Abstract
It is widely accepted that oxidative stress (OS) coming from a wide variety of causes has detrimental effects on male fertility. Antioxidants could have a significant role in the treatment of male infertility, and the current systematic review on the role of melatonin to ameliorate OS clearly shows that improvement of semen parameters follows melatonin supplementation. Although melatonin has considerable promise, further studies are needed to clarify its ability to preserve or restore semen quality under stress conditions in varied species. The present review examines the actions of melatonin via receptor subtypes and its function in the context of OS across male vertebrates.
1. Introduction
The term stress means both systemic and local stress that is created after a disturbance of homeostasis. Usually, a specific stressor causes a specific type of local stress; meanwhile, when the intensity of stress exceeds a threshold, the hypothalamic–pituitary–adrenal axis is activated, resulting in the systematic stress response [1,2]. Additionally, as the concept of homeostasis becomes more specific, so does the concept of stress [3]. As an example, oxidative stress (OS) refers exclusively to the disruption of redox signaling and control [4,5].
Our common knowledge on oxidative stress has evolved substantially over the years, being focused mostly on the fundamental chemical reactions and the most relevant chemical species [6]. Thus, reactive oxygen species (ROS) and reactive nitrogen species (RNS) were identified as key players in initiating, mediating, and regulating the cellular and biochemical complexity of oxidative stress either as physiological or as pathogenic processes [6]. RS (reactive species) is defined as the disruption of redox signaling and control caused by RONS (both ROS and RNS), while the OS response is defined as the efforts made to restore homeostasis. This response ultimately leads to specific biological consequences that constitute the effects of this stress [3].
RONS are characterized as reactive molecules and can damage various biomolecules such as proteins, lipids, and nucleic acids [7]. The first studies conducted on the effect of RONS showed that they cause the pathogenesis of various diseases and the aging process [8]. Although excessive production of RONS can cause damage, when present in certain concentrations, they are essential for the regulation of life processes [7,9,10,11]. Consequently, many researchers conclude that an optimal level of stress is vital for maintaining health, as its absence can impair growth body composition, and development as well as lead to pathological conditions [1].
Male infertility has been linked to various types of stress as well as stressful life events [12,13,14,15]. Several studies, for example, have linked male (sub/in) fertility to stressful lifestyles [12,16,17,18]. Because mitochondria are the major energy source for sperm, they have been suggested as an important link between sperm motility and their response to stress [1,17,19,20]. Additionally, stress hormone receptors activate specific signaling pathways each time that are particularly important for both mitochondrial network homeostasis and sperm functionality [14,15,21]. Therefore, mitochondria are a particularly important organelle that contributes to sperm functionality and fertility [17].
Many strategies (using different sources of antioxidants) have been suggested to prevent the generation of oxidative agents. Among them, the use of melatonin (N-acetyl-5-methoxytryptamine), the principal hormone secreted by the pineal gland, has been suggested as a free radical scavenger and antioxidant [22]. Mainly due to its amphiphilic nature that allows it to pass through all morphophysiological barriers of the cell, it is one of the most effective antioxidants protecting cells from OS caused by reactive species [23]. In addition, its lipophilic nature allows it to easily cross cell membranes and act directly in various organs including those of the reproductive system [24,25]. Of particular interest is the fact that melatonin’s metabolites, which are formed when the hormone functions as a scavenger, are likewise equally as good or better than the parent molecule in neutralizing toxic oxygen-based and nitrogen-based reactants [26].
In this review, we describe the adverse influence of different stressful stimuli (e.g., heat stress, assisted reproductive technologies, pollution, and cryopreservation) on semen quality and its relation to mitochondrial function in the semen of various animals, from humans and fishes. Furthermore, the role of melatonin in ameliorating OS in sperm cells and future research directions are discussed.
2. Sources of RONS in Semen
There is a plethora of sources that produce reactive oxygen species (RONS) in semen. Among them are different qualities of sperm (immature, dead, and dysfunctional), organelles such as mitochondria, and factors associated with lifestyles and environmental agents.
Leukocytes: Leukocytes in small quantities are a normal finding in ejaculation and often pass through the male genital tract. The World Health Organization defines leukocytospermia as peroxidase-positive leukocyte concentration >1 × 106 per mL of semen, and it occurs in about 10–20% of infertile men [27,28]. Polymorphonuclear neutrophils are the most common type of seminal leukocytes and the primary endogenous source of RONS in semen [29]. The activation of these leukocytes during genitourinary or accessory gland infections provokes an important increase in oxygen consumption and RONS generation [30,31].
Immature sperm: When there is a lot of cytoplasmic retention along the midpiece of the spermatozoa, there is a risk of functionally defective or immature sperm. These cytoplasmic residues contain an increased number of enzymes, particularly glucose-6-phosphate dehydrogenase (G6PDH) and NADPH oxidase [32]. The excessive presence of G6PD results in the generation of a significant amount of NADPH, which represents a primary source of electrons that are necessary for the reduction in molecular oxygen to O2•− [31]. Several studies have suggested a correlation between cytoplasmic droplet retention and severe peroxidation damage to the sperm membrane. Creatinine kinase (CK) and G6PDH were used as biomarkers of this retention [33,34,35,36]. In addition, it has been reported that in an environment of increased RONS levels, immature sperm can cause OS in neighboring mature ones during their transit through the epididymis [37,38,39]. Also, mature sperm could be an additional source of RONS due to cellular metabolism [40].
Lifestyles and environmental factors: A list of factors associated with lifestyles and environmental agents could contribute to the severe generation of RONS in sperm. Cigarette smoking, alcohol consumption, electromagnetic radiation, obesity, microorganism mutations, or sexually transmitted diseases are actively involved [28,41,42,43].
Mitochondrial metabolism: Mitochondria are an important source of RONS, but at the same time, they have a system to control the levels of RONS to ensure that they do not exceed some critical limits and do not cause cell damage or apoptosis [44]. Superoxide dismutase is one of the most studied enzymes involved in mitochondrial RONS scavenging. Although excessive production of RONS is associated with OS, their presence at specific levels aids many cellular processes, while their lack in developmental and somatic cells can lead to impairments in cell differentiation, immune response, and autophagy [45,46,47].
Dysfunctional sperm: This kind of sperm presents excess residual cytoplasm, retention of histones, and poor protamination, resulting from differentiation failure during spermiogenesis [48]. There is a positive correlation between RONS production, anomalies in sperm morphology, and DNA damage [31,38,49]. High levels of expression of the isoform of NADPH oxidase (NOX)—NOX5, generators of RONS in sperm—have been reported in both teratozoospermia [50] and asthenozoospermic human ejaculates [37].
Dead sperm: Increased levels of dead spermatozoa in ejaculation are alarming during techniques of assisted reproductive technologies (ARTs) related to sperm, especially during cryopreservation. Dead sperm are high RONS producers [51], and their contribution to RONS generation varies according to the species. Dead sperm produce RONS through an aromatic L-amino acid oxidase (LAAO) pathway in bull semen [52]. In male species such as bulls, rams, and stallions, L-amino acid oxidase is increased following sperm death [29,52,53,54,55].
3. Effects of RONS on Sperm
Redox balance is substantial for maintaining different crucial factors of semen functionality [56]. On the other hand, an imbalance of redox status negatively affects semen quality due to oxidative damage [32]. Prolonged exposure and/or increased RONS concentrations cause extensive injury to miscellaneous biomolecules, such as proteins, lipids, and nucleic acids, which in the end hinder multiple cellular functions [30,32,40,57]. The adverse events include mitochondrial dysfunction, impaired sperm motility, loss of membrane integrity, DNA damage, and apoptosis [40,58]. A significant increase in RONS levels has been detected in 25–40% of semen samples of men who are infertile [59,60]. The severity of oxidative injury to sperm among infertile men depends on the length of exposure, concentrations, properties of reactive molecules, and antioxidant efficiency as well as surrounding temperature and oxygen tension [56]. In farm animals, the impact of OS is associated with reduced sperm quality and fertility [61,62]. Recently, Wyck et al. [63] found that bull sperm exposed to OS showed increased DNA damage, which was associated with impaired DNA demethylation during epigenetic reprogramming in early embryonic development. Ribas-Maynou et al. [64] found that the impact of OS at the sperm level translates into poorer fertilization and blastocyst rates after ARTs, such as in vitro fertilization (IVF) or intracytoplasmic sperm injection (ICSI).
The sperm plasma membrane consists of large amounts of polyunsaturated fatty acids (PUFAs), especially docosahexaenoic acids, which improve its fluidity that is necessary for multiple membrane fusion events [56]. At the same time, however, these fatty acids make sperm more susceptible to oxidative damage as they act as potential peroxidation substrates because they contain methylene groups with highly reactive hydrogen atoms [65]. There are marked differences among species in the composition of a sperm plasma membrane PUFAs. The most abundant PUFAs is either DHA (ω-3 PUFAs) or DPA (ω-6 PUFAs). From a broad perspective, differences in the sperm plasma membrane lipid composition among species explain differences in the response to semen processing (e.g., during cooling and cryopreservation) [66]. These differences may be attributable to variations in plasma membrane compositions [67], for instance, the content of lipids in the bilayer, degree of hydrocarbon chain saturation, cholesterol/phospholipids ratio, and protein/phospholipid ratio [68].
In addition, studies have shown that the excessive presence of RONS, especially nitric oxide (NO•), O2•−, and OH•, leads to the attachment of the above molecules to sperm DNA, causing excessive modifications and deletions in nucleotide bases and strand breakages, along with other multiple genotoxic effects. Sperm DNA, due to poor chromatin compaction and incomplete protamination, is particularly susceptible to oxidative damage [43,69]. Damage to sperm DNA due to RONS was experimentally demonstrated by the positive correlation between sperm ROS levels and sperm DNA fragmentation in humans [70,71].
RONS have also been considered one of the main apoptotic stimuli triggering mitochondria to produce specific signaling molecules crucial for the activation of programmed cell death [72]. More specifically, mature sperm from patients with elevated RONS levels showed intense apoptotic activity compared to the control group [73]. Reduction in this activity due to OS can be observed after antioxidant therapy with cinnamon bark oil consumption in asthenozoospermic men [74].
Several studies have correlated OS markers with fertility [75,76,77] and important sperm quality parameters such as morphology [78,79] and motility [80,81] in humans. Rapid progressive sperm motility is a highly sensitive indicator of lipid peroxidation and can be detected before the deterioration of other sperm kinematic parameters [56]. Additionally, the negative correlation between intracellular sperm O2•− levels and average-path velocity (VAP) is important for understanding the potential role of free radicals in limiting the actual rate of sperm forward movement within the female reproductive tract, which is vital for successful fertilization [82]. Likewise, the positive correlation between superoxide dismutase (SOD) activity, curvilinear velocity (VCL), and amplitude of lateral head displacement (ALH) led to a quantification of the role of OS in the development of spontaneous and premature hyperactivated sperm motility during ejaculation [82]. Although a clear link between oxidative stress and male fertility disorders has been demonstrated in humans and laboratory animals, less information is available about the implications of this condition in the male fertility of domestic animals [83,84].
4. The Role of Melatonin
The major pathway of melatonin biosynthesis consists, in any organism or cell type tested, of tryptophan 5-hydroxylation, decarboxylation, N-acetylation, and O-methylation, in this order. Alternately, but at lower flux rates, melatonin can be formed via O-methylation of serotonin and subsequent N-acetylation of 5-methoxytryptamine or by O-methylation of tryptophan followed by decarboxylation and N-acetylation [85].
Melatonin has an impact working as either an antioxidant [86] or as a hormone due to its amphiphilic properties. Melatonin was discovered as a secretory product of the pineal gland and is now known to be generated in many organs in the body [87,88]. The hydrophilic and lipophilic properties of melatonin permit it to rapidly pass through all morphophysiological barriers easily and disperse in organs and fluids [89,90]. The important advantage of melatonin among other antioxidants is that in addition to melatonin itself, its metabolic derivatives formed during antioxidant reactions, including cyclic 3-hydroxymelatonin (3-OHM), N1-acetyl-N2-formyl-5-methoxykynuramine (AFMK), and N1-acetyl-5-methoxykynuramine (AMK), are all excellent free radical scavengers that contribute to the reduction in OS [26,88,91].
Melatonin involves multiple mechanisms to control cellular physiology, including via membrane receptors, nuclear binding sites, and interaction with cytosolic molecules [92]. Melatonin exerts most of its major physiological actions by reacting with the two membrane receptors MT1 and MT2, which belong to the G protein-coupled receptor superfamily [92,93,94].
The receptors of melatonin are expressed in several central nervous tissues and numerous peripheral tissues, including the testis and ovary [95]. Specifically, melatonin is involved in the modulation of the hypothalamic–pituitary–gonadal (HPG) axis, which is a very important regulatory center for animal reproduction, both in seasonal breeding animals and in non-seasonally breeding animals, including humans [96]. Some investigations have revealed that the MT1 receptor is widely distributed in endocrine tissues and brain regions, which are the main response sites of melatonin-induced physiological and circadian effects [97]. However, the MT2 receptor is generally absent in the mammalian hypothalamus and pituitary gland [97]. It is obvious that MT1 receptor is the more important receptor for melatonin-modulated reproductive regulation in mammals.
To date, the use of melatonin in human physiology has been restricted to the improvement of sleep quality, the alleviation of feelings of jet lag, and the reduction in sleep onset latency [98]. However, numerous studies have concluded that melatonin could also be associated with the prevention of several diseases related to aging and oxidative stress, including type 2 diabetes, cardiovascular and neurodegenerative diseases, or cancer [99,100,101,102,103,104,105,106]. Simple and composite formulations of synthetic melatonin come in a wide range of forms such as tablets, pills, sublingual drops, liquids, gels, creams, and even suppositories, as well as at dosages from 0.1 mg to 400 mg. In the case of composite formulations, synthetic melatonin is presented together with other compounds such as tryptophan, vitamins (C and B6), minerals, and even collagen and hyaluronic acid in the case of creams [98].
