
Nutrient Solution Flowing Environment Affects Metabolite Synthesis Inducing Root Thigmomorphogenesis of Lettuce (Lactuca sativa L.) in Hydroponics

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
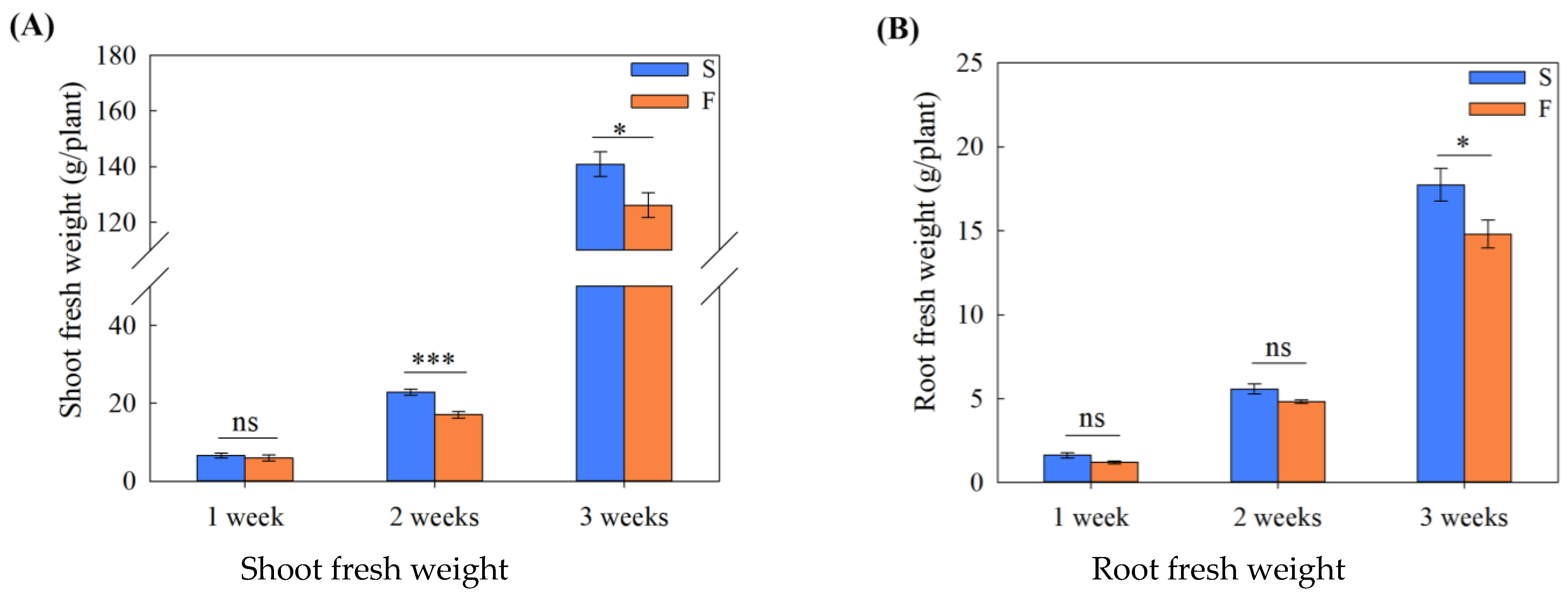
2.1. Effects of High-Flowing Environment on Plant Growth and Root Morphology with Growth Stage
2.2. Widely-Targeted Metabolite Detection
2.3. Key Differential Metabolites and Metabolic Pathways
3. Discussion
4. Materials and Methods
4.1. Cultivation
4.2. Measurement and Statistical Analysis
4.2.1. Fresh Weight and Root Morphology
4.2.2. Metabolite Extraction and Detection
4.2.3. Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pomoni, D.I.; Koukou, M.K.; Vrachopoulos, M.G.; Vasiliadis, L. A review of hydroponics and conventional agriculture based on energy and water consumption, environmental impact, and land use. Energies 2023, 16, 1690. [Google Scholar] [CrossRef]
- Khan, S.; Purohit, A.; Vadsaria, N. Hydroponics: Current and future state of the art in farming. J. Plant Nutr. 2021, 44, 1515–1538. [Google Scholar] [CrossRef]
- Singh, S.R.; Rajan, S. Vertical Hydroponics: A Future Technology for Urban Horticulture; Indian Horticulture; Indian Council of Agricultural Research: New Delhi, India, 2022. [Google Scholar]
- Baiyin, B.; Tagawa, K.; Yamada, M.; Wang, X.; Yamada, S.; Yamamoto, S.; Ibaraki, Y. Study on plant growth and nutrient uptake under different aeration intensity in hydroponics with the application of particle image velocimetry. Agriculture 2021, 11, 1140. [Google Scholar] [CrossRef]
- Baiyin, B.; Tagawa, K.; Yamada, M.; Wang, X.; Yamada, S.; Yamamoto, S.; Ibaraki, Y. Effect of the flow rate on plant growth and flow visualization of nutrient solution in hydroponics. Horticulturae 2021, 7, 225. [Google Scholar] [CrossRef]
- Baiyin, B.; Tagawa, K.; Yamada, M.; Wang, X.; Yamada, S.; Shao, Y.; An, P.; Yamamoto, S.; Ibaraki, Y. Effect of nutrient solution flow rate on hydroponic plant growth and root morphology. Plants. 2021, 10, 1840. [Google Scholar] [CrossRef]
- Chehab, E.W.; Eich, E.; Braam, J. Thigmomorphogenesis: A complex plant response to mechano-stimulation. J. Exp. Bot. 2009, 60, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Braam, J.; Chehab, E.W. Thigmomorphogenesis. Curr. Biol. 2017, 27, R863–R864. [Google Scholar] [CrossRef]
- Coutand, C. Mechanosensing and thigmomorphogenesis, a physiological and biomechanical point of view. Plant Sci. 2010, 179, 168–182. [Google Scholar] [CrossRef]
- Börnke, F.; Rocksch, T. Thigmomorphogenesis—Control of plant growth by mechanical stimulation. Sci. Hortic. 2018, 234, 344–353. [Google Scholar] [CrossRef]
- Ishihara, K.L.; Lee, E.K.W.; Borthakur, D. Thigmomorphogenesis: Changes in morphology, biochemistry, and levels of transcription in response to mechanical stress in Acacia koa. Can. J. For. Res. 2017, 47, 583–593. [Google Scholar] [CrossRef]
