
Modulation of Caecal Microbiota and Metabolome Profile in Salmonella-Infected Broilers by Phage Therapy
 ,
,  ,
,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
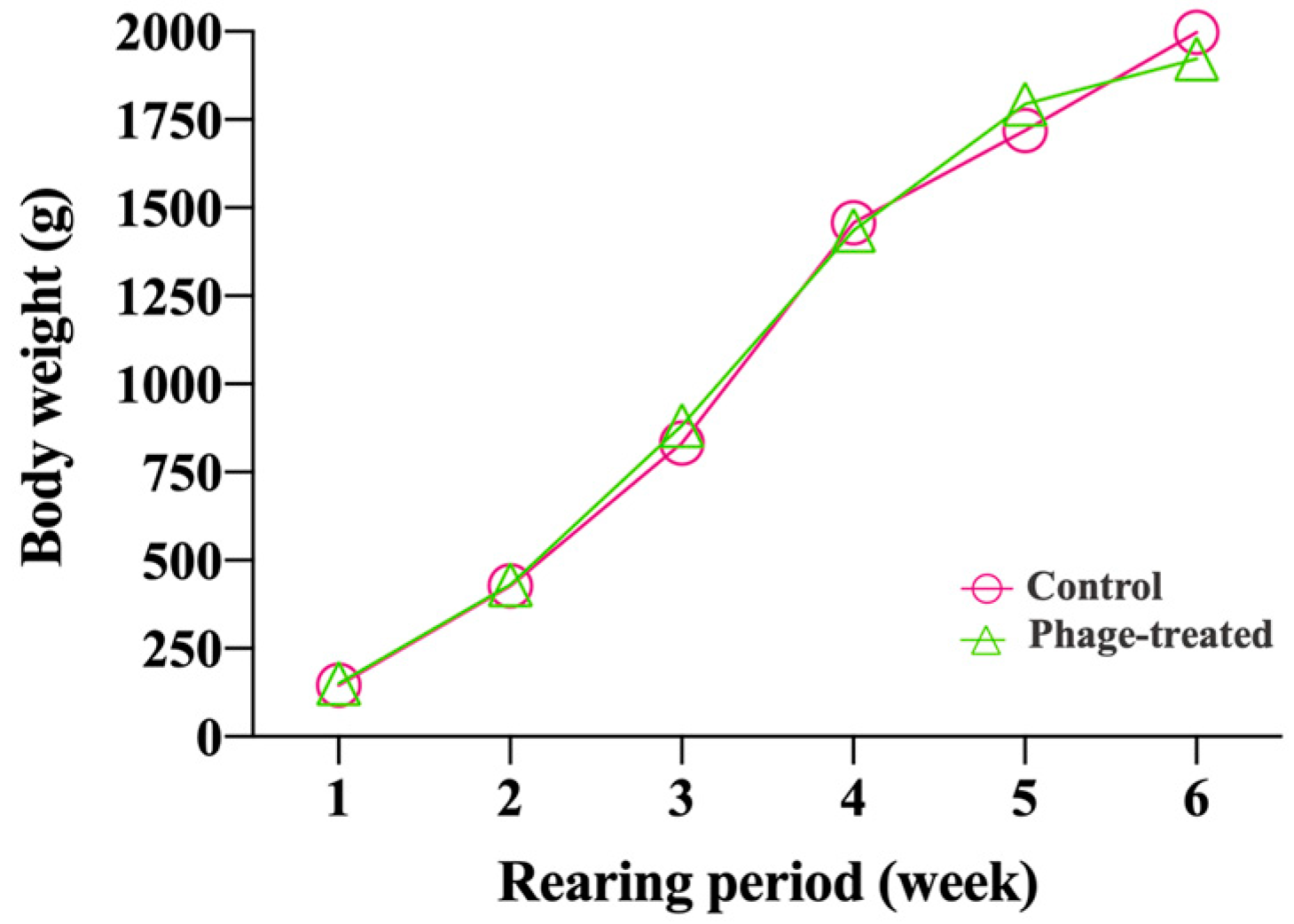
2.1. Effects of Phage on Body Weight
2.2. Effects of Phage on Caecal Microbiota
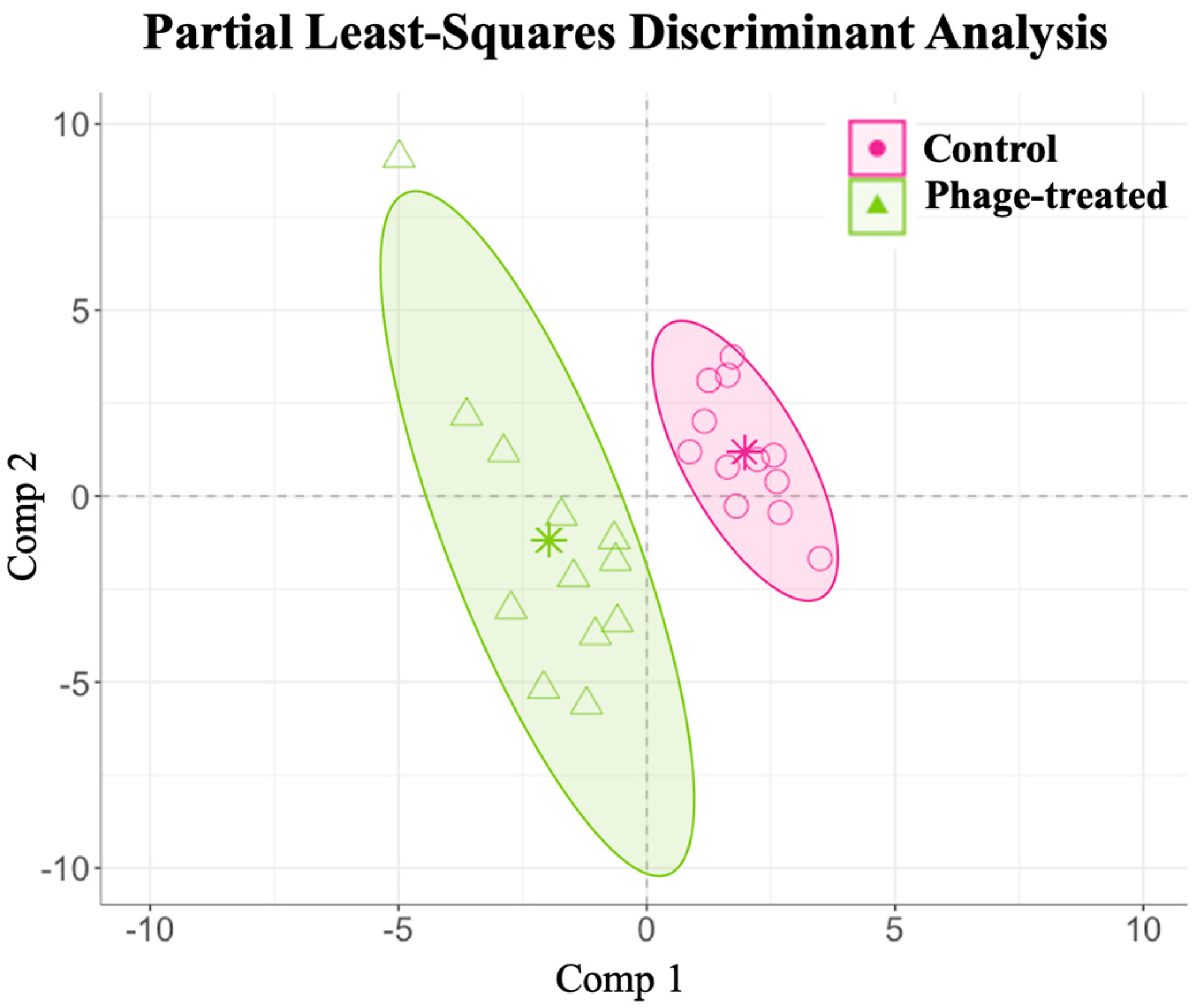
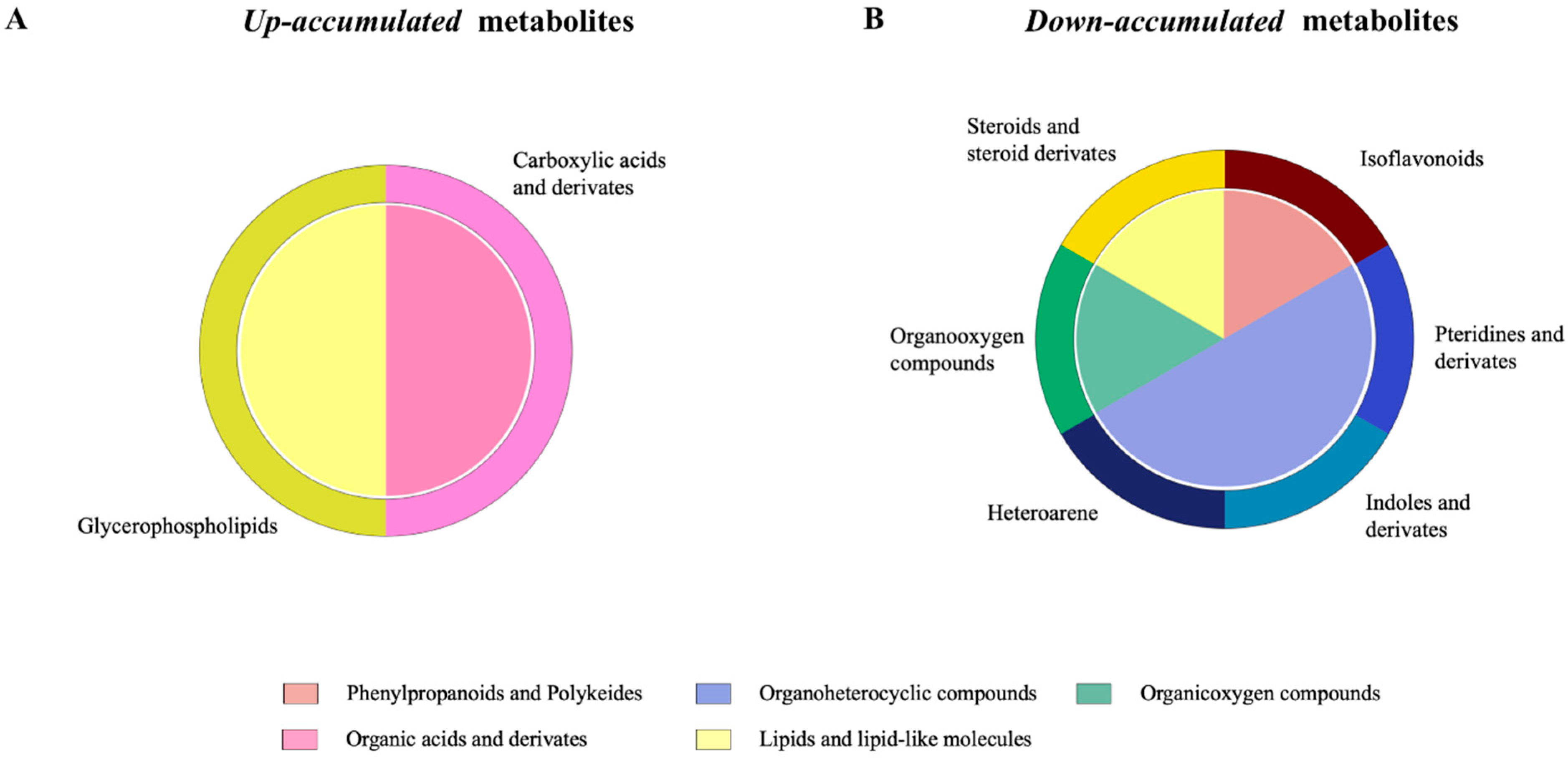
2.3. Effects of Phage on Gut Metabolome
2.4. Effects of Phage on Serum Metabolome
3. Discussion
4. Materials and Methods
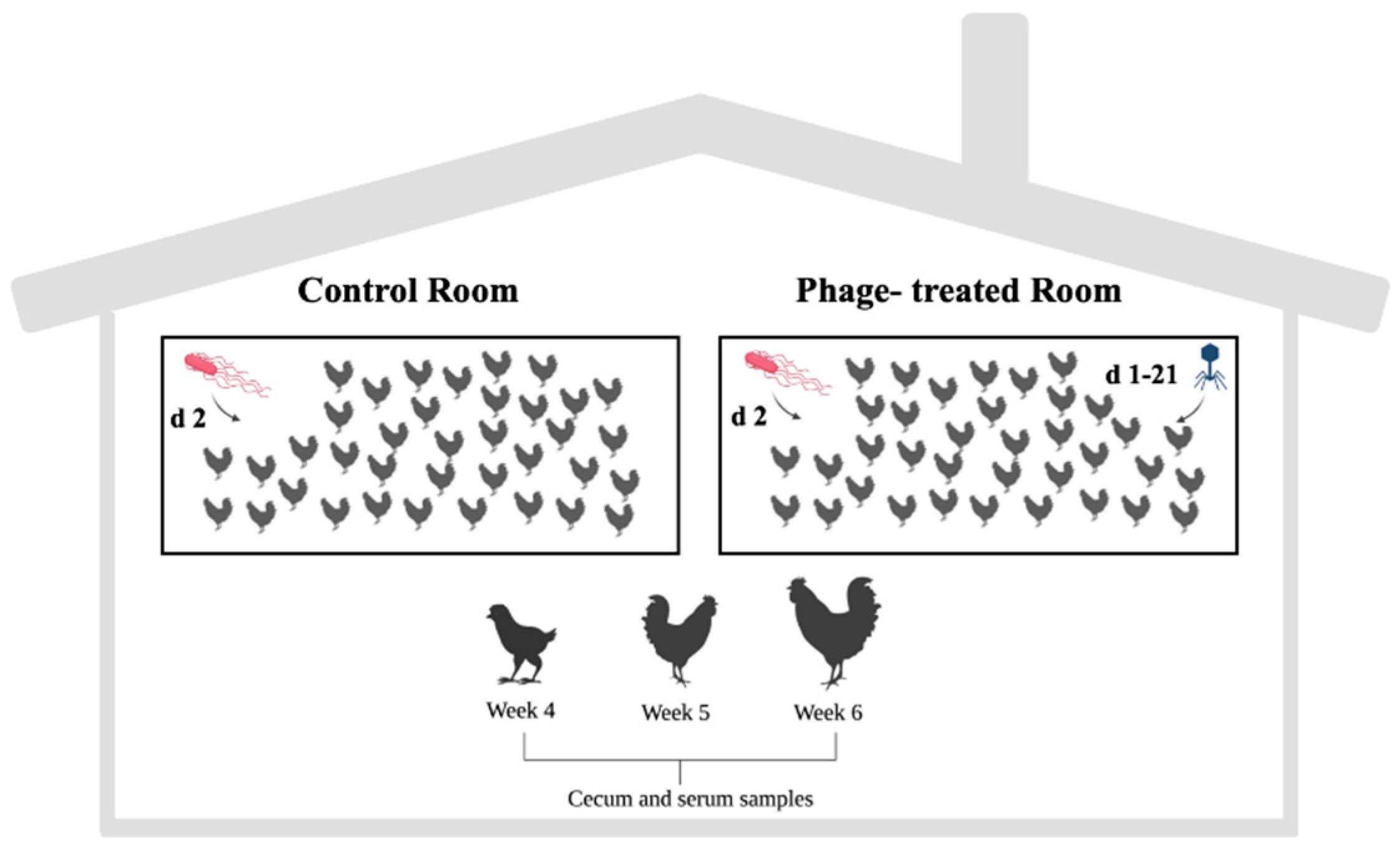
4.1. Caecal and Serum Content Origin
4.2. Microbiota Analysis
4.2.1. DNA Extraction, 16S rRNA Gene Amplification and MiSeq Sequencing
4.2.2. Bioinformatic Analysis
4.3. Metabolomics Analysis
4.3.1. Sample Preparations
4.3.2. LC–ESI–HRMS Analysis
4.4. Statistical Analysis
4.4.1. Body Weight Statistical Analysis
4.4.2. Caecal Microbiome and Caecal and Serum Metabolome Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tzani, M.; Mandilara, G.; Dias, J.G.; Sideroglou, T.; Chrysostomou, A.; Mellou, K. Impact of Salmonella Control Programmes in Poultry on Human Salmonellosis Burden in Greece. Antibiotics 2021, 10, 121. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Abd El-Mageed, T.A.; Soliman, S.M.; Khafaga, A.F.; Swelum, A.A.; Ahmed, A.E.; Alshammari, F.A.; Abd El-Hack, M.E. The Control of Poultry Salmonellosis Using Organic Agents: An Updated Overview. Poult. Sci. 2022, 101, 101716. [Google Scholar] [CrossRef]
- EFSA. ECDC The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Marin, C.; Cerdà-Cuéllar, M.; González-Bodi, S.; Lorenzo-Rebenaque, L.; Vega, S. Research Note: Persistent Salmonella Problems in Slaughterhouses Related to Clones Linked to Poultry Companies. Poult. Sci. 2022, 101, 101968. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Taha, A.E.; Soliman, S.M.; Ahmed, A.E.; El-kott, A.F.; et al. Alternatives to Antibiotics for Organic Poultry Production: Types, Modes of Action and Impacts on Bird’s Health and Production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef]
- Ayalew, H.; Zhang, H.; Wang, J.; Wu, S.; Qiu, K.; Qi, G.; Tekeste, A.; Wassie, T.; Chanie, D. Potential Feed Additives as Antibiotic Alternatives in Broiler Production. Front. Vet. Sci. 2022, 9, 916473. [Google Scholar] [CrossRef]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage Therapy to Combat Bacterial Infections in Poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Ruvalcaba-Gómez, J.M.; Villagrán, Z.; Valdez-Alarcón, J.J.; Martínez-Núñez, M.; Gomez-Godínez, L.J.; Ruesga-Gutiérrez, E.; Anaya-Esparza, L.M.; Arteaga-Garibay, R.I.; Villarruel-López, A. Non-Antibiotics Strategies to Control Salmonella Infection in Poultry. Animals 2022, 12, 102. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Y.; Lv, P.; Huang, J.; Tai, R.; Jin, X.; Wang, J.; Wang, X. Salmonella Phages Affect the Intestinal Barrier in Chicks by Altering the Composition of Early Intestinal Flora: Association with Time of Phage Use. Front. Microbiol. 2022, 13, 947640. [Google Scholar] [CrossRef]
- Clavijo, V.; Morales, T.; Vives-Flores, M.J.; Reyes Muñoz, A. The Gut Microbiota of Chickens in a Commercial Farm Treated with a Salmonella Phage Cocktail. Sci. Rep. 2022, 12, 991. [Google Scholar] [CrossRef]
- Cieplak, T.; Soffer, N.; Sulakvelidze, A.; Sandris Nielsen, D. Gut Microbes A Bacteriophage Cocktail Targeting Escherichia Coli Reduces E. Coli in Simulated Gut Conditions, While Preserving a Non-Targeted Representative Commensal Normal Microbiota. Gut Microbes 2018, 9, 391–399. [Google Scholar] [CrossRef]
- Gindin, M.; Febvre, H.P.; Rao, S.; Wallace, T.C.; Weir, T.L. Bacteriophage for Gastrointestinal Health (PHAGE) Study: Evaluating the Safety and Tolerability of Supplemental Bacteriophage Consumption. J. Am. Coll. Nutr. 2019, 38, 68–75. [Google Scholar] [CrossRef]
