
Fibroblast Growth Factor 21: A Fascinating Perspective on the Regulation of Muscle Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
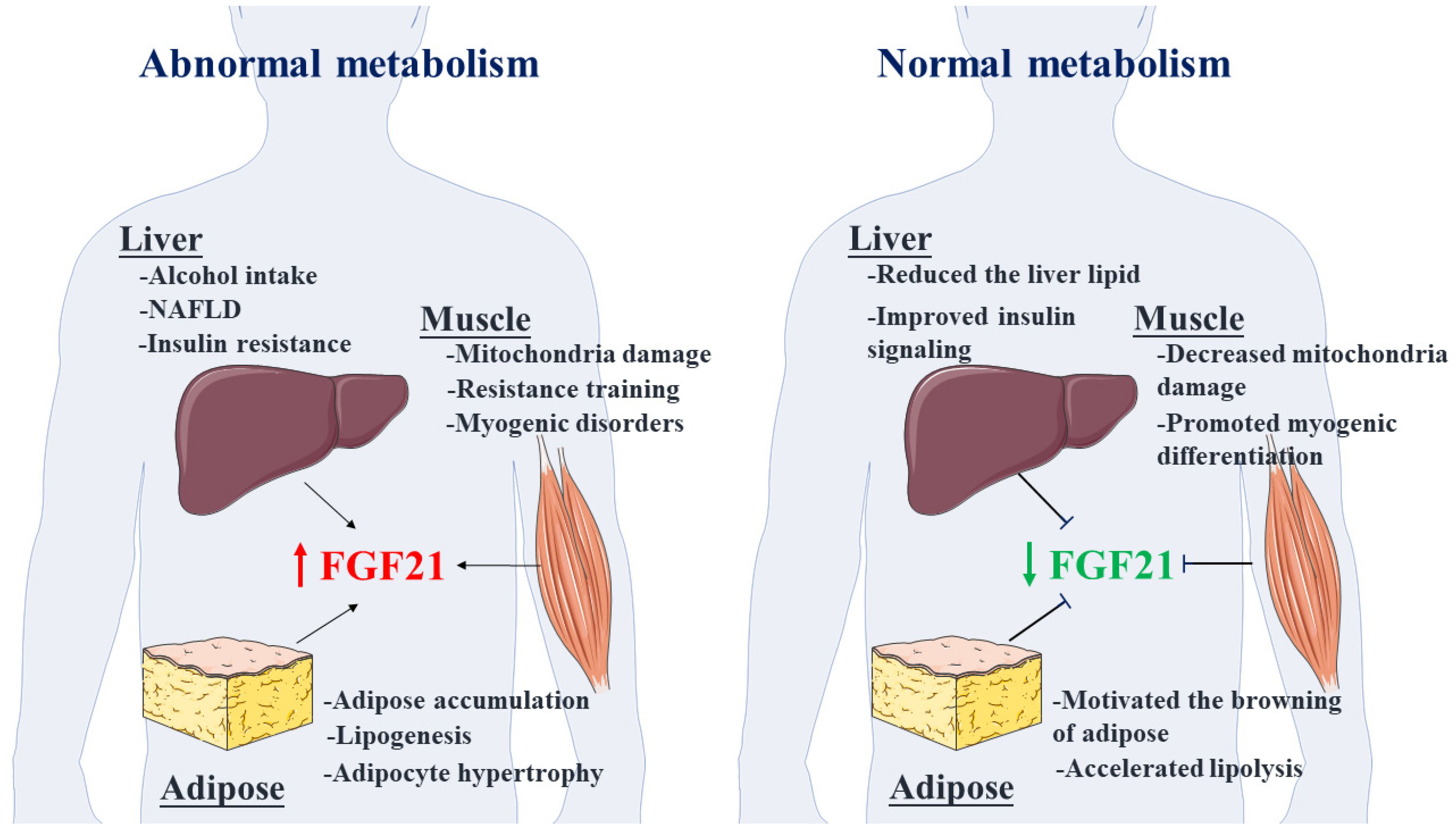
:1. Introduction
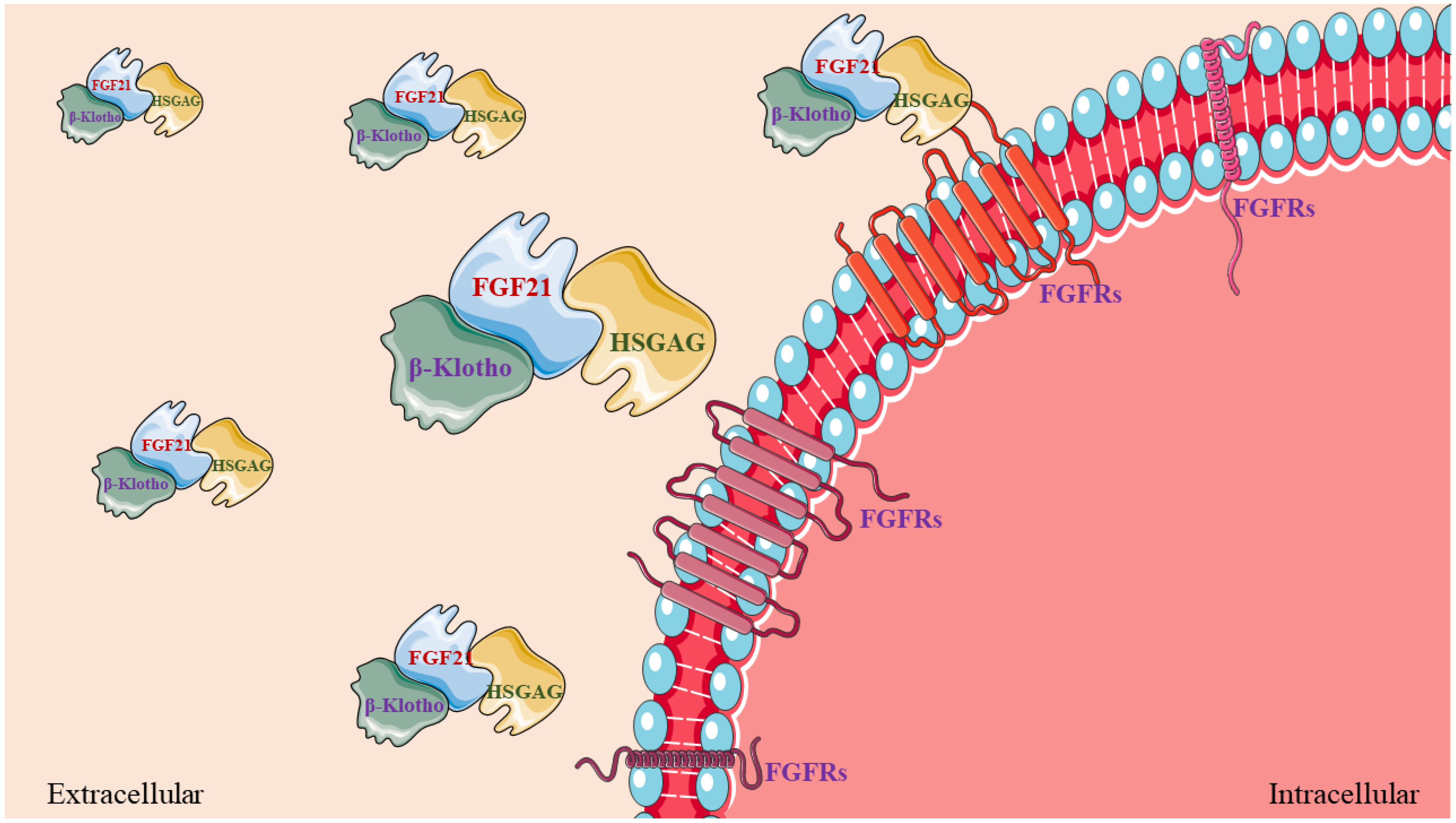
2. FGF21-Mediated Regulation of Metabolism Networks
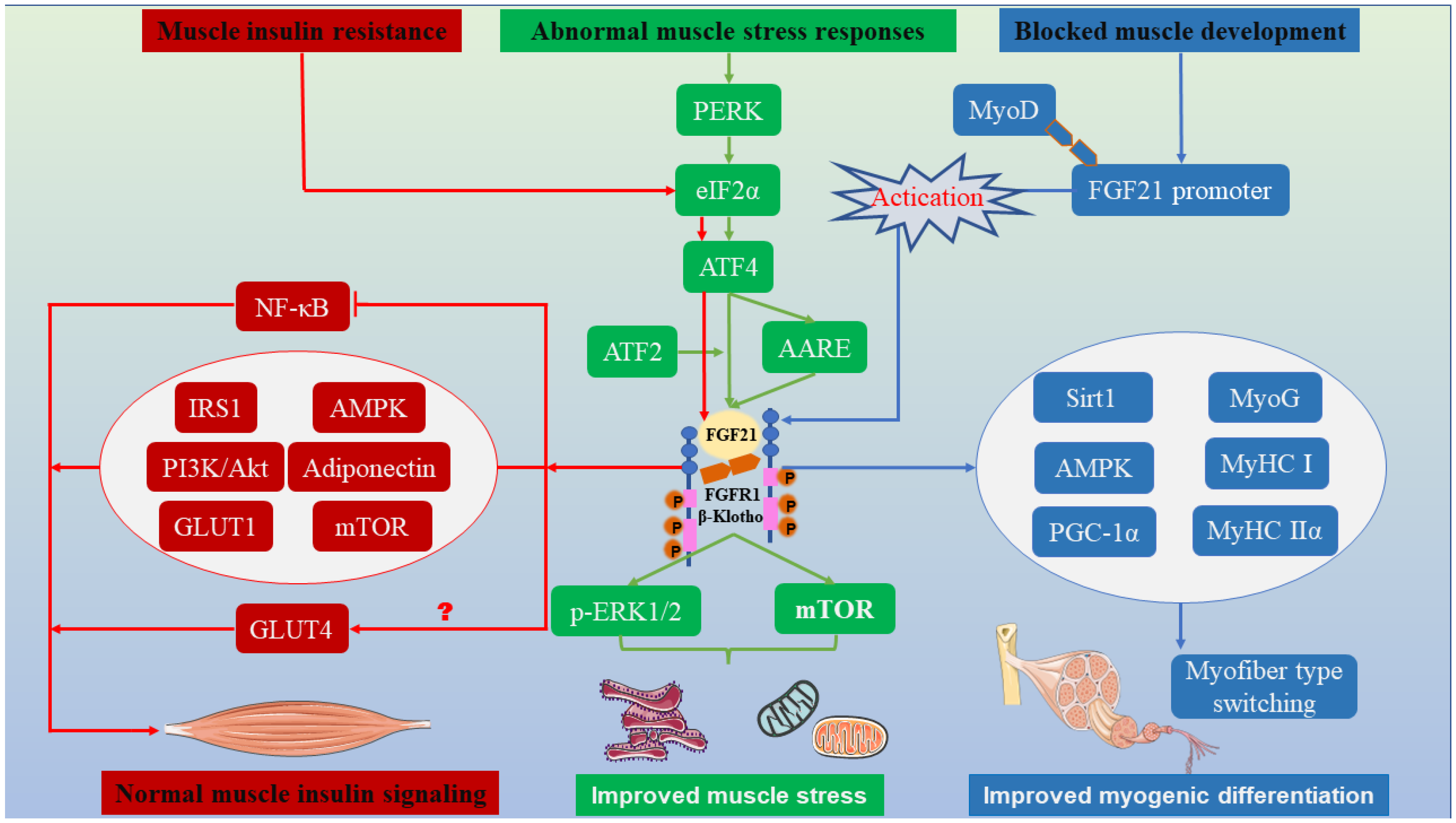
3. Muscle Stress Response
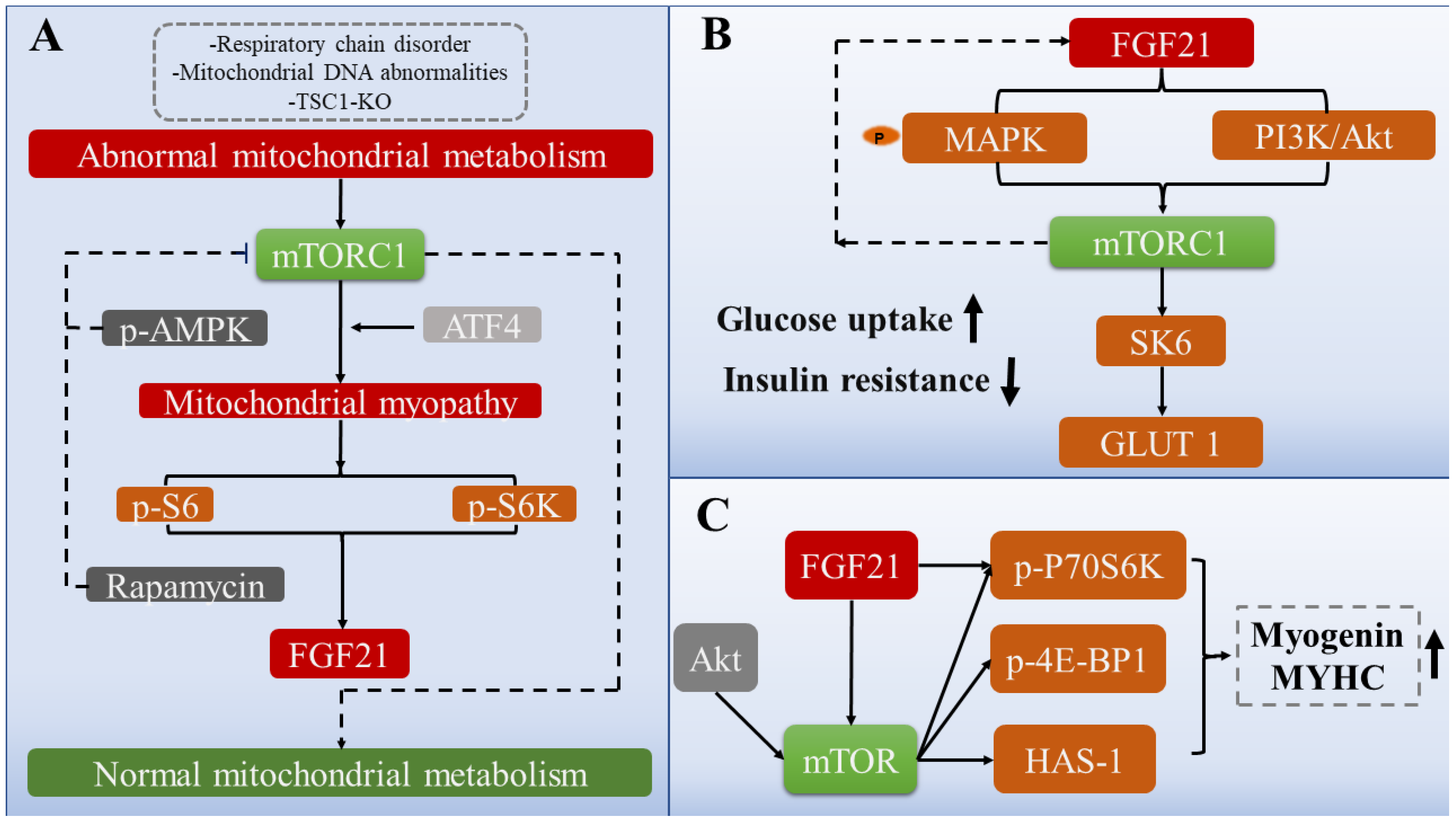
3.1. FGF21 Alleviates Muscle Mitochondrial Dysfunction
3.2. The Role of FGF21 in Endoplasmic Reticulum Stress in Muscle
4. Muscle Glucose Metabolism
4.1. FGF21 Preserves Muscle Glucose Metabolism in an Insulin-Dependent Manner
4.2. FGF21 Preserves Muscle Glucose Metabolism in an Insulin-Independent Manner
5. Muscle Development
FGF21 Regulates Multiple Pathways to Mediate Myogenic Differentiation
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip Rev. Syst. Biol. Med. 2020, 12, e1462. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, M.A.A. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef] [PubMed]
- Damluji, A.A.; Alfaraidhy, M.; AlHajri, N.; Rohant, N.N.; Kumar, M.; Al Malouf, C.; Bahrainy, S.; Ji Kwak, M.; Batchelor, W.B.; Forman, D.E.; et al. Sarcopenia and Cardiovascular Diseases. Circulation 2023, 147, 1534–1553. [Google Scholar] [CrossRef] [PubMed]
- Berezin, A.E.; Berezin, A.A.; Lichtenauer, M. Myokines and Heart Failure: Challenging Role in Adverse Cardiac Remodeling, Myopathy, and Clinical Outcomes. Dis. Markers 2021, 2021, 6644631. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Nakatake, Y.; Konishi, M.; Itoh, N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim. Biophys. Acta 2000, 1492, 203–206. [Google Scholar] [CrossRef]
- Jin, L.; Yang, R.; Geng, L.; Xu, A. Fibroblast Growth Factor-Based Pharmacotherapies for the Treatment of Obesity-Related Metabolic Complications. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 359–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, L.; Xu, X.; Tang, F.; Yi, P.; Qiu, B.; Hao, Y. A review of fibroblast growth factor 21 in diabetic cardiomyopathy. Heart Fail. Rev. 2019, 24, 1005–1017. [Google Scholar] [CrossRef]
- Liu, C.; Schonke, M.; Zhou, E.; Li, Z.; Kooijman, S.; Boon, M.R.; Larsson, M.; Wallenius, K.; Dekker, N.; Barlind, L.; et al. Pharmacological treatment with FGF21 strongly improves plasma cholesterol metabolism to reduce atherosclerosis. Cardiovasc. Res. 2022, 118, 489–502. [Google Scholar] [CrossRef]
- Li, S.; Zou, T.; Chen, J.; Li, J.; You, J. Fibroblast growth factor 21: An emerging pleiotropic regulator of lipid metabolism and the metabolic network. Genes Dis. 2023, 324, E409–E424. [Google Scholar] [CrossRef]
- Crooks, D.R.; Natarajan, T.G.; Jeong, S.Y.; Chen, C.; Park, S.Y.; Huang, H.; Ghosh, M.C.; Tong, W.H.; Haller, R.G.; Wu, C.; et al. Elevated FGF21 secretion, PGC-1alpha and ketogenic enzyme expression are hallmarks of iron-sulfur cluster depletion in human skeletal muscle. Hum. Mol. Genet. 2014, 23, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Chun, H.J.; Ahmad, K.; Shaikh, S.; Lim, J.H.; Ali, S.; Han, S.S.; Hur, S.J.; Sohn, J.H.; Lee, E.J.; et al. The roles of growth factors and hormones in the regulation of muscle satellite cells for cultured meat production. J. Anim. Sci. Technol. 2023, 65, 16–31. [Google Scholar] [CrossRef]
- Ribas, F.; Villarroya, J.; Hondares, E.; Giralt, M.; Villarroya, F. FGF21 expression and release in muscle cells: Involvement of MyoD and regulation by mitochondria-driven signalling. Biochem. J. 2014, 463, 191–199. [Google Scholar] [CrossRef]
- Arias-Calderón, M.; Casas, M.; Balanta-Melo, J.; Morales-Jiménez, C.; Hernández, N.; Llanos, P.; Jaimovich, E.; Buvinic, S. Fibroblast growth factor 21 is expressed and secreted from skeletal muscle following electrical stimulation via extracellular ATP activation of the PI3K/Akt/mTOR signaling pathway. Front. Endocrinol. 2023, 14, 1059020. [Google Scholar] [CrossRef]
- Li, H.; Sun, H.; Qian, B.; Feng, W.; Carney, D.; Miller, J.; Hogan, M.V.; Wang, L. Increased Expression of FGF-21 Negatively Affects Bone Homeostasis in Dystrophin/Utrophin Double Knockout Mice. J. Bone Miner. Res. 2019, 35, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.T.; Wang, C.H.; Lin, Y.L.; Hsu, B.G. Association of serum fibroblast growth factor 21 levels with skeletal muscle mass and mortality in chronic hemodialysis patients. J. Formos. Med. Assoc. 2022, 121, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Vankrunkelsven, W.; Thiessen, S.; Derde, S.; Vervoort, E.; Derese, I.; Pintelon, I.; Matheussen, H.; Jans, A.; Goossens, C.; Langouche, L.; et al. Development of muscle weakness in a mouse model of critical illness: Does fibroblast growth factor 21 play a role? Skelet. Muscle 2023, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Hou, L.; Xiong, Y.; Zhao, S. Fibroblast Growth Factor 21 (FGF21) Promotes Formation of Aerobic Myofibers via the FGF21-SIRT1-AMPK-PGC1alpha Pathway. J. Cell. Physiol. 