- Brenya, E.; Pervin, M.; Zhong-Hua, C.; Tissue, D.T.; Johnson, S.; Braam, J.; Cazzonelli, C.I. Mechanical stress acclimation in plants: Linking hormones and somatic memory to thigmomorphogenesis. Plant Cell Environ. 2022, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Barber, S.A.; Silberbush, M. Plant root morphology and nutrient uptake. ASA Spec. Publ. 1984, 49, 65–87. [Google Scholar]
- Baiyin, B.; Tagawa, K.; Yamada, M.; Wang, X.; Yamada, S.; Yamamoto, S.; Ibaraki, Y. Effect of substrate flow rate on nutrient uptake and use efficiency in hydroponically grown Swiss chard (Beta vulgaris L. ssp. cicla “Seiyou Shirokuki”). Agronomy 2021, 11, 2050. [Google Scholar] [CrossRef]
- Langer, M.; Hegge, E.; Speck, T.; Speck, O. Acclimation to wind loads and/or contact stimuli? A biomechanical study of peltate leaves of Pilea peperomioides. J. Exp. Bot. 2022, 73, 1236–1252. [Google Scholar] [CrossRef]
- Braam, J.; Davis, R.W. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell 1990, 60, 357–364. [Google Scholar] [CrossRef]
- Battaglia, M.; Lee, C.; Thomason, W.; Fike, J.; Sadeghpour, A. Hail damage impacts on corn productivity: A review. Crop Sci. 2019, 59, 1–14. [Google Scholar] [CrossRef]
- Braam, J. In touch: Plant responses to mechanical stimuli. New Phytol. 2005, 165, 373–389. [Google Scholar] [CrossRef]
- Biro, R.L.; Jaffe, M.J. Thigmomorphogenesis: Ethylene evolution and its role in the changes observed in mechanically perturbed bean plants. Physiologia Plantarum. 2010, 62, 289–296. [Google Scholar] [CrossRef]
- Jaffe, M.J.; Telewski, F.W.; Cooke, P.W. Thigmomorphogenesis: On the mechanical properties of mechanically perturbed bean plants. Physiol. Plant. 2010, 62, 73–78. [Google Scholar] [CrossRef]
- Telewski, F.W. Mechanosensing and plant growth regulators elicited during the thigmomorphogenetic response. Front. For. Glob. Change 2021, 3, 574096. [Google Scholar] [CrossRef]
- Wang, L.; Ma, C.; Wang, S.; Yang, F.; Sun, Y.; Tang, J.; Luo, J.; Wu, J. Ethylene and jasmonate signaling converge on gibberellin catabolism during thigmomorphogenesis in Arabidopsis. Plant Physiol. 2023, kiad556. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yang, Z.; Qing, D.; Ren, F.; Liu, S.; Zheng, Q.; Liu, J.; Zhang, W.; Dai, C.; Wu, M.; et al. Quantitative and functional posttranslational modification proteomics reveals that TREPH1 plays a role in plant touch-delayed bolting. Proc. Natl. Acad. Sci. USA 2018, 115, E10265–E10274. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, Y.; Lyu, M.; Luo, Y.; Shi, H.; Zhong, S. Touch-induced seedling morphological changes are determined by ethylene-regulated pectin degradation. Sci. Adv. 2020, 6, eabc9294. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, D.J.; Talon, G.-C. Plant phenotype demarcation using nontargeted lc-ms and gc-ms metabolite profiling. Arbona. J. Agric. Food Chem. 2009, 57, 7338–7347. [Google Scholar]
