

Evaluation of an Image-Derived Input Function for Kinetic Modeling of Nicotinic Acetylcholine Receptor-Binding PET Ligands in Mice

 , , ,
, , ,
Abstract
:1. Introduction
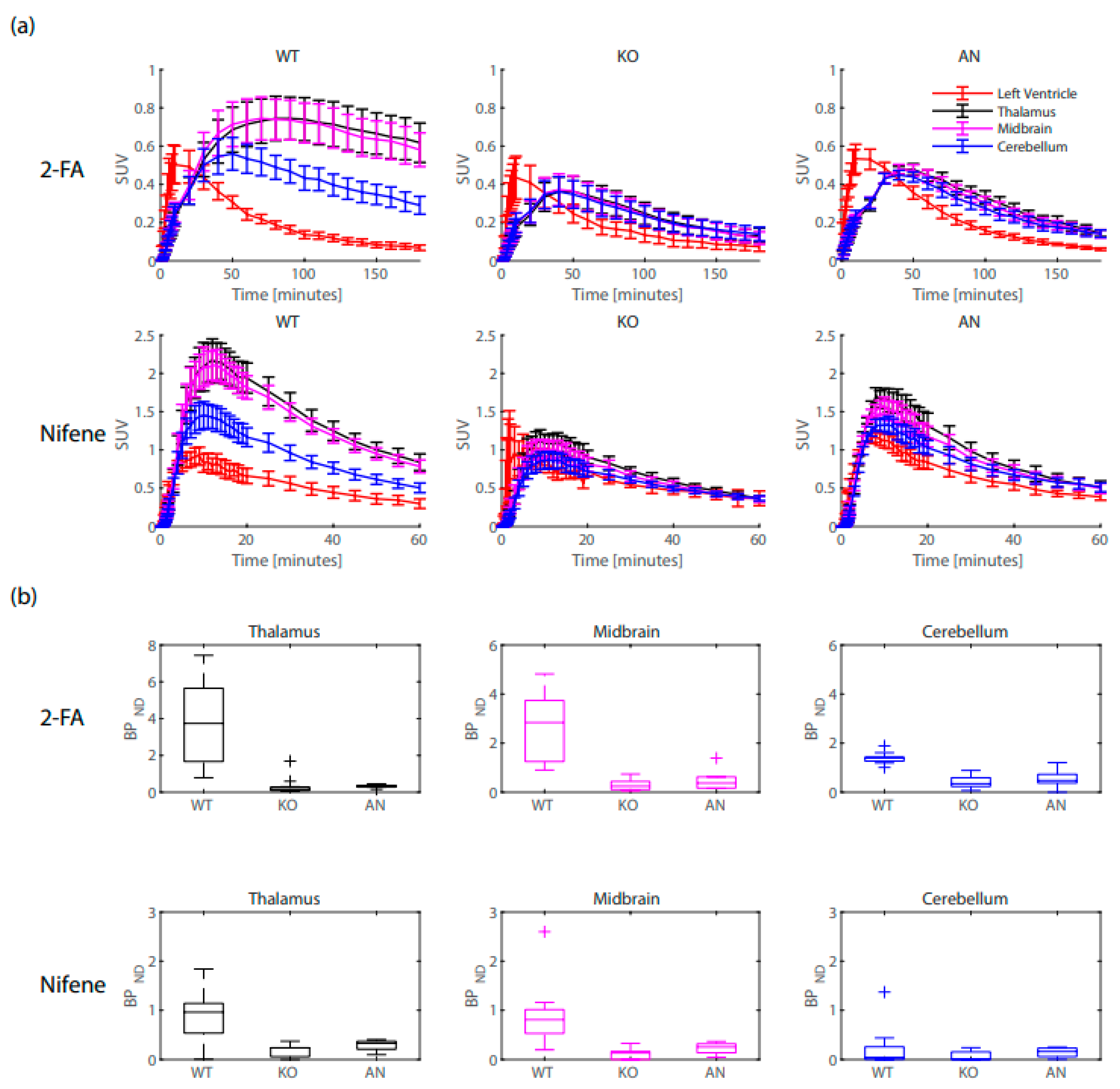
2. Results
2.1. Left Ventricle Comparisons
2.2. Binding Potential Comparisons
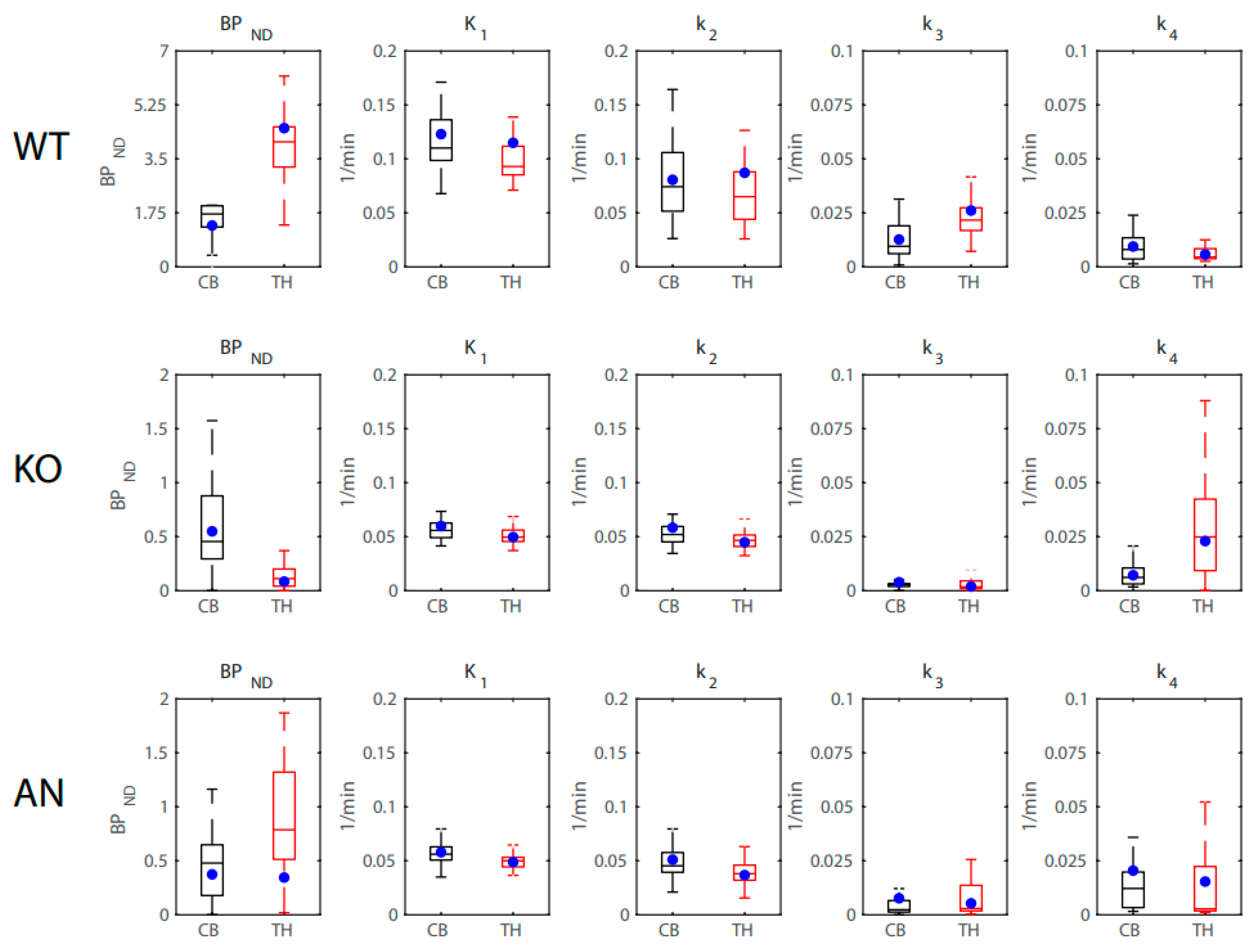
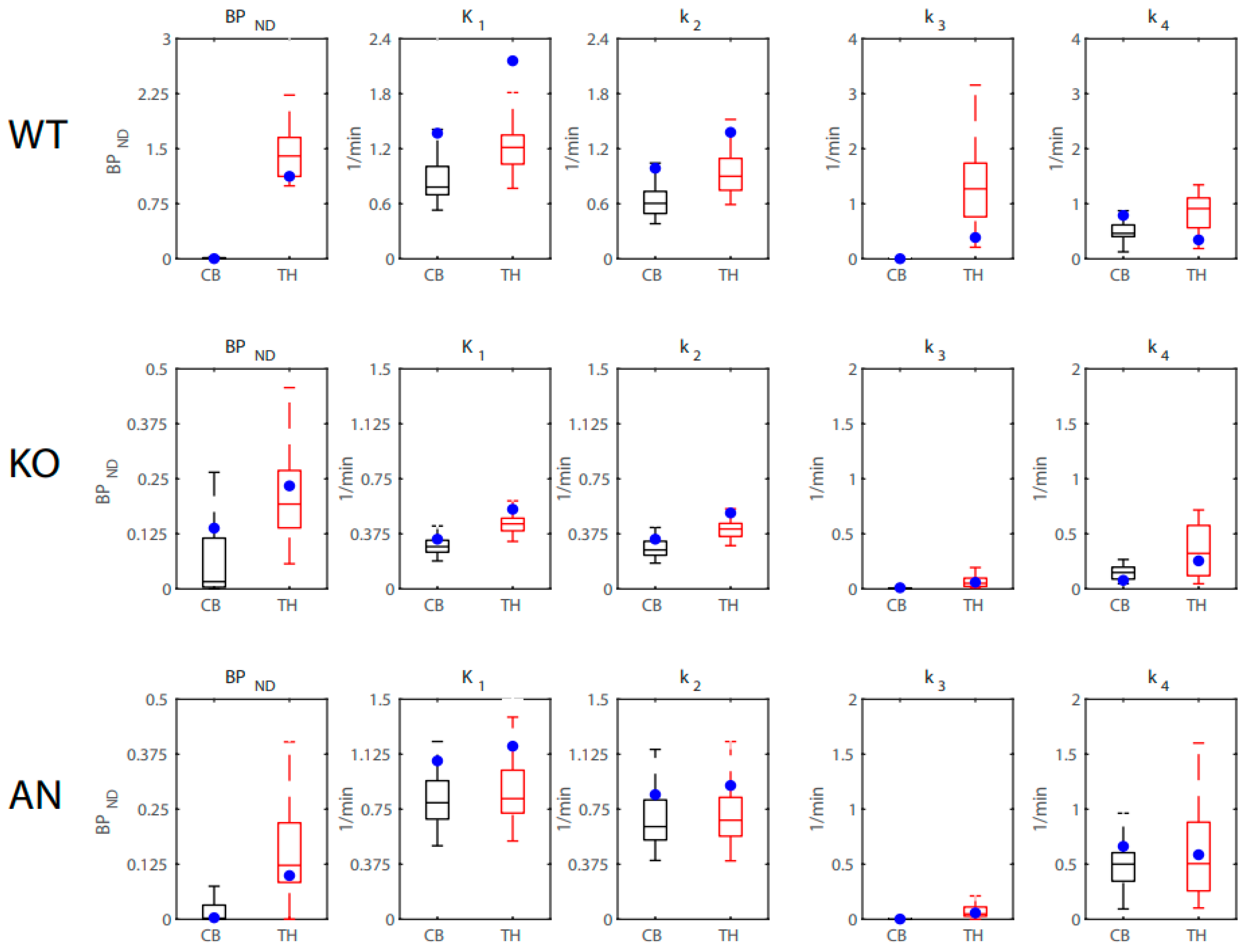
2.3. 2TCM Rate Constants
2.4. Rate Constant Comparisons between In-House Python Solver and PMOD Solver
2.5. 2TCM Comparisons to Logan Graphical Analysis
2.6. Simulations
3. Discussion
Limitations to the Study
4. Materials and Methods
4.1. Animals
4.2. Radioligand Syntheses
4.3. PET/CT Imaging
4.4. Image Quantification
4.5. Radioligand Kinetic Modeling
4.6. 2TCM Simulations
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- SurgeonGeneral The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General (2014). 2014. Available online: https://stacks.cdc.gov/view/cdc/21569 (accessed on 4 September 2023).
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian Nicotinic Acetylcholine Receptors: From Structure to Function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Benwell, M.E.; Balfour, D.J.; Anderson, J.M. Evidence That Tobacco Smoking Increases the Density of (-)-[3H]Nicotine Binding Sites in Human Brain. J. Neurochem. 1988, 50, 1243–1247. [Google Scholar] [CrossRef]
- Breese, C.R.; Marks, M.J.; Logel, J.; Adams, C.E.; Sullivan, B.; Collins, A.C.; Leonard, S. Effect of Smoking History on [3H]Nicotine Binding in Human Postmortem Brain. J. Pharmacol. Exp. Ther. 1997, 282, 7–13. [Google Scholar] [PubMed]
- Marks, M.J.; Burch, J.B.; Collins, A.C. Effects of Chronic Nicotine Infusion on Tolerance Development and Nicotinic Receptors. J. Pharmacol. Exp. Ther. 1983, 226, 817–825. [Google Scholar] [PubMed]
- Schwartz, R.D.; Kellar, K.J. Nicotinic Cholinergic Receptor Binding Sites in the Brain: Regulation In Vivo. Science 1983, 220, 214–216. [Google Scholar] [CrossRef]
- Lewis, A.S.; Picciotto, M.R. High-Affinity Nicotinic Acetylcholine Receptor Expression and Trafficking Abnormalities in Psychiatric Illness. Psychopharmacology 2013, 229, 477–485. [Google Scholar] [CrossRef]
- Vezina, P.; McGehee, D.S.; Green, W.N. Exposure to Nicotine and Sensitization of Nicotine-Induced Behaviors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; Garthwaite, J. Intracellular pH and the Distribution of Weak Acids and Bases in Isolated Rat Superior Cervical Ganglia. J. Physiol. 1979, 297, 597–620. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zammit, M.; Kao, C.-M.; Govind, A.P.; Mitchell, S.; Holderman, N.; Bhuiyan, M.; Freifelder, R.; Kucharski, A.; Zhuang, X.; et al. Trapping of Nicotinic Acetylcholine Receptor Ligands Assayed by In Vitro Cellular Studies and In Vivo PET Imaging. J. Neurosci. 2023, 43, 2–13. [Google Scholar] [CrossRef]
