
Contribution of Telomere Length to Systemic Sclerosis Onset: A Mendelian Randomization Study
 , , , , and
, , , , and
Abstract
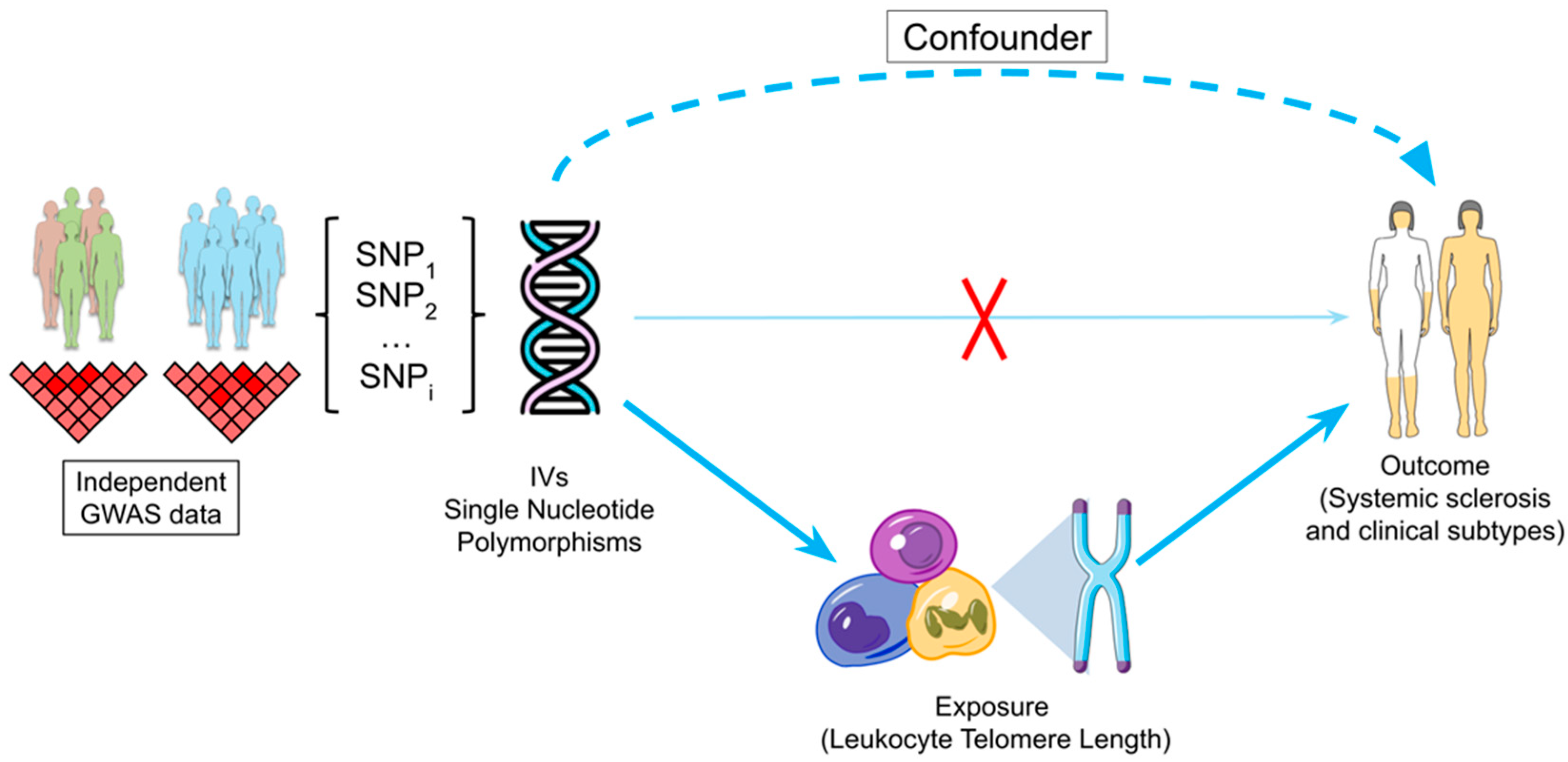
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Genetic Data Sources and IV Selection
4.2. MR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denton, C.P.; Khanna, D. Systemic Sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic Sclerosis. Nat. Rev. Dis. Primers 2015, 1, 15002. [Google Scholar] [CrossRef] [PubMed]
- López-Isac, E.; Acosta-Herrera, M.; Kerick, M.; Assassi, S.; Satpathy, A.T.; Granja, J.; Mumbach, M.R.; Beretta, L.; Simeón, C.P.; Carreira, P.; et al. GWAS for Systemic Sclerosis Identifies Multiple Risk Loci and Highlights Fibrotic and Vasculopathy Pathways. Nat. Commun. 2019, 10, 4955. [Google Scholar] [CrossRef] [PubMed]
- Terao, C.; Kawaguchi, T.; Dieude, P.; Varga, J.; Kuwana, M.; Hudson, M.; Kawaguchi, Y.; Matucci-Cerinic, M.; Ohmura, K.; Riemekasten, G.; et al. Transethnic Meta-Analysis Identifies and as Susceptibility Genes to Systemic Sclerosis. Ann. Rheum. Dis. 2017, 76, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Mayes, M.D.; Bossini-Castillo, L.; Gorlova, O.; Martin, J.E.; Zhou, X.; Chen, W.V.; Assassi, S.; Ying, J.; Tan, F.K.; Arnett, F.C.; et al. Immunochip Analysis Identifies Multiple Susceptibility Loci for Systemic Sclerosis. Am. J. Hum. Genet. 2014, 94, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Martín, G.; Martín, J.; Bossini-Castillo, L. Recent Advances in Elucidating the Genetic Basis of Systemic Sclerosis. Curr. Opin. Rheumatol. 2022, 34, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Beretta, L.; Barturen, G.; Vigone, B.; Bellocchi, C.; Hunzelmann, N.; De Langhe, E.; Cervera, R.; Gerosa, M.; Kovács, L.; Ortega Castro, R.; et al. Genome-Wide Whole Blood Transcriptome Profiling in a Large European Cohort of Systemic Sclerosis Patients. Ann. Rheum. Dis. 2020, 79, 1218–1226. [Google Scholar] [CrossRef]
- Fang, D.; Chen, B.; Lescoat, A.; Khanna, D.; Mu, R. Immune Cell Dysregulation as a Mediator of Fibrosis in Systemic Sclerosis. Nat. Rev. Rheumatol. 2022, 18, 683–693. [Google Scholar] [CrossRef]
- Keret, S.; Rimar, D.; Lansiaux, P.; Feldman, E.; Lescoat, A.; Milman, N.; Farge, D.; MATHEC Working Group. Differentially Expressed Genes in Systemic Sclerosis: Towards Predictive Medicine with New Molecular Tools for Clinicians. Autoimmun. Rev. 2023, 22, 103314. [Google Scholar] [CrossRef]
- Heba, A.-C.; Toupance, S.; Arnone, D.; Peyrin-Biroulet, L.; Benetos, A.; Ndiaye, N.C. Telomeres: New Players in Immune-Mediated Inflammatory Diseases? J. Autoimmun. 2021, 123, 102699. [Google Scholar] [CrossRef]