- Iason, G.; Moore, B.; Lennon, J.; Beaton, J. Plant Secondary Metabolite Polymorphisms and the Extended Chemical Phenotype. In The Ecology of Plant Secondary Metabolites: From Genes to Global Processes; Cambridge University Press: New York, NY, USA, 2012; pp. 247–268. [Google Scholar]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Zulak, K.G.; Liscombe, D.K.; Ashihara, H.; Facchini, P.J. Plant Secondary Metabolites; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root plasticity under abiotic stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis root development. Annu. Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef]
- Salazar-Henao, J.E.; Vélez-Bermúdez, I.C.; Schmidt, W. The regulation and plasticity of root hair patterning and morphogenesis. Development 2016, 143, 1848–1858. [Google Scholar] [CrossRef]
- Dehghanian, Z.; Habibi, K.; Dehghanian, M.; Aliyar, S.; Lajayer, B.A.; Astatkie, T.; Minkina, T.; Keswani, C. Reinforcing the bulwark: Unravelling the efficient applications of plant phenolics and tannins against environmental stresses. Heliyon 2022, 8, e09094. [Google Scholar] [CrossRef]
- Vaughan, D.; Ord, B. Influence of phenolic acids on morphological changes in roots of Pisum sativum. J. Sci. Food Agric. 1990, 52, 289–299. [Google Scholar] [CrossRef]
- Kawade, K.; Tabeta, H.; Ferjani, A.; Hirai, M.Y. The roles of functional amino acids in plant growth and development. Plant Cell Physiol. 2023, pcad071. [Google Scholar] [CrossRef]
- VanEtten, C.H.; Miller, R.W.; Wolff, I.A.; Jones, Q. Nutrients in seeds, amino acid composition of seeds from 200 angiospermous plant species. J. Agric. Food Chem. 1963, 11, 399–410. [Google Scholar] [CrossRef]
- Galili, G. New insights into the regulation and functional significance of lysine metabolism in plants. Annu. Rev. Plant Biol. 2002, 53, 27–43. [Google Scholar] [CrossRef]
- Baqir, H.A.; Zeboon, N.H.; Al-Behadili, A.A.J. The role and importance of amino acids within plants: A review. Plant Arch. 2019, 19, 1402–1410. [Google Scholar]
- Slocum, R.D. Genes, enzymes and regulation of arginine biosynthesis in plants. Plant Physiol. Biochem. 2005, 43, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, S.; Marathe, G.K. What we know about plant arginases? Plant Physiol. Biochem. 2020, 156, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Frémont, N.; Riefler, M.; Stolz, A.; Schmülling, T. The Arabidopsis TUMOR PRONE5 gene encodes an acetylornithine aminotransferase required for arginine biosynthesis and root meristem maintenance in blue light. Plant Physiol. 2013, 161, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Yamaji, N.; Ma, J.F. An appropriate concentration of arginine is required for normal root growth in rice. Plant Signal. Behav. 2014, 9, e28717. [Google Scholar] [CrossRef]