- Andreatti Filho, R.L.; Higgins, J.P.; Higgins, S.E.; Gaona, G.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Ability of Bacteriophages Isolated from Different Sources to Reduce Salmonella Enterica Serovar Enteritidis in Vitro and in Vivo. Poult. Sci. 2007, 86, 1904–1909. [Google Scholar] [CrossRef] [PubMed]
- Borie, C.; Sánchez, M.L.; Navarro, C.; Ramírez, S.; Morales, M.A.; Retamales, J.; Robeson, J. El Tratamiento Por Aerosol Con Bacteriófagos y Exclusión Competitiva Reduce La Infección Con Salmonella Enteritidis En Pollos. Avian Dis. 2009, 53, 250–254. [Google Scholar] [CrossRef]
- Lim, T.H.; Kim, M.S.; Lee, D.H.; Lee, Y.N.; Park, J.K.; Youn, H.N.; Lee, H.J.; Yang, S.Y.; Cho, Y.W.; Lee, J.B.; et al. Use of Bacteriophage for Biological Control of Salmonella Enteritidis Infection in Chicken. Res. Vet. Sci. 2012, 93, 1173–1178. [Google Scholar] [CrossRef]
- Grabowski, Ł.; Węgrzyn, G.; Węgrzyn, A.; Podlacha, M. Highly Different Effects of Phage Therapy and Antibiotic Therapy on Immunological Responses of Chickens Infected with Salmonella Enterica Serovar Typhimurium. Front. Immunol. 2022, 13, 956833. [Google Scholar] [CrossRef]
- Lorenzo-Rebenaque, L.; Malik, D.J.; Catalá-Gregori, P.; Torres, J.; Marin, C.; Sevilla-Navarro, S. Microencapsulated Bacteriophages Incorporated in Feed for Salmonella Control in Broilers. Vet. Microbiol. 2022, 274, 109579. [Google Scholar] [CrossRef] [PubMed]
- Peh, E.; Szott, V.; Reichelt, B.; Friese, A.; Rösler, U.; Plötz, M.; Kittler, S. Bacteriophage Cocktail Application for Campylobacter Mitigation—from in Vitro to in Vivo. BMC Microbiol. 2023, 23, 209. [Google Scholar] [CrossRef]
- Sarrami, Z.; Sedghi, M.; Mohammadi, I.; Bedford, M.; Miranzadeh, H.; Ghasemi, R. Effects of Bacteriophage on Salmonella Enteritidis Infection in Broilers. Sci. Rep. 2023, 13, 12198. [Google Scholar] [CrossRef] [PubMed]
- Thanki, A.M.; Hooton, S.; Whenham, N.; Salter, M.G.; Bedford, M.R.; O’Neill, H.V.M.; Clokie, M.R.J. A Bacteriophage Cocktail Delivered in Feed Significantly Reduced Salmonella Colonization in Challenged Broiler Chickens. Emerg. Microbes Infect. 2023, 12, 2217947. [Google Scholar] [CrossRef] [PubMed]
- Baümler, A.J.; Sperandio, V. Interactions between the Microbiota and Pathogenic Bacteria in the Gut. Nature 2016, 535, 85. [Google Scholar] [CrossRef]
- Kosznik-Kwaśnicka, K.; Podlacha, M.; Grabowski, Ł.; Stasiłojć, M.; Nowak-Zaleska, A.; Ciemińska, K.; Cyske, Z.; Dydecka, A.; Gaffke, L.; Mantej, J.; et al. Biological Aspects of Phage Therapy versus Antibiotics against Salmonella Enterica Serovar Typhimurium Infection of Chickens. Front. Cell Infect. Microbiol. 2022, 12, 941867. [Google Scholar] [CrossRef]
- Chen, M.X.; Wang, S.Y.; Kuo, C.H.; Tsai, I.L. Metabolome Analysis for Investigating Host-Gut Microbiota Interactions. J. Formos. Med. Assoc. 2019, 118, S10–S22. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.Z.; Chen, G.; Hong, Q.; Huang, S.; Smith, H.M.; Shah, R.D.; Scholz, M.; Ferguson, J.F. Multi-Omic Analysis of the Microbiome and Metabolome in Healthy Subjects Reveals Microbiome-Dependent Relationships between Diet and Metabolites. Front. Genet. 2019, 10, 454. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.F.; Zhou, B.; Ressom, H.W. Metabolite Identification and Quantitation in LC-MS/MS-Based Metabolomics. TrAC—Trends Anal. Chem. 2012, 32, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zhou, L.; Zhang, Y.; Xue, S.; Gan, Q.F.; Fang, S. Effect of Host Breeds on Gut Microbiome and Serum Metabolome in Meat Rabbits. BMC Vet. Res. 2021, 17, 24. [Google Scholar] [CrossRef]
- Grzymajlo, K. The Game for Three: Salmonella–Host–Microbiota Interaction Models. Front. Microbiol. 2022, 13, 1082. [Google Scholar] [CrossRef]
