adipocyte Size and Number) through Inhibition of CD36/SR-B2 Signalling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. FAT/CD36 Is Involved in Lipid Storage but Reduces Lipogenesis
2.2. Transcriptional Regulations by Fatty Acid in Absence of FAT/CD36: Emerging Role of FABP4
2.3. Intracellular FABP4 Regulates Oleic Acid-Induced PPARG Activity in Adipocytes
2.4. Extracellular FABP4 Inhibits the Recruitment of Adipogenic Precursors by Oleic Acid-Induced FAT/CD36 Signalling
3. Discussion
4. Material & Methods
4.1. Preparation of Explants, Cell Cultures, and Treatments
4.2. Size Determination, Triglyceride, and FAT/CD36 Content Analyses
4.3. Analysis of FABP4 Binding on 3T3-MBX Cells
4.4. Gene Transcription Analyzes
4.5. Bioinformatic Gene Dataset Analyses
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Géloën, A.; Collet, A.J.; Guay, G.; Bukowiecki, L.J. Insulin Stimulates in Vivo Cell Proliferation in White Adipose Tissue. Am. J. Physiol.-Cell Physiol. 1989, 256, C190–C196. [Google Scholar] [CrossRef] [PubMed]
- Walewski, J.L.; Ge, F.; Gagner, M.; Inabnet, W.B.; Pomp, A.; Branch, A.D.; Berk, P.D. Adipocyte Accumulation of Long-Chain Fatty Acids in Obesity Is Multifactorial, Resulting from Increased Fatty Acid Uptake and Decreased Activity of Genes Involved in Fat Utilization. Obes. Surg. 2010, 20, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlamov, O.; Somwar, R.; Cornea, A.; Kievit, P.; Grove, K.L.; Roberts, C.T. Single-Cell Analysis of Insulin-Regulated Fatty Acid Uptake in Adipocytes. Am. J. Physiol.-Endocrinol. Metab. 2010, 299, E486–E496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrinelli, V.; Carobbio, S.; Vidal-Puig, A. Adipose Tissue Plasticity: How Fat Depots Respond Differently to Pathophysiological Cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef] [Green Version]
- Stenkula, K.G.; Erlanson-Albertsson, C. Adipose Cell Size: Importance in Health and Disease. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2018, 315, R284–R295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Samovski, D.; Okunade, A.L.; Stahl, P.D.; Abumrad, N.A.; Su, X. CD36 Level and Trafficking Are Determinants of Lipolysis in Adipocytes. FASEB J. 2012, 26, 4733–4742. [Google Scholar] [CrossRef] [Green Version]
- Azuma, K.; Heilbronn, L.K.; Albu, J.B.; Smith, S.R.; Ravussin, E.; Kelley, D.E. Adipose Tissue Distribution in Relation to Insulin Resistance in Type 2 Diabetes Mellitus. Am. J. Physiol.-Endocrinol. Metab. 2007, 293, E435–E442. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, T.; Lamendola, C.; Coghlan, N.; Liu, T.C.; Lerner, K.; Sherman, A.; Cushman, S.W. Subcutaneous Adipose Cell Size and Distribution: Relationship to Insulin Resistance and Body Fat: Adipose Cell Size and Insulin Resistance. Obesity 2014, 22, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Muir, L.A.; Neeley, C.K.; Meyer, K.A.; Baker, N.A.; Brosius, A.M.; Washabaugh, A.R.; Varban, O.A.; Finks, J.F.; Zamarron, B.F.; Flesher, C.G.; et al. Adipose Tissue Fibrosis, Hypertrophy, and Hyperplasia: Correlations with Diabetes in Human Obesity: Adipose Tissue Fibrosis in Human Obesity. Obesity 2016, 24, 597–605. [Google Scholar] [CrossRef]