- Govind, A.P.; Jeyifous, O.; Russell, T.A.; Yi, Z.; Weigel, A.V.; Ramaprasad, A.; Newell, L.; Ramos, W.; Valbuena, F.M.; Casler, J.C.; et al. Activity-Dependent Golgi Satellite Formation in Dendrites Reshapes the Neuronal Surface Glycoproteome. eLife 2021, 10, e68910. [Google Scholar] [CrossRef]
- Cosgrove, K.P.; Esterlis, I.; Sandiego, C.; Petrulli, R.; Morris, E.D. Imaging Tobacco Smoking with PET and SPECT. In The Neuropharmacology of Nicotine Dependence; Balfour, D.J.K., Munafò, M.R., Eds.; Current Topics in Behavioral Neurosciences; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–17. ISBN 978-3-319-13482-6. [Google Scholar]
- Lundqvist, H.; Nordberg, A.; Hartvig, P.; Långström, B. (S)-(-)-[11C]Nicotine Binding Assessed by PET: A Dual Tracer Model Evaluated in the Rhesus Monkey Brain. Alzheimer Dis. Assoc. Disord. 1998, 12, 238–246. [Google Scholar] [CrossRef]
- Mazière, M.; Delforge, J. PET Imaging of [11C]Nicotine: Historical Aspects. In Brain Imaging of Nicotine and Tobacco Smoking; NPP Books: Ann Arbor, MI, USA, 1995; pp. 13–28. [Google Scholar]
- Sihver, W.; Nordberg, A.; Långström, B.; Mukhin, A.G.; Koren, A.O.; Kimes, A.S.; London, E.D. Development of Ligands for In Vivo Imaging of Cerebral Nicotinic Receptors. Behav. Brain Res. 2000, 113, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Horti, A.; Ravert, H.T.; London, E.D.; Dannals, R.F. Synthesis of a Radiotracer for Studying Nicotinic Acetylcholine Receptors: (+/−)-Exo-2-(2-[18F]Fluoro-5-Pyridyl)-7-Azabicyclo [2.2.1]Heptane. J. Label. Compd. Radiopharm. 1996, 38, 355–365. [Google Scholar] [CrossRef]
- Chefer, S.I.; London, E.D.; Koren, A.O.; Pavlova, O.A.; Kurian, V.; Kimes, A.S.; Horti, A.G.; Mukhin, A.G. Graphical Analysis of 2-[18F]FA Binding to Nicotinic Acetylcholine Receptors in Rhesus Monkey Brain. Synapse 2003, 48, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Lao, P.J.; Betthauser, T.J.; Samra, G.K.; Pan, M.-L.; Patel, I.H.; Liang, C.; Metherate, R.; Christian, B.T. Human Brain Imaging of Nicotinic Acetylcholine A4β2* Receptors Using [18F]Nifene: Selectivity, Functional Activity, Toxicity, Aging Effects, Gender Effects, and Extrathalamic Pathways. J. Comp. Neurol. 2018, 526, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, D.B.; Stein, E.A.; Mukhin, A.G. Quantification of A4β2* Nicotinic Receptors in the Rat Brain with microPET® and 2-[18F]F-A-85380. NeuroImage 2007, 34, 1352–1362. [Google Scholar] [CrossRef]