2017, 232, 1893–1906. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, D.; Sui, M.; Zhou, M.; Wang, B.; Qi, Q.; Wang, T.; Zhang, G.; Wan, F.; Zhang, B. CRISPRa-based activation of Fgf21 and Fndc5 ameliorates obesity by promoting adipocytes browning. Clin. Transl. Med. 2023, 13, e1326. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Huang, S.; Zheng, F.; Liu, Y. The CEBPA-FGF21 regulatory network may participate in the T2DM-induced skeletal muscle atrophy by regulating the autophagy-lysosomal pathway. Acta Diabetol. 2023, 60, 1491–1503. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, W.; Zeng, L.Q.; Bai, H.; Li, J.; Zhou, J.; Zhou, G.Y.; Fang, C.W.; Wang, F.; Qin, X.J. Exercise and dietary intervention ameliorate high-fat diet-induced NAFLD and liver aging by inducing lipophagy. Redox Biol. 2020, 36, 101635. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, M.; Wu, X.; Zhang, Y.; Xia, Y. Muscle-to-tumor crosstalk: The effect of exercise-induced myokine on cancer progression. Biochim. Et Biophys. Acta (BBA)—Rev. Cancer 2022, 1877, 188761. [Google Scholar] [CrossRef]
- Romanello, V.; Sandri, M. The connection between the dynamic remodeling of the mitochondrial network and the regulation of muscle mass. Cell. Mol. Life Sci. 2021, 78, 1305–1328. [Google Scholar] [CrossRef]
- Lehtonen, J.M.; Forsstrom, S.; Bottani, E.; Viscomi, C.; Baris, O.R.; Isoniemi, H.; Hockerstedt, K.; Osterlund, P.; Hurme, M.; Jylhava, J.; et al. FGF21 is a biomarker for mitochondrial translation and mtDNA maintenance disorders. Neurology 2016, 87, 2290–2299. [Google Scholar] [CrossRef]
- Ferrer-Curriu, G.; Guitart-Mampel, M.; Rupérez, C.; Zamora, M.; Crispi, F.; Villarroya, F.; Fernández-Solà, J.; Garrabou, G.; Planavila, A. The protective effect of fibroblast growth factor-21 in alcoholic cardiomyopathy: A role in protecting cardiac mitochondrial function. J. Pathol. 2020, 253, 198–208. [Google Scholar] [CrossRef]
- Chen, L.; Fu, L.; Sun, J.; Huang, Z.; Fang, M.; Zinkle, A.; Liu, X.; Lu, J.; Pan, Z.; Wang, Y.; et al. Structural basis for FGF hormone signalling. Nature 2023, 618, 862–870. [Google Scholar] [CrossRef]
- Sun, H.; Sherrier, M.; Li, H. Skeletal Muscle and Bone—Emerging Targets of Fibroblast Growth Factor-21. Front. Physiol. 2021, 12, 625287. [Google Scholar] [CrossRef]
- Dolegowska, K.; Marchelek-Mysliwiec, M.; Nowosiad-Magda, M.; Slawinski, M.; Dolegowska, B. FGF19 subfamily members: FGF19 and FGF21. J. Physiol. Biochem. 2019, 75, 229–240. [Google Scholar] [CrossRef]
- Ying, L.; Wang, L.; Guo, K.; Hou, Y.; Li, N.; Wang, S.; Liu, X.; Zhao, Q.; Zhou, J.; Zhao, L.; et al. Paracrine FGFs target skeletal muscle to exert potent anti-hyperglycemic effects. Nat. Commun. 2021, 12, 7256. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, Y.; Liu, Y.; Zhang, B. Moderate-Intensity Continuous Training Improves FGF21 and KLB Expression in Obese Mice. Biochemistry 2020, 85, 938–946. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Prado Neto, E.V.; De Alvares Goulart, R.; Bechara, M.D.; Baisi Chagas, E.F.; Audi, M.; Guissoni Campos, L.M.; Landgraf Guiger, E.; Buchaim, R.L.; Buchaim, D.V.; et al. Myokines: A descriptive review. J. Sports Med. Phys. Fit. 2020, 60, 1583–1590. [Google Scholar] [CrossRef]
- Balboa-Vasquez, J.; Dominguez-Reyes, A.; Escandon-San Martin, Y.; Pavicic, F.; Ehrenfeld, P.; Martinez-Huenchullan, S. Relationship between fibroblast growth factor-21, muscle mass, and function outcomes in overweight and obese older adults living in the community. An exploratory study. Rev. Esp. Geriatr. Gerontol. 2021, 56, 81–86. [Google Scholar] [CrossRef]
- Meng, F.; Cao, Y.; Khoso, M.H.; Kang, K.; Ren, G.; Xiao, W.; Li, D. Therapeutic effect and mechanism of combined use of FGF21 and insulin on diabetic nephropathy. Arch. Biochem. Biophys. 2021, 713, 109063. [Google Scholar] [CrossRef]
- Hojman, P.; Pedersen, M.; Nielsen, A.R.; Krogh-Madsen, R.; Yfanti, C.; Akerstrom, T.; Nielsen, S.; Pedersen, B.K. Fibroblast growth factor-21 is induced in human skeletal muscles by hyperinsulinemia. Diabetes 2009, 58, 2797–2801. [Google Scholar] [CrossRef]
- Nasser, S.; Solé, T.; Vega, N.; Thomas, T.; Balcerczyk, A.; Strigini, M.; Pirola, L. Ketogenic diet administration to mice after a high-fat-diet regimen promotes weight loss, glycemic normalization and induces adaptations of ketogenic pathways in liver and kidney. Mol. Metab. 2022, 65, 101578. [Google Scholar] [CrossRef]