- Shi, B.; Tsou, P.-S.; Ma, F.; Mariani, M.P.; Mattichak, M.N.; LeBrasseur, N.K.; Chini, E.N.; Lafyatis, R.; Khanna, D.; Whitfield, M.L.; et al. Senescent Cells Accumulate in Systemic Sclerosis Skin. J. Investig. Dermatol. 2023, 143, 661–664.e5. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.M.; Park, K.-S.; Kim, K.-J. Integrative Analysis of Lung Molecular Signatures Reveals Key Drivers of Systemic Sclerosis-Associated Interstitial Lung Disease. Ann. Rheum. Dis. 2022, 81, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rane, G.; Dai, X.; Shanmugam, M.K.; Arfuso, F.; Samy, R.P.; Lai, M.K.P.; Kappei, D.; Kumar, A.P.; Sethi, G. Ageing and the Telomere Connection: An Intimate Relationship with Inflammation. Ageing Res. Rev. 2016, 25, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Rossiello, F.; Jurk, D.; Passos, J.F.; d’Adda di Fagagna, F. Telomere Dysfunction in Ageing and Age-Related Diseases. Nat. Cell Biol. 2022, 24, 135–147. [Google Scholar] [CrossRef]
- Usategui, A.; Municio, C.; Arias-Salgado, E.G.; Martín, M.; Fernández-Varas, B.; Del Rey, M.J.; Carreira, P.; González, A.; Criado, G.; Perona, R.; et al. Evidence of Telomere Attrition and a Potential Role for DNA Damage in Systemic Sclerosis. Immun. Ageing 2022, 19, 7. [Google Scholar] [CrossRef]
- Artlett, C.M.; Black, C.M.; Briggs, D.C.; Stevens, C.O.; Welsh, K.I. Telomere Reduction in Scleroderma Patients: A Possible Cause for Chromosomal Instability. Br. J. Rheumatol. 1996, 35, 732–737. [Google Scholar] [CrossRef]
- Lakota, K.; Hanumanthu, V.S.; Agrawal, R.; Carns, M.; Armanios, M.; Varga, J. Short Lymphocyte, but Not Granulocyte, Telomere Length in a Subset of Patients with Systemic Sclerosis. Ann. Rheum. Dis. 2019, 78, 1142–1144. [Google Scholar] [CrossRef]
- Liu, S.; Chung, M.P.; Ley, B.; French, S.; Elicker, B.M.; Fiorentino, D.F.; Chung, L.S.; Boin, F.; Wolters, P.J. Peripheral Blood Leucocyte Telomere Length Is Associated with Progression of Interstitial Lung Disease in Systemic Sclerosis. Thorax 2021, 76, 1186–1192. [Google Scholar] [CrossRef]
- MacIntyre, A.; Brouilette, S.W.; Lamb, K.; Radhakrishnan, K.; McGlynn, L.; Chee, M.M.; Parkinson, E.K.; Freeman, D.; Madhok, R.; Shiels, P.G. Association of Increased Telomere Lengths in Limited Scleroderma, with a Lack of Age-Related Telomere Erosion. Ann. Rheum. Dis. 2008, 67, 1780–1782. [Google Scholar] [CrossRef]
- Boehm, F.J.; Zhou, X. Statistical Methods for Mendelian Randomization in Genome-Wide Association Studies: A Review. Comput. Struct. Biotechnol. J. 2022, 20, 2338–2351. [Google Scholar] [CrossRef]
