
Virome Analysis of Aconitum carmichaelii Reveals Infection by Eleven Viruses, including Two Potentially New Species

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Viruses Detected in A. carmichaelii Plants
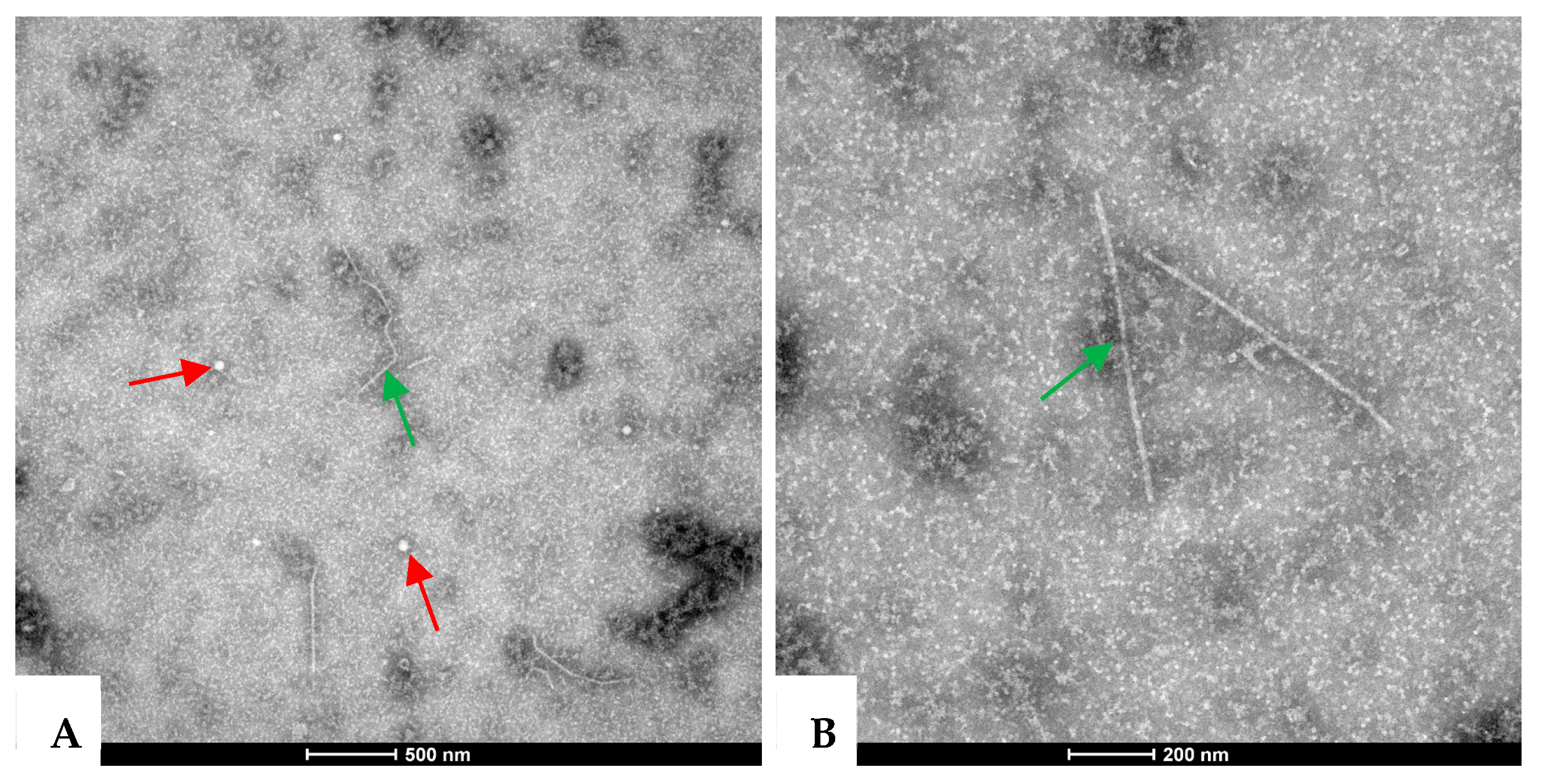
2.2. Virus Particles in the Symptomatic A. carmichaelii Samples
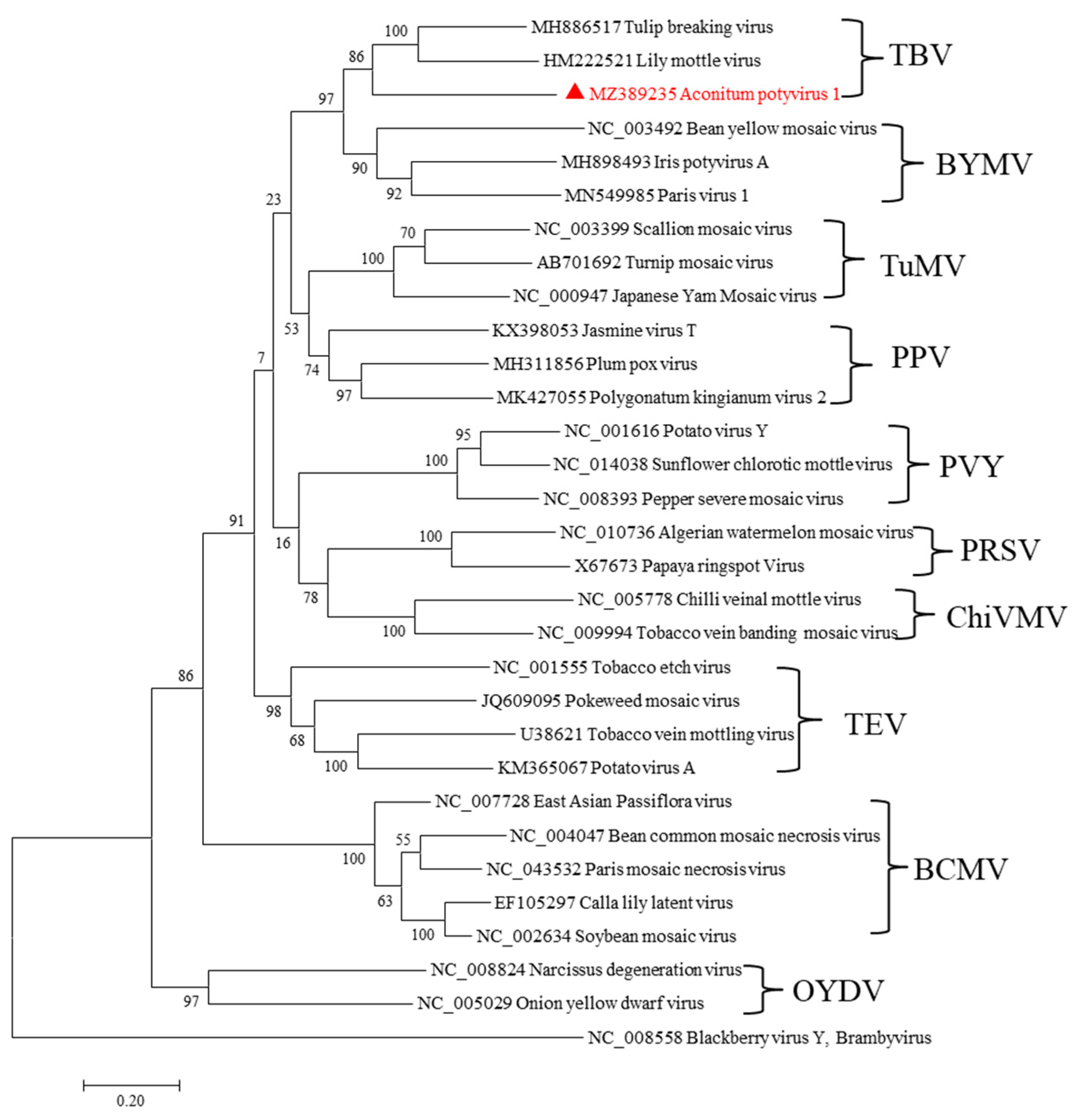
2.3. A Novel Potyvirus Infecting A. carmichaelii Plants
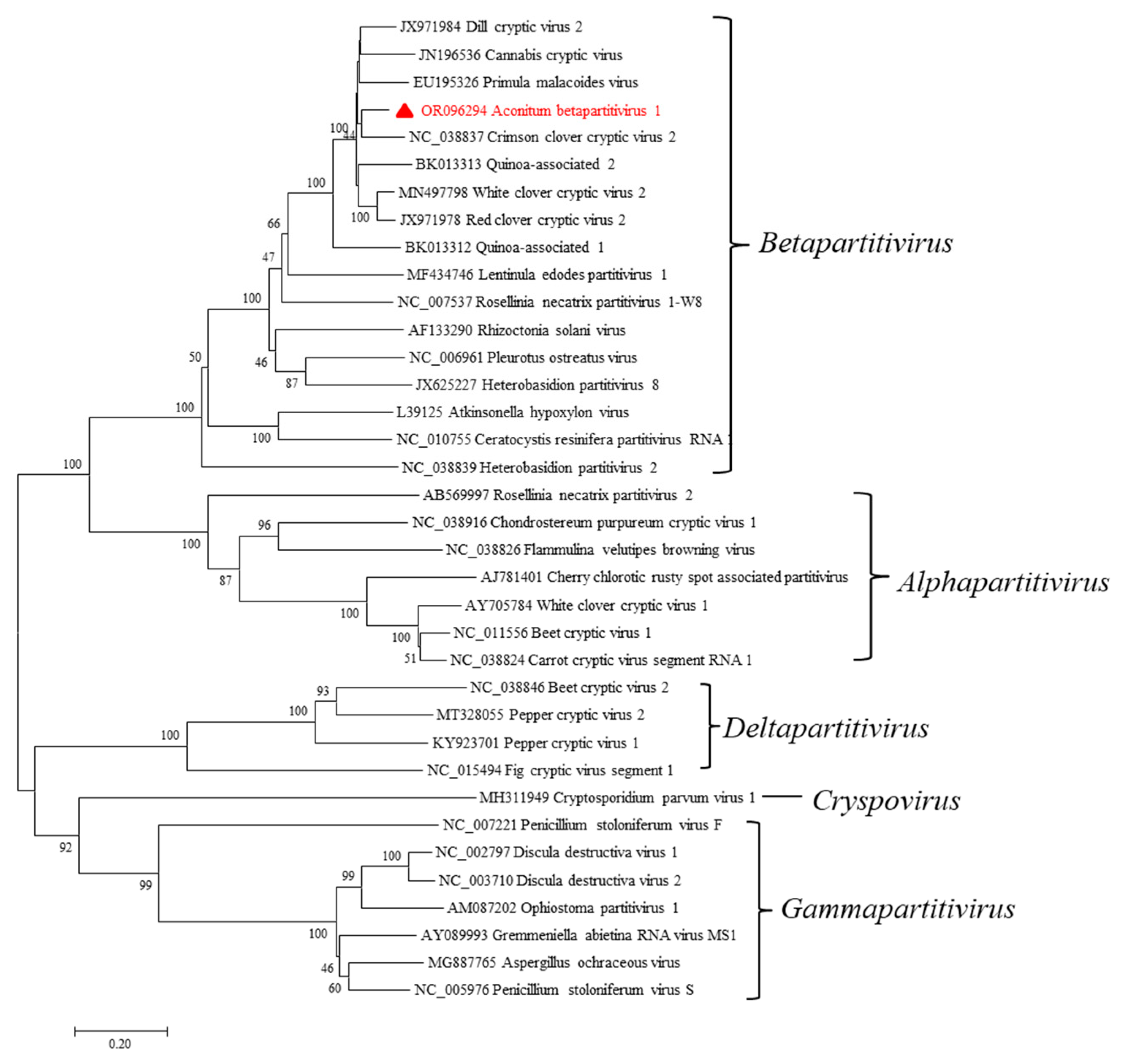
2.4. A Novel Betapartitivirus in A. carmichaelii Plants
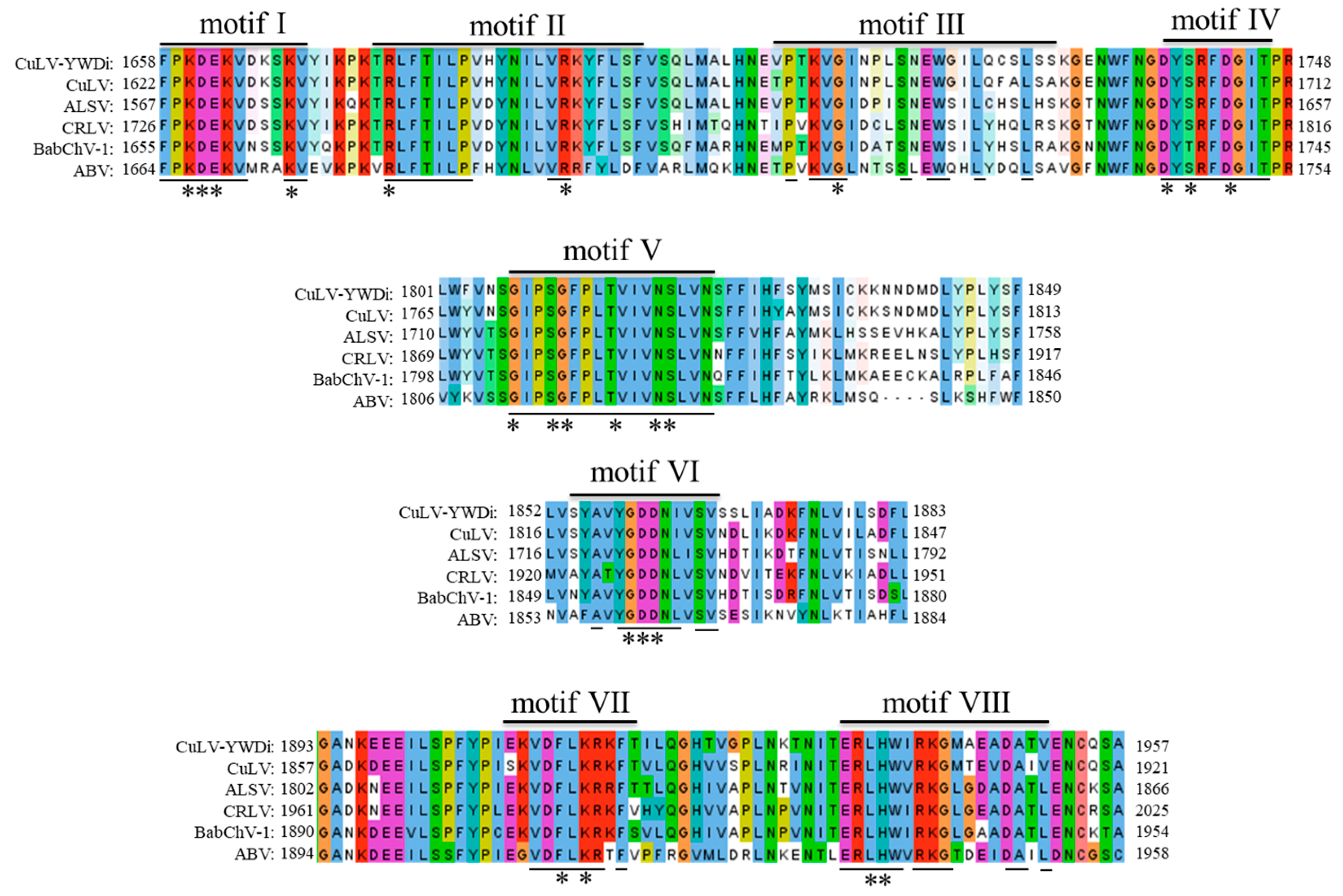
2.5. Diversity Analysis of Currant Latent Virus
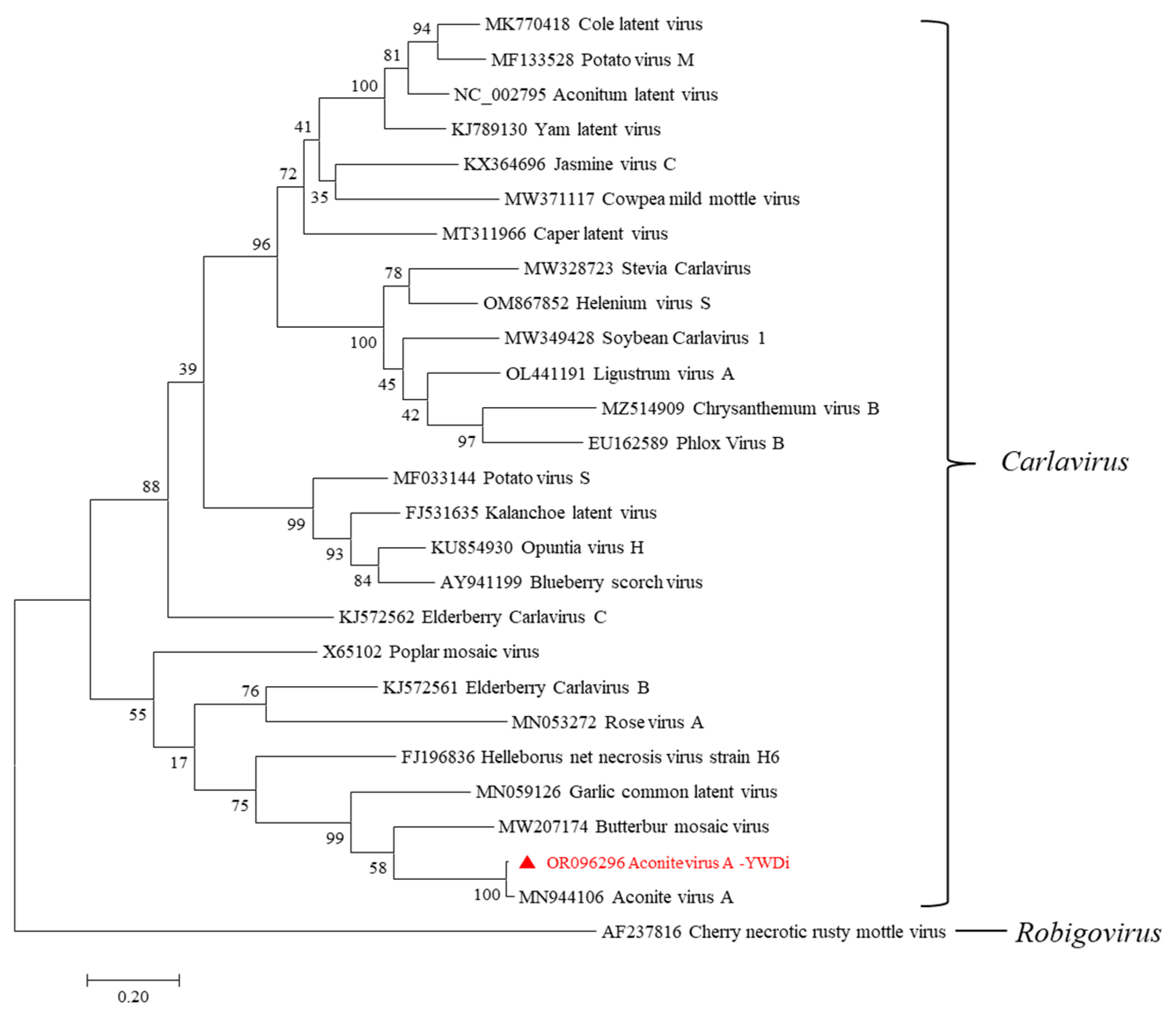
2.6. Diversity Analysis of Aconite Virus A
2.7. Incidence of Eleven Viruses in Aconitum carmichaelii Plants
2.8. Seed Transmission of CMV, AcoPV-1, AcVA, and AcoAV-1
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Virus Particle Observation
4.3. RNA Extraction, HTS, and Data Processing
4.4. Reconstruction of Virus Genomes
4.5. Virus Incidence
4.6. Genomic and Phylogenetic Analyses
4.7. Seed Transmission Test of Viruses Identified in A. carmichaelii
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Yang, Y.X.; Shu, X.Y.; Huang, J.; Hou, D.B. Aconitum carmichaelii Debeaux, cultivated as a medicinal plant in western China. Genet. Resour. Crop Evol. 2016, 63, 919–924. [Google Scholar] [CrossRef]
- Fu, Y.P.; Zou, Y.F.; Lei, F.Y.; Wangensteen, H.; Inngjerdingen, K.T. Aconitum carmichaelii Debeaux: A systematic review on traditional use, and the chemical structures and pharmacological properties of polysaccharides and phenolic compounds in the roots. J. Ethnopharmacol. 2022, 291, 115148. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Tang, L.; Zhou, X.; Wang, T.; Kou, Z.; Wang, Z. A review on phytochemistry and pharmacological activities of the processed lateral root of Aconitum carmichaelii Debeaux. J. Ethnopharmacol. 2015, 160, 173–193. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Wang, J.; Cui, Y.; Wu, X. Pharmacological effects of Chinese herb aconite (fuzi) on cardiovascular system. J. Tradit. Chin. Med. 2012, 32, 308–313. [Google Scholar] [CrossRef]