- Johnson, K.V.A.; Burnet, P.W.J. Microbiome: Should We Diversify from Diversity? Gut Microbes 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Jeong, J.; Lee, J.; Kim, S.; Min, W.; Myung, H. Therapeutic Effects of Bacteriophages against Salmonella Gallinarum Infection in Chickens. J. Microbiol. Biotechnol. 2013, 23, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Amir Karimi Torshizi, M.; Rahimi, S.; Dennehy, J.J. Prophylactic Bacteriophage Administration More Effective than Post-Infection Administration in Reducing Salmonella Enterica Serovar Enteritidis Shedding in Quail. Front. Microbiol. 2016, 7, 1253. [Google Scholar] [CrossRef]
- Adhikari, P.A.; Cosby, D.E.; Cox, N.A.; Lee, J.H.; Kim, W.K. Effect of Dietary Bacteriophage Supplementation on Internal Organs, Fecal Excretion, and Ileal Immune Response in Laying Hens Challenged by Salmonella Enteritidis. Poult. Sci. 2017, 96, 3264–3271. [Google Scholar] [CrossRef]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the Impact of Bacteriophages in the Treatment of Salmonella in Broiler Chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef]
- Vaz, C.S.L.; Voss-Rech, D.; Alves, L.; Coldebella, A.; Brentano, L.; Trevisol, I.M. Effect of Time of Therapy with Wild-Type Lytic Bacteriophages on the Reduction of Salmonella Enteritidis in Broiler Chickens. Vet. Microbiol. 2020, 240, 108527. [Google Scholar] [CrossRef] [PubMed]
- Sorour, H.K.; Gaber, A.F.; Hosny, R.A. Evaluation of the Efficiency of Using Salmonella Kentucky and Escherichia Coli O119 Bacteriophages in the Treatment and Prevention of Salmonellosis and Colibacillosis in Broiler Chickens. Lett. Appl. Microbiol. 2020, 71, 345–350. [Google Scholar] [CrossRef]
- Sausset, R.; Petit, M.A.; Gaboriau-Routhiau, V.; de Paepe, M. New Insights into Intestinal Phages. Mucosal Immunol. 2020, 13, 205–215. [Google Scholar] [CrossRef]
- Galtier, M.; de Sordi, L.; Maura, D.; Arachchi, H.; Volant, S.; Dillies, M.A.; Debarbieux, L. Bacteriophages to Reduce Gut Carriage of Antibiotic Resistant Uropathogens with Low Impact on Microbiota Composition. Environ. Microbiol. 2016, 18, 2237–2245. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.V.; Ruggles, K.V.; Zhou, H.; Heguy, A.; Tsirigos, A.; Tetz, V. Bacteriophages as Potential New Mammalian Pathogens. Sci. Rep. 2017, 7, 7043. [Google Scholar] [CrossRef]
- Dissanayake, U.; Ukhanova, M.; Moye, Z.D.; Sulakvelidze, A.; Mai, V. Bacteriophages Reduce Pathogenic Escherichia Coli Counts in Mice Without Distorting Gut Microbiota. Front. Microbiol. 2019, 10, 1984. [Google Scholar] [CrossRef]
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic Modulation of the Gut Microbiota and Metabolome by Bacteriophages in a Mouse Model. Cell Host Microbe 2019, 25, 803–814.e5. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Rebenaque, L.; Casto-Rebollo, C.; Diretto, G.; Frusciante, S.; Rodríguez, J.C.; Ventero, M.-P.; Molina-Pardines, C.; Vega, S.; Marin, C.; Marco-Jiménez, F. Examining the Effects of Salmonella Phage on the Caecal Microbiota and Metabolome Features in Salmonella-Free Broilers. Front. Genet. 2022, 13, 1060713. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, Q.; Huang, X.; Jiang, G.; Li, C.; Zhang, X.; Liu, S.; He, L.; Liu, Y.; Dai, Q.; et al. Effects of Dietary Ferulic Acid on the Intestinal Microbiota and the Associated Changes on the Growth Performance, Serum Cytokine Profile, and Intestinal Morphology in Ducks. Front. Microbiol. 2021, 12, 1800. [Google Scholar] [CrossRef]
- Sarker, S.A.; McCallin, S.; Barretto, C.; Berger, B.; Pittet, A.C.; Sultana, S.; Krause, L.; Huq, S.; Bibiloni, R.; Bruttin, A.; et al. Oral T4-like Phage Cocktail Application to Healthy Adult Volunteers from Bangladesh. Virology 2012, 434, 222–232. [Google Scholar] [CrossRef]