4.1. Melatonin Receptors
In vertebrates, there are two melatonin receptοrs: MT1 (or Mel1a, gene MTNR1A) and MT2 (or Mel1b, gene MTNR1B) [107]. The melatonin receptor family belongs to the class A rhodopsin-like G protein-coupled receptor (GPCR) subfamily [94,95]. The MT1 receptor is composed of 350 amino acids, while the MT2 receptor is composed of 362 amino acids, showing 55% and 70% homology in the transmembrane domain [108]. Interestingly, ruminants such as cows, sheep, and camels have MT1 and MT2 receptors that are about 10 amino acids longer than those in humans [109], although there is no report of significant functional differences among species. The two receptors have 60% homology among mammalian species, including humans, rats, mice, and ruminants [110,111]. An additional member, MT3, also termed Mel1c, has been identified but only in nonmammalian species, such as birds, amphibians, and fish [96,112].
4.2. Melatonin Subtype Receptors
Melatonin’s effects on reproduction are likely acting as a hormone that binds subtype receptors [86]. Differences in regulatory functions of melatonin-binding proteins have procured categories of melatonin receptor subtypes [113]. Melatonin subtype receptors include membrane G protein-coupled receptors and nuclear orphan receptors [113]. There is scientific evidence that nuclear orphan receptors might be primary targets downstream in the membrane receptor signaling pathway [114]. Also, many studies have proven that melatonin’s functions via membrane receptors regulate metabolic, cardiovascular, immune, and reproductive systems [115].
4.3. Melatonin-Receptor-Mediated Biological Effects
Melatonin receptors play a significant role in miscellaneous physiological activities such as the regulation of circadian rhythm, stem-cell survival, maturation, differentiation, antioxidation, apoptosis, and cancer [116]. Melatonin receptors are also involved in antioxidant and antiapoptotic effects [117]. The biological effects that are mediated with melatonin receptors are represented schematically in Figure 1.
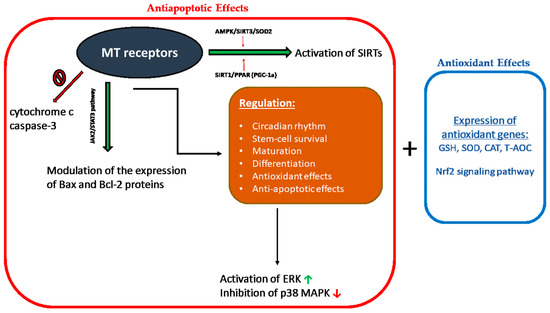
Figure 1.
Illustration of the biological effects that are mediated with melatonin receptors. Red frame (antiapoptotic effects) and orange frame (antioxidant effects). MT: melatonin; SIRTs: certain histone deacetylases; Nrf-2: nuclear factor erythroid 2-related factor; GSH: glutathione peroxidase; SOD: superoxide dismutase; CAT: catalase; T-AOC: total antioxidant capacity; ERK: extracellular signal-regulated kinase; and p38 MAPK: mitogen-activated protein kinases.
4.3.1. Antioxidant Effects
The effects of melatonin and its metabolites on RONS levels and mitochondrial protection are expected to be linked to the activation of basic signaling pathways such as stress-activated/mitogen-activated protein kinases (SAP/MAPKs). These protein kinases are mediators of signal transmission from the cell surface to the nucleus and practically control most of the physiological activities in eukaryotic cells [118]. Nuclear factor erythroid 2-related factor 2 (Nrf-2) and heme oxygenase-1 1 (HO-1) regulate the antioxidant potential of cells by regulating the transcription and translation of various antioxidant enzymes [119].
Melatonin protects the cell from OS by inducing the expression of antioxidant genes such as glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), catalase (CAT) and by increasing the total antioxidant capacity (T-AOC) in the presence of a stimulation of respiratory complexes activity [120]. These effects are mediated by the stimulation of transcriptional proteins, such as nuclear factor erythroid 2 (Nrf2), the activity of which is particularly important for the cryoprotection and stress adaptation effects of melatonin [121].
4.3.2. Antiapoptotic Effects
Melatonin modulates the expression of the proteins Bax and Bcl-2 and inhibits the release of cytochrome c and the activation of caspase-3. Melatonin-induced Bax/Bcl-2 translocation occurs via the JAK2/STAT3 pathway, and its antiapoptotic effect is mediated via ERK activation and p38 MAPK inhibition [122,123,124]. In addition, melatonin-activated signaling cascades also include activation of certain histone deacetylases (SIRTs) through AMPK/SIRT3/SOD2 and SIRT1/PPAR- coactivator (PGC-1α) [125,126]. The regulation of transcription factor PGC-1α is MT1-receptor-dependent [127]. The blockage of melatonin receptors significantly promotes hCG-induced apoptosis in mice [109].
4.4. Melatonin Effects on Testicular Physiology
Sperm motility in humans seems to be unaffected by melatonin [128], inhibited by high concentrations of melatonin in Wistar rats [129]. These varying effects of melatonin on sperm motility might be due to the time of day the tissue was harvested, photoperiodicity of these mammals, or even melatonin-receptor expression in sperm [113]. Melatonin implants in Rasa Aragonesa rams during the non-reproductive season (spring) increased seminal plasma testosterone (T) concentrations after four weeks and 17α-estradiol (E2) after eight weeks [130]. From this, we can predict that for seasonally breeding males that respond to melatonin to time their breeding, melatonin administration can override other endogenous circannual rhythms to activate the mammalian reproductive axis [113]. Melatonin administration for fifteen weeks during the breeding season compared to the non-breeding season significantly increased plasma T concentrations in both winter and spring seasons in Chios rams, and this increase was higher in winter [131]. In Syrian hamsters (long-day breeders), testes collected at different reproductive states (photostimulated and photorefractory) showed that melatonin concentration is significantly higher in the testes of hamsters kept on short days (photorefractory) relative to their long-day (photostimulated) counterparts [132].
In addition to the implication of seasonal changes in the synthesis of melatonin in testes, Mukherjee and Haldar [132] also found that the MT1 receptor was significantly higher in testes collected from the short-day group. Across a variety of breeding types as long-day, short-day, and non-seasonal breeders [133], MT1 and MT2 were expressed in the sperm of these types in a receptor-mediated manner affecting semen [113]. In Table 1, the effects of melatonin supplementation in different media for different species on sperm cells are summarized.

Table 1.
Effects of melatonin supplementation on sperm quality under heat stress conditions.
4.5. Melatonin and Sperm Quality
Melatonin is now considered a potential improver of sperm motility [107]. In in vivo experiments performed on mammals such as rams [140], bucks [141], and buffalos [142], subcutaneous implantation of melatonin resulted in better sperm quality and improved both total and rapid motility as well as sperm viability. In vitro experiments with melatonin showed a significant reduction in sperm deformations and an improvement in its stability, viability, and fertilizing ability, including non-sorted and sex-sorted sperm [143,144].
Despite the positive effects of melatonin due to its antioxidant properties and its receptor-mediated signaling transduction, its mechanism of action is still not fully clear [107]. This hormone can scavenge high levels of RONS in sperm, thereby reducing DNA damage and sperm apoptosis. It can also remove nitrogen-based reactants and toxic oxygen but also increase the activities of antioxidant enzymes, such as catalase (CAT), glutathione peroxidase (GPx), and superoxide dismutase (SOD) [107].
Melatonin has improved the mobility and velocity parameters of sperm because of its interaction with calmodulin [145], which influences the cytoskeletal elements of the sperm. Melatonin also stimulates the cellular influx of Ca2+ into sperm cells enhancing their motility [146]. Calcium and its channels control the transition of flagella wave forms from symmetric to asymmetric [147]. Calmodulin acts as an intracellular regulator of Ca2+ function and has been identified both in the head and flagellar parts of spermatozoa [148], and it is believed to be a major signal transducer of Ca2+ in regulating motility. Additionally, it modulates the levels of the second messenger cAMP [149], which acts both via the axoneme of the sperm tail [150] and via cell-membrane-dependent pathways [151], thereby improving sperm motility and velocity. The negative effects of different types of stress and the positive effects of melatonin (MEL) on sperm are illustrated in Figure 2.
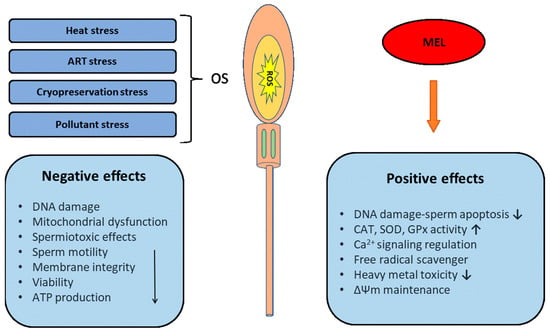
Figure 2.
Illustration of the negative effects of different types of stress and the positive effects of melatonin (MEL) on sperm. ROS: reactive oxygen species; SOD: superoxide dismutase; CAT: catalase; GPX: glutathioneperoxidase; and ΔΨm: mitochondrial membrane potential.
5. Melatonin Alleviates Heat-Stress-Induced OS
5.1. The Problem of Heat Stress Conditions
In the case of heat stress (HS), an increase in RONS levels is caused due to the initiation of OS [152]. Sperm functionality is negatively affected by OS due to the reduction in motility parameters, DNA integrity, and mitochondrial dysfunction [152]. Characteristically, HS induces dysfunctions in mitochondria involving reduced activities of the mitochondrial respiratory chain (complex I and IV) and sperm membrane potential, leading to reduced ATP content [152].
Gong et al. [153] reported that under HS conditions, sperm mitochondrial function is affected mainly due to interferences in mitochondrial remodeling and mitochondrial protein transport. Additionally, HS affects the induction of glycogen synthase kinase 3 (GSK3) activity, which modulates the permeability of the exterior membrane of mitochondria, leading to decreases in the mitochondrial ability to release ATP [153]. Furthermore, increased mitochondrial ROS production has been linked to decreased sperm motility and loss of mitochondrial membrane potential (MMP) [154]. Specifically, Rahman et al. [155] induced heat stress in cryopreserved bovine semen by exposing it to 41.0 °C and observed a decrease in plasma membrane integrity and mitochondrial potential. Finally, the abnormal function of sperm mitochondria due to heat stress reduces its functionality, leading to male infertility [156].
5.2. Melatonin’s Anti-Heat Stress Action
To enhance and improve sperm mitochondrial function due to adverse environmental stress such as HS, several antioxidants have been proposed and used [77,139,157,158] In addition, an extensive study of the negative effects of HS on sperm mitochondrial function could help develop new techniques to mitigate them using mitochondrial enhancers [157].
Zhao et al. [134] reported that 1000 μM of melatonin acts against HS-induced (the sperm aliquot was incubated in an incubator at 42 °C for 3 h in a humidified air atmosphere with 5% of CO2) oxidative damage in human sperm and specifically improves sperm motility, reduces mitochondrial RONS production, stabilizes mitochondrial membrane potential balance, reduces lipid peroxidation, helps maintain DNA integrity, and reduces apoptosis. Also, exposure of human sperm suspensions to 1000 µM of melatonin for 30 min led to an improvement in motility and a decrease in the number of immobile spermatozoa [135]. Accordingly, treatment with 2 mM of melatonin for 120 min in human sperm improved motility and reduced NO and ROS generation [136].
Recently, Qin et al. [137] reported that melatonin can effectively confront different endoplasmic reticulum stress (ERS) signaling pathways and reduce mouse spermatocyte apoptosis [159]. The ERS is a critically crucial factor in many pathologies, and several studies have linked ERS to heat stress [137]. Melatonin actions on the mouse spermatocytes during high temperature include the inhibition of signaling pathways Atf6 and Perk, down-regulation of the expression of Chop and Caspase12, and reduction in apoptosis [137]. The role of melatonin in the mouse spermatocyte (cell line GC2) was dependent on both MT1 and MT2 receptors [137]. In contrast, melatonin activity in mammals was mediated through its interactions with the G protein-coupled MT1 and MT2 receptors [115].
Guo et al. [138] confirmed experimentally in vivo that exposure to HS causes loss of mouse germ cells (CD-1) and disruption of testicular histomorphology. Apoptotic germ cells were experimentally cleared, resulting in a reduction in all stages of these cells in the seminiferous tubules, leading to hollow seminiferous tubules and decreased testicular weight. Subsequently, the seminiferous tubules gradually filled with regenerated germ cells, and testicular weight was restored. The use of melatonin in the above experiment failed to prevent testicular damage due to HS but significantly accelerated testicular recovery [138]. The above work is the first report that showed that the prolonged use of melatonin accelerates the repair of heat-stress-induced testicular damage in mice. This action of melatonin promotes the clearance of apoptotic germ cells through RAC1-mediated phagocytosis via Sertoli cells and supports the regeneration of germ cells by restoring tight and gap junctions. Additionally, melatonin improves the phagocytotic capacity of Sertoli cells by upregulating the Cx43 gap junction protein Connexin43 [138].
In 2022, Shahat et al. [139] induced mild HS (scrotal neck isolation) in rams and studied whether melatonin use affected testicular blood flow and sperm quality. It was experimentally shown that from 2 to 7 weeks after stress induction, the group receiving melatonin (subcutaneously) showed higher and better progressive and total motility. Also, compared to the control group, it showed fewer sperm abnormalities and higher acrosome integrity. The above work is the first report of the positive effects of melatonin in testicular protection against HS in farm animals and confirms the improving role of this hormone in testicular blood flow as well as the protective role in sperm motility and morphology in rams that have been exposed to HS. There are also studies showing that injectable melatonin in rams [160] and goats [161] improved sperm quality during the non-breeding period. The above positive effects are considered to be due to the systemic or local action roles of melatonin in the testis but also to the increase in the blood flow of the testicles, which probably also acts as an initiation of their endocrine function [162]. Another theory claims that its improving effect is due to its antioxidant and antiapoptotic properties in the testicles and sperm [163]. Therefore, based on the above results, melatonin is considered a substance that can significantly mitigate the negative effects of testicular HS under field conditions. In Table 1, the effects of melatonin supplementation on sperm are summarized against the negative impacts of heat stress.