- Tillman, E.J.; Rolph, T. FGF21: An Emerging Therapeutic Target for Non-Alcoholic Steatohepatitis and Related Metabolic Diseases. Front. Endocrinol. 2020, 11, 601290. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Ma, C.; Lin, J.; Du, J. Alternate-day fasting alleviates high fat diet induced non-alcoholic fatty liver disease through controlling PPARalpha/Fgf21 signaling. Mol. Biol. Rep. 2022, 49, 3113–3122. [Google Scholar] [CrossRef]
- Miyake, M.; Nomura, A.; Ogura, A.; Takehana, K.; Kitahara, Y.; Takahara, K.; Tsugawa, K.; Miyamoto, C.; Miura, N.; Sato, R.; et al. Skeletal muscle-specific eukaryotic translation initiation factor 2alpha phosphorylation controls amino acid metabolism and fibroblast growth factor 21-mediated non-cell-autonomous energy metabolism. FASEB J. 2016, 30, 798–812. [Google Scholar] [CrossRef]
- Kim, C.S.; Joe, Y.; Choi, H.S.; Back, S.H.; Park, J.W.; Chung, H.T.; Roh, E.; Kim, M.S.; Ha, T.Y.; Yu, R. Deficiency of fibroblast growth factor 21 aggravates obesity-induced atrophic responses in skeletal muscle. J. Inflamm. 2019, 16, 17. [Google Scholar] [CrossRef]
- Xu, Q.; Lin, S.; Li, Q.; Lin, Y.; Xiong, Y.; Zhu, J.; Wang, Y. Fibroblast growth factor 21 regulates lipid accumulation and adipogenesis in goat intramuscular adipocyte. Anim. Biotechnol. 2021, 32, 318–326. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Hou, L.; Wu, W.; Zhao, S.; Xiong, Y. Fibroblast Growth Factor 21 Suppresses Adipogenesis in Pig Intramuscular Fat Cells. Int. J. Mol. Sci. 2015, 17, 11. [Google Scholar] [CrossRef] [PubMed]
- Croon, M.; Szczepanowska, K.; Popovic, M.; Lienkamp, C.; Senft, K.; Brandscheid, C.P.; Bock, T.; Gnatzy-Feik, L.; Ashurov, A.; Acton, R.J.; et al. FGF21 modulates mitochondrial stress response in cardiomyocytes only under mild mitochondrial dysfunction. Sci. Adv. 2022, 8, eabn7105. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, J.M.; Auranen, M.; Darin, N.; Sofou, K.; Bindoff, L.; Hikmat, O.; Uusimaa, J.; Vieira, P.; Tulinius, M.; Lonnqvist, T.; et al. Diagnostic value of serum biomarkers FGF21 and GDF15 compared to muscle sample in mitochondrial disease. J. Inherit. Metab. Dis. 2021, 44, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Huddar, A.; Govindaraj, P.; Chiplunkar, S.; Deepha, S.; Jessiena Ponmalar, J.N.; Philip, M.; Nagappa, M.; Narayanappa, G.; Mahadevan, A.; Sinha, S.; et al. Serum fibroblast growth factor 21 and growth differentiation factor 15: Two sensitive biomarkers in the diagnosis of mitochondrial disorders. Mitochondrion 2021, 60, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Torre-Villalvazo, I.; Aleman-Escondrillas, G.; Valle-Rios, R.; Noriega, L.G. Protein intake and amino acid supplementation regulate exercise recovery and performance through the modulation of mTOR, AMPK, FGF21, and immunity. Nutr. Res. 2019, 72, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Battaglioni, S.; Benjamin, D.; Walchli, M.; Maier, T.; Hall, M.N. mTOR substrate phosphorylation in growth control. Cell 2022, 185, 1814–1836. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, B.; Li, L.; Mitchell, S.E.; Green, C.L.; D’Agostino, G.; Wang, G.; Wang, L.; Li, M.; Li, J.; et al. Very-low-protein diets lead to reduced food intake and weight loss, linked to inhibition of hypothalamic mTOR signaling, in mice. Cell Metab. 2021, 33, 888–904.e886. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Hu, W.; Zhang, L.; Jiang, F.; Mao, X.; Yang, G.; Li, L. FGF21 facilitates autophagy in prostate cancer cells by inhibiting the PI3K-Akt-mTOR signaling pathway. Cell Death Dis. 2021, 12, 303. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C.; Akimoto, T.; Blaauw, B. Molecular Mechanisms of Skeletal Muscle Hypertrophy. J. Neuromuscul. Dis. 2021, 8, 169–183. [Google Scholar] [CrossRef]
- Cangelosi, A.L.; Puszynska, A.M.; Roberts, J.M.; Armani, A.; Nguyen, T.P.; Spinelli, J.B.; Kunchok, T.; Wang, B.; Chan, S.H.; Lewis, C.A.; et al. Zonated leucine sensing by Sestrin-mTORC1 in the liver controls the response to dietary leucine. Science 2022, 377, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Kuang, Y.; Bo, W.; Liang, Q.; Zhu, W.; Cai, M.; Tian, Z. Exercise Training Alleviates Cardiac Fibrosis through Increasing Fibroblast Growth Factor 21 and Regulating TGF-beta1-Smad2/3-MMP2/9 Signaling in Mice with Myocardial Infarction. Int. J. Mol. Sci. 2021, 22, 12341. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, N.; Koitabashi, N.; Matsui, H.; Sunaga, H.; Umbarawan, Y.; Syamsunarno, M.; Yamaguchi, A.; Obokata, M.; Hanaoka, H.; Yokoyama, T.; et al. DPP-4 inhibitor induces FGF21 expression via sirtuin 1 signaling and improves myocardial energy metabolism. Heart Vessel. 