- Liu, J.; Liang, M.; Lin, T.; Zhao, Q.; Wang, H.; Yang, S.; Guo, Q.; Wang, X.; Guo, H.; Cui, L. A LAMP-based toolbox developed for detecting the major pathogens affecting the production and quality of the Chinese medicinal crop Aconitum carmichaelii. Plant Dis. 2023, 107, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Cao, L.L.; Yang, Y.X.; Guan, L.L.; Gou, L.L.; Shu, X.Y.; Huang, J.; Liu, D.; Zhang, H.; Hou, D.B. Genetic diversity and marker–trait association analysis for agronomic traits in Aconitum carmichaelii Debeaux. Biotechnol. Biotechnol. Equip. 2017, 31, 905–911. [Google Scholar] [CrossRef]
- Cohen, J.; Zeidan, M.; Rosner, A.; Gera, A. Biological and molecular characterization of a new carlavirus isolated from an Aconitum sp. Phytopathology 2000, 90, 340–344. [Google Scholar] [CrossRef]
- Fumiyoshi, F.; Shin-ichi, F.; Kouichi, S.; Masahide, I. Cucumber mosaic virus isolated from Aconitum spp. in Japan. J. Gen. Plant Pathol. 2008, 74, 88–90. [Google Scholar] [CrossRef]
- Wang, R.; Chen, B.; Li, Y.; Cao, M.; Ding, W. Complete nucleotide sequence of a new carlavirus infecting Aconitum carmichaelii in China. Arch. Virol. 2021, 166, 1513–1515. [Google Scholar] [CrossRef]
- Yang, J.; Lan, P.X.; Li, J.M.; Chen, X.J.; Tan, G.L.; Wei, T.Y.; Li, R.H.; Li, F. Complete genome sequence of Aconitum amalgavirus 1, a distinct member of the genus Amalgavirus. Arch. Virol. 2022, 167, 2351–2353. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Jordan, R.; Kreuze, J.; Li, F.; López-Moya, J.J.; Mäkinen, K.; Ohshima, K.; Wylie, S.J.; Consortium, I.R. ICTV virus taxonomy profile: Potyviridae 2022. J. Gen. Virol. 