- Zanotti-Fregonara, P.; Chen, K.; Liow, J.-S.; Fujita, M.; Innis, R.B. Image-Derived Input Function for Brain PET Studies: Many Challenges and Few Opportunities. J. Cereb. Blood Flow. Metab. 2011, 31, 1986–1998. [Google Scholar] [CrossRef]
- Ferl, G.Z.; Zhang, X.; Wu, H.-M.; Kreissl, M.C.; Huang, S.-C. Estimation of the 18F-FDG Input Function in Mice by Use of Dynamic Small-Animal PET and Minimal Blood Sample Data. J. Nucl. Med. 2007, 48, 2037–2045. [Google Scholar] [CrossRef]
- Tantawy, M.N.; Peterson, T.E. Simplified [18F]FDG Image-Derived Input Function Using the Left Ventricle, Liver, and One Venous Blood Sample. Mol. Imaging 2010, 9, 76–86. [Google Scholar] [CrossRef]
- Hernandez, A.M.; Huber, J.S.; Murphy, S.T.; Janabi, M.; Zeng, G.L.; Brennan, K.M.; O’Neil, J.P.; Seo, Y.; Gullberg, G.T. Longitudinal Evaluation of Left Ventricular Substrate Metabolism, Perfusion, and Dysfunction in the Spontaneously Hypertensive Rat Model of Hypertrophy Using Small-Animal PET/CT Imaging. J. Nucl. Med. 2013, 54, 1938–1945. [Google Scholar] [CrossRef]
- Weinberg, I.N.; Huang, S.C.; Hoffman, E.J.; Araujo, L.; Nienaber, C.; McKay, M.G.; Dahlbom, M.; Schelbert, H. Validation of PET-Acquired Input Functions for Cardiac Studies. J. Nucl. Med. 1988, 29, 241–247. [Google Scholar] [PubMed]
- Dunnwald, L.K.; Doot, R.K.; Specht, J.M.; Gralow, J.R.; Ellis, G.K.; Livingston, R.B.; Linden, H.M.; Gadi, V.K.; Kurland, B.F.; Schubert, E.K.; et al. PET Tumor Metabolism in Locally Advanced Breast Cancer Patients Undergoing Neoadjuvant Chemotherapy: Value of Static versus Kinetic Measures of Fluorodeoxyglucose Uptake. Clin. Cancer Res. 2011, 17, 2400–2409. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, A.T.; Wooten, D.W.; Slesarev, M.S.; Ahlers, E.O.; Barnhart, T.E.; Murali, D.; Schneider, M.L.; Mukherjee, J.; Christian, B.T. PET Imaging of A4β2* Nicotinic Acetylcholine Receptors: Quantitative Analysis of 18F-Nifene Kinetics in the Nonhuman Primate. J. Nucl. Med. 2012, 53, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, A.T.; Wooten, D.W.; Slesarev, M.S.; Ahlers, E.O.; Barnhart, T.E.; Schneider, M.L.; Mukherjee, J.; Christian, B.T. Measuring A4β2* Nicotinic Acetylcholine Receptor Density In Vivo with [18F]Nifene PET in the Nonhuman Primate. J. Cereb. Blood Flow. Metab. 2013, 33, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.D.; Lockman, P.R.; Roder, K.E.; Dwoskin, L.P.; Crooks, P.A. Active Transport of High-Affinity Choline and Nicotine Analogs into the Central Nervous System by the Blood-Brain Barrier Choline Transporter. J. Pharmacol. Exp. Ther. 2003, 304, 1268–1274. [Google Scholar] [CrossRef] [PubMed]