2021, 36, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Jia, C.; Wu, D.; Jin, H.; Lin, Z.; Pan, J.; Li, X.; Wang, W. Fibroblast growth factor 21 (FGF21) alleviates senescence, apoptosis, and extracellular matrix degradation in osteoarthritis via the SIRT1-mTOR signaling pathway. Cell Death Dis. 2021, 12, 865. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Zheng, J.; Lv, J.; Xu, J.; Ji, X.; Luo, Y.B.; Li, W.; Zhao, Y.; Yan, C. Skeletal muscle increases FGF21 expression in mitochondrial disorders to compensate for energy metabolic insufficiency by activating the mTOR-YY1-PGC1alpha pathway. Free Radic. Biol. Med. 2015, 84, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as Modulator of Metabolism in Health and Disease. Front. Physiol. 2019, 10, 419. [Google Scholar] [CrossRef]
- Post, A.; Groothof, D.; Schutten, J.C.; Kelly, D.; Swarte, J.C.; Flores-Guerrero, J.L.; van der Veen, Y.; Kema, I.P.; Ozyilmaz, A.; Enya, A.; et al. Fibroblast growth factor 21 and protein energy wasting in hemodialysis patients. Clin. Nutr. 2021, 40, 4216–4224. [Google Scholar] [CrossRef]
- Stone, K.P.; Wanders, D.; Orgeron, M.; Cortez, C.C.; Gettys, T.W. Mechanisms of increased in vivo insulin sensitivity by dietary methionine restriction in mice. Diabetes 2014, 63, 3721–3733. [Google Scholar] [CrossRef]
- Jung, H.W.; Park, J.H.; Kim, D.A.; Jang, I.Y.; Park, S.J.; Lee, J.Y.; Lee, S.; Kim, J.H.; Yi, H.S.; Lee, E.; et al. Association between serum FGF21 level and sarcopenia in older adults. Bone 2021, 145, 115877. [Google Scholar] [CrossRef]
- Delaye, J.B.; Lanznaster, D.; Veyrat-Durebex, C.; Fontaine, A.; Bacle, G.; Lefevre, A.; Hergesheimer, R.; Lecron, J.C.; Vourc’h, P.; Andres, C.R.; et al. Behavioral, Hormonal, Inflammatory, and Metabolic Effects Associated with FGF21-Pathway Activation in an ALS Mouse Model. Neurotherapeutics 2020, 18, 297–308. [Google Scholar] [CrossRef]
- Khan, N.A.; Nikkanen, J.; Yatsuga, S.; Jackson, C.; Wang, L.; Pradhan, S.; Kivela, R.; Pessia, A.; Velagapudi, V.; Suomalainen, A. mTORC1 Regulates Mitochondrial Integrated Stress Response and Mitochondrial Myopathy Progression. Cell Metab. 2017, 26, 419–428 e415. [Google Scholar] [CrossRef]
- Elia, I.; Realini, G.; Di Mauro, V.; Borghi, S.; Bottoni, L.; Tornambe, S.; Vitiello, L.; Weiss, S.J.; Chiariello, M.; Tamburrini, A.; et al. SNAI1 is upregulated during muscle regeneration and represses FGF21 and ATF3 expression by directly binding their promoters. FASEB J. 2022, 36, e22401. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Soltany, A.; Visavadiya, N.P.; Burtscher, M.; Millet, G.P.; Khoramipour, K.; Khamoui, A.V. Mitochondrial stress and mitokines in aging. Aging Cell 2023, 22, e13770. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Mokhtar, R.; Bayliss, J.; Parkington, H.C.; Suturin, V.M.; Bruce, C.R.; Watt, M.J. Perilipin 5 Deletion Unmasks an Endoplasmic Reticulum Stress-Fibroblast Growth Factor 21 Axis in Skeletal Muscle. Diabetes 2018, 67, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Romanello, V.; Sandri, M. Implications of mitochondrial fusion and fission in skeletal muscle mass and health. Semin. Cell Dev. Biol. 2023, 143, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Song, W. Resistance training increases fibroblast growth factor-21 and irisin levels in the skeletal muscle of Zucker diabetic fatty rats. J. Exerc. Nutr. Biochem. 2017, 21, 50–54. [Google Scholar] [CrossRef]
- Keipert, S.; Ost, M.; Johann, K.; Imber, F.; Jastroch, M.; van Schothorst, E.M.; Keijer, J.; Klaus, S. Skeletal muscle mitochondrial uncoupling drives endocrine cross-talk through the induction of FGF21 as a myokine. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E469–E482. [Google Scholar] [CrossRef]
- Boardman, N.T.; Trani, G.; Scalabrin, M.; Romanello, V.; Wüst, R.C.I. Intracellular to Interorgan Mitochondrial Communication in Striated Muscle in Health and Disease. Endocr. Rev. 2023, 44, 668–692. [Google Scholar] [CrossRef]
- Yan, B.; Mei, Z.; Tang, Y.; Song, H.; Wu, H.; Jing, Q.; Zhang, X.; Yan, C.; Han, Y. FGF21-FGFR1 controls mitochondrial homeostasis in cardiomyocytes by modulating the degradation of OPA1. Cell Death Dis. 2023, 14. [Google Scholar] [CrossRef]
- Colina-Tenorio, L.; Horten, P.; Pfanner, N.; Rampelt, H. Shaping the mitochondrial inner membrane in health and disease. J. Intern. Med. 2020, 287, 645–664. [Google Scholar] [CrossRef]
- Rashid, T.; Nemazanyy, I.; Paolini, C.; Tatsuta, T.; Crespin, P.; de Villeneuve, D.; Brodesser, S.; Benit, P.; Rustin, P.; Baraibar, M.A.; et al. Lipin1 deficiency causes sarcoplasmic reticulum stress and chaperone-responsive myopathy. EMBO J. 2019, 38, e99576. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, L. Fibroblast growth factor 21, a stress regulator, inhibits Drp1 activation to alleviate skeletal muscle ischemia/reperfusion injury. Lab. Investig. 2022, 102, 979–988. [Google Scholar] [CrossRef]