6. Assisted Reproductive Techniques (ARTs) and OS
6.1. Reproductive Technologies and OS
Sperm during ARTs receive a variety of in vitro manipulations in human and domestic animals [164]. Also, OS during ARTs could arise from several exogenous factors such as exposure to cryopreservation, visible light, O2 tension, centrifugation, culture media, pH, and temperature, which generates iatrogenic damage [164,165].
The results that the various reproductive techniques will give are affected by several factors, the most important of which is sperm OS. Potential oxidative damage to sperm DNA can lead to negative effects on embryo development or cause mutations in the offspring [166]. During ARTs, sperm are subjected to altered microenvironments, shearing forces, and a variety of exogenous factors that differ significantly from the physicochemical microenvironments of the female reproductive tract [164]. Sperm cryopreservation, laboratory handling, and washing techniques could increase the level of OS in sperm, especially in the case that the initial sperm ROS levels are already elevated [164].
Many studies have proven that the prolonged incubation of sperm during IVF (in vitro fertilization) procedures induces sperm DNA fragmentation (SDF) and OS both in human and animal models [167,168,169,170,171]. Characteristically, after incubation of sperm from healthy donors, reduced motility and increased RONS levels were observed starting from 6 h and reached a peak at 48 and 24 h, respectively [172].
Sperm concentration during ARTs also affects RONS levels, viability, and SDF in many species [164]. Post-thaw incubation of ram sperm at 6 to 100 × 106/mL for 6 h increased SDF, with lower sperm concentrations responding better [173]. In bovine, the level of sperm concentration during the storage of semen in a fresh extender affects the level of RONS, with higher concentrations of sperm pressing the sperm cell viability and increasing OS compared to lower sperm concentrations [174].
Oxygen tension during an IVF procedure could influence the levels of ROS in sperm. According to the usual method, sperm are incubated in 20% O2, but samples with high ROS levels could benefit from reduced O2 tensions [164]. A study reported no differences in sperm function for normozoospermic sperm samples after incubation under 5 versus 20% O2, whereas 5% O2 incubation was found beneficial for oligozoospermic samples that are known to have higher levels of ROS than fertile samples [175].
6.2. Beneficial Role of Melatonin
As an antioxidant, the use of melatonin, as a radical scavenger and antiapoptotic factor [176,177,178,179], has already been investigated. Indeed, many studies reported that melatonin reduced the rate of OS in human sperm, having beneficial effects on quality parameters, such as motility, DNA integrity, and lipid peroxidation [134,135,180,181,182,183,184].
Monllor et al. [184] suggested that certain levels of melatonin in the sperm environment from spermatogenesis to capacitation could be an effective strategy to avoid RONS-induced infertility. Melatonin, unlike other antioxidant compounds, reduces RONS levels to some extent, and low levels of RONS act as second messengers of capacitation [185].
Very recently, the use of melatonin to preserve human sperm quality, using cadmium (Cd) as an inductor of OS in vitro, was examined by Minucci and Venditti [186]. It is well known that testis is one of the main targets of this heavy metal, and many papers focused on its ability to generate OS, as well as other damages, compromising the normal spermatogenesis and decreasing sperm quality in both animal models [187,188] and humans [189,190]. Minucci and Venditti [186] further expanded the information concerning the protective action of melatonin when RONS generation is favored, using Cd as a pro-oxidative agent in human sperm in vitro.
According to the above researchers, the negative effects of cadmium-induced OS were counteracted by co-treatment with melatonin. In particular, its use improved sperm motility after 30 min of incubation compared to the control group, and at 24 h, it prevented the physiological alteration in terms of motility, DNA integrity, and apoptosis [186]. The above data lead to the conclusion that melatonin can be used to improve the quality of human gametes under stressful conditions [186].
Caspase-3 participates in cell apoptosis, being a good marker of cell death, and active caspase-3 is exclusively found in the midpiece of spermatozoa [191]. According to Monllor et al. [184], the use of melatonin in the sperm incubation medium before ART (storage at 37 °C for 30 min), in both normal and oligozoospermic samples, prevented an increase in the activity of caspase-3 as well as subsequent DNA fragmentation. Although the mechanism of action of melatonin for the above positive effects is not yet known, Espino et al. [192] reported that melatonin could reverse the apoptotic events caused by OS or elevated intracellular Ca2+ levels.
In human sperm, the expression of CD46, which is located in the inner acrosomal membrane [193], has an important role in the fusion process of the spermatozoon with the oocyte membrane [194] and therefore is considered an indicator of the fertilizing capacity of the sperm [184]. Based on the above, Monllor and colleagues [184] studied the effect melatonin had on CD46 expression in sperm samples. The results saw that melatonin increased the ratio of adequate sperm both in normal and oligozoospermic groups.
During the late stages of spermatogenesis in mammals, the sperm nucleus acquires high levels of chromatin condensation [184] to protect DNA from oxidative damage. When sperm DNA exhibits reduced condensation, chromatin is much more susceptible to OS and deletions, mutations, DNA cross-links, and chromosomal rearrangements [195], and it has been associated with subfertility in several species [196]. Melatonin, although unable to improve DNA compaction during the short time of sperm preparation for ART, can enhance the migration of sperm with compacted DNA and prevent its fragmentation [184]. In Table 2, the effects of melatonin supplementation on sperm are summarized against the negative impacts of ARTs.
Table 2.
Effects of melatonin supplementation on sperm against the negative impacts of ART stress.
7. Effects of Melatonin on Cryopreservation-Induced OS
7.1. Cryopreservation and OS
The cryopreservation of reproductive cells is known to be an important tool in human- and animal-assisted reproduction. Despite the benefits of cryopreservation, during this process, an imbalance is observed between the production of RONS and the antioxidant defense mechanism of the cells, known as OS [197]. Although all cells possess specific defense mechanisms against RONS, when these are overwhelmed, RONS can affect various cellular functions and processes [198]. Specifically, sperm are unable to cope with extreme RONS production mainly due to two reasons. The first is the low amount of antioxidant compounds due to the reduction in the cytoplasm in the final phases of spermatogenesis [199,200,201], and the second is the reduction in the antioxidant capacity of sperm due to the dilution of the seminal plasma, which is endowed with enzymatic and non-enzymatic antioxidants [202]. The above two reasons, as well as the high content of the sperm cell membrane in unsaturated fatty acids, make these cells vulnerable to RONS, leading to irreversible cell changes [203].
Oxidative stress is the main cause of most of the structural and molecular damages that occur in sperm during both freezing and thawing. Specifically, they lead to lipid peroxidation, DNA fragmentation, mitochondrial damage and dysfunction, protein oxidation, and loss or inactivation of enzymes associated with sperm motility [203]. OS is, also, one of the main factors associated with male infertility and reduced sperm viability and reproductive capacity. Additionally, OS has been linked to reduced rates of fertilization and in vitro embryo development in non-human mammals [29].
During the freezing–thawing process, motility is most affected [204,205], and this is due to mitochondrial damage and physical changes that occur in the tail of the spermatozoa [206]. In particular, the damage observed in the mitochondrial membrane interrupts the energy production process, thus reducing ATP production while the irreversible coiling of the flagellum caused by cryopreservation prevents the propulsive movement of the tail [206]. In addition, the greatest percentage of peroxidation and membrane damage takes place in the mitochondria, resulting in a reduction in ATP, loss of sperm motility, reduction in sperm–egg fusion, and DNA damage [202].
During the cryopreservation process, changes occur in the fluidity of the mitochondrial membrane, resulting in an increase in mitochondrial membrane potential (ΔΨm) and, by extension, the release of RONS [29]. In turn, these cause DNA damage leading to single/double-strand DNA breaks [29]. From ovine studies, it is implied that freezing–thawing injury to sperm mitochondria significantly reduces the ability of sperm to migrate through the cervix and survive in the female reproductive tract [29]. Accordingly, studies in bulls confirmed that mitochondrial dysfunction is due to the opening of the mitochondrial permeability transition pore in response to intracellular Ca2+ increases, and this was associated with the loss of ∆Ψm, decreased ATP content, increased RONS levels, and deterioration of plasma membrane integrity [29].
According to Figueroa et al. [207], significant structural alterations were detected in the midpiece and mitochondria in the cryopreserved sperm of fish. These alterations reduce the functionality of the mitochondria and the energy reserves of the cell, creating problems in cellular osmoregulation, ion exchange, lipid peroxidation, and enzymatic mechanisms that regulate motility [207]. According to Cabrita et al. [208] and Figueroa et al. [209], the mitochondria of cryopreserved sperm in the species Dicentrarchus labrax, Acipenser ruthenus, Cyprinus carpio L., Oncorhynchus mykiss, Salvelinus fontinalis, and Sparus aurata show increased sensitivity to frost damage. Additionally, in trout and Atlantic salmon, mitochondria show 40–50% of the mitochondrial membrane potential, while Atlantic salmon shows 61% of the mitochondrial membrane potential [207]. A positive correlation between mitochondrial membrane potential and fertilization rate in Onchorynchus mykiss and Salmo salar has also been reported, a fact possibly related to the reduced motility and fertilization capacity presented by the cryopreserved sperm of these two species [207].
7.2. Beneficial Role of Melatonin
Melatonin has been shown to protect sperm from oxidative damage, maintain sperm viability, reduce morphological abnormalities, and prevent DNA fragmentation [29]. The in vitro use of this hormone can improve the quality characteristics of human, ram, and pig sperm, while its use as an antioxidant agent in cryopreservation improves the quality of sperm after thawing [197].
Alevra et al. [197] reported the effects of the hormone melatonin both on the cryopreserved sperm of humans and of productive animals and fish. The addition of melatonin to cryopreserved solutions of buffalos [210,211], bovine [212], sheep [213], human [214,215], fish [216,217,218,219], and pig [220] sperm increased its viability after thawing and reduced morphological abnormalities. Unlike sperm enrichment with melatonin, no differences were observed during cryopreservation in sperm quality of goat semen [221]. Specifically, in several studies, it was shown that the administration of melatonin on both fresh and frozen sperm improved membrane integrity, motility and velocity, capacitation, antioxidant protein quantity, and developmental competence of sperm [222,223,224,225,226,227,228,229,230,231,232,233,234].
In cryopreserved bull sperm, the administration of 0.25 mM of melatonin increased VAP and VSL velocities, while the administration of 0.1 mM protected the plasma membrane and acrosome region and maintained the ultrastructure integrity of the sperm [229]. The administration of 0.2 mg/mL of melatonin to Mediterranean buffalo sperm improved its antioxidant capacity, motility, and morphology during cryopreservation [230]. Furthermore, the addition of 1 mM of melatonin in swamp buffalo bull sperm protected it from damage during cryopreservation [231]. In frozen–thawed pig sperm, 1.0 μM of melatonin presented higher viability and acrosome integrity, lower levels of peroxynitrite, ⋅O2−, and lipid peroxidation and diminished the levels of total ROS [220]. On the other hand, melatonin supplementation in canine sperm had no effects on it [232].
Our group [219], tested the effects of melatonin (0.5 mM, 1 mM, 1.5 mM, and 2 mM) on sea bream (Sparus aurata) sperm, stored at −196 °C and 4 °C. During short-term storage (4 °C), the melatonin improved sperm motility, allowing the sperm to remain motile for a longer storage period compared to fresh sperm and the control group. On the contrary, in the cryopreserved semen, no improvement was observed in the kinematic parameters of sperm [219].
Melatonin has the ability to maintain ΔΨm and preserve various mitochondrial functions. This is achieved by scavenging ROS and RNS and inhibiting the mitochondrial permeability transition pore (mPTP) opening. In addition, this hormone has the ability to regulate the expression of various antioxidant enzymes and genes that deal with stress [29].
Studies have shown that the use of melatonin as an antioxidant agent in cryopreserved ram sperm inhibits mPTP opening, thus improving sperm motility and viability, ATP synthesis, and oxygen consumption as well as the function of key OXPHOS enzymes [233,234]. Moreover, in ram sperm, the use of melatonin led to the suppression of mPTP opening, thus protecting the mitochondria and improving its quality characteristics during cryopreservation [234]. Additionally, the use of this hormone during the sperm equilibration period before cryopreservation led to an increase in plasma membrane integrity, ΔΨm, and mitochondrial Cyt C concentration, while correspondingly it helped to inhibit the mPTP opening and reduce enzymatic activity of Cyclophilin D (key mediator of the mPTP opening) [29]. Finally, the use of melatonin is shown to enhance the expression of the antiapoptotic genes Bcl-2 and heat shock protein 90 (HSP90), thereby conferring resistance to stressors in cryopreserved sperm [29]. In Table 3, the effects of melatonin supplementation on sperm are summarized against the negative impacts of cryopreservation.
Table 3.
Effects of melatonin supplementation on sperm against negative impacts of cryopreservation.
8. Antioxidant Role of Melatonin against Environmental Pollutants
Reproductive functions and embryo development procedures appear to be affected by chemical compounds, which are known as endocrine disruptors, resulting in degradations during the reproductive process in various species of birds, reptiles, fish, and mammals [237].
Chemical elements, such as organotin, whose use is mainly for biocides in antifouling paints, are considered responsible for reproductive disorders in the male genital organs of marine gastropods [237]. According to an experiment that was carried out on two different marine invertebrates, some nanoparticles such as nickel and copper oxide had spermiotoxic effects [237]. More specifically, these nanoparticles had negative effects on several semen parameters, such as concentration, morphology, motility, mitochondrial potential, and the generation of OS, which in turn affected the membrane and genome integrity [237].