- Wen, P.; Li, J.-M.; Qun, L.; Xing, Z. Effects of amino acid on growth and secondary metabolites contents of adventitious roots of Tripterygium wilfordii. Zhongguo Zhong Yao Za Zhi = China J. Chin. Mater. Med. 2014, 12, 2267–2274. [Google Scholar]
- Glian’ko, A.K.; Ishchenko, A.A. Level nitric oxide (NO) and growth of roots of etiolated pea seedlings. Izv. Akad. Nauk Ser. Biol. 2013, 6, 689–695. [Google Scholar] [CrossRef]
- Reiter, C.D.; Tend, R.L.; Beckman, J.S. Superoxidereacts with nitric oxide to nitrate tyrosine at physiological pH peroxinitrite. J. Biol. Chem. 2000, 275, 32460–32466. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Hancock, J.T. Nitric oxide signaling in plants. New Phytol. 2003, 159, 11–35. [Google Scholar] [CrossRef]
- Pagnussat, G.C.; Lanteri, M.L.; Lombardo, M.C.; Lamattina, L. Nitric oxide mediates the indole acetic acid induction activation of a mitogen-activated protein kinase cascade involved in adventitious root development. Plant Physiol. 2004, 135, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Correa-Aragunde, N.; Graziano, M.; Lamattina, L. Nitric oxide plays a central role in determining lateral root development in tomato. Planta. 2004, 218, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Gouvêa, C.M.C.P.; Souza, J.F.; Magalhães, A.C.N.; Martins, I.S. NO- releasing substances that induce growth elongation in maize root segments. Plant Growth Regul. 1997, 21, 183–187. [Google Scholar] [CrossRef]
- Morris, S.M., Jr. Recent advances in arginine metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 45–51. [Google Scholar] [CrossRef]
- Morris, S.M., Jr. Enzymes of arginine metabolism. J. Nutr. 2004, 134, 2743S–2747S; discussion 2765S. [Google Scholar] [CrossRef]
- Satriano, J. Arginine pathways and the inflammatory response: Interregulation of nitric oxide and polyamines: Review article. Amino Acids 2004, 26, 321–329. [Google Scholar] [CrossRef]
- Gao, J.; Zhuang, S.; Zhang, Y.; Qian, Z. Exogenously applied spermidine alleviates hypoxia stress in Phyllostachys praecox seedlings via changes in endogenous hormones and gene expression. BMC Plant Biol. 2022, 22, 200. [Google Scholar] [CrossRef]
- Kalac, P.; Krausová, P.A. A review of dietary polyamines: Formation, implications for growth and health and occurrence in foods. Food Chem. 2005, 90, 219–230. [Google Scholar] [CrossRef]
- Yang, H.Q.; Jie, Y.L.; Jia, W.S.; Zhang, D.P. The signaling cascades of water loss to abscisic acid accumulation in apple roots. Acta Hortic. 2004, 636, 221–226. [Google Scholar]