- Sarker, S.A.; Berger, B.; Deng, Y.; Kieser, S.; Foata, F.; Moine, D.; Descombes, P.; Sultana, S.; Huq, S.; Bardhan, P.K.; et al. Oral Application of Escherichia Coli Bacteriophage: Safety Tests in Healthy and Diarrheal Children from Bangladesh. Environ. Microbiol. 2017, 19, 237–250. [Google Scholar] [CrossRef]
- McCallin, S.; Alam Sarker, S.; Barretto, C.; Sultana, S.; Berger, B.; Huq, S.; Krause, L.; Bibiloni, R.; Schmitt, B.; Reuteler, G.; et al. Safety Analysis of a Russian Phage Cocktail: From Metagenomic Analysis to Oral Application in Healthy Human Subjects. Virology 2013, 443, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea with Two Coliphage Preparations: A Randomized Trial in Children from Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef]
- Rubinelli, P.M.; Kim, S.A.; Park, S.H.; Roto, S.M.; Nealon, N.J.; Ryan, E.P.; Ricke, S.C. Differential Effects of Rice Bran Cultivars to Limit Salmonella Typhimurium in Chicken Cecal in Vitro Incubations and Impact on the Cecal Microbiome and Metabolome. PLoS ONE 2017, 12, e0185002. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.D.; Ipharraguerre, I.R.; Arsenault, R.J.; Lyte, M.; Lyte, J.M.; Humphrey, B.; Angel, R.; Korver, D.R. Informal Nutrition Symposium: Leveraging the Microbiome (and the Metabolome) for Poultry Production. Poult. Sci. 2022, 101, 101588. [Google Scholar] [CrossRef]
- Aljahdali, N.H.; Sanad, Y.M.; Han, J.; Foley, S.L. Current Knowledge and Perspectives of Potential Impacts of Salmonella Enterica on the Profile of the Gut Microbiota. BMC Microbiol. 2020, 20, 101588. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chousalkar, K.K. Salmonella Typhimurium Infection Disrupts but Continuous Feeding of Bacillus Based Probiotic Restores Gut Microbiota in Infected Hens. J. Anim. Sci. Biotechnol. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Gao, H.; Qin, W.; Song, P.; Wang, H.; Zhang, J.; Liu, D.; Wang, D.; Zhang, T. Marked Seasonal Variation in Structure and Function of Gut Microbiota in Forest and Alpine Musk Deer. Front. Microbiol. 2021, 12, 699797. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, H.; Sun, G.; Duan, Y.; Guo, Y.; Xie, L.; Ding, X. Alterations in the Gut Microbiome and Metabolome Profiles of Septic Rats Treated with Aminophylline. J. Transl. Med. 2022, 20, 69. [Google Scholar] [CrossRef]
- Strnad, P.; Tacke, F.; Koch, A.; Trautwein, C. Liver-Guardian, Modifier and Target of Sepsis. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Memon, F.U.; Yang, Y.; Zhang, G.; Leghari, I.H.; Lv, F.; Wang, Y.; Laghari, F.; Khushk, F.A.; Si, H. Chicken Gut Microbiota Responses to Dietary Bacillus Subtilis Probiotic in the Presence and Absence of Eimeria Infection. Microorganisms 2022, 10, 1548. [Google Scholar] [CrossRef]
- Gilroy, R.; Ravi, A.; Getino, M.; Pursley, I.; Horton, D.L.; Alikhan, N.F.; Baker, D.; Gharbi, K.; Hall, N.; Watson, M.; et al. Extensive Microbial Diversity within the Chicken Gut Microbiome Revealed by Metagenomics and Culture. PeerJ 2021, 9, e10941. [Google Scholar] [CrossRef]
- Tenea, G.N.; Lara, M.I. Antimicrobial Compounds Produced by Weissella Confusa Cys2-2 Strain Inhibit Gram-Negative Bacteria Growth. CYTA J. Food 2019, 17, 105–111. [Google Scholar] [CrossRef]
- Cupi, D.; Elvig-Jørgensen, S.G. Safety Assessment of Weissella Confusa—A Direct-Fed Microbial Candidate. Regul. Toxicol. Pharmacol. 2019, 107, 104414. [Google Scholar] [CrossRef] [PubMed]
- O’Cuív, P.; Klaassens, E.S.; Durkin, A.S.; Harkins, D.M.; Foster, L.; McCorrison, J.; Torralba, M.; Nelson, K.E.; Morrison, M. Draft Genome Sequence of Turicibacter Sanguinis PC909, Isolated from Human Feces. J. Bacteriol. 2011, 193, 1288–1289. [Google Scholar] [CrossRef]