Ribeiro et al. [238] supported that several heavy metals have a toxic effect on the male reproductive system, causing a reduction in semen quality and the production of RONS. Metals, such as iron (Fe), Cadmium (Cd), and Mercury (Hg), have been associated with the reduction in progressive motility and other motility parameters in species such as buffalo, bull, ram, and goat [238].
Generally, in the male reproductive system, an essential role of melatonin is the prevention of testicular damage, and it is effective in maintaining the blood–testis barrier in testes [239]. Concerning heavy metals, it is likely that melatonin plays an essential role in the reduction of negative impacts caused by Cadmium (Cd) in the male reproductive system, improving the DNA damage and suspending autophagy in sperm through the ATM/AMPK/mTOR signaling pathway [240].
Based on several studies carried out in recent years on the use of melatonin against metal-induced cytotoxicity, it appears that melatonin has a protective role against OS by counteracting free radicals and enhancing the antioxidant enzyme activity [239].
According to Li and colleagues [239], melatonin can be effective in toxicity reduction, induced by metals such as copper, iron, and molybdenum, due to its high lipophilicity, low toxicity, and ability to penetrate fast into cells [239]. The same research group studied the exposure of haexavalent chromium in spermatogonia and the role of melatonin in the protection against haexavalent chromium toxicity. They prove that melatonin can eliminate RONS by suppressing ATM p53 phosphorylation and the mitogen-activated protein kinase (MAPK) [239].
Dehdari Ebrahimi et al. [241] studied 38 publications, 31 of which were included in meta-analysis. The results of most studies presented useful data on the role of melatonin in the histopathological features of testicular tissue. Specifically, 20 toxic compounds were studied, some of which belonged to heavy metals while others to environmental pollutants [241]. The general conclusion reached by the researchers of this review, studying the effects of various chemical compounds in animal models, is that melatonin treatment significantly improved spermatological parameters (increased sperm count, motility, viability, and spermatozoa diameter), serum steroid hormone levels (testosterone and luteinizing hormone), antioxidant enzyme levels (superoxide dismutase and glutathione), and sperm morphometrics (epididymis weight and testicular weight) [241]. Despite this, the action of melatonin was found to be at lower levels, in relation to the apoptotic index and the treatment of different morphological changes in the structure of the sperm [241].
Except heavy metals, melatonin (MEL) showed a strong antioxidant action against Bisphenol S (BPS), an endocrine disrupting chemical, which contributes to cell apoptosis and causes negative effects in testis histomorphology (histological degenerative changes) and sperm quality parameters in adult male golden hamsters (Mesocricetus auratus) [242]. Reduction in serum levels of testosterone, melatonin, and testicular oxidative stress (TOS) was also associated with ΒPS exposure [242]. During treatment with MEL (10 mg/kg BW/day), there was an improvement in sperm quality parameters (viability and sperm count), restoration of the testicles to their original state, enhanced activity of antioxidant enzymes SOD and CAT, and a decrease in proteins NF-kB/COX-2, which are responsible for inflammation [242]. Finally, MEL seems to act as a stimulator in signaling pathways Nrf-2/HO-1 and SIRT-1/ FOXO-1, which are responsible for regulating antioxidant proteins [242].
Regarding the effect of melatonin in minimizing the toxicity of heavy metals in the testicles, Venditti et al. [243] concluded for the first time that the co-administration of melatonin (3 mg MEL/L) to cadmium-exposed rats weakened cadmium toxicity against genes DAAM1 and PREP expression in the testes [243]. In addition to this, the protective effect of melatonin against cadmium toxicity is confirmed by the reduction in oxidative stress and apoptosis as well as the restoration of histological and biomolecular changes (alterations in CAT and SOD activity) in normal levels, before causing damage to the testicles, due to exposure to cadmium [243]. It is considered appropriate to mention that the treatment exclusively used melatonin, and it was observed that this antioxidant agent is characterized by the ability to cause increased expression of PREP, which regulates the concentration of the hormone’s progesterone and GnRH, while its role is linked to microtubule processes [243]. Finally, the researchers presented new evidence on the mode of action of melatonin as a protective factor in other studied testicular parameters, such as in the improvement of impaired steroidogenesis (reduced protein level of StAR and 3β HSD) [243]. In Table 4, the effects of melatonin supplementation on sperm are summarized against the negative impacts of pollutants.
Table 4.
Effects of melatonin supplementation on sperm against stress of pollutants.
9. Future Perspectives
In the future, research emphasis should be placed on the mechanisms that activate the endogenous production of antioxidants, such as melatonin, to naturally protect sperm from OS [197]. An interesting case is the endogenous production of melatonin in the digestive tract through feeding protocols where varied plans of feeding schedules, different qualities of supplied food, and different sources of natural plant extracts (phytomelatonin) will be tested [244,245,246,247].
Additionally, the use of tryptophan as a dietary supplement could increase melatonin synthesis in the gut and by extension in the blood [245,248]. The important thing, in this case, is that the synthesis of melatonin does not depend on darkness and that the availability and quality of the food provided can affect its synthesis within the gut tissue [249]. On the other hand, the presence of increased levels of phytomelatonin in the blood is accompanied by an increase in serum antioxidant capacity [98,250,251]. More recently, Victoria Peña-Delgado et al. [252] found, for the first time, that a phytomelatonin-rich diet (including a mix of grape pulp, pomegranate, and tomato pomaces) can increase melatonin levels in seminal plasma of ram sperm, improving sperm viability and morphology and protecting sperm cells against oxidative damage.
Interestingly, from the nutrigenomics point of view, nutrients are feeding signals, which are detected with the cellular system of sensors and influence the expression of genes and proteins, and in consequence, the production of metabolites [253]. Nowadays, nutrigenomics using omic’s technologies investigates molecular relationships between nutrients and genes to identify how even minor modifications could potentially alter animal health and performance [254,255]. However, such types of experiments using gut melatonin and phytomelatonin by emphasizing nutrigenomics, reproduction, and antioxidant capacity in different species is still missing in the available literature.
Another approach, which is quite interesting, is the protective effect of endogenous-produced melatonin on sperm quality. The moment of the highest melatonin concentration in the bloodstream and seminal plasma varies between species, but normally, it rises during the dark period and falls to basal levels during the day [256,257,258]. Félix et al. [257] suggested that endogenously produced melatonin at mid-dark moment of the day may contribute to the improvement of some sperm parameters. In this way, this allows the aquaculture and livestock sectors to select sperm quality sperm by choosing the best moment of the day.
10. Conclusions
There are a variety of situations (heat stress, assisted reproductive technologies, and cryopreservation) that generate RONS in male germ cells. The increase in RONS above a threshold is not physiological and is harmful to the antioxidant defense system of males susceptible to OS, and antioxidants such as melatonin should be a significant factor in preserving the functional integrity of the sperm. Notably, the diet enriched by tryptophan and phytomelatonin supplementation and the suitable time of sperm collection may develop as useful techniques to increase endogenous melatonin in the gut and serum, respectively. Thereby, more carefully controlled research using animals representing different groups of vertebrates is required in the field of the use of endogenous and exogenous melatonin as an antioxidant.
Author Contributions
Conceptualization, S.P., A.I.A. and A.M.; methodology, S.P.; software, A.I.A.; validation, A.E.; formal analysis, S.P.; investigation, S.P., A.M., A.I.A. and A.E.; resources, S.P. and A.E.; data curation, S.P., A.M. and A.I.A.; writing—original draft preparation, S.P., A.I.A. and A.M.; writing—review and editing, S.P. and A.E.; visualization, S.P., A.M. and A.I.A.; supervision, S.P.; project administration, A.E.; funding acquisition, A.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P.; Gold, P.W. The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wei, F.; Li, G. The evolution of the concept of stress and the framework of the stress system. Cell Stress 2021, 5, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N. Roles and Functions of ROS and RNS in Cellular Physiology and Pathology. Cells 2020, 9, 767. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Bräuner, E.V.; Nordkap, L.; Priskorn, L.; Hansen, Å.M.; Bang, A.K.; Holmboe, S.A.; Schmidt, L.; Jensen, T.K.; Jørgensen, N. Psychological stress, stressful life events, male factor infertility, and testicular function: A cross-sectional study. Fertil. Steril. 2020, 113, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Gak, I.A.; Radovic, S.M.; Dukic, A.R.; Janjic, M.M.; Stojkov-Mimic, N.J.; Kostic, T.S.; Andric, S.A. Stress triggers mitochondrial biogenesis to preserve steroidogenesis in Leydig cells. Biochim. Biophys. Acta 2015, 1853, 2217–2227. [Google Scholar] [CrossRef] [PubMed]
- Starovlah, I.M.; Radovic Pletikosic, S.M.; Kostic, T.S.; Andric, S.A. Reduced spermatozoa functionality during stress is the consequence of adrenergic-mediated disturbance of mitochondrial dynamics markers. Sci. Rep. 2020, 10, 16813. [Google Scholar] [CrossRef]
- Starovlah, I.M.; Radovic Pletikosic, S.M.; Kostic, T.S.; Andric, S.A. Mitochondrial Dynamics Markers and Related Signaling Molecules Are Important Regulators of Spermatozoa Number and Functionality. Int. J. Mol. Sci. 2021, 22, 5693. [Google Scholar] [CrossRef]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Barbagallo, F.; La Vignera, S.; Cannarella, R.; Aversa, A.; Calogero, A.E.; Condorelli, R.A. Evaluation of Sperm Mitochondrial Function: A Key Organelle for Sperm Motility. J. Clin. Med. 2020, 9, 363. [Google Scholar] [CrossRef]
- Nordkap, L.; Jensen, T.K.; Hansen, Å.M.; Lassen, T.H.; Bang, A.K.; Joensen, U.N.; Jensen, M.B.; Skakkebæk, N.E.; Jørgensen, N. Psychological stress and testicular function: A cross-sectional study of 1,215 Danish men. Fertil. Steril. 2016, 105, 174–187.e171–172. [Google Scholar] [CrossRef]
- Manoli, I.; Alesci, S.; Blackman, M.R.; Su, Y.A.; Rennert, O.M.; Chrousos, G.P. Mitochondria as key components of the stress response. Trends Endocrinol. Metab. 2007, 18, 190–198. [Google Scholar] [CrossRef]
- Selye, H. Stress and disease. Science 1955, 122, 625–631. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S.; Lozach, A.; Habert, R.; Tanneux, M.; Guigon, C.; Brailly-Tabard, S.; Maltier, J.P.; Legrand-Maltier, C. Fertility and spermatogenesis are altered in α1b-adrenergic receptor knockout male mice. J. Endocrinol. 2007, 195, 281–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.Y.; Xu, D.P.; Li, H.B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A Pleiotropic, Orchestrating Regulator Molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Maitra, S.K.; Hasan, K.N. The Role of Melatonin as a Hormone and an Antioxidant in the Control of Fish Reproduction. Front. Endocrinol. 2016, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the Free Radical Scavenging Activities of Melatonin’s Metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Saleh, R.A.; Agarwal, A. Oxidative stress and male infertility: From research bench to clinical practice. J. Androl. 2002, 23, 737–752. [Google Scholar]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. [Google Scholar] [CrossRef]