- Deuther-Conrad, W.; Patt, J.T.; Lockman, P.R.; Allen, D.D.; Patt, M.; Schildan, A.; Ganapathy, V.; Steinbach, J.; Sabri, O.; Brust, P. Norchloro-Fluoro-Homoepibatidine (NCFHEB)—A Promising Radioligand for Neuroimaging Nicotinic Acetylcholine Receptors with PET. Eur. Neuropsychopharmacol. 2008, 18, 222–229. [Google Scholar] [CrossRef]
- Govind, A.P.; Vallejo, Y.F.; Stolz, J.R.; Yan, J.Z.; Swanson, G.T.; Green, W.N. Selective and Regulated Trapping of Nicotinic Receptor Weak Base Ligands and Relevance to Smoking Cessation. eLife 2017, 6, e25651. [Google Scholar] [CrossRef]
- Lippiello, P.M.; Sears, S.B.; Fernandes, K.G. Kinetics and Mechanism of L-[3H]Nicotine Binding to Putative High Affinity Receptor Sites in Rat Brain. Mol. Pharmacol. 1987, 31, 392–400. [Google Scholar]
- Whiteaker, P.; Sharples, C.G.V.; Wonnacott, S. Agonist-Induced Up-Regulation of A4β2 Nicotinic Acetylcholine Receptors in M10 Cells: Pharmacological and Spatial Definition. Mol. Pharmacol. 1998, 53, 950–962. [Google Scholar]
- Logan, J.; Fowler, J.S.; Volkow, N.D.; Wang, G.-J.; Ding, Y.-S.; Alexoff, D.L. Distribution Volume Ratios without Blood Sampling from Graphical Analysis of PET Data. J. Cereb. Blood Flow. Metab. 1996, 16, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Zoli, M.; Léna, C.; Bessis, A.; Lallemand, Y.; Le Novère, N.; Vincent, P.; Pich, E.M.; Brûlet, P.; Changeux, J.P. Abnormal Avoidance Learning in Mice Lacking Functional High-Affinity Nicotine Receptor in the Brain. Nature 1995, 374, 65–67. [Google Scholar] [CrossRef]
- Horti, A.G.; Chefer, S.I.; Mukhin, A.G.; Koren, A.O.; Gündisch, D.; Links, J.M.; Kurian, V.; Dannals, R.F.; London, E.D. 6-[18F]Fluoro-A-85380, a Novel Radioligand for In Vivo Imaging of Central Nicotinic Acetylcholine Receptors. Life Sci. 2000, 67, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, S.; Blankemeyer, E.; Mollet, P.; Surti, S.; Van Holen, R.; Karp, J.S. Performance Evaluation of the MOLECUBES β-CUBE-a High Spatial Resolution and High Sensitivity Small Animal PET Scanner Utilizing Monolithic LYSO Scintillation Detectors. Phys. Med. Biol. 2018, 63, 155013. [Google Scholar] [CrossRef] [PubMed]