- Chen, C.; Wang, P.; Zhang, R.-D.; Fang, Y.; Jiang, L.-Q.; Fang, X.; Zhao, Y.; Wang, D.-G.; Ni, J.; Pan, H.-F. Mendelian randomization as a tool to gain insights into the mosaic causes of autoimmune diseases. Autoimmun. Rev. 2022, 21, 103210. [Google Scholar] [CrossRef] [PubMed]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of Seven Loci Affecting Mean Telomere Length and Their Association with Disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Butterworth, A.; Thompson, S.G. Mendelian Randomization Analysis with Multiple Genetic Variants Using Summarized Data. Genet. Epidemiol. 2013, 37, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Luo, P.; Liu, L.; Wei, X.; Bai, X.; Li, J.; Wu, L.; Luo, M. Immune-Mediated Inflammatory Diseases and Leukocyte Telomere Length: A Mendelian Randomization Study. Front. Genet. 2023, 14, 1129247. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhang, W.; Qian, Y.; Huang, H.; Wu, D.J.H.; He, Z.; Ye, D.; Mao, Y.; Wen, C. Association of Telomere Length with Risk of Rheumatoid Arthritis: A Meta-Analysis and Mendelian Randomization. Rheumatology 2020, 59, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Xu, W.-J.; Wang, F.-F.; Leng, R.; Yang, X.-K.; Ling, H.-Z.; Fan, Y.-G.; Tao, J.-H.; Shuai, Z.-W.; Zhang, L.; et al. Telomere Length and Development of Systemic Lupus Erythematosus: A Mendelian Randomization Study. Arthritis Rheumatol. 2022, 74, 1984–1990. [Google Scholar] [CrossRef]
- Fingerlin, T.E.; Murphy, E.; Zhang, W.; Peljto, A.L.; Brown, K.K.; Steele, M.P.; Loyd, J.E.; Cosgrove, G.P.; Lynch, D.; Groshong, S.; et al. Genome-Wide Association Study Identifies Multiple Susceptibility Loci for Pulmonary Fibrosis. Nat. Genet. 2013, 45, 613–620. [Google Scholar] [CrossRef]
- Alder, J.K.; Armanios, M. Telomere-Mediated Lung Disease. Physiol. Rev. 2022, 102, 1703–1720. [Google Scholar] [CrossRef]
- Li, G.; Larregina, A.T.; Domsic, R.T.; Stolz, D.B.; Medsger, T.A., Jr.; Lafyatis, R.; Fuschiotti, P. Skin-Resident Effector Memory CD8+CD28− T Cells Exhibit a Profibrotic Phenotype in Patients with Systemic Sclerosis. J. Investig. Dermatol. 2017, 137, 1042–1050. [Google Scholar] [CrossRef]
- O’Donovan, A.; Pantell, M.S.; Puterman, E.; Dhabhar, F.S.; Blackburn, E.H.; Yaffe, K.; Cawthon, R.M.; Opresko, P.L.; Hsueh, W.-C.; Satterfield, S.; et al. Cumulative Inflammatory Load Is Associated with Short Leukocyte Telomere Length in the Health, Aging and Body Composition Study. PLoS ONE 2011, 6, e19687. [Google Scholar] [CrossRef]
- Rodier, F.; Coppé, J.-P.; Patil, C.K.; Hoeijmakers, W.A.M.; Muñoz, D.P.; Raza, S.R.; Freund, A.; Campeau, E.; Davalos, A.R.; Campisi, J. Persistent DNA Damage Signalling Triggers Senescence-Associated Inflammatory Cytokine Secretion. Nat. Cell Biol. 2009, 11, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lieberman, P.M. The Crosstalk of Telomere Dysfunction and Inflammation through Cell-Free TERRA Containing Exosomes. RNA Biol. 2016, 13, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Adler, B.L.; Boin, F.; Wolters, P.J.; Bingham, C.O.; Shah, A.A.; Greider, C.; Casciola-Rosen, L.; Rosen, A. Autoantibodies Targeting Telomere-Associated Proteins in Systemic Sclerosis. Ann. Rheum. Dis. 2021, 80, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base Platform Supports Systematic Causal Inference across the Human Phenome. Elife 2018, 7, e34408. [Google Scholar] [CrossRef]