- Lackner, J.E.; Agarwal, A.; Mahfouz, R.; du Plessis, S.S.; Schatzl, G. The association between leukocytes and sperm quality is concentration dependent. Reprod. Biol. Endocrinol. 2010, 8, 12. [Google Scholar] [CrossRef]
- Rengan, A.K.; Agarwal, A.; van der Linde, M.; du Plessis, S.S. An investigation of excess residual cytoplasm in human spermatozoa and its distinction from the cytoplasmic droplet. Reprod. Biol. Endocrinol. 2012, 10, 92. [Google Scholar] [CrossRef]
- Kothari, S.; Thompson, A.; Agarwal, A.; du Plessis, S.S. Free radicals: Their beneficial and detrimental effects on sperm function. Indian J. Exp. Biol. 2010, 48, 425–435. [Google Scholar]
- Huszar, G.; Corrales, M.; Vigue, L. Correlation between sperm creatine phosphokinase activity and sperm concentrations in normospermic and oligospermic men. Gamete Res. 1988, 19, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Huszar, G.; Vigue, L.; Corrales, M. Sperm creatine phosphokinase activity as a measure of sperm quality in normospermic, variablespermic, and oligospermic men. Biol. Reprod. 1988, 38, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Huszar, G.; Vigue, L.; Corrales, M. Sperm creatine kinase activity in fertile and infertile oligospermic men. J. Androl. 1990, 11, 40–46. [Google Scholar]
- Aitken, J.; Krausz, C.; Buckingham, D. Relationships between biochemical markers for residual sperm cytoplasm, reactive oxygen species generation, and the presence of leukocytes and precursor germ cells in human sperm suspensions. Mol. Reprod. Dev. 1994, 39, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Buckingham, D.W.; Brindle, J.; Lanzafame, F.; Irvine, D.S.; Aitken, R.J. Development of an image analysis system to monitor the retention of residual cytoplasm by human spermatozoa: Correlation with biochemical markers of the cytoplasmic space, oxidative stress, and sperm function. J. Androl. 1996, 17, 276–287. [Google Scholar]
- Gil-Guzman, E.; Ollero, M.; Lopez, M.C.; Sharma, R.K.; Alvarez, J.G.; Thomas, A.J., Jr.; Agarwal, A. Differential production of reactive oxygen species by subsets of human spermatozoa at different stages of maturation. Hum. Reprod. 2001, 16, 1922–1930. [Google Scholar] [CrossRef]
- Ollero, M.; Gil-Guzman, E.; Lopez, M.C.; Sharma, R.K.; Agarwal, A.; Larson, K.; Evenson, D.; Thomas, A.J., Jr.; Alvarez, J.G. Characterization of subsets of human spermatozoa at different stages of maturation: Implications in the diagnosis and treatment of male infertility. Hum. Reprod. 2001, 16, 1912–1921. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef]
- Das, S.K.; Vasudevan, D.M. Alcohol-induced oxidative stress. Life Sci. 2007, 81, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Singh, K.V.; Nirala, J.; Murmu, N.N.; Meena, R.; Rajamani, P. Oxidative stress-mediated alterations on sperm parameters in male Wistar rats exposed to 3G mobile phone radiation. Andrologia 2019, 51, e13201. [Google Scholar] [CrossRef] [PubMed]
- Iommiello, V.M.; Albani, E.; Di Rosa, A.; Marras, A.; Menduni, F.; Morreale, G.; Levi, S.L.; Pisano, B.; Levi-Setti, P.E. Ejaculate oxidative stress is related with sperm DNA fragmentation and round cells. Int. J. Endocrinol. 2015, 2015, 321901. [Google Scholar] [CrossRef]
- Almansa-Ordonez, A.; Bellido, R.; Vassena, R.; Barragan, M.; Zambelli, F. Oxidative Stress in Reproduction: A Mitochondrial Perspective. Biology 2020, 9, 269. [Google Scholar] [CrossRef]
- Dan Dunn, J.; Alvarez, L.A.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxidative Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef]
- Koppers, A.J.; Garg, M.L.; Aitken, R.J. Stimulation of mitochondrial reactive oxygen species production by unesterified, unsaturated fatty acids in defective human spermatozoa. Free Radic. Biol. Med. 2010, 48, 112–119. [Google Scholar] [CrossRef]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef]
- Ghani, E.; Keshtgar, S.; Habibagahi, M.; Ghannadi, A.; Kazeroni, M. Expression of NOX5 in human teratozoospermia compared to normozoospermia. Andrologia 2013, 45, 351–356. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, J.K.; Srivastava, N.; Ghosh, S.K. Strategies to Minimize Various Stress-Related Freeze-Thaw Damages during Conventional Cryopreservation of Mammalian Spermatozoa. Biopreserv. Biobank. 2019, 17, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Upreti, G.C.; Jensen, K.; Munday, R.; Duganzich, D.M.; Vishwanath, R.; Smith, J.F. Studies on aromatic amino acid oxidase activity in ram spermatozoa: Role of pyruvate as an antioxidant. Anim. Reprod. Sci. 1998, 51, 275–287. [Google Scholar] [CrossRef]
- Roca, J.; Martinez-Alborcia, M.J.; Gil, M.A.; Parrilla, I.; Martinez, E.A. Dead spermatozoa in raw semen samples impair in vitro fertilization outcomes of frozen-thawed spermatozoa. Fertil. Steril. 2013, 100, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Curson, B. Toxic effect and action of dead sperm on diluted bovine semen. J. Dairy Sci. 1972, 55, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Curson, B. Site of aromatic L-amino acid oxidase in dead bovine spermatozoa and determination of between-bull differences in the percentage of dead spermatozoa by oxidase activity. J. Reprod. Fertil. 1982, 64, 469–473. [Google Scholar] [CrossRef]
- Ayad, B.; Omolaoye, T.S.; Louw, N.; Ramsunder, Y.; Skosana, B.T.; Oyeipo, P.I.; Du Plessis, S.S. Oxidative Stress and Male Infertility: Evidence from a Research Perspective. Front. Reprod. Health 2022, 4, 822257. [Google Scholar] [CrossRef]
- Agarwal, A.; Mulgund, A.; Sharma, R.; Sabanegh, E. Mechanisms of oligozoospermia: An oxidative stress perspective. Syst. Biol. Reprod. Med. 2014, 60, 206–216. [Google Scholar] [CrossRef]
- Ritchie, C.; Ko, E.Y. Oxidative stress in the pathophysiology of male infertility. Andrologia 2021, 53, e13581. [Google Scholar] [CrossRef]
- de Lamirande, E.; Jiang, H.; Zini, A.; Kodama, H.; Gagnon, C. Reactive oxygen species and sperm physiology. Rev. Reprod. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Padron, O.F.; Brackett, N.L.; Sharma, R.K.; Lynne, C.M.; Thomas, A.J., Jr.; Agarwal, A. Seminal reactive oxygen species and sperm motility and morphology in men with spinal cord injury. Fertil. Steril. 1997, 67, 1115–1120. [Google Scholar] [CrossRef]
- Tvrdá, E.; Kňažická, Z.; Bárdos, L.; Massányi, P.; Lukáč, N. Impact of oxidative stress on male fertility—A review. Acta Vet. Hung. 2011, 59, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Simões, R.; Feitosa, W.B.; Siqueira, A.F.; Nichi, M.; Paula-Lopes, F.F.; Marques, M.G.; Peres, M.A.; Barnabe, V.H.; Visintin, J.A.; Assumpção, M.E. Influence of bovine sperm DNA fragmentation and oxidative stress on early embryo in vitro development outcome. Reproduction 2013, 146, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Wyck, S.; Herrera, C.; Requena, C.E.; Bittner, L.; Hajkova, P.; Bollwein, H.; Santoro, R. Oxidative stress in sperm affects the epigenetic reprogramming in early embryonic development. Epigenet Chromatin 2018, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; Yeste, M.; Salas-Huetos, A. The Relationship between Sperm Oxidative Stress Alterations and IVF/ICSI Outcomes: A Systematic Review from Nonhuman Mammals. Biology 2020, 9, 178. [Google Scholar] [CrossRef]
- Oborna, I.; Fingerova, H.; Novotny, J.; Brezinova, J.; Svobodova, M.; Aziz, N. Reactive oxygen species in human semen in relation to leukocyte contamination. Biomed. Pap. Med. Fac. Palacky Univ. Olomouc 2009, 153, 53–57. [Google Scholar] [CrossRef]
- Gautier, C.; Aurich, C. “Fine feathers make fine birds”—The mammalian sperm plasma membrane lipid composition and effects on assisted reproduction. Anim. Reprod. Sci. 2022, 246, 106884. [Google Scholar] [CrossRef]
- Bailey, J.L.; Blodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon minireview. J. Androl. 2000, 21, 1–7. [Google Scholar]
- Medeiros, C.M.; Forell, F.; Oliveira, A.T.; Rodrigues, J.L. Current status of sperm cryopreservation: Why isn’t it better? Theriogenology 2000, 57, 327–344. [Google Scholar] [CrossRef]
- Cocuzza, M.; Sikka, S.C.; Athayde, K.S.; Agarwal, A. Clinical relevance of oxidative stress and sperm chromatin damage in male infertility: An evidence based analysis. Int. Braz. J. Urol. 2007, 33, 603–621. [Google Scholar] [CrossRef]
- Vaughan, D.A.; Tirado, E.; Garcia, D.; Datta, V.; Sakkas, D. DNA fragmentation of sperm: A radical examination of the contribution of oxidative stress and age in 16 945 semen samples. Hum. Reprod. 2020, 35, 2188–2196. [Google Scholar] [CrossRef]
- Elbardisi, H.; Finelli, R.; Agarwal, A.; Majzoub, A.; Henkel, R.; Arafa, M. Predictive value of oxidative stress testing in semen for sperm DNA fragmentation assessed by sperm chromatin dispersion test. Andrology 2020, 8, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, R.Z.; du Plessis, S.S.; Aziz, N.; Sharma, R.; Sabanegh, E.; Agarwal, A. Sperm viability, apoptosis, and intracellular reactive oxygen species levels in human spermatozoa before and after induction of oxidative stress. Fertil. Steril. 2010, 93, 814–821. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef]
- Yüce, A.; Türk, G.; Çeribaşi, S.; Sönmez, M.; Çiftçi, M.; Güvenç, M. Effects of cinnamon (Cinnamomum zeylanicum) bark oil on testicular antioxidant values, apoptotic germ cell and sperm quality. Andrologia 2013, 45, 248–255. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Barik, G.; Chaturvedula, L.; Bobby, Z. Role of Oxidative Stress and Antioxidants in Male Infertility: An Interventional Study. J. Hum. Reprod. Sci. 2019, 12, 204–209. [Google Scholar] [CrossRef]
- De Luca, M.N.; Colone, M.; Gambioli, R.; Stringaro, A.; Unfer, V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants 2021, 10, 1283. [Google Scholar] [CrossRef] [PubMed]
- Ammar, O.; Mehdi, M.; Muratori, M. Teratozoospermia: Its association with sperm DNA defects, apoptotic alterations, and oxidative stress. Andrology 2020, 8, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Saleh, R.A.; Sharma, R.K.; Lewis-Jones, I.; Esfandiari, N.; Thomas, A.J., Jr.; Agarwal, A. Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil. Steril. 2004, 81, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Dobrakowski, M.; Kasperczyk, S.; Horak, S.; Chyra-Jach, D.; Birkner, E.; Kasperczyk, A. Oxidative stress and motility impairment in the semen of fertile males. Andrologia 2017, 49, e12783. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Nixon, B. Molecular Changes Induced by Oxidative Stress that Impair Human Sperm Motility. Antioxidants 2020, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Ayad, B.M.; Oyeyipo, I.P.; Van der Horst, G.; Du Plessis, S.S. Cementing the relationship between conventional advanced semen parameters. Middle East Fertil. Soc. J. 2021, 9, 134. [Google Scholar] [CrossRef]
- Barranco, I.; Rubio, C.P.; Tvarijonaviciute, A.; Rodriguez-Martinez, H.; Roca, J. Measurement of oxidative stress index in seminal plasma can predict in vivo fertility of liquid-stored porcine artificial insemination semen doses. Antioxidants 2021, 10, 1203. [Google Scholar] [CrossRef]
- Vigolo, V.; Giaretta, E.; da Dalt, L.; Damiani, J.; Gabai, G.; Bertuzzo, F.; Falomo, M.E. Relationships between Biomarkers of Oxidative Stress in Seminal Plasma and Sperm Motility in Bulls before and after Cryopreservation. Animals 2022, 12, 2534. [Google Scholar] [CrossRef]
- Sprenger, J.; Hardeland, R.; Fuhrberg, B.; Han, S.-Z. Melatonin and Other 5-Methoxylated Indoles in Yeast: Presence in High Concentrations and Dependence on Tryptophan Availability. Cytologia 1999, 64, 209–213. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. Camb. Philos. Soc. 2010, 85, 607–623. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Manchester, L.C.; Tan, D.X. Peripheral reproductive organ health and melatonin: Ready for prime time. Int. J. Mol. Sci. 2013, 14, 7231–7272. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Rosales-Corral, S.A. Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology. Hum. Reprod. Update 2014, 20, 293–307. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; et al. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res. 2012, 5, 5. [Google Scholar] [CrossRef]
- Cruz, M.H.; Leal, C.L.; Cruz, J.F.; Tan, D.X.; Reiter, R.J. Essential actions of melatonin in protecting the ovary from oxidative damage. Theriogenology 2014, 82, 925–932. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; et al. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar]
- Chan, K.H.; Wong, Y.H. A molecular and chemical perspective in defining melatonin receptor subtype selectivity. Int. J. Mol. Sci. 2013, 14, 18385–18406. [Google Scholar] [CrossRef] [PubMed]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on melatonin receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [PubMed]