- Hesterman, J.; Ghayoor, A.; Novicki, A.; Wang, X.; Cadoret, Y.; Becerra, L.; Gunn, R.; Avants, B. Multi-Atlas Approaches for Image Segmentation across Modality, Species and Application Area. Konica Minolta Technol. Rep. 2019, 16, 5. [Google Scholar]
- Mazur, C.; Powers, B.; Zasadny, K.; Sullivan, J.M.; Dimant, H.; Kamme, F.; Hesterman, J.; Matson, J.; Oestergaard, M.; Seaman, M.; et al. Brain Pharmacology of Intrathecal Antisense Oligonucleotides Revealed through Multimodal Imaging. JCI Insight 2019, 4, e129240. [Google Scholar] [CrossRef]
- Slavine, N.V.; Kulkarni, P.V.; McColl, R.W. Iterative Image Processing for Early Diagnostic of Beta-Amyloid Plaque Deposition in Pre-Clinical Alzheimer’s Disease Studies. J. Appl. Bioinforma Comput. Biol. 2017, 6, 134. [Google Scholar] [CrossRef]
- Lammertsma, A.A.; Hume, S.P. Simplified Reference Tissue Model for PET Receptor Studies. NeuroImage 1996, 4, 153–158. [Google Scholar] [CrossRef]
- Gunn, R.N.; Gunn, S.R.; Cunningham, V.J. Positron Emission Tomography Compartmental Models. J. Cereb. Blood Flow. Metab. 2001, 21, 635–652. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Radioligand | Region | Rate Constant (1/min) | WT | KO | AN | ANOVA F(df) | p-Value | η2 |
|---|---|---|---|---|---|---|---|---|
| 2-FA | Thalamus | K1 | 0.098 (0.013) | 0.066 (0.0061) | 0.060 (0.0036) | 3.88(2) | 0.035 | 0.24 [0.00, 0.45] |
| k2 | 0.075 (0.015) | 0.055 (0.0066) | 0.050 (0.0046) | 1.37(2) | 0.27 | 0.10 [0.00, 0.31] | ||
| k3 | 0.029 (0.0064) | 0.0049 (0.0016) | 0.0054 (0.00080) | 9.75(2) | 0.00080 | 0.45 [0.11, 0.62] | ||
| k4 | 0.0077 (0.0013) | 0.020 (0.0037) | 0.016 (0.0017) | 5.48(2) | 0.011 | 0.31 [0.02, 0.51] | ||
| Midbrain | K1 | 0.11 (0.017) | 0.076 (0.0087) | 0.062 (0.0051) | 2.67(2) | 0.090 | 0.18 [0.00, 0.39] | |
| k2 | 0.076 (0.020) | 0.066 (0.0087) | 0.051 (0.0055) | 0.54(2) | 0.59 | 0.04 [0.00, 0.21] | ||
| k3 | 0.024 (0.0062) | 0.0060 (0.0012) | 0.0031 (0.00070) | 6.70(2) | 0.0049 | 0.36 [0.05, 0.55] | ||
| k4 | 0.0084 (0.0016) | 0.026 (0.0037) | 0.0093 (0.0022) | 11.70(2) | 0.00029 | 0.49 [0.16, 0.65] | ||
| Cerebellum | K1 | 0.10 (0.016) | 0.077 (0.0072) | 0.063 (0.0053) | 2.53(2) | 0.10 | 0.17 [0.00,0.39] | |
| k2 | 0.080 (0.016) | 0.068 (0.0069) | 0.058 (0.0057) | 0.68(2) | 0.52 | 0.05 [0.00, 0.23] | ||
| k3 | 0.011 (0.0025) | 0.0056 (0.00083) | 0.0036 (0.0010) | 4.29(2) | 0.026 | 0.26 [0.00, 0.47] | ||
| k4 | 0.0081 (0.0017) | 0.017 (0.0040) | 0.015 (0.0059) | 1.534(2) | 0.24 | 0.11 [0.00, 0.31] | ||
| Nifene | Thalamus | K1 | 1.66 (0.24) | 0.55 (0.089) | 1.25 (0.072) | 7.82(2) | 0.0027 | 0.42 [0.08, 0.60] |