- Zhou, Q.; Lan, F.; Li, X.; Yan, W.; Sun, C.; Li, J.; Yang, N.; Wen, C. The Spatial and Temporal Characterization of Gut Microbiota in Broilers. Front. Vet. Sci. 2021, 8, 712226. [Google Scholar] [CrossRef]
- Baniel, A.; Amato, K.R.; Beehner, J.C.; Bergman, T.J.; Mercer, A.; Perlman, R.F.; Petrullo, L.; Reitsema, L.; Sams, S.; Lu, A.; et al. Seasonal Shifts in the Gut Microbiome Indicate Plastic Responses to Diet in Wild Geladas. Microbiome 2021, 9, 26. [Google Scholar] [CrossRef]
- Zhang, S.; Zhong, G.; Shao, D.; Wang, Q.; Hu, Y.; Wu, T.; Ji, C.; Shi, S. Dietary Supplementation with Bacillus Subtilis Promotes Growth Performance of Broilers by Altering the Dominant Microbial Community. Poult. Sci. 2021, 100, 100935. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Zheng, H.M.; Zhang, G.X.; Chen, F.L.; Chen, L.D.; Yang, Z.C. High Oscillospira Abundance Indicates Constipation and Low BMI in the Guangdong Gut Microbiome Project. Sci. Rep. 2020, 10, 9364. [Google Scholar] [CrossRef]
- Reyer, H.; Sjöberg, P.J.R.; Oster, M.; Wubuli, A.; Murani, E.; Ponsuksili, S.; Wolf, P.; Wimmers, K. Mineral Phosphorus Supply in Piglets Impacts the Microbial Composition and Phytate Utilization in the Large Intestine. Microorganisms 2021, 9, 1197. [Google Scholar] [CrossRef]
- Mon, K.K.Z.; Saelao, P.; Halstead, M.M.; Chanthavixay, G.; Chang, H.C.; Garas, L.; Maga, E.A.; Zhou, H. Salmonella Enterica Serovars Enteritidis Infection Alters the Indigenous Microbiota Diversity in Young Layer Chicks. Front. Vet. Sci. 2015, 2, 61. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, X.; Ma, X.; Geng, S.; Jiang, X.; Huang, Q.; Hu, C.; Han, X. Intestinal Microbiome-Metabolome Responses to Essential Oils in Piglets. Front. Microbiol. 2018, 9, 1988. [Google Scholar] [CrossRef] [PubMed]
- Cazals, A.; Estellé, J.; Bruneau, N.; Coville, J.L.; Menanteau, P.; Rossignol, M.N.; Jardet, D.; Bevilacqua, C.; Rau, A.; Bed’Hom, B.; et al. Differences in Caecal Microbiota Composition and Salmonella Carriage between Experimentally Infected Inbred Lines of Chickens. Genet. Sel. Evol. 2022, 54, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lilburn, M.; Yu, Z. Intestinal Microbiota of Broiler Chickens as Affected by Litter Management Regimens. Front. Microbiol. 2016, 7, 593. [Google Scholar] [CrossRef]
- Kumar, S.; Adhikari, P.; Oakley, B.; Kim, W.K. Changes in Cecum Microbial Community in Response to Total Sulfur Amino Acid (TSAA: DL-Methionine) in Antibiotic-Free and Supplemented Poultry Birds. Poult. Sci. 2019, 98, 5809–5819. [Google Scholar] [CrossRef]
- Bindari, Y.R.; Gerber, P.F. Centennial Review: Factors Affecting the Chicken Gastrointestinal Microbial Composition and Their Association with Gut Health and Productive Performance. Poult. Sci. 2022, 101, 101612. [Google Scholar] [CrossRef]
- Calik, A.; Ergün, A. Effect of Lactulose Supplementation on Growth Performance, Intestinal Histomorphology, Cecal Microbial Population, and Short-Chain Fatty Acid Composition of Broiler Chickens. Poult. Sci. 2015, 94, 2173–2182. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current Perspectives of the Chicken Gastrointestinal Tract and Its Microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Flórez, M.J.V. The Gastrointestinal Microbiome and Its Association with the Control of Pathogens in Broiler Chicken Production: A Review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Yang, D.; Zhao, D.; Shah, S.Z.A.; Wu, W.; Lai, M.; Zhang, X.; Li, J.; Guan, Z.; Zhao, H.; Li, W.; et al. Implications of Gut Microbiota Dysbiosis and Metabolic Changes in Prion Disease. Neurobiol. Dis. 2020, 135, 104704. [Google Scholar] [CrossRef]