- Alkozi, H.A.; Sanchez Montero, J.M.; Doadrio, A.L.; Pintor, J. Docking studies for melatonin receptors. Expert Opin. Drug Discov. 2018, 13, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Smith, D.G.; Hardeland, R.; Yang, M.Y.; Xu, H.L.; Zhang, L.; Yin, H.D.; Zhu, Q. Melatonin receptor genes in vertebrates. Int. J. Mol. Sci. 2013, 14, 11208–11223. [Google Scholar] [CrossRef]
- Munley, K.M.; Dutta, S.; Jasnow, A.M.; Demas, G.E. Adrenal MT1 melatonin receptor expression is linked with seasonal variation in social behavior in male Siberian hamsters. Horm. Behav. 2022, 138, 105099. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. The potential of phytomelatonin as a nutraceutical. Molecules 2018, 23, 238. [Google Scholar] [CrossRef]
- Jan, J.E.; Reiter, R.J.; Wasdell, M.B.; Bax, M. The role of the thalamus in sleep, pineal melatonin production, and circadian rhythm sleep disorders. J. Pineal Res. 2009, 46, 1–7. [Google Scholar] [CrossRef]
- Paredes, S.D.; Marchena, A.M.; Bejarano, I.; Espino, J.; Barriga, C.; Rial, R.V.; Reiter, R.J.; Rodríguez, A.B. Melatonin and tryptophan affect the activity-rest rhythm, core and peripheral temperatures, and interleukin levels in the ringdove: Changes with age. J. Gerontol.-Ser. A Biol. Sci. Med. Sci. 2009, 64, 340–350. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in Aging and Disease-Multiple Consequences of Reduced Secretion, Options and Limits of Treatment. Aging Dis. 2012, 3, 194–225. [Google Scholar] [PubMed]
- Witt-Enderby, P.A.; Radio, N.M.; Doctor, J.S.; Davis, V.L. Therapeutic treatments potentially mediated by melatonin receptors: Potential clinical uses in the prevention of osteoporosis, cancer and as an adjuvant therapy. J. Pineal Res. 2006, 41, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Mociková, K.; Mníchová, M.; Kubatka, P.; Bojková, B.; Ahlers, I.; Ahlersová, E. Mammary carcinogenesis induced in Wistarhan rats by the combination of ionizing radiation and dimethylbenz(a)anthracene: Prevention with melatonin. Neoplasma 2000, 47, 227–229. [Google Scholar]
- Claustrat, B.; Brun, J.; Chazot, G. The basic physiology and pathophysiology of melatonin. Sleep Med. Rev. 2005, 9, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Pandi-Perumal, S.R.; Maestroni, G.J.M.; Esquifino, A.I.; Hardeland, R.; Cardinali, D.P. Role of melatonin in neurodegenerative diseases. Neurotoxicol. Res. 2005, 7, 293–318. [Google Scholar] [CrossRef]
- Waterhouse, J.; Reilly, T.; Atkinson, G. Jet lag. Lancet 1997, 350, 1611–1616. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, S.; Zhang, Y.; Zhang, Q. Melatonin Receptors: A Key Mediator in Animal Reproduction. Vet. Sci. 2022, 9, 309. [Google Scholar] [CrossRef]
- Oishi, A.; Karamitri, A.; Gerbier, R.; Lahuna, O.; Ahmad, R.; Jockers, R. Orphan GPR61, GPR62 and GPR135 receptors and the melatonin MT(2) receptor reciprocally modulate their signaling functions. Sci. Rep. 2017, 7, 8990. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, X.; Zhao, S.; Zhang, Y.; Ma, H.; Yang, Z.; Yang, W.; Zhao, C.; Wang, L.; Zhang, Q. Melatonin receptor depletion suppressed hCG-induced testosterone expression in mouse Leydig cells. Cell. Mol. Biol. Lett. 2019, 24, 21. [Google Scholar] [CrossRef]
- Chabra, A.; Shokrzadeh, M.; Naghshvar, F.; Salehi, F.; Ahmadi, A. Melatonin ameliorates oxidative stress and reproductive toxicity induced by cyclophosphamide in male mice. Hum. Exp. Toxicol. 2014, 33, 185–195. [Google Scholar] [CrossRef]
- Vargas, A.; Bustos-Obregón, E.; Hartley, R. Effects of hypoxia on epididymal sperm parameters and protective role of ibuprofen and melatonin. Biol. Res. 2011, 44, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Tang, K.Q.; Fu, C.Z.; Riaz, H.; Zhang, Q.; Zan, L.S. Melatonin regulates the development and function of bovine Sertoli cells via its receptors MT1 and MT2. Anim. Reprod. Sci. 2014, 147, 10–16. [Google Scholar] [CrossRef] [PubMed]
- ViviD, D.; Bentley, G.E. Seasonal Reproduction in Vertebrates: Melatonin Synthesis, Binding, and Functionality Using Tinbergen’s Four Questions. Molecules 2018, 23, 652. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C. Gene regulation by melatonin. Ann. N. Y. Acad. Sci. 2000, 917, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Delagrange, P.; Krause, D.N.; Sugden, D.; Cardinali, D.P.; Olcese, J. International Union of Basic and Clinical Pharmacology. LXXV. Nomenclature, classification, and pharmacology of G protein-coupled melatonin receptors. Pharmacol. Rev. 2010, 62, 343–380. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 Melatonin Receptors: A Therapeutic Perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A Mitochondrial Targeting Molecule Involving Mitochondrial Protection and Dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef]
- McCubrey, J.A.; LaHair, M.M.; Franklin, R.A. Reactive oxygen species-induced activation of the MAP kinase signaling pathways. Antioxid. Redox Signal. 2006, 8, 1775–1789. [Google Scholar] [CrossRef]
- Franco, R.; Navarro, G.; Martínez-Pinilla, E. Antioxidant defense mechanisms in erythrocytes and in the central nervous system. Antioxidants 2019, 8, 46. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.-X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Duan, W.; Jin, Z.; Yi, W.; Yan, J.; Zhang, S.; Wang, N.; Liang, Z.; Li, Y.; Chen, W.; et al. JAK2/STAT3 activation by melatonin attenuates the mitochondrial oxidative damage induced by myocardial ischemia/reperfusion injury. J. Pineal Res. 2013, 55, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Betti, M.; Canonico, B.; Arcangeletti, M.; Ferri, P.; Galli, F.; Papa, S. ERK MAPK activation mediates the antiapoptotic signaling of melatonin in UVB-stressed U937 cells. Free Radic. Biol. Med. 2009, 46, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J. 2010, 24, 3603–3624. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Linkova, N.; Polyakova, V.; Mironova, E.; Hashimova, U.; Gadzhiev, A.; Safikhanova, K.; Krylova, T.K.J.; Tarquini, R.; Mazzoccoli, G.; et al. Melatonin and Sirtuins in Buccal Epithelium: Potential Biomarkers of Aging and Age-Related Pathologies. Int. J. Mol. Sci. 2020, 21, 8134. [Google Scholar] [CrossRef]
- Mayo, J.C.; Sainz, R.M.; Gonzalez Menendez, P.; Cepas, V.; Tan, D.X.; Reiter, R.J. Melatonin and sirtuins: A “not-so unexpected” relationship. J. Pineal Res. 2017, 62, e12391. [Google Scholar] [CrossRef]
- Kunst, S.; Wolloscheck, T.; Kelleher, D.K.; Wolfrum, U.; Sargsyan, S.A.; Iuvone, P.M.; Baba, K.; Tosini, G.; Spessert, R. Pgc-1alpha and Nr4a1 Are Target Genes of Circadian Melatonin and Dopamine Release in Murine Retina. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6084–6094. [Google Scholar] [CrossRef]
- Bornman, M.; Oosthuizen, J.; Barnard, H.; Schulenburg, G.; Boomker, D.; Reif, S. Melatonin and sperm motility/melatonin und spermatozoenmotilität. Andrologia 1989, 21, 483–485. [Google Scholar] [CrossRef]
- Gwayi, N.; Bernard, R. The effects of melatonin on sperm motility in vitro in Wistar rats. Andrologia 2002, 34, 391–396. [Google Scholar] [CrossRef]
- Casao, A.; Pérez-Pé, R.; Abecia, J.A.; Forcada, F.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á. The effect of exogenous melatonin during the non-reproductive season on the seminal plasma hormonal profile and the antioxidant defense system of Rasa Aragonesa rams. Anim. Reprod. Sci. 2013, 138, 168–174. [Google Scholar] [CrossRef]
- Kokolis, N.; Theodosiadou, E.; Tsantarliotou, M.; Rekkas, C.; Goulas, P.; Smokovitis, A. The effect of melatonin implants on blood testosterone and acrosin activity in spermatozoa of the ram. Andrologia 2000, 32, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Haldar, C. Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and steroidogenesis in the testis of adult golden hamster, Mesocricetus auratus. J. Photochem. Photobiol. B Biol. 2014, 140, 374–380. [Google Scholar] [CrossRef] [PubMed]
- González-Arto, M.; Vicente-Carrillo, A.; Martínez-Pastor, F.; Fernández-Alegre, E.; Roca, J.; Miró, J.; Rigau, T.; Rodríguez-Gil, J.E.; Pérez-Pé, R.; Muiño-Blanco, T. Melatonin receptors MT 1 and MT 2 are expressed in spermatozoa from several seasonal and nonseasonal breeder species. Theriogenology 2016, 86, 1958–1968. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Whiting, S.; Lambourne, S.; Aitken, R.J.; Sun, Y.P. Melatonin alleviates heat stress-induced oxidative stress and apoptosis in human spermatozoa. Free Radic. Biol. Med. 2021, 164, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Espino, J.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Pariente, J.A.; Rodríguez, A.B. High endogenous melatonin concentrations enhance sperm quality and short-term in vitro exposure to melatonin improves aspects of sperm motility. J. Pineal Res. 2011, 50, 132–139. [Google Scholar] [CrossRef]
- du Plessis, S.S.; Hagenaar, K.; Lampiao, F. The in vitro effects of melatonin on human sperm function and its scavenging activities on NO and ROS. Andrologia 2010, 42, 112–116. [Google Scholar] [CrossRef]
- Qin, D.Z.; Cai, H.; He, C.; Yang, D.H.; Sun, J.; He, W.L.; Li, B.L.; Hua, J.L.; Peng, S. Melatonin relieves heat-induced spermatocyte apoptosis in mouse testes by inhibition of ATF6 and PERK signaling pathways. Zool. Res. 2021, 42, 514–524. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, H.; Wang, Q.J.; Qi, X.; Li, Q.; Fu, W.; Huang, J.; Yao, C.Y.; Liu, Z.Y.; Wang, M.Z.; et al. Prolonged melatonin treatment promote testicular recovery by enhancing RAC1-mediated apoptotic cell clearance and cell junction-dependent spermatogensis after heat stress. Theriogenology 2021, 162, 22–31. [Google Scholar] [CrossRef]
- Shahat, A.M.; Thundathil, J.C.; Kastelic, J.P. Melatonin or L-arginine in semen extender mitigate reductions in quality of frozen-thawed sperm from heat-stressed rams. Theriogenology 2022, 188, 163–169. [Google Scholar] [CrossRef]
- Egerszegi, I.; Sarlós, P.; Rátky, J.; Solti, L.; Faigl, V.; Kulcsár, M.; Cseh, S. Effect of melatonin treatment on semen parameters and endocrine function in Black Racka rams out of the breeding season. Small Rumin. Res. 2014, 116, 192–198. [Google Scholar] [CrossRef]
- Vince, S.; Valpotic, H.; Berta, V.; Milinkovic-Tur, S.; Samardzija, M.; Grizelj, J.; Spoljaric, B.; Ðuricic, D.; Nazansky, I.; Zura Zaja, I. Monitoring of libido and semen quality parameters in melatonin-treated French alpine bucks during the non-breeding season. Reprod. Domest. Anim. 2017, 52, 953–961. [Google Scholar] [CrossRef]
- Ramadan, T.A.; Kumar, D.; Ghuman, S.S.; Singh, I. Melatonin-improved buffalo semen quality during nonbreeding season under tropical condition. Domest. Anim. Endocrinol. 2019, 68, 119–125. [Google Scholar] [CrossRef]
- da Silva, C.M.; Macías-García, B.; Miró-Morán, A.; González-Fernández, L.; Morillo-Rodriguez, A.; Ortega-Ferrusola, C.; Gallardo-Bolaños, J.M.; Stilwell, G.; Tapia, J.A.; Peña, F.J. Melatonin reduces lipid peroxidation and apoptotic-like changes in stallion spermatozoa. J. Pineal Res. 2011, 51, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin protects ram spermatozoa from cryopreservation injuries in a dose-dependent manner. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Benitez-King, G.; Anton-Tay, F. Calmodulin mediates melatonin cytoskeletal effects. Experientia 1993, 49, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Delgadillo, L.H.; Tay, F.A.A.; King, G.B. Effect of melatonin on microtubule assembly depend on hormone concentration: Role of melatonin as a calmodulin antagonist. J. Pineal Res. 1994, 17, 50–58. [Google Scholar]
- Si, Y. Temperature-dependent hyperactivated movement of hamster spermatozoa. Biol. Reprod. 1997, 57, 1407–1412. [Google Scholar] [CrossRef][Green Version]
- Tash, J.S.; Means, A.R. Cyclic adenosine 3′, 5′ monosphate, calcium and protein phosphorylation in flagellar motility. Biol. Reprod. 1983, 28, 75–104. [Google Scholar] [CrossRef]
- Yung, L.Y.; Tsim, S.T.; Wong, Y.H. Stimulation of cAMP accumulation by the cloned Xenopus melatonin receptor through G1 AND G2 proteins. FFBS Lett. 1995, 372, 99–102. [Google Scholar] [CrossRef]
- Lindamann, C.B. A cAMP-induced increase in the motility of demembranated bull sperm models. Cell 1978, 13, 9–18. [Google Scholar] [CrossRef]
- Garbers, D.L.; Kopf, G.S. The Regulation of Spermatozoa by Calcium and Cyclic Nucleotides; Advances in Cyclic Nucleotide Research; Robinson, G.A., Ed.; Raven Press: New York, NY, USA, 1980; pp. 251–306. [Google Scholar]