| k2 | 1.23 (0.26) | 0.51 (0.074) | 1.09 (0.068) | 3.05(2) | 0.068 | 0.22 [0.00, 0.44] | ||
| k3 | 0.43 (0.11) | 0.047 (0.011) | 0.14 (0.024) | 4.96(2) | 0.017 | 0.31 [0.01, 0.52] | ||
| k4 | 0.50 (0.12) | 0.27 (0.034) | 0.51 (0.12) | 1.71(2) | 0.20 | 0.13 [0.00, 0.35] | ||
| Midbrain | K1 | 1.94 (0.31) | 0.50 (0.080) | 1.28 (0.099) | 8.00(2) | 0.0024 | 0.42 [0.08, 0.60] | |
| k2 | 1.52 (0.35) | 0.47 (0.069) | 1.14 (0.061) | 3.54(2) | 0.047 | 0.24 [0.00, 0.46] | ||
| k3 | 0.50 (0.13) | 0.029 (0.0085) | 0.12 (0.026) | 5.75(2) | 0.0098 | 0.34 [0.03, 0.54] | ||
| k4 | 0.60 (0.14) | 0.26 (0.046) | 0.60 (0.077) | 2.21(2) | 0.13 | 0.17 [0.00, 0.39] | ||
| Cerebellum | K1 | 1.56 (0.33) | 0.33 (0.061) | 0.91 (0.10) | 5.19(2) | 0.014 | 0.32 [0.02, 0.52] | |
| k2 | 1.19 (0.32) | 0.31 (0.053) | 0.83 (0.072) | 2.81(2) | 0.082 | 0.20 [0.00, 0.42] | ||
| k3 | 0.10 (0.055) | 0.010 (0.0052) | 0.033 (0.013) | 1.21(2) | 0.32 | 0.10 [0.00, 0.31] | ||
| k4 | 0.62 (0.18) | 0.19 (0.046) | 0.34 (0.14) | 2.00(2) | 0.16 | 0.16 [0.00, 0.37] |
| Group | Rate Constant | Region | 2TCM Python | 2TCM PMOD | T-Value | p-Value |
|---|---|---|---|---|---|---|
| WT | K1 | Thalamus | 0.098 (0.013) | 0.13 (0.022) | 1.17 | 0.27 |
| Midbrain | 0.11 (0.017) | 0.12 (0.014) | 1.23 | 0.25 | ||
| k2 | Thalamus | 0.075 (0.015) | 0.19 (0.085) | 1.24 | 0.24 | |
| Midbrain | 0.076 (0.020) | 0.13 (0.028) | 1.61 | 0.14 | ||
| k3 | Thalamus | 0.029 (0.0064) | 0.071 (0.021) | 1.69 | 0.12 | |
| Midbrain | 0.024 (0.0062) | 0.043 (0.0076) | 2.26 | 0.051 | ||
| k4 | Thalamus | 0.0077 (0.0013) | 0.026 (0.017) | 0.98 | 0.35 | |
| Midbrain | 0.0084 (0.0016) | 0.013 (0.0032) | 1.28 | 0.23 | ||
| BPND | Thalamus | 3.84 (0.70) | 5.70 (1.25) | 1.42 | 0.19 | |
| Midbrain | 2.66 (0.41) | 3.78 (0.55) | 1.94 | 0.084 | ||
| KO | K1 | Thalamus | 0.066 (0.0061) | 0.14 (0.030) | 2.39 | 0.038 |
| Midbrain | 0.076 (0.0087) | 0.18 (0.041) | 2.46 | 0.034 | ||
| k2 | Thalamus | 0.055 (0.0066) | 0.44 (0.18) | 1.97 | 0.077 | |
| Midbrain | 0.066 (0.0087) | 0.54 (0.20) | 2.18 | 0.054 | ||
| k3 | Thalamus | 0.0049 (0.0016) | 0.090 (0.023) | 3.49 | 0.0059 | |
| Midbrain | 0.0060 (0.0012) | 0.092 (0.027) | 2.98 | 0.014 | ||
| k4 | Thalamus | 0.020 (0.0037) | 0.049 (0.0060) | 5.39 | 0.00030 | |
| Midbrain | 0.026 (0.0037) | 0.044 (0.0080) | 3.02 | 0.013 | ||
| BPND | Thalamus | 0.34 (0.14) | 2.07 (0.63) | 2.44 | 0.035 | |
| Midbrain | 0.29 (0.064) | 1.92 (0.54) | 2.66 | 0.024 | ||
| AN | K1 | Thalamus | 0.060 (0.0036) | 0.34 (0.13) | 1.85 | 0.12 |