- Verboven, K.; Wouters, K.; Gaens, K.; Hansen, D.; Bijnen, M.; Wetzels, S.; Stehouwer, C.D.; Goossens, G.H.; Schalkwijk, C.G.; Blaak, E.E.; et al. Abdominal Subcutaneous and Visceral Adipocyte Size, Lipolysis and Inflammation Relate to Insulin Resistance in Male Obese Humans. Sci. Rep. 2018, 8, 4677. [Google Scholar] [CrossRef]
- Okazaki, H.; Osuga, J.; Tamura, Y.; Yahagi, N.; Tomita, S.; Shionoiri, F.; Iizuka, Y.; Ohashi, K.; Harada, K.; Kimura, S.; et al. Lipolysis in the Absence of Hormone-Sensitive Lipase. Diabetes 2002, 51, 3368–3375. [Google Scholar] [CrossRef] [Green Version]
- Berger, E.; Héraud, S.; Mojallal, A.; Lequeux, C.; Weiss-Gayet, M.; Damour, O.; Géloën, A. Pathways Commonly Dysregulated in Mouse and Human Obese Adipose Tissue: FAT/CD36 Modulates Differentiation and Lipogenesis. Adipocyte 2015, 4, 161–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, J.K.; Vidal-Puig, A.J. Thematic Review Series: Adipocyte Biology. Adipose Tissue Function and Plasticity Orchestrate Nutritional Adaptation. J. Lipid Res. 2007, 48, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. Organismal Carbohydrate and Lipid Homeostasis. Cold Spring Harb. Perspect. Biol. 2012, 4, a006031. [Google Scholar] [CrossRef]
- Roberts, R.; Hodson, L.; Dennis, A.L.; Neville, M.J.; Humphreys, S.M.; Harnden, K.E.; Micklem, K.J.; Frayn, K.N. Markers of de Novo Lipogenesis in Adipose Tissue: Associations with Small Adipocytes and Insulin Sensitivity in Humans. Diabetologia 2009, 52, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Glatz, J.F.C.; Luiken, J.J.F.P. From Fat to FAT (CD36/SR-B2): Understanding the Regulation of Cellular Fatty Acid Uptake. Biochimie 2017, 136, 21–26. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Yang, P.; Chen, Z.; Tang, N.; Ruan, X.Z.; Chen, Y. Genome-Wide Transcriptome Analysis of CD36 Overexpression in HepG2.2.15 Cells to Explore Its Regulatory Role in Metabolism and the Hepatitis B Virus Life Cycle. PLoS ONE 2016, 11, e0164787. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, L.; Laviolette, M.; Rodrigue-Way, A.; Sow, B.; Brochu, M.; Caron, V.; Tremblay, A. The CD36-PPARγ Pathway in Metabolic Disorders. Int. J. Mol. Sci. 2018, 19, 1529. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, J.; Cui, W.; Silverstein, R.L. CD36, a Signaling Receptor and Fatty Acid Transporter That Regulates Immune Cell Metabolism and Fate. J. Exp. Med. 2022, 219, e20211314. [Google Scholar] [CrossRef] [PubMed]
- Géloën, A.; Helin, L.; Geeraert, B.; Malaud, E.; Holvoet, P.; Marguerie, G. CD36 Inhibitors Reduce Postprandial Hypertriglyceridemia and Protect against Diabetic Dyslipidemia and Atherosclerosis. PLoS ONE 2012, 7, e37633. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Graves, R.A.; Budavari, A.I.; Spiegelman, B.M. MPPAR Gamma 2: Tissue-Specific Regulator of an Adipocyte Enhancer. Genes Dev. 1994, 8, 1224–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauma, L.; Franck, N.; Paulsson, J.F.; Westermark, G.T.; Kjølhede, P.; Strålfors, P.; Söderström, M.; Nystrom, F.H. Peroxisome Proliferator Activated Receptor Gamma Activity Is Low in Mature Primary Human Visceral Adipocytes. Diabetologia 2006, 50, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, E.; Géloën, A. Insulin Prevents Fatty Acid Induced Increase of Adipocyte Size. Adipocyte 2022, 11, 510–528. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, V.; Van Hul, M.; Lijnen, H.R.; Scroyen, I. CD36 Promotes Adipocyte Differentiation and Adipogenesis. Biochim. Biophys. Acta BBA—Gen. Subj. 2012, 1820, 949–956. [Google Scholar] [CrossRef]