- Takahashi, T.; Kakehi, J.I. Polyamines: Ubiquitous polycations with unique roles in growth and stress responses. Ann. Bot. 2010, 105, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Newton, R.J. Polyamines promote root elongation and growth by increasing root cell division in regenerated Virginia pine (Pinus virginiana Mill.) plantlets. Plant Cell Rep. 2005, 24, 581–589. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, T.; Wang, Q.; Guo, Y.; Zhang, P.; Xie, H.; Liu, J.; Li, L.; Zhang, C.; Qin, P. Mechanisms of Resistance to Spot Blotch in Yunnan Iron Shell Wheat Based on Metabolome and Transcriptomics. Int. J. Mol. Sci. 2022, 23, 5184. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, L.; Chen, S.; Qin, Y.; Ding, X.; Li, J.; Pan, Y.; Zhang, X. Pterostilbene production of tomato transformed with resveratrol synthase and resveratrol O-methyltransferase genes. Plant Sci. 2022, 322, 111343. [Google Scholar] [CrossRef]
- Yu, X.; Li, J.; Yang, Y.; Zhu, J.; Yuan, H.; Jiang, Y. Comprehensive investigation on flavonoids metabolites of Longjing tea in different cultivars, geographical origins, and storage time. Heliyon 2023, 9, e17305. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant. 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Wu, D.; Zhuang, F.; Wang, J.; Gao, R.; Zhang, Q.; Wang, X.; Zhang, G.; Fang, M.; Zhang, Y.; Li, Y.; et al. Metabolomics and Transcriptomics Revealed a Comprehensive Understanding of the Biochemical and Genetic Mechanisms Underlying the Color Variations in Chrysanthemums. Metabolites 2023, 13, 742. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Concentration |
|---|---|
| Ca(NO₃)₂·4H2O | 945 mg/L |
| KNO₃ | 607 mg/L |
| NH₄H₂PO₄ | 115 mg/L |
| MgSO₄·7H₂O | 493 mg/L |
| Na2Fe(EDTA) | 20–40 mg/L |
| H₃BO₃ | 2.86 mg/L |
| MnSO4·4H₂O | 2.13 mg/L |
| ZnSO₄·7H₂O | 0.22 mg/L |
| CuSO₄·5H₂O | 0.08 mg/L |
| (NH4)6Mo7O24·4H2O | 0.02 mg/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baiyin, B.; Xiang, Y.; Hu, J.; Tagawa, K.; Son, J.E.; Yamada, S.; Yang, Q. Nutrient Solution Flowing Environment Affects Metabolite Synthesis Inducing Root Thigmomorphogenesis of Lettuce (Lactuca sativa L.) in Hydroponics. Int. J. Mol. Sci. 2023, 24, 16616. https://doi.org/10.3390/ijms242316616
Baiyin B, Xiang Y, Hu J, Tagawa K, Son JE, Yamada S, Yang Q. Nutrient Solution Flowing Environment Affects Metabolite Synthesis Inducing Root Thigmomorphogenesis of Lettuce (Lactuca sativa L.) in Hydroponics. International Journal of Molecular Sciences. 2023; 24(23):16616. https://doi.org/10.3390/ijms242316616
Chicago/Turabian StyleBaiyin, Bateer, Yue Xiang, Jiangtao Hu, Kotaro Tagawa, Jung Eek Son, Satoshi Yamada, and Qichang Yang. 2023. "Nutrient Solution Flowing Environment Affects Metabolite Synthesis Inducing Root Thigmomorphogenesis of Lettuce (Lactuca sativa L.) in Hydroponics" International Journal of Molecular Sciences 24, no. 23: 16616. https://doi.org/10.3390/ijms242316616
APA StyleBaiyin, B., Xiang, Y., Hu, J., Tagawa, K., Son, J. E., Yamada, S., & Yang, Q. (2023). Nutrient Solution Flowing Environment Affects Metabolite Synthesis Inducing Root Thigmomorphogenesis of Lettuce (Lactuca sativa L.) in Hydroponics. International Journal of Molecular Sciences, 24(23), 16616. https://doi.org/10.3390/ijms242316616