- Fu, T.; Xu, Z.; Liu, L.; Guo, Q.; Wu, H.; Liang, X.; Zhou, D.; Xiao, L.; Liu, L.; Liu, Y.; et al. Mitophagy Directs Muscle-Adipose Crosstalk to Alleviate Dietary Obesity. Cell Rep. 2018, 23, 1357–1372. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wu, K.; Li, B.; Liu, Y. lncRNA UCA1 inhibits mitochondrial dysfunction of skeletal muscle in type 2 diabetes mellitus by sequestering miR-143-3p to release FGF21. Cell Tissue Res. 2023, 391, 561–575. [Google Scholar] [CrossRef]
- Maruyama, R.; Shimizu, M.; Hashidume, T.; Inoue, J.; Itoh, N.; Sato, R. FGF21 Alleviates Hepatic Endoplasmic Reticulum Stress under Physiological Conditions. J. Nutr. Sci. Vitaminol. 2018, 64, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Haider, A.; Alvarez-Guaita, A.; Bidault, G.; El-Sayed Moustafa, J.S.; Guiu-Jurado, E.; Tadross, J.A.; Warner, J.; Harrison, J.; Virtue, S.; et al. Combined genetic deletion of GDF15 and FGF21 has modest effects on body weight, hepatic steatosis and insulin resistance in high fat fed mice. Mol. Metab. 2022, 65, 101589. [Google Scholar] [CrossRef]
- Cadena Sandoval, M.; Heberle, A.M.; Rehbein, U.; Barile, C.; Ramos Pittol, J.M.; Thedieck, K. mTORC1 Crosstalk with Stress Granules in Aging and Age-Related Diseases. Front. Aging 2021, 2, 761333. [Google Scholar] [CrossRef]
- de la Cruz Lopez, K.G.; Toledo Guzman, M.E.; Sanchez, E.O.; Garcia Carranca, A. mTORC1 as a Regulator of Mitochondrial Functions and a Therapeutic Target in Cancer. Front. Oncol. 2019, 9, 1373. [Google Scholar] [CrossRef]
- Mounier, R.; Lantier, L.; Leclerc, J.; Sotiropoulos, A.; Foretz, M.; Viollet, B. Antagonistic control of muscle cell size by AMPK and mTORC1. Cell Cycle 2014, 10, 2640–2646. [Google Scholar] [CrossRef]
- Osataphan, S.; Macchi, C.; Singhal, G.; Chimene-Weiss, J.; Sales, V.; Kozuka, C.; Dreyfuss, J.M.; Pan, H.; Tangcharoenpaisan, Y.; Morningstar, J.; et al. SGLT2 inhibition reprograms systemic metabolism via FGF21-dependent and -independent mechanisms. JCI Insight 2019, 4, e123130. [Google Scholar] [CrossRef]
- Nakashima, K.; Ishida, A. AMP-activated Protein Kinase Activation Suppresses Protein Synthesis and mTORC1 Signaling in Chick Myotube Cultures. J. Poult. Sci. 2022, 59, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Vandanmagsar, B.; Warfel, J.D.; Wicks, S.E.; Ghosh, S.; Salbaum, J.M.; Burk, D.; Dubuisson, O.S.; Mendoza, T.M.; Zhang, J.; Noland, R.C.; et al. Impaired Mitochondrial Fat Oxidation Induces FGF21 in Muscle. Cell Rep. 2016, 15, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Castets, P.; Lin, S.; Rion, N.; Di Fulvio, S.; Romanino, K.; Guridi, M.; Frank, S.; Tintignac, L.A.; Sinnreich, M.; Ruegg, M.A. Sustained activation of mTORC1 in skeletal muscle inhibits constitutive and starvation-induced autophagy and causes a severe, late-onset myopathy. Cell Metab. 2013, 17, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Abdon, B.; Liang, Y.; da Luz Scheffer, D.; Torres, M.; Shrestha, N.; Reinert, R.B.; Lu, Y.; Pederson, B.; Bugarin-Lapuz, A.; Kersten, S.; et al. Muscle specific ER-associated degradation maintains postnatal muscle hypertrophy and systemic energy metabolism. JCI Insight 2023, 8, e170387. [Google Scholar] [CrossRef]
- Kreß, J.K.C.; Jessen, C.; Hufnagel, A.; Schmitz, W.; Xavier da Silva, T.N.; Ferreira dos Santos, A.; Mosteo, L.; Goding, C.R.; Friedmann Angeli, J.P.; Meierjohann, S. The integrated stress response effector ATF4 is an obligatory metabolic activator of NRF2. Cell Rep. 2023, 42, 112724. [Google Scholar] [CrossRef]
- Wan, X.S.; Lu, X.H.; Xiao, Y.C.; Lin, Y.; Zhu, H.; Ding, T.; Yang, Y.; Huang, Y.; Zhang, Y.; Liu, Y.L.; et al. ATF4- and CHOP-dependent induction of FGF21 through endoplasmic reticulum stress. Biomed. Res. Int. 2014, 2014, 807874. [Google Scholar] [CrossRef]
- Liang, P.; Zhong, L.; Gong, L.; Wang, J.; Zhu, Y.; Liu, W.; Yang, J. Fibroblast growth factor 21 protects rat cardiomyocytes from endoplasmic reticulum stress by promoting the fibroblast growth factor receptor 1-extracellular signalregulated kinase 1/2 signaling pathway. Int. J. Mol. Med. 2017, 40, 1477–1485. [Google Scholar] [CrossRef]
- Tang, H.; Inoki, K.; Brooks, S.V.; Okazawa, H.; Lee, M.; Wang, J.; Kim, M.; Kennedy, C.L.; Macpherson, P.C.D.; Ji, X.; et al. mTORC1 underlies age-related muscle fiber damage and loss by inducing oxidative stress and catabolism. Aging Cell 2019, 18, e12943. [Google Scholar] [CrossRef]
- Guridi, M.; Tintignac, L.A.; Lin, S.; Kupr, B.; Castets, P.; Ruegg, M.A. Activation of mTORC1 in skeletal muscle regulates whole-body metabolism through FGF21. Sci. Signal. 2015, 8, ra113. [Google Scholar] [CrossRef]