2022, 103, 1738. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.A.; Hayward, A.C.; Fletcher, S.J.; Mitter, N. Molecular characterization and analysis of conserved potyviral motifs in bean common mosaic virus (BCMV) for RNAi-mediated protection. Arch. Virol. 2019, 164, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M.; Consortium, I.R. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Petrzik, K.; Koloniuk, I.; Přibylová, J.; Špak, J. Complete genome sequence of currant latent virus (genus Cheravirus, family Secoviridae). Arch. Virol. 2016, 161, 491–493. [Google Scholar] [CrossRef]
- Li, C.; Yoshikawa, N.; Takahashi, T.; Ito, T.; Yoshida, K.; Koganezawa, H. Nucleotide sequence and genome organization of apple latent spherical virus: A new virus classified into the family Comoviridae. J. Gen. Virol. 2000, 81, 541–547. [Google Scholar] [CrossRef]
- James, D.; Upton, C. Genome segment RNA-1 of a flat apple isolate of cherry rasp leaf virus: Nucleotide sequence analysis and RT-PCR detection. Arch. Virol. 2005, 150, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.; Glover, R.; Souza-Richards, R.; Bennett, S.; Hany, U.; Boonham, N. Complete genome sequence of arracacha virus B: A novel cheravirus. Arch. Virol. 2013, 158, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Petrzik, K.; Přibylová, J.; Špak, J.; Havelka, J. Partial genome sequence of currant latent virus, a new chera-like virus related to apple latent spherical virus. J. Gen. Plant Pathol. 2015, 81, 142–145. [Google Scholar] [CrossRef]
- Khalili, M.; Candresse, T.; Brans, Y.; Faure, C.; Audergon, J.M.; Decroocq, V.; Roch, G.; Marais, A. The molecular characterization of a new prunus-infecting cheravirus and complete genome sequence of stocky prune virus. Viruses 2022, 14, 2325. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Koonin, E.V.; Wolf, Y.I. A new superfamily of putative NTP-binding domains encoded by genomes of small DNA and RNA viruses. FEBS Lett. 1990, 262, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Dolja, V.V.; Morris, T.J. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef]