- Carroll-Portillo, A.; Lin, H.C. Bacteriophage and the Innate Immune System: Access and Signaling. Microorganisms 2019, 7, 625. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Maurice, C.F. Bacteriophages: Uncharacterized and Dynamic Regulators of the Immune System. Mediators Inflamm. 2019, 2019, 3730519. [Google Scholar] [CrossRef]
- van Belleghem, J.D.; Dąbrowska, K.; Vaneechoutte, M.; Barr, J.J.; Bollyky, P.L. Interactions between Bacteriophage, Bacteria, and the Mammalian Immune System. Viruses 2019, 11, 10. [Google Scholar] [CrossRef]
- Nguyen, S.; Baker, K.; Padman, B.S.; Patwa, R.; Dunstan, R.A.; Weston, T.A.; Schlosser, K.; Bailey, B.; Lithgow, T.; Lazarou, M.; et al. Bacteriophage Transcytosis Provides a Mechanism to Cross Epithelial Cell Layers. mBio 2017, 8, 10–1128. [Google Scholar] [CrossRef]
- Aldars-García, L.; Gisbert, J.P.; Chaparro, M. Metabolomics Insights into Inflammatory Bowel Disease: A Comprehensive Review. Pharmaceuticals 2021, 14, 1190. [Google Scholar] [CrossRef]
- Vila, A.; Hu, S.; Andreu-Sánchez, S.; Collij, V.; Jansen, B.H.; Augustijn, H.E.; Bolte, L.; AAA Ruigrok, R.; Abu-Ali, G.; Giallourakis, C.; et al. Faecal metabolome and its determinants in inflammatory bowel disease. Gut 2023, 72, 1472–1485. [Google Scholar] [CrossRef]
- Ma, F.; Luo, L.; Gao, X. Metabolite and Transcriptome Analyses Revealed the Modulation of Fructo-Oligosaccharide on Ileum Metabolism of Taiping Chickens. J. Appl. Microbiol. 2022, 132, 2249–2261. [Google Scholar] [CrossRef] [PubMed]
- Fobofou, S.A.; Savidge, T. Microbial Metabolites: Cause or Consequence in Gastrointestinal Disease? Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G535–G552. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; McConn, B.R.; Liu, D.; Cline, M.A.; Gilbert, E.R. The Effects of Dietary Macronutrient Composition on Lipid Metabolism-Associated Factor Gene Expression in the Adipose Tissue of Chickens Are Influenced by Fasting and Refeeding. BMC Obes. 2017, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Noor, M.; Yasmin Runa, N.; Husna, A.; Rahman, M.; Rajib, D.M.; Mahbub-e-Elahi, A.; Masudur Rahman, M. Evaluation of the Effect of Dietary Supplementation of Bacteriophage on Production Performance and Excreta Microflora of Commercial Broiler and Layer Chickens in Bangladesh. MOJ Proteom. Bioinform. 2020, 9, 27–31. [Google Scholar] [CrossRef]
- Sarrami, Z.; Sedghi, M.; Mohammadi, I.; Kim, W.K.; Mahdavi, A.H. Effects of Bacteriophage Supplement on the Growth Performance, Microbial Population, and PGC-1α and TLR4 Gene Expressions of Broiler Chickens. Sci. Rep. 2022, 12, 14391. [Google Scholar] [CrossRef] [PubMed]
- Sevilla-Navarro, S.; Catalá-Gregori, P.; Marin, C. Salmonella Bacteriophage Diversity According to Most Prevalent Salmonella Serovars in Layer and Broiler Poultry Farms from Eastern Spain. Animals 2020, 10, 1456. [Google Scholar] [CrossRef]
- Lorenzo-Rebenaque, L.; Malik, D.J.; Catalá-Gregori, P.; Marin, C.; Sevilla-Navarro, S. In Vitro and In Vivo Gastrointestinal Survival of Non-Encapsulated and Microencapsulated Salmonella Bacteriophages: Implications for Bacteriophage Therapy in Poultry. Pharmaceuticals 2021, 14, 434. [Google Scholar] [CrossRef]
- Malik, D.J. Bacteriophage Encapsulation Using Spray Drying for Phage Therapy. Curr. Issues Mol. Biol. 2021, 40, 303–316. [Google Scholar] [CrossRef]
- Ijaz, U.Z.; Sivaloganathan, L.; McKenna, A.; Richmond, A.; Kelly, C.; Linton, M.; Stratakos, A.C.; Lavery, U.; Elmi, A.; Wren, B.W.; et al. Comprehensive Longitudinal Microbiome Analysis of the Chicken Cecum Reveals a Shift from Competitive to Environmental Drivers and a Window of Opportunity for Campylobacter. Front. Microbiol. 2018, 9, 2452. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Babraham Bioinformatics FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 13 September 2022).