| Midbrain | 0.062 (0.0051) | 0.37 (0.12) | 2.26 | 0.073 | ||
| k2 | Thalamus | 0.050 (0.0046) | 1.23 (0.63) | 1.71 | 0.15 | |
| Midbrain | 0.051 (0.0055) | 1.19 (0.47) | 2.20 | 0.079 | ||
| k3 | Thalamus | 0.0054 (0.00080) | 0.087 (0.023) | 2.91 | 0.033 | |
| Midbrain | 0.0031 (0.00070) | 0.085 (0.028) | 2.67 | 0.045 | ||
| k4 | Thalamus | 0.016 (0.0017) | 0.030 (0.0028) | 3.14 | 0.026 | |
| Midbrain | 0.0093 (0.0022) | 0.031 (0.0024) | 6.03 | 0.0018 | ||
| BPND | Thalamus | 0.33 (0.037) | 2.84 (0.86) | 2.66 | 0.045 | |
| Midbrain | 0.52 (0.17) | 2.61 (0.77) | 2.77 | 0.039 |
| Group | Rate Constant | Region | 2TCM Python | 2TCM PMOD | T-Value | p-Value |
|---|---|---|---|---|---|---|
| WT | K1 | Thalamus | 1.66 (0.24) | 1.18 (0.13) | 3.60 | 0.0042 |
| Midbrain | 1.94 (0.31) | 1.45 (0.17) | 2.83 | 0.016 | ||
| k2 | Thalamus | 1.23 (0.26) | 0.61 (0.12) | 3.37 | 0.0062 | |
| Midbrain | 1.52 (0.35) | 0.85 (0.15) | 2.33 | 0.040 | ||
| k3 | Thalamus | 0.43 (0.11) | 0.76 (0.51) | 0.72 | 0.49 | |
| Midbrain | 0.50 (0.13) | 0.36 (0.12) | 0.68 | 0.51 | ||
| k4 | Thalamus | 0.50 (0.12) | 1.63 (0.62) | 2.09 | 0.060 | |
| Midbrain | 0.60 (0.14) | 1.42 (0.56) | 1.77 | 0.10 | ||
| BPND | Thalamus | 0.90 (0.14) | 0.57 (0.18) | 2.28 | 0.043 | |
| Midbrain | 0.88 (0.17) | 0.63 (0.22) | 1.10 | 0.30 | ||
| KO | K1 | Thalamus | 0.55 (0.089) | 0.52 (0.12) | 0.73 | 0.48 |
| Midbrain | 0.50 (0.080) | 0.50 (0.11) | 0.0096 | 0.99 | ||
| k2 | Thalamus | 0.51 (0.074) | 0.63 (0.29) | 0.52 | 0.62 | |
| Midbrain | 0.47 (0.069) | 0.45 (0.098) | 0.33 | 0.75 | ||
| k3 | Thalamus | 0.047 (0.011) | 0.075 (0.037) | 0.62 | 0.55 | |
| Midbrain | 0.029 (0.0085) | 0.068 (0.043) | 0.87 | 0.41 | ||
| k4 | Thalamus | 0.27 (0.034) | 2.64 (1.04) | 2.16 | 0.062 | |
| Midbrain | 0.26 (0.046) | 0.65 (0.35) | 1.01 | 0.34 | ||
| BPND | Thalamus | 0.18 (0.038) | 0.030 (0.010) | 4.13 | 0.0033 | |
| Midbrain | 0.12 (0.034) | 0.11 (0.047) | 0.15 | 0.88 | ||
| AN | K1 | Thalamus | 1.25 (0.072) | 1.00 (0.050) | 1.92 | 0.15 |
| Midbrain | 1.28 (0.099) | 1.25 (0.083) | 0.20 | 0.86 | ||
| k2 | Thalamus | 1.09 (0.068) | 0.86 (0.15) | 1.09 | 0.35 | |
| Midbrain | 1.14 (0.061) | 1.29 (0.21) | 0.57 | 0.61 | ||
| k3 | Thalamus | 0.14 (0.024) | 0.65 (0.39) | 1.19 | 0.32 | |
| Midbrain | 0.12 (0.026) | 1.37 (1.04) | 1.02 | 0.38 | ||
| k4 | Thalamus | 0.51 (0.12) | 4.23 (1.73) | 1.90 | 0.15 | |
| Midbrain | 0.60 (0.077) | 3.83 (1.62) | 1.80 | 0.17 | ||
| BPND | Thalamus | 0.29 (0.056) | 0.31 (0.17) | 0.11 | 0.91 | |