- Thompson, J.R.; Dasgupta, I.; Fuchs, M.; Iwanami, T.; Karasev, A.V.; Petrzik, K.; Sanfaçon, H.; Tzanetakis, I.; van der Vlugt, R.; Wetzel, T. ICTV virus taxonomy profile: Secoviridae. J. Gen. Virol. 2017, 98, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.F.; Melo, F.L.; Elena, S.F.; Candresse, T.; Sabanadzovic, S.; Tzanetakis, I.E.; Blouin, A.G.; Villamor, D.E.V.; Mollov, D.; Constable, F. Virus classification based on in-depth sequence analyses and development of demarcation criteria using the Betaflexiviridae as a case study. J. Gen. Virol. 2022, 103, 1806. [Google Scholar] [CrossRef] [PubMed]
- Beck-Okins, A.L.; Del Río Mendoza, L.E.; Burrows, M.; Simons, K.J.; Pasche, J.S. Pea seed-borne mosaic virus (PSbMV) risk analysis of field pea based on susceptibility, yield loss, and seed transmission. Plant Dis. 2022, 106, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.J.; Nyland, G.; McElroy, F.; Stace-Smith, R. Cherry rasp leaf disease in North America. Phytopathology 1974, 64, 721–727. [Google Scholar] [CrossRef]
- Jones, R. Tests for transmission of four potato viruses through potato true seed. Ann. Appl. Biol. 1982, 100, 315–320. [Google Scholar] [CrossRef]
- Brito, M.; Fernández-Rodríguez, T.; Garrido, M.J.; Mejías, A.; Romano, M.; Marys, E. First report of cowpea mild mottle carlavirus on yardlong bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela. Viruses 2012, 4, 3804–3811. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Valverde, R.A.; Brown, J.K.; Martin, R.R.; Tzanetakis, I.E. Southern tomato virus: The link between the families Totiviridae and Partitiviridae. Virus Res. 2009, 140, 130–137. [Google Scholar] [CrossRef]