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Campos, P.M.; Darwish, N.; Shao, J.; Proszkowiec-Weglarz, M. Research Note: Choice of Microbiota Database Affects Data Analysis and Interpretation in Chicken Cecal Microbiota. Poult. Sci. 2022, 101, 101971. [Google Scholar] [CrossRef] [PubMed]
- Grosso, V.; Farina, A.; Giorgi, D.; Nardi, L.; Diretto, G.; Lucretti, S. A High-Throughput Flow Cytometry System for Early Screening of in Vitro Made Polyploids in Dendrobium Hybrids. Plant Cell Tissue Organ. Cult. 2018, 132, 57–70. [Google Scholar] [CrossRef]
- Garcia-Dominguez, X.; Diretto, G.; Frusciante, S.; Vicente, J.S.; Marco-Jiménez, F. Metabolomic Analysis Reveals Changes in Preimplantation Embryos Following Fresh or Vitrified Transfer. Int. J. Mol. Sci. 2020, 21, 7116. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Tan, P.; Zhuang, Z.; Wang, Z.; Zhu, L.; Qiu, R.; Xu, H. Untargeted Metabolomic Approach to Study the Serum Metabolites in Women with Polycystic Ovary Syndrome. BMC Med. Genom. 2021, 14, 206. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; van Ommen, B.; Smilde, A.K. Large-Scale Human Metabolomics Studies: A Strategy for Data (Pre-) Processing and Validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef]
- Greenacre, M.; Martínez-Álvaro, M.; Blasco, A. Compositional Data Analysis of Microbiome and Any-Omics Datasets: A Validation of the Additive Logratio Transformation. Front. Microbiol. 2021, 12, 2625. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.-A.L. MixOmics: An R Package for ’omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Galindo-Prieto, B.; Eriksson, L.; Trygg, J. Variable Influence on Projection (VIP) for Orthogonal Projections to Latent Structures (OPLS). J. Chemom. 2014, 28, 623–632. [Google Scholar] [CrossRef]
- Blasco, A. Bayesian Data Analysis for Animal Scientists: The Basics. In Bayesian Data Analysis for Animal Scientists: The Basics; Springer: New York, NY, USA, 2017; pp. 1–275. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Family | Genus | HPD95 | P0 | D |
|---|---|---|---|---|---|
| Firmicutes | Streptococcaceae | Streptococcus | [−0.03, 1.59] | 96.71 | 0.76 |
| Staphylococcaceae | Faecalibacterium | [−1.51, 0.15] | 94.89 | −0.68 | |
| Ruminococcaceae | Paludicola | [−0.2, 1.47] | 92.69 | 0.61 | |
| Peptostreptococcaceae | Romboutsia | [0.11, 1.68] | 98.62 | 0.90 | |
| Oscillospirales | Hydrogenoanaerobacterium | [0.04, 1.65] | 97.99 | 0.84 | |
| Oscillospiraceae | UCG005 | [0.66, 1.97] | 99.98 | 1.31 | |
| Monoglobaceae | Monoglobus | [−1.49, 0.16] | 94.64 | −0.67 | |
| Leuconostocaceae | Weissella | [−0.18, 1.5] | 92.95 | 0.62 | |
| Lachnospiraceae | Frisingicoccus | [0.63, 1.95] | 99.97 | 1.29 | |
| Marvinbryantia | [0.55, 1.94] | 99.89 | 1.22 | ||
| Erysipelotrichaceae | Turicibacter | [0.77, 2.01] | 99.99 | 1.40 | |
| Erysipelatoclostridiaceae | Erysipelatoclostridium | [−1.65, −0.09] | 98.64 | −0.89 | |
| Anaerovoracaceae | Family_XIII_UCG001 | [−0.25, 1.43] | 92.26 | 0.60 | |
| Bacteroidota | Bacteroidaceae | Bacteroides | [0.7, 1.98] | 99.99 | 1.33 |
| Superclass | Class | Subclass | Metabolite | HPD95 Phage-Control | P0 Phage-Control | D Phage-Control |
|---|---|---|---|---|---|---|
| Benzenoids | Benzene and substituted derivates | Phenyl methyl carbamates | 2-(Ethylsulfonylmethyl)phenyl methyl carbamate-like | [0.3, 1.8] | 99.47 | 1.03 |
| Lipids and lipid-like molecules | Fatty acyls | Eicosanoids | 9-deoxy-9-methylene-PGE2 | [−1.43, 0.27] | 92.19 | −0.60 |
| Fatty alcohols | Persenone A-like | [0.54, 1.93] | 99.88 | 1.21 | ||
| Stigmasterols and C24-ethyl derivatives | 5alpha,8alpha-epidioxy-stigmasta-6,9(11),22E-trien-3beta-ol-like | [−1.73, −0.18] | 98.92 | −0.93 | ||
| Cholesterol and derivates | 9,11alpha-epoxy-6alpha-acetoxy-cholest-7-en-3beta,5alpha,19-triol | [0.37, 1.82] | 99.80 | 1.11 | ||
| Steroids and steroid derivates | Steroid ester | Estra-1,3,5(10)-triene-3,6beta,17beta-triol triacetate | [0.53, 1.91] | 99.93 | 1.22 | |
| Sulfate steroids | Pregnanolone sulfate | [−1.82, −0.35] | 99.70 | −1.08 | ||
| Bile acids, alcohols and derivates | Perulactone | [1.05, 2.11] | 100.00 | 1.57 | ||
| Stigmastane and derivates | 7-Oxostigmasterol-like | [0.82, 2.03] | 100.00 | 1.42 | ||
| Sphingolipids | Phosphosphingolipids | SM(d18:1/0:0) | [−1.82, −0.35] | 99.69 | −1.08 | |
| Prenol lipids | Isoprenoids | (+)-3beta-Hydroxy-ursan-28-oic acid-like | [−1.71, −0.14] | 98.85 | −0.92 | |
| Glycerophospholipids | Glycerophosphoethanolamines | PE(14:0/0:0) | [0.37, 1.81] | 99.80 | 1.09 | |