- Gao, H.; Volat, F.; Sandhow, L.; Galitzky, J.; Nguyen, T.; Esteve, D.; Åström, G.; Mejhert, N.; Ledoux, S.; Thalamas, C.; et al. CD36 Is a Marker of Human Adipocyte Progenitors with Pronounced Adipogenic and Triglyceride Accumulation Potential. Stem Cells 2017, 35, 1799–1814. [Google Scholar] [CrossRef] [Green Version]
- Samovski, D.; Sun, J.; Pietka, T.; Gross, R.W.; Eckel, R.H.; Su, X.; Stahl, P.D.; Abumrad, N.A. Regulation of AMPK Activation by CD36 Links Fatty Acid Uptake to β-Oxidation. Diabetes 2015, 64, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Gaidhu, M.P.; Fediuc, S.; Ceddia, R.B. 5-Aminoimidazole-4-Carboxamide-1-β-D-Ribofuranoside-Induced AMP-Activated Protein Kinase Phosphorylation Inhibits Basal and Insulin-Stimulated Glucose Uptake, Lipid Synthesis, and Fatty Acid Oxidation in Isolated Rat Adipocytes. J. Biol. Chem. 2006, 281, 25956–25964. [Google Scholar] [CrossRef] [Green Version]
- Hebbachi, A.; Saggerson, D. Acute Regulation of 5′-AMP-Activated Protein Kinase by Long-Chain Fatty Acid, Glucose and Insulin in Rat Primary Adipocytes. Biosci. Rep. 2013, 33, e00007. [Google Scholar] [CrossRef]
- Kim, S.-J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase To Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol. Cell. Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Vroegrijk, I.O.C.M.; van Klinken, J.B.; van Diepen, J.A.; van den Berg, S.A.A.; Febbraio, M.; Steinbusch, L.K.M.; Glatz, J.F.C.; Havekes, L.M.; Voshol, P.J.; Rensen, P.C.N.; et al. CD36 Is Important for Adipocyte Recruitment and Affects Lipolysis: CD36 Affects Adipocyte Lipolysis. Obesity 2013, 21, 2037–2045. [Google Scholar] [CrossRef]
- Coe, N.R.; Simpson, M.A.; Bernlohr, D.A. Targeted Disruption of the Adipocyte Lipid-Binding Protein (AP2 Protein) Gene Impairs Fat Cell Lipolysis and Increases Cellular Fatty Acid Levels. J. Lipid Res. 1999, 40, 967–972. [Google Scholar] [CrossRef]
- Hofer, P.; Boeszoermenyi, A.; Jaeger, D.; Feiler, U.; Arthanari, H.; Mayer, N.; Zehender, F.; Rechberger, G.; Oberer, M.; Zimmermann, R.; et al. Fatty Acid-Binding Proteins Interact with Comparative Gene Identification-58 Linking Lipolysis with Lipid Ligand Shuttling. J. Biol. Chem. 2015, 290, 18438–18453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyamfi, J.; Yeo, J.H.; Kwon, D.; Min, B.S.; Cha, Y.J.; Koo, J.S.; Jeong, J.; Lee, J.; Choi, J. Interaction between CD36 and FABP4 Modulates Adipocyte-Induced Fatty Acid Import and Metabolism in Breast Cancer. Npj Breast Cancer 2021, 7, 129. [Google Scholar] [CrossRef]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 Attenuates PPARγ and Adipogenesis and Is Inversely Correlated With PPARγ in Adipose Tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soula, H.A.; Julienne, H.; Soulage, C.O.; Géloën, A. Modelling Adipocytes Size Distribution. J. Theor. Biol. 2013, 332, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.Z.; Richard, G.; Gévry, N.; Tchernof, A.; Carpentier, A.C. Fat Cell Size: Measurement Methods, Pathophysiological Origins, and Relationships with Metabolic Dysregulations. Endocr. Rev. 2022, 43, 35–60. [Google Scholar] [CrossRef]