- Samarskaya, V.O.; Ryabov, E.V.; Gryzunov, N.; Spechenkova, N.; Kuznetsova, M.; Ilina, I.; Suprunova, T.; Taliansky, M.E.; Ivanov, P.A.; Kalinina, N.O. The temporal and geographical dynamics of potato virus Y diversity in Russia. Int. J. Mol. Sci. 2023, 24, 14833. [Google Scholar] [CrossRef]
- Gildow, F.; Shah, D.; Sackett, W.; Butzler, T.; Nault, B.; Fleischer, S. Transmission efficiency of cucumber mosaic virus by aphids associated with virus epidemics in snap bean. Phytopathology 2008, 98, 1233–1241. [Google Scholar] [CrossRef]
- Shah, H.; Yasmin, T.; Fahim, M.; Hameed, S.; Haque, M. Transmission and host range studies of Pakistani isolate of chilli veinal mottle virus. Pak. J. Bot. 2008, 40, 2669–2681. [Google Scholar]
- Chen, X.J.; Luo, H.M.; Zhang, J.Y.; Ma, Y.; Li, K.H.; Xiong, F.; Yang, Y.H.; Yang, J.Z.; Lan, P.X.; Wei, T.Y.; et al. Synergism among the four tobacco bushy top disease causal agents in symptom induction and aphid transmission. Front. Microbiol. 2022, 13, 846857. [Google Scholar] [CrossRef]
- Olmedo-Velarde, A.; Wilson, J.R.; Stallone, M.; DeBlasio, S.L.; Chappie, J.S.; Heck, M. Potato leafroll virus molecular interactions with plants and aphids: Gaining a new tactical advantage on an old foe. Physiol. Mol. Plant Pathol. 2023, 125, 102015. [Google Scholar] [CrossRef]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid transmission of Potyvirus: The largest plant-infecting RNA virus genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Machado Caballero, J.E.; Lockhart, B.E.; Mason, S.L.; Daughtrey, M. Identification and properties of a carlavirus causing chlorotic mottle of florists’ Hydrangea (H. macrophylla) in the United States. Plant Dis. 2009, 93, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Park, C.H.; Seo, E.Y.; Kim, J.K.; Hammond, J.; Lim, H.S. Occurrence of apple stem grooving virus in commercial apple seedlings and analysis of its coat protein sequence. Korean J. Agric. Sci. 2016, 43, 21–27. [Google Scholar] [CrossRef]