| PC(18:2(2E,4E)/0:0) | [−1.62, −0.03] | 97.83 | −0.83 | |||
| Organic acids and derivates | Carboxylic acids and derivates | Amino acids, peptides and analogues | Yersiniabactin | [1.73, 2.12] | 100.00 | 1.91 |
| Monocarboxylic acid | 1-(3,4-Dihydroxyphenyl)-1-decene-3,5-dione | [−1.11, 0.6] | 72.98 | −0.26 | ||
| Carboxylic acid derivates | (S,E)-Lyratol propanoate | [0.68, 1.98] | 99.98 | 1.33 | ||
| Organic nitrogen compounds | Organonitrogen compounds | Organic nitrous compounds | 3-[(3-Methylbutyl)nitrosamine]-2-butanone | [0.82, 2.03] | 99.99 | 1.43 |
| Organic oxygen compounds | Organooxygen compounds | Carbohydrates and carbohydrate conjugates | D-Glucosamine 1-phosphate | [0.59, 1.94] | 99.97 | 1.28 |
| Carbohydrates and carbohydrate conjugates | Glucosyl (E)-2,6-Dimethyl-2,5-heptadienoate | [−1.88, −0.46] | 99.90 | −1.18 | ||
| Organoheterocyclic compounds | Tetrapyrroles and derivates | Bilirubins | Mesobilirubinogen | [0.18, 1.71] | 99.22 | 0.95 |
| Phenylpropanoids and polyketides | Stilbenes | Stilbenes | Batatasin III-like | [−1.84, −0.37] | 99.73 | −1.09 |
| Flavonoids | Flavans | Kaempferol 7,4′-dimethyl ether 3-(6″-(E)-p-coumarylglucoside)-like | [1.27, 2.15] | 100.00 | 1.71 |
| Superclass | Class | Subclass | Metabolite | HPD95 Phage-Control | P0 Phage-Control | D Phage-Control |
|---|---|---|---|---|---|---|
| Organoheterocyclic compounds | Indoles and derivates | Tryptamines and derivates | 5-Methoxytryptophan | [−1.8, −0.3] | 99.63 | −1.05 |
| Pteridines and derivates | Pterins and derivates | 6-Lactoyltetrahydropterin | [−1.63, −0.03] | 97.69 | −0.82 | |
| Heteroarene | Polycyclic heteroarene | Indolylmethylthiohydroximate | [−1.88, −0.48] | 99.92 | −1.2 | |
| Organic oxygen compounds | Organooxygen compounds | Carbohydrates and carbohydrate conjugates | D-Mannitol | [−1.7, −0.14] | 98.8 | −0.92 |
| Organic acids and derivates | Carboxylic acids and derivates | Amino acids, peptides, and analogues | L-Ornithuric acid | [0.35, 1.8] | 99.69 | 1.08 |
| Prolyl-Tyrosine | [−0.61, 1.13] | 69.97 | 0.22 | |||
| Lipids and lipid-like molecules | Steroids and steroid derivates | Bile acids, alcohols and derivates | Murocholic acid-like | [−1.41, 0.27] | 91.6 | −0.58 |
| Glycerophospholipids | Glycerophosphoserines | PE(17:0/0:0) | [−0.26, 1.42] | 92.76 | 0.61 | |
| Phenylpropanoids and polykeides | Isoflavonoids | O-methylated isoflavonoids | Homoferreirin | [−1.43, 0.27] | 91.27 | −0.57 |
| Non-identified metabolite 50 | [−1.7, −0.16] | 98.85 | −0.93 | |||
| Non-identified metabolite 322 | [−1.81, −0.32] | 99.63 | −1.06 | |||
| Non-identified metabolite 323 | [−1.87, −0.42] | 99.81 | −1.12 | |||
| Non-identified metabolite 346 | [−1.78, −0.29] | 99.62 | −1.06 | |||
| Non-identified metabolite 395 | [−1.81, −0.34] | 99.65 | −1.05 | |||
| Non-identified metabolite 458 | [−1.81, −0.32] | 99.59 | −1.05 | |||
| Non-identified metabolite 461 | [0.27, 1.76] | 99.51 | 1.02 | |||
| Non-identified metabolite 490 | [−0.3, 1.39] | 90.14 | 0.54 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenzo-Rebenaque, L.; Casto-Rebollo, C.; Diretto, G.; Frusciante, S.; Rodríguez, J.C.; Ventero, M.-P.; Molina-Pardines, C.; Vega, S.; Marin, C.; Marco-Jiménez, F. Modulation of Caecal Microbiota and Metabolome Profile in Salmonella-Infected Broilers by Phage Therapy. Int. J. Mol. Sci. 2023, 24, 15201. https://doi.org/10.3390/ijms242015201
Lorenzo-Rebenaque L, Casto-Rebollo C, Diretto G, Frusciante S, Rodríguez JC, Ventero M-P, Molina-Pardines C, Vega S, Marin C, Marco-Jiménez F. Modulation of Caecal Microbiota and Metabolome Profile in Salmonella-Infected Broilers by Phage Therapy. International Journal of Molecular Sciences. 2023; 24(20):15201. https://doi.org/10.3390/ijms242015201
Chicago/Turabian StyleLorenzo-Rebenaque, Laura, Cristina Casto-Rebollo, Gianfranco Diretto, Sarah Frusciante, Juan Carlos Rodríguez, María-Paz Ventero, Carmen Molina-Pardines, Santiago Vega, Clara Marin, and Francisco Marco-Jiménez. 2023. "Modulation of Caecal Microbiota and Metabolome Profile in Salmonella-Infected Broilers by Phage Therapy" International Journal of Molecular Sciences 24, no. 20: 15201. https://doi.org/10.3390/ijms242015201
APA StyleLorenzo-Rebenaque, L., Casto-Rebollo, C., Diretto, G., Frusciante, S., Rodríguez, J. C., Ventero, M.-P., Molina-Pardines, C., Vega, S., Marin, C., & Marco-Jiménez, F. (2023). Modulation of Caecal Microbiota and Metabolome Profile in Salmonella-Infected Broilers by Phage Therapy. International Journal of Molecular Sciences, 24(20), 15201. https://doi.org/10.3390/ijms242015201