- Capelo-Diz, A.; Lachiondo-Ortega, S.; Fernández-Ramos, D.; Cañas-Martín, J.; Goikoetxea-Usandizaga, N.; Serrano-Maciá, M.; González-Rellan, M.J.; Mosca, L.; Blazquez-Vicens, J.; Tinahones-Ruano, A.; et al. Hepatic levels of S-adenosylmethionine regulate the adaptive response to fasting. Cell Metab. 2023, 35, 1373–1389.e1378. [Google Scholar] [CrossRef] [PubMed]
- Warrier, M.; Paules, E.M.; Silva-Gomez, J.; Friday, W.B.; Bramlett, F.; Kim, H.; Zhang, K.; Trujillo-Gonzalez, I. Homocysteine-induced endoplasmic reticulum stress activates FGF21 and is associated with browning and atrophy of white adipose tissue in Bhmt knockout mice. Heliyon 2023, 9, e13216. [Google Scholar] [CrossRef] [PubMed]
- Post, A.; Dam, W.A.; Sokooti, S.; Groothof, D.; Gloerich, J.; van Gool, A.J.; Kremer, D.; Gansevoort, R.T.; van den Born, J.; Kema, I.P.; et al. Circulating FGF21 Concentration, Fasting Plasma Glucose, and the Risk of Type 2 Diabetes: Results From the PREVEND Study. J. Clin. Endocrinol. Metab. 2023, 108, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Chavez, A.O.; Molina-Carrion, M.; Abdul-Ghani, M.A.; Folli, F.; Defronzo, R.A.; Tripathy, D. Circulating fibroblast growth factor-21 is elevated in impaired glucose tolerance and type 2 diabetes and correlates with muscle and hepatic insulin resistance. Diabetes Care 2009, 32, 1542–1546. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Stanislaus, S.; Chinookoswong, N.; Lau, Y.Y.; Hager, T.; Patel, J.; Ge, H.; Weiszmann, J.; Lu, S.-C.; Graham, M.; et al. Acute glucose-lowering and insulin-sensitizing action of FGF21 in insulin-resistant mouse models—Association with liver and adipose tissue effects. Am. J. Physiol.-Endocrinol. Metab. 2009, 297, E1105–E1114. [Google Scholar] [CrossRef] [PubMed]
- Flippo, K.H.; Potthoff, M.J. Metabolic Messengers: FGF21. Nat. Metab. 2021, 3, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Stohr, O.; Tao, R.; Miao, J.; Copps, K.D.; White, M.F. FoxO1 suppresses Fgf21 during hepatic insulin resistance to impair peripheral glucose utilization and acute cold tolerance. Cell Rep. 2021, 34, 108893. [Google Scholar] [CrossRef]
- Lovadi, E.; Csereklyei, M.; Merkli, H.; FuLop, K.; Sebok, A.; Karcagi, V.; Komoly, S.; Pal, E. Elevated FGF 21 in myotonic dystrophy type 1 and mitochondrial diseases. Muscle Nerve 2017, 55, 564–569. [Google Scholar] [CrossRef]
- Yang, M.; Wei, D.; Mo, C.; Zhang, J.; Wang, X.; Han, X.; Wang, Z.; Xiao, H. Saturated fatty acid palmitate-induced insulin resistance is accompanied with myotube loss and the impaired expression of health benefit myokine genes in C2C12 myotubes. Lipids Health Dis. 2013, 12, 104. [Google Scholar] [CrossRef]
- Jeon, J.Y.; Choi, S.E.; Ha, E.S.; Kim, T.H.; Jung, J.G.; Han, S.J.; Kim, H.J.; Kim, D.J.; Kang, Y.; Lee, K.W. Association between insulin resistance and impairment of FGF21 signal transduction in skeletal muscles. Endocrine 2016, 53, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, H.J.; Song, J.H.; Shin, Y.K.; Abd El-Aty, A.M.; Ramadan, A.; Hacimuftuoglu, A.; Jeong, J.H.; Jung, T.W. Dimethyl itaconate attenuates palmitate-induced insulin resistance in skeletal muscle cells through the AMPK/FGF21/PPARdelta-mediated suppression of inflammation. Life Sci. 2021, 287, 120129. [Google Scholar] [CrossRef] [PubMed]
- Mashili, F.L.; Austin, R.L.; Deshmukh, A.S.; Fritz, T.; Caidahl, K.; Bergdahl, K.; Zierath, J.R.; Chibalin, A.V.; Moller, D.E.; Kharitonenkov, A.; et al. Direct effects of FGF21 on glucose uptake in human skeletal muscle: Implications for type 2 diabetes and obesity. Diabetes Metab. Res. Rev. 2011, 27, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Yamaguchi, K.; Seko, Y.; Okishio, S.; Ishiba, H.; Tochiki, N.; Takahashi, A.; Kataoka, S.; Okuda, K.; Liu, Y.; et al. Hepatocyte-specific fibroblast growth factor 21 overexpression ameliorates high-fat diet-induced obesity and liver steatosis in mice. Lab. Investig. 2022, 102, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Choi, S.E.; Ha, E.S.; An, S.Y.; Kim, T.H.; Han, S.J.; Kim, H.J.; Kim, D.J.; Kang, Y.; Lee, K.W. Fibroblast growth factor-21 protects human skeletal muscle myotubes from palmitate-induced insulin resistance by inhibiting stress kinase and NF-kappaB. Metabolism 2012, 61, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Tian, H.; Lam, K.S.; Lin, S.; Hoo, R.C.; Konishi, M.; Itoh, N.; Wang, Y.; Bornstein, S.R.; Xu, A.; et al. Adiponectin mediates the metabolic effects of FGF21 on glucose homeostasis and insulin sensitivity in mice. Cell Metab. 2013, 17, 779–789. [Google Scholar] [CrossRef]