- Ito, T.; Namba, N.; Ito, T. Distribution of citrus viroids and apple stem grooving virus on citrus trees in Japan using multiplex reverse transcription polymerase chain reaction. J. Gen. Plant Pathol. 2003, 69, 205–207. [Google Scholar] [CrossRef]
- Lan, P.X.; He, P.; Yang, J.; Zhou, G.H.; Chen, X.J.; Wei, T.Y.; Li, C.R.; Gu, R.; Li, R.H.; Li, F. High-throughput sequencing reveals the presence of novel and known viruses in diseased Paris yunnanensis. Front. Microbiol. 2022, 13, 1045750. [Google Scholar] [CrossRef]
- Tan, S.T.; Liu, F.; Lv, J.; Liu, Q.L.; Luo, H.M.; Xu, Y.; Ma, Y.; Chen, X.J.; Lan, P.X.; Chen, H.R. Identification of two novel poleroviruses and the occurrence of tobacco bushy top disease causal agents in natural plants. Sci. Rep. 2021, 11, 21045. [Google Scholar] [CrossRef]
- Postler, T.S.; Rubino, L.; Adriaenssens, E.M.; Dutilh, B.E.; Harrach, B.; Junglen, S.; Kropinski, A.M.; Krupovic, M.; Wada, J.; Crane, A. Guidance for creating individual and batch latinized binomial virus species names. J. Gen. Virol. 2022, 103, 1800. [Google Scholar] [CrossRef]
- Li, R.H.; Mock, R.; Huang, Q.; Abad, J.; Hartung, J.; Kinard, G. A reliable and inexpensive method of nucleic acid extraction for the PCR-based detection of diverse plant pathogens. J. Virol. Methods 2008, 154, 48–55. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Genus | Length (nt) | Isolate Name | Region | GenBank Accession |
|---|---|---|---|---|---|
| Aconitum potyvirus 1 (AcoPV-1) | Potyvirus | 9453 | YZYi | Complete genome | MZ389235 |
| Aconitum betapartitivirus 1 (AcoBPV-1) | Betapartitivirus | 2363 | YWDi | Near-complete RNA1 | OR096294 |