| Midbrain | 0.23 (0.056) | 0.40 (0.14) | 0.88 | 0.44 |
| Radioligand | Group | Region | BPND + 1 | DVR | T-Value | p-Value |
|---|---|---|---|---|---|---|
| 2-FA | WT | Thalamus | 4.84 (0.70) | 2.35 (0.15) | 3.94 | 0.0034 |
| Midbrain | 3.66 (0.41) | 1.96 (0.080) | 4.15 | 0.0025 | ||
| KO | Thalamus | 1.34 (0.14) | 0.97 (0.019) | 2.57 | 0.028 | |
| Midbrain | 1.29 (0.064) | 0.97 (0.011) | 4.78 | 0.00075 | ||
| AN | Thalamus | 1.33 (0.037) | 1.15 (0.051) | 2.73 | 0.042 | |
| Midbrain | 1.52 (0.17) | 1.05 (0.023) | 2.40 | 0.062 | ||
| Nifene | WT | Thalamus | 1.90 (0.14) | 1.79 (0.079) | 0.65 | 0.53 |
| Midbrain | 1.88 (0.17) | 1.54 (0.092) | 1.82 | 0.095 | ||
| KO | Thalamus | 1.18 (0.038) | 0.94 (0.039) | 5.03 | 0.0010 | |
| Midbrain | 1.12 (0.034) | 0.98 (0.015) | 3.11 | 0.015 | ||
| AN | Thalamus | 1.29 (0.056) | 0.93 (0.016) | 5.79 | 0.010 | |
| Midbrain | 1.23 (0.056) | 0.99 (0.0049) | 3.57 | 0.037 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zammit, M.; Kao, C.-M.; Zhang, H.J.; Tsai, H.-M.; Holderman, N.; Mitchell, S.; Tanios, E.; Bhuiyan, M.; Freifelder, R.; Kucharski, A.; et al. Evaluation of an Image-Derived Input Function for Kinetic Modeling of Nicotinic Acetylcholine Receptor-Binding PET Ligands in Mice. Int. J. Mol. Sci. 2023, 24, 15510. https://doi.org/10.3390/ijms242115510
Zammit M, Kao C-M, Zhang HJ, Tsai H-M, Holderman N, Mitchell S, Tanios E, Bhuiyan M, Freifelder R, Kucharski A, et al. Evaluation of an Image-Derived Input Function for Kinetic Modeling of Nicotinic Acetylcholine Receptor-Binding PET Ligands in Mice. International Journal of Molecular Sciences. 2023; 24(21):15510. https://doi.org/10.3390/ijms242115510
Chicago/Turabian StyleZammit, Matthew, Chien-Min Kao, Hannah J. Zhang, Hsiu-Ming Tsai, Nathanial Holderman, Samuel Mitchell, Eve Tanios, Mohammed Bhuiyan, Richard Freifelder, Anna Kucharski, and et al. 2023. "Evaluation of an Image-Derived Input Function for Kinetic Modeling of Nicotinic Acetylcholine Receptor-Binding PET Ligands in Mice" International Journal of Molecular Sciences 24, no. 21: 15510. https://doi.org/10.3390/ijms242115510
APA StyleZammit, M., Kao, C.-M., Zhang, H. J., Tsai, H.-M., Holderman, N., Mitchell, S., Tanios, E., Bhuiyan, M., Freifelder, R., Kucharski, A., Green, W. N., Mukherjee, J., & Chen, C.-T. (2023). Evaluation of an Image-Derived Input Function for Kinetic Modeling of Nicotinic Acetylcholine Receptor-Binding PET Ligands in Mice. International Journal of Molecular Sciences, 24(21), 15510. https://doi.org/10.3390/ijms242115510