- Zhang, L.; Li, G.; She, Y.; Zhang, Z. Low levels of spexin and adiponectin may predict insulin resistance in patients with non-alcoholic fatty liver. Pract. Lab. Med. 2021, 24, e00207. [Google Scholar] [CrossRef]
- Jung, J.G.; Yi, S.A.; Choi, S.E.; Kang, Y.; Kim, T.H.; Jeon, J.Y.; Bae, M.A.; Ahn, J.H.; Jeong, H.; Hwang, E.S.; et al. TM-25659-Induced Activation of FGF21 Level Decreases Insulin Resistance and Inflammation in Skeletal Muscle via GCN2 Pathways. Mol. Cells 2015, 38, 1037–1043. [Google Scholar] [CrossRef]
- Rosales-Soto, G.; Diaz-Vegas, A.; Casas, M.; Contreras-Ferrat, A.; Jaimovich, E. Fibroblast growth factor-21 potentiates glucose transport in skeletal muscle fibers. J. Mol. Endocrinol. 2020, 65, 85–95. [Google Scholar] [CrossRef]
- Xiang, C.; Zhang, Y.; Chen, Q.; Sun, A.; Peng, Y.; Zhang, G.; Zhou, D.; Xie, Y.; Hou, X.; Zheng, F.; et al. Increased glycolysis in skeletal muscle coordinates with adipose tissue in systemic metabolic homeostasis. J. Cell. Mol. Med. 2021, 25, 7840–7854. [Google Scholar] [CrossRef]
- Habtemichael, E.N.; Li, D.T.; Camporez, J.P.; Westergaard, X.O.; Sales, C.I.; Liu, X.; López-Giráldez, F.; DeVries, S.G.; Li, H.; Ruiz, D.M.; et al. Insulin-stimulated endoproteolytic TUG cleavage links energy expenditure with glucose uptake. Nat. Metab. 2021, 3, 378–393. [Google Scholar] [CrossRef]
- Minard, A.Y.; Tan, S.X.; Yang, P.; Fazakerley, D.J.; Domanova, W.; Parker, B.L.; Humphrey, S.J.; Jothi, R.; Stockli, J.; James, D.E. mTORC1 Is a Major Regulatory Node in the FGF21 Signaling Network in Adipocytes. Cell Rep. 2016, 17, 29–36. [Google Scholar] [CrossRef]
- Mu, J.; Pinkstaff, J.; Li, Z.; Skidmore, L.; Li, N.; Myler, H.; Dallas-Yang, Q.; Putnam, A.M.; Yao, J.; Bussell, S.; et al. FGF21 analogs of sustained action enabled by orthogonal biosynthesis demonstrate enhanced antidiabetic pharmacology in rodents. Diabetes 2012, 61, 505–512. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Phytochemicals from the Cocoa Shell Modulate Mitochondrial Function, Lipid and Glucose Metabolism in Hepatocytes via Activation of FGF21/ERK, AKT, and mTOR Pathways. Antioxidants 2022, 11, 136. [Google Scholar] [CrossRef]
- Berthou, F.; Sobolewski, C.; Abegg, D.; Fournier, M.; Maeder, C.; Dolicka, D.; Correia de Sousa, M.; Adibekian, A.; Foti, M. Hepatic PTEN Signaling Regulates Systemic Metabolic Homeostasis through Hepatokines-Mediated Liver-to-Peripheral Organs Crosstalk. Int. J. Mol. Sci. 2022, 23, 3959. [Google Scholar] [CrossRef]
- Mengeste, A.M.; Rustan, A.C.; Lund, J. Skeletal muscle energy metabolism in obesity. Obesity 2021, 29, 1582–1595. [Google Scholar] [CrossRef]
- Schnyder, S.; Handschin, C. Skeletal muscle as an endocrine organ: PGC-1alpha, myokines and exercise. Bone 2015, 80, 115–125. [Google Scholar] [CrossRef]
- Luo, Y.; McKeehan, W.L. Stressed Liver and Muscle Call on Adipocytes with FGF21. Front. Endocrinol. 2013, 4, 194. [Google Scholar] [CrossRef]
- Vicente-Garcia, C.; Hernandez-Camacho, J.D.; Carvajal, J.J. Regulation of myogenic gene expression. Exp. Cell Res. 2022, 419, 113299. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, C.; Gui, Y.; Zou, T.; Xi, S.; Guo, X. Fluoride regulates the differentiation and atrophy through FGF21/ERK signaling pathway in C2C12 cells. Ecotoxicol. Environ. Saf. 2023, 252, 114626. [Google Scholar] [CrossRef]
- Luo, X.; Zhang, H.; Cao, X.; Yang, D.; Yan, Y.; Lu, J.; Wang, X.; Wang, H. Endurance Exercise-Induced Fgf21 Promotes Skeletal Muscle Fiber Conversion through TGF-β1 and p38 MAPK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 11401. [Google Scholar] [CrossRef]
- Roh, E.; Hwang, S.Y.; Yoo, H.J.; Baik, S.H.; Cho, B.; Park, Y.S.; Kim, H.J.; Lee, S.G.; Kim, B.J.; Jang, H.C.; et al. Association of plasma FGF21 levels with muscle mass and muscle strength in a national multicentre cohort study: Korean Frailty and Aging Cohort Study. Age Ageing 2021, 50, 1971–1978. [Google Scholar] [CrossRef]
- Jia, W.H.; Wang, N.Q.; Yin, L.; Chen, X.; Hou, B.Y.; Wang, J.H.; Qiang, G.F.; Chan, C.B.; Yang, X.Y.; Du, G.H. Effects of fasting on the expression pattern of FGFs in different skeletal muscle fibre types and sexes in mice. Biol. Sex Differ. 2020, 11, 9. [Google Scholar] [CrossRef]
- Bazhan, N.; Jakovleva, T.; Feofanova, N.; Denisova, E.; Dubinina, A.; Sitnikova, N.; Makarova, E. Sex Differences in Liver, Adipose Tissue, and Muscle Transcriptional Response to Fasting and Refeeding in Mice. Cells 2019, 8, 1529. [Google Scholar] [CrossRef]
- Suomalainen, A.; Elo, J.M.; Pietiläinen, K.H.; Hakonen, A.H.; Sevastianova, K.; Korpela, M.; Isohanni, P.; Marjavaara, S.K.; Tyni, T.; Kiuru-Enari, S.; et al. FGF-21 as a biomarker for muscle-manifesting mitochondrial respiratory chain deficiencies: A diagnostic study. Lancet Neurol. 2011, 10, 806–818. [Google Scholar] [CrossRef]