| 2279 | YWDi | Complete RNA2 | OR096295 | ||
| Currant latent virus (CuLV) | Cheravirus | 6683 | YWDi | Complete RNA1 | OR096292 |
| 3070 | YWDi | Near-complete RNA2 | OR096293 | ||
| Aconite virus A (AcVA) | Carlavirus | 8833 | YWDi | Complete genome | OR096296 |
| Cucumber mosaic virus (CMV) | Cucumovirus | 657 | YWDi | Complete coat protein | MZ389240 |
| Aconitum amalgavirus 1 (AcoAV-1) | Amalgavirus | 1061 | YWDi | Partial fusion protein | OR096297 |
| Apple stem grooving virus (ASGV) | Capillovirus | 893 | YWDi | Partial polyprotein | OR096298 |
| Tobacco vein distorting virus (TVDV) | Polerovirus | 501 | YZYi | Partial coat protein | OR096301 |
| Tomato spotted wilt orthotospovirus (TSWV) | Orthotospovirus | 504 | YZYi | Partial segment M | OR096302 |
| Chilli veinal mottle virus (ChiVMV) | Potyvirus | 919 | YWDi | Partial polyprotein | OR096299 |
| Potato leafroll virus (PLRV) | Polerovirus | 599 | YZYi | Partial capsid protein | OR096300 |
| Virus | County and Collection Year | Zhanyi 2017 | Wuding 2020 | Wuding 2021 | Total |
|---|---|---|---|---|---|
| Sample Number | 17 | 71 | 54 | 142 | |
| Cucumber mosaic virus (CMV) | Positive samples | 11 | 69 | 39 | 119 |
| Incidence | 64.7% | 97.2% | 72.2% | 83.8% | |
| Aconite virus A (AcVA) | Positive samples | 0 | 65 | 35 | 100 |
| Incidence | 0 | 91.6% | 64.8% | 70.4% | |
| Aconitum potyvirus 1 (AcoPV-1) | Positive samples | 15 | 46 | 27 | 88 |
| Incidence | 88.3% | 64.8% | 50% | 62% | |
| Aconitum amalgavirus 1 (AcoAV-1) | Positive samples | 2 | 25 | 19 | 46 |
| Incidence | 11.78% | 35.2% | 35.2% | 32.4% | |
| Currant latent virus (CuLV) | Positive samples | 0 | 28 | 9 | 37 |
| Incidence | 0 | 39.5% | 16.7% | 26.1% | |
| Aconitum betapartitivirus 1 (AcoBPV-1) | Positive samples | 0 | 26 | 11 | 37 |
| Incidence | 0 | 36.6% | 20.4% | 26.1% | |
| Apple stem grooving virus (ASGV) | Positive samples | 0 | 22 | 4 | 26 |