- Fang, L.; Guo, F.; Zhou, L.; Stahl, R.; Grams, J. The Cell Size and Distribution of Adipocytes from Subcutaneous and Visceral Fat Is Associated with Type 2 Diabetes Mellitus in Humans. Adipocyte 2015, 4, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Hames, K.C.; Vella, A.; Kemp, B.J.; Jensen, M.D. Free Fatty Acid Uptake in Humans with CD36 Deficiency. Diabetes 2014, 63, 3606–3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daquinag, A.C.; Gao, Z.; Fussell, C.; Immaraj, L.; Pasqualini, R.; Arap, W.; Akimzhanov, A.M.; Febbraio, M.; Kolonin, M.G. Fatty Acid Mobilization from Adipose Tissue Is Mediated by CD36 Posttranslational Modifications and Intracellular Trafficking. JCI Insight 2021, 6, e147057. [Google Scholar] [CrossRef]
- Gumbilai, V.; Ebihara, K.; Aizawa-Abe, M.; Ebihara, C.; Zhao, M.; Yamamoto, Y.; Mashimo, T.; Hosoda, K.; Serikawa, T.; Nakao, K. Fat Mass Reduction With Adipocyte Hypertrophy and Insulin Resistance in Heterozygous PPARγ Mutant Rats. Diabetes 2016, 65, 2954–2965. [Google Scholar] [CrossRef]
- López, I.P.; Marti, A.; Milagro, F.I.; Zulet, M.D.L.A.; Moreno-Aliaga, M.J.; Martinez, J.A.; De Miguel, C. DNA Microarray Analysis of Genes Differentially Expressed in Diet-Induced (Cafeteria) Obese Rats. Obes. Res. 2003, 11, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Takasawa, K.; Kubota, N.; Terauchi, Y.; Kadowaki, T. Impact of Increased PPARgamma Activity in Adipocytes in Vivo on Adiposity, Insulin Sensitivity and the Effects of Rosiglitazone Treatment. Endocr. J. 2008, 55, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S.; Bernlohr, D.A. Metabolic Functions of FABPs—Mechanisms and Therapeutic Implications. Nat. Rev. Endocrinol. 2015, 11, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Trojnar, M.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Leszczyńska-Gorzelak, B.; Mosiewicz, J. Associations between Fatty Acid-Binding Protein 4–A Proinflammatory Adipokine and Insulin Resistance, Gestational and Type 2 Diabetes Mellitus. Cells 2019, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Micaelo, N.; Rodríguez-Calvo, R.; Guaita-Esteruelas, S.; Heras, M.; Girona, J.; Masana, L. Extracellular FABP4 Uptake by Endothelial Cells Is Dependent on Cytokeratin 1 Expression. Biochim. Biophys. Acta BBA—Mol. Cell Biol. Lipids 2019, 1864, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Ertunc, M.E.; Sikkeland, J.; Fenaroli, F.; Griffiths, G.; Daniels, M.P.; Cao, H.; Saatcioglu, F.; Hotamisligil, G.S. Secretion of Fatty Acid Binding Protein AP2 from Adipocytes through a Nonclassical Pathway in Response to Adipocyte Lipase Activity. J. Lipid Res. 2015, 56, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.J.; Sridhar, K.; Bernlohr, D.A.; Kraemer, F.B. Interaction of Rat Hormone-Sensitive Lipase with Adipocyte Lipid-Binding Protein. Proc. Natl. Acad. Sci. USA 1999, 96, 5528–5532. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.R.; Mazurkiewicz-Muñoz, A.M.; Suttles, J.; Carter-Su, C.; Bernlohr, D.A. Interaction of Adipocyte Fatty Acid-Binding Protein (AFABP) and JAK2. J. Biol. Chem. 2009, 284, 13473–13480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amri, E.; Bertrand, B.; Ailhaud, G.; Grimaldi, P. Regulation of Adipose Cell Differentiation. I. Fatty Acids Are Inducers of the AP2 Gene Expression. J. Lipid Res. 1991, 32, 1449–1456. [Google Scholar] [CrossRef]
- Yamamoto, T.; Furuhashi, M.; Sugaya, T.; Oikawa, T.; Matsumoto, M.; Funahashi, Y.; Matsukawa, Y.; Gotoh, M.; Miura, T. Transcriptome and Metabolome Analyses in Exogenous FABP4- and FABP5-Treated Adipose-Derived Stem Cells. PLoS ONE 2016, 11, e0167825. [Google Scholar] [CrossRef]
- Dou, H.-X.; Wang, T.; Su, H.-X.; Gao, D.-D.; Xu, Y.-C.; Li, Y.-X.; Wang, H.-Y. Exogenous FABP4 Interferes with Differentiation, Promotes Lipolysis and Inflammation in Adipocytes. Endocrine 2020, 67, 587–596. [Google Scholar] [CrossRef]
- Berger, E.; Rome, S.; Vega, N.; Ciancia, C.; Vidal, H. Transcriptome Profiling in Response to Adiponectin in Human Cancer-Derived Cells. Physiol. Genomics 2010, 42A, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Berger, E.; Vega, N.; Weiss-Gayet, M.; Géloën, A. Gene Network Analysis of Glucose Linked Signaling Pathways and Their Role in Human Hepatocellular Carcinoma Cell Growth and Survival in HuH7 and HepG2 Cell Lines. BioMed Res. Int. 2015, 2015, 821761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabaouni, I.; Vannier, B.; Moussa, A.; Ibrahimi, A. Microarray integrated analysis of a gene network for the CD36 myocardial phenotype. Bioinformation 2016, 12, 332–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, D.; Araki, H.; Tamada, Y.; Dunmore, B.; Sanders, D.; Humphreys, S.; Affara, M.; Imoto, S.; Yasuda, K.; Tomiyasu, Y.; et al. Gene Network Inference and Visualization Tools for Biologists: Application to New Human Transcriptome Datasets. Nucleic Acids Res. 2012, 40, 2377–2398. [Google Scholar] [CrossRef]
- Yang, R.; Castriota, G.; Chen, Y.; Cleary, M.A.; Ellsworth, K.; Shin, M.K.; Tran, J.-L.; Vogt, T.F.; Wu, M.; Xu, S.; et al. RNAi-Mediated Germline Knockdown of FABP4 Increases Body Weight but Does Not Improve the Deranged Nutrient Metabolism of Diet-Induced Obese Mice. Int. J. Obes. 2011, 35, 217–225. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, E.; Géloën, A. FABP4 Controls Fat Mass Expandability (Adipocyte Size and Number) through Inhibition of CD36/SR-B2 Signalling. Int. J. Mol. Sci. 2023, 24, 1032. https://doi.org/10.3390/ijms24021032
Berger E, Géloën A. FABP4 Controls Fat Mass Expandability (Adipocyte Size and Number) through Inhibition of CD36/SR-B2 Signalling. International Journal of Molecular Sciences. 2023; 24(2):1032. https://doi.org/10.3390/ijms24021032
Chicago/Turabian StyleBerger, Emmanuelle, and Alain Géloën. 2023. "FABP4 Controls Fat Mass Expandability (Adipocyte Size and Number) through Inhibition of CD36/SR-B2 Signalling" International Journal of Molecular Sciences 24, no. 2: 1032. https://doi.org/10.3390/ijms24021032