- Cao, F.; Wang, S.; Cao, X.; Liu, X.; Fu, K.; Hao, P.; Liu, J. Fibroblast growth factor 21 attenuates calcification of vascular smooth muscle cells in vitro. J. Pharm. Pharmacol. 2017, 69, 1802–1816. [Google Scholar] [CrossRef]
- Li, Y.; He, S.; Wang, C.; Jian, W.; Shen, X.; Shi, Y.; Liu, J. Fibroblast growth factor 21 inhibits vascular calcification by ameliorating oxidative stress of vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2023, 650, 39–46. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Zhao, S.; Li, X. Fibroblast Growth Factor 21 Promotes C2C12 Cells Myogenic Differentiation by Enhancing Cell Cycle Exit. Biomed. Res. Int. 2017, 2017, 1648715. [Google Scholar] [CrossRef]
- Oost, L.J.; Kustermann, M.; Armani, A.; Blaauw, B.; Romanello, V. Fibroblast growth factor 21 controls mitophagy and muscle mass. J. Cachexia Sarcopenia Muscle 2019, 10, 630–642. [Google Scholar] [CrossRef]
- Carrasco-Rozas, A.; Fernandez-Simon, E.; Suarez-Calvet, X.; Pinol-Jurado, P.; Alonso-Perez, J.; de Luna, N.; Schoser, B.; Meinke, P.; Dominguez-Gonzalez, C.; Hernandez-Lain, A.; et al. BNIP3 Is Involved in Muscle Fiber Atrophy in Late-Onset Pompe Disease Patients. Am. J. Pathol. 2022, 192, 1151–1166. [Google Scholar] [CrossRef]
- Touvier, T.; De Palma, C.; Rigamonti, E.; Scagliola, A.; Incerti, E.; Mazelin, L.; Thomas, J.L.; D’Antonio, M.; Politi, L.; Schaeffer, L.; et al. Muscle-specific Drp1 overexpression impairs skeletal muscle growth via translational attenuation. Cell Death Dis. 2015, 6, e1663. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Araya, Y.; Stenhouse, C.; Salavati, M.; Dan-Jumbo, S.O.; Ho, W.; Ashworth, C.J.; Clark, E.; Esteves, C.L.; Donadeu, F.X. KLB dysregulation mediates disrupted muscle development in intrauterine growth restriction. J. Physiol. 2022, 600, 1771–1790. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Q.; Zhang, Z.; Fu, R.; Zhou, T.; Long, C.; He, T.; Yang, D.; Li, Z.; Peng, S. Magnesium supplementation enhances mTOR signalling to facilitate myogenic differentiation and improve aged muscle performance. Bone 2021, 146, 115886. [Google Scholar] [CrossRef]
- Benoit, B.; Meugnier, E.; Castelli, M.; Chanon, S.; Vieille-Marchiset, A.; Durand, C.; Bendridi, N.; Pesenti, S.; Monternier, P.A.; Durieux, A.C.; et al. Fibroblast growth factor 19 regulates skeletal muscle mass and ameliorates muscle wasting in mice. Nat. Med. 2017, 23, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.J. What’s So Special about FGF19-Unique Effects Reported on Skeletal Muscle Mass and Function. Cell Metab. 2017, 26, 287–288. [Google Scholar] [CrossRef]
- Kharitonenkov, A.; Larsen, P. FGF21 reloaded: Challenges of a rapidly growing field. Trends Endocrinol. Metab. 2011, 22, 81–86. [Google Scholar] [CrossRef]
- Tezze, C.; Romanello, V.; Desbats, M.A.; Fadini, G.P.; Albiero, M.; Favaro, G.; Ciciliot, S.; Soriano, M.E.; Morbidoni, V.; Cerqua, C.; et al. Age-Associated Loss of OPA1 in Muscle Impacts Muscle Mass, Metabolic Homeostasis, Systemic Inflammation, and Epithelial Senescence. Cell Metab. 2017, 25, 1374–1389 e1376. [Google Scholar] [CrossRef]
- de Oliveira Dos Santos, A.R.; de Oliveira Zanuso, B.; Miola, V.F.B.; Barbalho, S.M.; Santos Bueno, P.C.; Flato, U.A.P.; Detregiachi, C.R.P.; Buchaim, D.V.; Buchaim, R.L.; Tofano, R.J.; et al. Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. Int. J. Mol. Sci. 2021, 22, 2639. [Google Scholar] [CrossRef]
- Abel, E.D. Mitochondrial Dynamics and Metabolic Regulation in Cardiac and Skeletal Muscle. Trans. Am. Clin. Climatol. Assoc. 2018, 129, 266–278. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Chen, J.; Wei, P.; Zou, T.; You, J. Fibroblast Growth Factor 21: A Fascinating Perspective on the Regulation of Muscle Metabolism. Int. J. Mol. Sci. 2023, 24, 16951. https://doi.org/10.3390/ijms242316951
Li S, Chen J, Wei P, Zou T, You J. Fibroblast Growth Factor 21: A Fascinating Perspective on the Regulation of Muscle Metabolism. International Journal of Molecular Sciences. 2023; 24(23):16951. https://doi.org/10.3390/ijms242316951
Chicago/Turabian StyleLi, Shuo, Jun Chen, Panting Wei, Tiande Zou, and Jinming You. 2023. "Fibroblast Growth Factor 21: A Fascinating Perspective on the Regulation of Muscle Metabolism" International Journal of Molecular Sciences 24, no. 23: 16951. https://doi.org/10.3390/ijms242316951
APA StyleLi, S., Chen, J., Wei, P., Zou, T., & You, J. (2023). Fibroblast Growth Factor 21: A Fascinating Perspective on the Regulation of Muscle Metabolism. International Journal of Molecular Sciences, 24(23), 16951. https://doi.org/10.3390/ijms242316951