| Incidence | 0 | 31% | 7.4% | 18.3% | |
| Tobacco vein distorting virus (TVDV) | Positive samples | 7 | 0 | 0 | 7 |
| Incidence | 41.2% | 0 | 0 | 4.9% | |
| Tomato spotted wilt orthotospovirus (TSWV) | Positive samples | 5 | 0 | 0 | 5 |
| Incidence | 29.4% | 0 | 0 | 3.5% | |
| Chilli veinal mottle virus (ChiVMV) | Positive samples | 0 | 3 | 0 | 3 |
| Incidence | 0 | 4.2% | 0 | 2.1% | |
| Potato leafroll virus (PLRV) | Positive samples | 2 | 0 | 0 | 2 |
| Incidence | 11.8% | 0 | 0 | 1.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Lan, P.-X.; Wang, Y.; Li, J.-M.; Li, R.; Wylie, S.; Chen, X.-J.; Yang, G.-H.; Cai, H.; Li, F. Virome Analysis of Aconitum carmichaelii Reveals Infection by Eleven Viruses, including Two Potentially New Species. Int. J. Mol. Sci. 2023, 24, 15558. https://doi.org/10.3390/ijms242115558
Yang J, Lan P-X, Wang Y, Li J-M, Li R, Wylie S, Chen X-J, Yang G-H, Cai H, Li F. Virome Analysis of Aconitum carmichaelii Reveals Infection by Eleven Viruses, including Two Potentially New Species. International Journal of Molecular Sciences. 2023; 24(21):15558. https://doi.org/10.3390/ijms242115558
Chicago/Turabian StyleYang, Jie, Ping-Xiu Lan, Yun Wang, Jin-Ming Li, Ruhui Li, Steve Wylie, Xiao-Jiao Chen, Gen-Hua Yang, Hong Cai, and Fan Li. 2023. "Virome Analysis of Aconitum carmichaelii Reveals Infection by Eleven Viruses, including Two Potentially New Species" International Journal of Molecular Sciences 24, no. 21: 15558. https://doi.org/10.3390/ijms242115558
APA StyleYang, J., Lan, P. -X., Wang, Y., Li, J. -M., Li, R., Wylie, S., Chen, X. -J., Yang, G. -H., Cai, H., & Li, F. (2023). Virome Analysis of Aconitum carmichaelii Reveals Infection by Eleven Viruses, including Two Potentially New Species. International Journal of Molecular Sciences, 24(21), 15558. https://doi.org/10.3390/ijms242115558