- Starkov, A.A. The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. N. Y. Acad. Sci. 2008, 1147, 37–52. [Google Scholar] [CrossRef]
- Gong, Y.; Guo, H.; Zhang, Z.; Zhou, H.; Zhao, R.; He, B. Heat Stress Reduces Sperm Motility via Activation of Glycogen Synthase Kinase-3α and Inhibition of Mitochondrial Protein Import. Front. Physiol. 2017, 8, 718. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.R.; Chen, G.W.; Shi, H.J.; O, W.S.; Martin-DeLeon, P.A.; Chen, H. Prohibitin involvement in the generation of mitochondrial superoxide at complex I in human sperm. J. Cell. Mol. Med. 2017, 21, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.B.; Vandaele, L.; Rijsselaere, T.; El-Deen, M.S.; Maes, D.; Shamsuddin, M.; Van Soom, A. Bovine spermatozoa react to in vitro heat stress by activating the mitogen-activated protein kinase 14 signalling pathway. Reprod. Fertil. Dev. 2014, 26, 245–257. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Koppers, A.J.; De Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of mitochondrial reactive oxygen species in the generation of oxidative stress in spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; Al-Gabri, N.A.; Hashem, N.M.; Gonzalez-Bulnes, A. Supplementation with Proline Improves Haemato-Biochemical and Reproductive Indicators in Male Rabbits Affected by Environmental Heat-Stress. Animals 2021, 11, 373. [Google Scholar] [CrossRef]
- Qiao, N.; Chen, H.; Du, P.; Kang, Z.; Pang, C.; Liu, B.; Zeng, Q.; Pan, J.; Zhang, H.; Mehmood, K.; et al. Acetyl-L-Carnitine Induces Autophagy to Promote Mouse Spermatogonia Cell Recovery after Heat Stress Damage. Biomed Res. Int. 2021, 2021, 8871328. [Google Scholar] [CrossRef]
- Cui, Y.; Ren, L.; Li, B.; Fang, J.; Zhai, Y.; He, X.; Du, E.; Miao, Y.; Hua, J.; Peng, S. Melatonin Relieves Busulfan-Induced Spermatogonial Stem Cell Apoptosis of Mouse Testis by Inhibiting Endoplasmic Reticulum Stress. Cell. Physiol. Biochem. 2017, 44, 2407–2421. [Google Scholar] [CrossRef]
- El-Shalofy, A.S.; Shahat, A.M.; Hedia, M.G. Effects of melatonin administration on testicular hemodynamics, echotexture, steroids production, and semen parameters during the non-breeding season in Ossimi rams. Theriogenology 2022, 184, 34–40. [Google Scholar] [CrossRef]
- Samir, H.; Nyametease, P.; Elbadawy, M.; Nagaoka, K.; Sasaki, K.; Watanabe, G. Administration of melatonin improves testicular blood flow, circulating hormones, and semen quality in Shiba goats. Theriogenology 2020, 146, 111–119. [Google Scholar] [CrossRef]
- Shahat, A.M.; Rizzoto, G.; Kastelic, J.P. Amelioration of heat stress-induced damage to testes and sperm quality. Theriogenology 2020, 158, 84–96. [Google Scholar] [CrossRef]
- Zhang, P.; Zheng, Y.; Lv, Y.; Li, F.; Su, L.; Qin, Y.; Zeng, W. Melatonin protects the mouse testis against heat-induced damage. Mol. Hum. Reprod. 2020, 26, 65–79. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Longobardi, S.; Di Rella, F.; Adiga, S.K.; Talevi, R. Sperm Oxidative Stress during In Vitro Manipulation and Its Effects on Sperm Function and Embryo Development. Antioxidants 2021, 10, 1025. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.H.; Leal, C.L.; da Cruz, J.F.; Tan, D.X.; Reiter, R.J. Role of melatonin on production and preservation of gametes and embryos: A brief review. Anim. Reprod. Sci. 2014, 145, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A. The Role of Genetics and Oxidative Stress in the Etiology of Male Infertility-A Unifying Hypothesis? Front. Endocrinol. 2020, 11, 581838. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Gibb, Z.; Mitchell, L.A.; Lambourne, S.R.; Connaughton, H.S.; De Iuliis, G.N. Sperm motility is lost in vitro as a consequence of mitochondrial free radical production and the generation of electrophilic aldehydes but can be significantly rescued by the presence of nucleophilic thiols. Biol. Reprod. 2012, 87, 110. [Google Scholar] [CrossRef]
- Simon, L.; Murphy, K.; Aston, K.I.; Emery, B.R.; Hotaling, J.M.; Carrell, D.T. Micro-electrophoresis: A noninvasive method of sperm selection based on membrane charge. Fertil. Steril. 2015, 103, 361–366.e363. [Google Scholar] [CrossRef]
- Banihani, S.; Sharma, R.; Bayachou, M.; Sabanegh, E.; Agarwal, A. Human sperm DNA oxidation, motility and viability in the presence of L-carnitine during in vitro incubation and centrifugation. Andrologia 2012, 44 (Suppl. S1), 505–512. [Google Scholar] [CrossRef]
- Gualtieri, R.; Barbato, V.; Fiorentino, I.; Braun, S.; Rizos, D.; Longobardi, S.; Talevi, R. Treatment with zinc, d-aspartate, and coenzyme Q10 protects bull sperm against damage and improves their ability to support embryo development. Theriogenology 2014, 82, 592–598. [Google Scholar] [CrossRef]
- Nematollahi, S.; Mehdizadeh, M.; Hosseini, S.; Kashanian, M.; Amjadi, F.S.; Salehi, M. DNA integrity and methylation changes of mouse spermatozoa following prolonged incubation. Andrologia 2019, 51, e13276. [Google Scholar] [CrossRef]
- Calamera, J.C.; Fernandez, P.J.; Buffone, M.G.; Acosta, A.A.; Doncel, G.F. Effects of long-term in vitro incubation of human spermatozoa: Functional parameters and catalase effect. Andrologia 2001, 33, 79–86. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández, C.; Johnston, S.D.; Fernández, J.L.; Wilson, R.J.; Gosálvez, J. Fragmentation dynamics of frozen-thawed ram sperm DNA is modulated by sperm concentration. Theriogenology 2010, 74, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Fahey, A.G.; Shafat, A.; Fair, S. Reducing sperm concentration is critical to limiting the oxidative stress challenge in liquid bull semen. J. Dairy Sci. 2013, 96, 4447–4454. [Google Scholar] [CrossRef] [PubMed]
- Griveau, J.F.; Grizard, G.; Boucher, D.; Le Lannou, D. Influence of oxygen tension on function of isolated spermatozoa from ejaculates of oligozoospermic patients and normozoospermic fertile donors. Hum. Reprod. 1998, 13, 3108–3113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galano, A.; Reiter, R.J. Melatonin and its metabolites vs. oxidative stress: From individual actions to collective protection. J. Pineal Res. 2018, 65, e12514. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Bernatoniene, J. Molecular Mechanisms of Melatonin-Mediated Cell Protection and Signaling in Health and Disease. Pharmaceutics 2021, 13, 129. [Google Scholar] [CrossRef]
- Loren, P.; Sánchez, R.; Arias, M.E.; Felmer, R.; Risopatrón, J.; Cheuquemán, C. Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals. Int. J. Mol. Sci. 2017, 18, 1119. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Qi, W. Biochemical reactivity of melatonin with reactive oxygen and nitrogen species: A review of the evidence. Cell Biochem. Biophys. 2001, 34, 237–256. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Xiong, Y.M.; Tan, Y.J.; Wang, L.; Li, R.; Zhang, Y.; Liu, X.M.; Lin, X.H.; Jin, L.; Hu, Y.T.; et al. Melatonin rescues impaired penetration ability of human spermatozoa induced by mitochondrial dysfunction. Reproduction 2019, 158, 465–475. [Google Scholar] [CrossRef]
- Karimfar, M.H.; Niazvand, F.; Haghani, K.; Ghafourian, S.; Shirazi, R.; Bakhtiyari, S. The protective effects of melatonin against cryopreservation-induced oxidative stress in human sperm. Int. J. Immunopathol. Pharmacol. 2015, 28, 69–76. [Google Scholar] [CrossRef]
- Bejarano, I.; Monllor, F.; Marchena, A.M.; Ortiz, A.; Lozano, G.; Jiménez, M.I.; Gaspar, P.; García, J.F.; Pariente, J.A.; Rodríguez, A.B.; et al. Exogenous melatonin supplementation prevents oxidative stress-evoked DNA damage in human spermatozoa. J. Pineal Res. 2014, 57, 333–339. [Google Scholar] [CrossRef]
- Malmir, M.; Naderi Noreini, S.; Ghafarizadeh, A.; Faraji, T.; Asali, Z. Ameliorative effect of melatonin on apoptosis, DNA fragmentation, membrane integrity and lipid peroxidation of spermatozoa in the idiopathic asthenoteratospermic men: In vitro. Andrologia 2021, 53, e13944. [Google Scholar] [CrossRef] [PubMed]
- Monllor, F.; Espino, J.; Marchena, A.M.; Ortiz, Á.; Lozano, G.; García, J.F.; Pariente, J.A.; Rodríguez, A.B.; Bejarano, I. Melatonin diminishes oxidative damage in sperm cells, improving assisted reproductive techniques. Turk. J. Biol. 2017, 41, 881–889. [Google Scholar] [CrossRef] [PubMed]
- de Lamirande, E.; Gagnon, C. Origin of a motility inhibitor within the male reproductive tract. J. Androl. 1984, 5, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Minucci, S.; Venditti, M. New Insight on the In Vitro Effects of Melatonin in Preserving Human Sperm Quality. Int. J. Mol. Sci. 2022, 23, 5128. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, T.; Cao, M.; Zhao, Z.; Zhao, Y.; Chen, L.; Chen, T.; Li, C.; Zhou, X. Damage to the Testicular Structure of Rats by Acute Oral Exposure of Cadmium. Int. J. Environ. Res. Public Health 2021, 18, 6038. [Google Scholar] [CrossRef]
- Bhardwaj, J.K.; Panchal, H.; Saraf, P. Cadmium as a testicular toxicant: A Review. J. Appl. Toxicol. 2021, 41, 105–117. [Google Scholar] [CrossRef]
- Santonastaso, M.; Mottola, F.; Iovine, C.; Cesaroni, F.; Colacurci, N.; Rocco, L. In Vitro Effects of Titanium Dioxide Nanoparticles (TiO2NPs) on Cadmium Chloride (CdCl2) Genotoxicity in Human Sperm Cells. Nanomaterials 2020, 10, 1118. [Google Scholar] [CrossRef]
- Marchiani, S.; Tamburrino, L.; Farnetani, G.; Muratori, M.; Vignozzi, L.; Baldi, E. Acute effects on human sperm exposed in vitro to cadmium chloride and diisobutyl phthalate. Reproduction 2019, 158, 281–290. [Google Scholar] [CrossRef]
- Oehninger, S.; Morshedi, M.; Weng, S.L.; Taylor, S.; Duran, H.; Beebe, S. Presence and significance of somatic cell apoptosis markers in human ejaculated spermatozoa. Reprod. Biomed. Online 2003, 7, 469–476. [Google Scholar] [CrossRef]
- Espino, J.; Bejarano, I.; Ortiz, A.; Lozano, G.M.; García, J.F.; Pariente, J.A.; Rodríguez, A.B. Melatonin as a potential tool against oxidative damage and apoptosis in ejaculated human spermatozoa. Fertil. Steril. 2010, 94, 1915–1917. [Google Scholar] [CrossRef]
- Anderson, D.J.; Michaelson, J.S.; Johnson, P.M. Trophoblast/Leukocyte-Common Antigen is Expressed by Human Testicular Germ Cells and Appears on the Surface of Acrosome-Reacted Sperm1. Biol. Reprod. 1989, 41, 285–293. [Google Scholar] [CrossRef]
- Taylor, C.T.; Biljan, M.M.; Kingsland, C.R.; Johnson, P.M. Inhibition of human spermatozoon-oocyte interaction in vitro by monoclonal antibodies to CD46 (membrane cofactor protein). Hum. Reprod. 1994, 9, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Schulte, R.T.; Ohl, D.A.; Sigman, M.; Smith, G.D. Sperm DNA damage in male infertility: Etiologies, assays, and outcomes. J. Assist. Reprod. Genet. 2010, 27, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hekmatdoost, A.; Lakpour, N.; Sadeghi, M.R. Sperm chromatin integrity: Etiologies and mechanisms of abnormality, assays, clinical importance, preventing and repairing damage. Avicenna J. Med. Biotechnol. 2009, 1, 147–160. [Google Scholar]
- Alevra, A.I.; Exadactylos, A.; Mente, E.; Papadopoulos, S. The Protective Role of Melatonin in Sperm Cryopreservation of Farm Animals and Human: Lessons for Male Fish Cryopreservation. Animals 2022, 12, 791. [Google Scholar] [CrossRef] [PubMed]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.W.; de França, L.R.; Lareyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- O’Donnell, L.; O’Bryan, M.K. Microtubules and spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 45–54. [Google Scholar] [CrossRef]
- Ciereszko, A.; Dietrich, M.A.; Nynca, J. Fish semen proteomics—New opportunities in fish reproductive research. Aquaculture 2017, 472, 81–92. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Vento, M.; Ciriminna, R.; Artini, P.G. Cryopreservation and oxidative stress in reproductive cells. Gynecol. Endocrinol. 2010, 26, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Vargas, L.; Silva Jiménez, M.; Risopatrón González, J.; Villalobos, E.F.; Cabrita, E.; Valdebenito Isler, I. Oxidative stress and use of antioxidants in fish semen cryopreservation. Rev. Aquac. 2021, 13, 365–387. [Google Scholar] [CrossRef]
- Donnelly, E.T.; McClure, N.; Lewis, S.E. Cryopreservation of human semen and prepared sperm: Effects on motility parameters and DNA integrity. Fertil. Steril. 2001, 76, 892–900. [Google Scholar] [CrossRef]