- Staley, J.R.; Blackshaw, J.; Kamat, M.A.; Ellis, S.; Surendran, P.; Sun, B.B.; Paul, D.S.; Freitag, D.; Burgess, S.; Danesh, J.; et al. PhenoScanner: A Database of Human Genotype-Phenotype Associations. Bioinformatics 2016, 32, 3207–3209. [Google Scholar] [CrossRef]
- Kamat, M.A.; Blackshaw, J.A.; Young, R.; Surendran, P.; Burgess, S.; Danesh, J.; Butterworth, A.S.; Staley, J.R. PhenoScanner V2: An Expanded Tool for Searching Human Genotype-Phenotype Associations. Bioinformatics 2019, 35, 4851–4853. [Google Scholar] [CrossRef]
- Machiela, M.J.; Chanock, S.J. LDlink: A Web-Based Application for Exploring Population-Specific Haplotype Structure and Linking Correlated Alleles of Possible Functional Variants. Bioinformatics 2015, 31, 3555–3557. [Google Scholar] [CrossRef]
- Lin, S.-H.; Brown, D.W.; Machiela, M.J. LDtrait: An Online Tool for Identifying Published Phenotype Associations in Linkage Disequilibrium. Cancer Res. 2020, 80, 3443–3446. [Google Scholar] [CrossRef]
- Brion, M.-J.A.; Shakhbazov, K.; Visscher, P.M. Calculating Statistical Power in Mendelian Randomization Studies. Int. J. Epidemiol. 2013, 42, 1497–1501. [Google Scholar] [CrossRef]
- Burgess, S.; Scott, R.A.; Timpson, N.J.; Davey Smith, G.; Thompson, S.G.; EPIC-InterAct Consortium. Using Published Data in Mendelian Randomization: A Blueprint for Efficient Identification of Causal Risk Factors. Eur. J. Epidemiol. 2015, 30, 543–552. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Interpreting Findings from Mendelian Randomization Using the MR-Egger Method. Eur. J. Epidemiol. 2017, 32, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Verbanck, M.; Chen, C.-Y.; Neale, B.; Do, R. Detection of Widespread Horizontal Pleiotropy in Causal Relationships Inferred from Mendelian Randomization between Complex Traits and Diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, F.P.; Davey Smith, G.; Bowden, J. Robust Inference in Summary Data Mendelian Randomization via the Zero Modal Pleiotropy Assumption. Int. J. Epidemiol. 2017, 46, 1985–1998. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| LTL | SSc | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| SNP | CHR | BP | Gene | Effect Allele | Other Allele | β | p | β | p |
| rs11125529 | 2 | 54,475,866 | ACYP2 | C | A | −0.056 | 4.48 × 10−8 | −0.021 | 0.478 |
| rs10936599 | 3 | 169,492,101 | TERC | T | C | −0.097 | 2.54 × 10−31 | 0.051 | 0.043 |
| rs7675998 | 4 | 164,007,820 | NAF1 | A | G | −0.074 | 4.35 × 10−16 | −0.004 | 0.910 |