- Gandini, L.; Lombardo, F.; Lenzi, A.; Spanò, M.; Dondero, F. Cryopreservation and sperm DNA integrity. Cell Tissue Bank. 2006, 7, 91–98. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, M.; McClure, N.; Lewis, S.E. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, E.; Lee-Estevez, M.; Valdebenito, I.; Watanabe, I.; Oliveira, R.P.S.; Romero, J.; Castillo, R.L.; Farías, J.G. Effects of cryopreservation on mitochondrial function and sperm quality in fish. Aquaculture 2019, 511, 634190. [Google Scholar] [CrossRef]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.G.; Sarasquete, C.; Herráez, M.P.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Figueroa, E.; Valdebenito, I.; Zepeda, A.B.; Figueroa, C.A.; Dumorné, K.; Castillo, R.L.; Farias, J.G. Effects of cryopreservation on mitochondria of fish spermatozoa. Rev. Aquac. 2017, 9, 76–87. [Google Scholar] [CrossRef]
- EL-Raey, M.; Badr, M.; Rawash, Z.; Darwish, G.M. Evidences for the Role of Melatonin as a Protective Additive during Buffalo Semen Freezing. Am. J. Anim. Vet. Sci. 2014, 9, 252–262. [Google Scholar] [CrossRef][Green Version]
- Chaudhary, S.C.; Aeksiri, N.; Liao, Y.-J.; Inyawilert, W. Effects of Melatonin on Cryopreserved Semen Parameters and Apoptosis of Thai Swamp Buffalo Bull (Bubalus bubalis) in Different Thawing Conditions. Adv. Anim. Vet. Sci. 2021, 9, 238–245. [Google Scholar] [CrossRef]
- Su, G.; Wu, S.; Wu, M.; Wang, L.; Yang, L.; Du, M.; Zhao, X.; Su, X.; Liu, X.; Bai, C.; et al. Melatonin improves the quality of frozen bull semen and influences gene expression related to embryo genome activation. Theriogenology 2021, 176, 54–62. [Google Scholar] [CrossRef]
- Pool, K.R.; Rickard, J.P.; de Graaf, S.P. Melatonin improves the motility and DNA integrity of frozen-thawed ram spermatozoa likely via suppression of mitochondrial superoxide production. Domest. Anim. Endocrinol. 2021, 74, 106516. [Google Scholar] [CrossRef]
- Deng, S.L.; Sun, T.C.; Yu, K.; Wang, Z.P.; Zhang, B.L.; Zhang, Y.; Wang, X.X.; Lian, Z.X.; Liu, Y.X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Najafi, A.; Adutwum, E.; Yari, A.; Salehi, E.; Mikaeili, S.; Dashtestani, F.; Abolhassani, F.; Rashki, L.; Shiasi, S.; Asadi, E. Melatonin affects membrane integrity, intracellular reactive oxygen species, caspase3 activity and AKT phosphorylation in frozen thawed human sperm. Cell Tissue Res. 2018, 372, 149–159. [Google Scholar] [CrossRef] [PubMed]
- de Lima Assis, I.; Palhares, P.C.; Machado, G.J.; da Silva Souza, J.G.; de Souza França, T.; de Oliveira Felizardo, V.; Murgas, L.D.S. Effect of Melatonin on Cryopreserved Sperm of Prochilodus lineatus (Characiformes). CryoLetters 2019, 40, 152–158. [Google Scholar] [PubMed]
- Ferrão, M.L.P. The Role of Melatonin in Sperm from Two Aquaculture Fish Species with Reproductive Problems: Solea senegalensis and Anguilla anguilla. Master’s Thesis, University of Algarve, Faro, Portugal, 2020. [Google Scholar]
- Palhares, P.C.; Assis, I.L.; Machado, G.J.; de Freitas, R.M.P.; de Freitas, M.B.D.; Paula, D.A.J.; Carneiro, W.F.; Motta, N.C.; Murgas, L.D.S. Sperm characteristics, peroxidation lipid and antioxidant enzyme activity changes in milt of Brycon orbignyanus cryopreserved with melatonin in different freezing curves. Theriogenology 2021, 176, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Alevra, A.I.; Makris, A.; Kanaki, A.P.; Exadactylos, A.; Mente, E.; Mylonas, C.C.; Papadopoulos, S. The effects of melatonin in preserving Gilthead Seabream (Sparus aurata) spermatozoa: Preliminary results. In Proceedings of the Aquaculture Europe 2022, Rimini, Italy, 22–27 September 2022; pp. 34–35. [Google Scholar]
- Pezo, F.; Zambrano, F.; Uribe, P.; Moya, C.; de Andrade, A.F.C.; Risopatron, J.; Yeste, M.; Burgos, R.A.; Sánchez, R. Oxidative and nitrosative stress in frozen-thawed pig spermatozoa. I: Protective effect of melatonin and butylhydroxytoluene on sperm function. Res. Vet. Sci. 2021, 136, 143–150. [Google Scholar] [CrossRef]
- Monteiro, M.M.; Silva, R.A.J.A.; Arruda, L.C.P.; Oliveira, A.S.d.; Mergulhão, F.C.d.C.; Júnior, P.L.J.M.; Seal, D.C.d.M.; Trevisan, M.; Batista, A.M.; Guerra, M.M.P. Effect of melatonin in different extenders on the quality of frozen semen of goats. Emerg. Anim. Species 2022, 5, 100015. [Google Scholar] [CrossRef]
- Ashrafi, I.; Kohram, H.; Naijian, H.; Bahreini, M.; Poorhamdollah, M. 2011. Protective effect of melatonin on sperm motility parameters on liquid storage of ram semen at 5 °C. Afr. J. Biotechnol. 2011, 10, 6670–6674. [Google Scholar] [CrossRef]
- Rateb, S.A.; Khalifa, M.A.; Abd El-Hamid, I.S.; Shedeed, H.A. Enhancing Liquid-Chilled Storage and Cryopreservation Ca-pacities of Ram Spermatozoa by Supplementing the Diluent with Different Additives. Asian-Australas J. Anim. Sci. 2020, 33, 1068–1076. [Google Scholar] [CrossRef]
- Medrano, A.; Contreras, C.B.; Herrera, F.; Alcantar-Rodriguez, A. Melatonin as an Antioxidant Preserving Sperm from Domestic Animals. Asian Pac. J. Reprod. 2017, 6, 241. [Google Scholar] [CrossRef]
- El-Battawy, K. Preservation of goat semen at 5οC with emphasis on its freezability and the impact of melatonin. Int. J. Vet. Sci. Res. 2019, 5, 035–038. [Google Scholar] [CrossRef]
- Pool, K.R.; Rickard, J.P.; Tumeth, E.; de Graaf, S.P. Treatment of Rams with Melatonin Implants in the Non-Breeding Season Improves Post-Thaw Sperm Progressive Motility and DNA Integrity. Anim. Reprod. Sci. 2020, 221, 106579. [Google Scholar] [CrossRef] [PubMed]
- Maldjian, A.; Pizzi, F.; Gliozzi, T.; Cerolini, S.; Penny, P.; Noble, R. Changes in Sperm Quality and Lipid Composition during Cryopreservation of Boar Semen. Theriogenology 2005, 63, 411–421. [Google Scholar] [CrossRef]
- Martín-Hidalgo, D.; Barón, F.J.; Bragado, M.J.; Carmona, P.; Robina, A.; García-Marín, L.J.; Gil, M.C. The Effect of Melatonin on the Quality of Extended Boar Semen after Long-Term Storage at 17 °C. Theriogenology 2011, 75, 1550–1560. [Google Scholar] [CrossRef]
- ChaithraShree, A.R.; Ingole, S.D.; Dighe, V.D.; Nagvekar, A.S.; Bharucha, S.V.; Dagli, N.R.; Kekan, P.M.; Kharde, S.D. Effect of melatonin on bovine sperm characteristics and ultrastructure changes following cryopreservation. Vet. Med. Sci. 2020, 6, 177–186. [Google Scholar] [CrossRef]
- Luo, X.; Wu, D.; Liang, M.; Huang, S.; Shi, D.; Li, X. The effects of melatonin, glutathione and vitamin E on semen cryopreservation of Mediterranean buffalo. Theriogenology 2022, 197, 94–100. [Google Scholar] [CrossRef]
- Inyawilert, W.; Rungruangsak, J.; Liao, Y.J.; Tang, P.C.; Paungsukpaibool, V. Melatonin supplementation improved cryopreserved Thai swamp buffalo semen. Reprod. Domest. Anim. 2021, 56, 83–88. [Google Scholar] [CrossRef]
- Monteiro, K.S.; Motta, N.C.; Cardoso, A.C.P.; Souza, S.P.; Murgas, L.D.S. Melatonin Supplementation for the Cryopreservation of Canine Sperm. Biopreserv. Biobank. 2022. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin Inhibits Formation of Mitochondrial Permeability Transition Pores and Improves Oxidative Phosphorylation of Frozen-Thawed Ram Sperm. Front. Endocrinol. 2019, 10, 896. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, C.; Xiang, H.; Jia, G.; Zhong, R. Melatonin improves cryopreservation of ram sperm by inhibiting mitochondrial permeability transition pore opening. Reprod. Domest. Anim. 2020, 55, 1240–1249. [Google Scholar] [CrossRef]
- Motta, N.C.; Egger, R.C.; Monteiro, K.S.; Vogel de Oliveira, A.; Solis Murgas, L.D. Effects of melatonin supplementation on the quality of cryopreserved sperm in the neotropical fish Prochilodus lineatus. Theriogenology 2022, 179, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Palhares, P.C.; Assis, I.L.; Souza, J.; França, T.S.; Egger, R.C.; Paula, D.A.J.; Murgas, L.D.S. Effect of melatonin supplementation to a cytoprotective medium on post-thawed Brycon orbignyanus sperm quality preserved during different freezing times. Cryobiology 2020, 96, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Tosti, E. Effects of ecosystem stress on reproduction and development. Mol. Reprod. Dev. 2019, 86, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.M.; Viana, A.G.d.A.; Carvalho, R.P.R.; Waddington, B.; Machado-Neves, M. Could metal exposure affect sperm parameters of domestic ruminants? A meta-analysis. Anim. Reprod. Sci. 2022, 244, 107050. [Google Scholar] [CrossRef]
- Li, T.; Zheng, Y.; Li, T.; Guo, M.; Wu, X.; Liu, R.; Liu, Q.; You, X.; Zeng, W.; Lv, Y. Potential dual protective effects of melatonin on spermatogonia against hexavalent chromium. Reprod. Toxicol. 2022, 111, 92–105. [Google Scholar] [CrossRef]
- Sun, T.; Song, L.; Ma, J.; Yu, H.; Zhou, S.; Wang, S.; Tian, L. Melatonin and its protective role against male reproductive toxicity induced by heavy metals, environmental pollutants, and chemotherapy: A review. Biocell 2020, 44, 479–485. [Google Scholar] [CrossRef]
- Dehdari Ebrahimi, N.; Parsa, S.; Nozari, F.; Shahlaee, M.A.; Maktabi, A.; Sayadi, M.; Sadeghi, A.; Azarpira, N. Protective effects of melatonin against the toxic effects of environmental pollutants and heavy metals on testicular tissue: A systematic review and meta-analysis of animal studies. Front. Endocrinol. 2023, 14, 1119553. [Google Scholar] [CrossRef]
- Kumar, J.; Verma, R.; Haldar, C. Melatonin ameliorates Bisphenol S induced testicular damages by modulating Nrf-2/HO-1 and SIRT-1/FOXO-1 expressions. Environ. Toxicol. 2021, 36, 396–407. [Google Scholar] [CrossRef]
- Venditti, M.; Rhouma, M.B.; Romano, M.Z.; Messaoudi, I.; Reiter, R.J.; Minucci, S. Altered expression of daam1 and prep induced by cadmium toxicity is counteracted by melatonin in the rat testis. Genes 2021, 12, 1016. [Google Scholar] [CrossRef]
- Mukherjee, S.; Maitra, S.K. Effects of starvation, re-feeding and timing of food supply on daily rhythm features of gut melatonin in carp (Catla catla). Chronobiol. Int. 2015, 32, 1264–1277. [Google Scholar] [CrossRef]
- Mukherjee, S.; Maitra, S.K. Daily profiles of serum and gastrointestinal melatonin in response to daytime or night-time supply of tryptophan-rich diet in carp (Catla catla). Biol. Rhythm. Res. 2018, 49, 315–327. [Google Scholar] [CrossRef]
- Falcón, J.; Migaud, H.; Muñoz-Cueto, J.A.; Carrillo, M. Current knowledge on the melatonin system in teleost fish. Gen. Comp. Endocrinol. 2010, 165, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Seth, M.; Maitra, S.K. Importance of light in temporal organization of photoreceptor proteins and melatonin-producing system in the pineal of carp Catla catla. Chronobiol. Int. 2010, 27, 463–486. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.K.; Hasan, K.N.; Maitra, S.K. Gut melatonin response to microbial infection in carp Catla catla. Fish Physiol. Biochem. 2016, 42, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, F.; Sutradhar, S.; Das, P.; Mukherjee, S. Gut melatonin: A potent candidate in the diversified journey of melatonin research. Gen. Comp. Endocrinol. 2021, 303, 113693. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Burkhardt, S.; Manchester, L.C. Melatonin in plants. Nutr. Rev. 2001, 59, 286–290. [Google Scholar] [CrossRef]
- Benot, S.; Goberna, R.; Reiter, R.J.; Garcia-Mauriño, S.; Osuna, C.; Guerrero, J.M. Physiological levels of melatonin contribute to the antioxidant capacity of human serum. J. Pineal Res. 1999, 27, 59–64. [Google Scholar] [CrossRef]
- Peña-Delgado, V.; Carvajal-Serna, M.; Fondevila, M.; Martín-Cabrejas, M.A.; Aguilera, Y.; Álvarez-Rivera, G.; Abecia, J.A.; Casao, A.; Pérez-Pe, R. Improvement of the Seminal Characteristics in Rams Using Agri-Food By-Products Rich in Phytomelatonin. Animals 2023, 13, 905. [Google Scholar] [CrossRef]
- Exadactylos, A. Nutrigenomics in Aquaculture Research. Fish. Aquac. J. 2014, 5, 1. [Google Scholar] [CrossRef]
- Haq, Z.U.; Saleem, A.; Khan, A.A.; Dar, M.A.; Ganaie, A.M.; Beigh, Y.A.; Hamadani, H.; Ahmad, S.M. Nutrigenomics in livestock sector and its human-animal interface-a review. Vet. Anim. Sci. 2022, 17, 100262. [Google Scholar] [CrossRef] [PubMed]
- Loor, J.J. Nutrigenomics in livestock: Potential role in physiological regulation and practical applications. Anim. Prod. Sci. 2022, 62, 901–912. [Google Scholar] [CrossRef]
- Bromage, N.; Porter, M.; Randall, C. The environmental regulation of maturation in farmed finfish with special reference to the role of photoperiod and melatonin. Aquaculture 2001, 197, 63–98. [Google Scholar] [CrossRef]
- Félix, F.; Antunes, R.; Vera, L.M.; Oliveira, C.C.V.; Cabrita, E. The protective effect of endogenous melatonin on gilthead seabream sperm during cryopreservation. Aquaculture 2023, 577, 739997. [Google Scholar] [CrossRef]
- Félix, F.; Gallego, V.; Mendes, A.; Soares, F.; Vera, L.M.; Cabrita, E.; Oliveira, C.C.V. Novel approaches on melatonin role: Presence of clock-hormone in fish seminal plasma. Aquaculture 2023, 573, 739578. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).