| rs2736100 | 5 | 1,286,516 | TERT | A | C | −0.078 | 4.38 × 10−19 | 0.034 | 0.145 |
| rs9420907 | 10 | 105,676,465 | OBFC1 | A | C | −0.069 | 6.90 × 10−11 | −0.002 | 0.974 |
| rs8105767 | 19 | 22,215,441 | ZNF208 | A | G | −0.048 | 1.11 × 10−9 | 0.039 | 0.297 |
| rs755017 | 20 | 62,421,622 | RTEL1 | A | G | −0.062 | 6.71 × 10−9 | 0.025 | 0.450 |
| MR Approach | nSNP | OR (95% CI) | p | p for Heterogeneity | p for Pleiotropy |
|---|---|---|---|---|---|
| Inverse-variance weighted FE | 7 | 0.716 (0.528–0.970) | 0.031 | 0.708 | NA |
| Maximum likelihood | 7 | 0.714 (0.525–0.970) | 0.031 | NA | NA |
| MR Egger | 7 | 0.397 (0.094–1.670) | 0.263 | 0.686 | 0.448 |
| Weighted median | 7 | 0.642 (0.438–0.941) | 0.023 | NA | NA |
| Weighted mode | 7 | 0.625 (0.411–0.952) | 0.071 | NA | NA |
| MR-PRESSO | 7 | 0.716 (0.563–0.911) | 0.035 | NA | NA |
| lcSSc | dcSSc | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| MR Approach | nSNP | OR (95% CI) | p | p for Heterogeneity | p for Pleiotropy | OR (95% CI) | p | p for Heterogeneity | p for Pleiotropy |
| Inverse-variance weighted FE | 7 | 0.669 (0.468–0.956) | 0.027 | 0.772 | NA | 0.771 (0.481–1.237) | 0.281 | 0.847 | NA |
| Maximum likelihood | 7 | 0.667 (0.466–0.957) | 0.028 | NA | NA | 0.770 (0.479–1.237) | 0.280 | NA | NA |
| MR Egger | 7 | 0.338 (0.062–1.845) | 0.266 | 0.755 | 0.457 | 0.389 (0.042–3.646) | 0.446 | 0.804 | 0.567 |
| Weighted median | 7 | 0.605 (0.393–0.932) | 0.023 | NA | NA | 0.647 (0.360–1.163) | 0.146 | NA | NA |
| Weighted mode | 7 | 0.574 (0.345–0.954) | 0.076 | NA | NA | 0.644 (0.329–1.262) | 0.247 | NA | NA |
| MR-PRESSO | 7 | 0.669 (0.514–0.872) | 0.025 | NA | NA | 0.771 (0.562–1.058) | 0.159 | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Martin, I.; Villanueva-Martin, G.; Guillen-Del-Castillo, A.; Ortego-Centeno, N.; Callejas, J.L.; Simeón-Aznar, C.P.; Martin, J.; Acosta-Herrera, M. Contribution of Telomere Length to Systemic Sclerosis Onset: A Mendelian Randomization Study. Int. J. Mol. Sci. 2023, 24, 15589. https://doi.org/10.3390/ijms242115589
Rodriguez-Martin I, Villanueva-Martin G, Guillen-Del-Castillo A, Ortego-Centeno N, Callejas JL, Simeón-Aznar CP, Martin J, Acosta-Herrera M. Contribution of Telomere Length to Systemic Sclerosis Onset: A Mendelian Randomization Study. International Journal of Molecular Sciences. 2023; 24(21):15589. https://doi.org/10.3390/ijms242115589
Chicago/Turabian StyleRodriguez-Martin, Inmaculada, Gonzalo Villanueva-Martin, Alfredo Guillen-Del-Castillo, Norberto Ortego-Centeno, José L. Callejas, Carmen P. Simeón-Aznar, Javier Martin, and Marialbert Acosta-Herrera. 2023. "Contribution of Telomere Length to Systemic Sclerosis Onset: A Mendelian Randomization Study" International Journal of Molecular Sciences 24, no. 21: 15589. https://doi.org/10.3390/ijms242115589