

Toward Transgene-Free Transposon-Mediated Biological Mutagenesis for Plant Breeding


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Genotype-Mediated TE Transposition Reactivation
2.1. Heterologous Expression of TEs via Plant Transformation
2.2. Implication of DNA-Methylation-Deficient Genotypes
2.3. Utilization of Mutagenic Plant Lines with Reactivated TEs
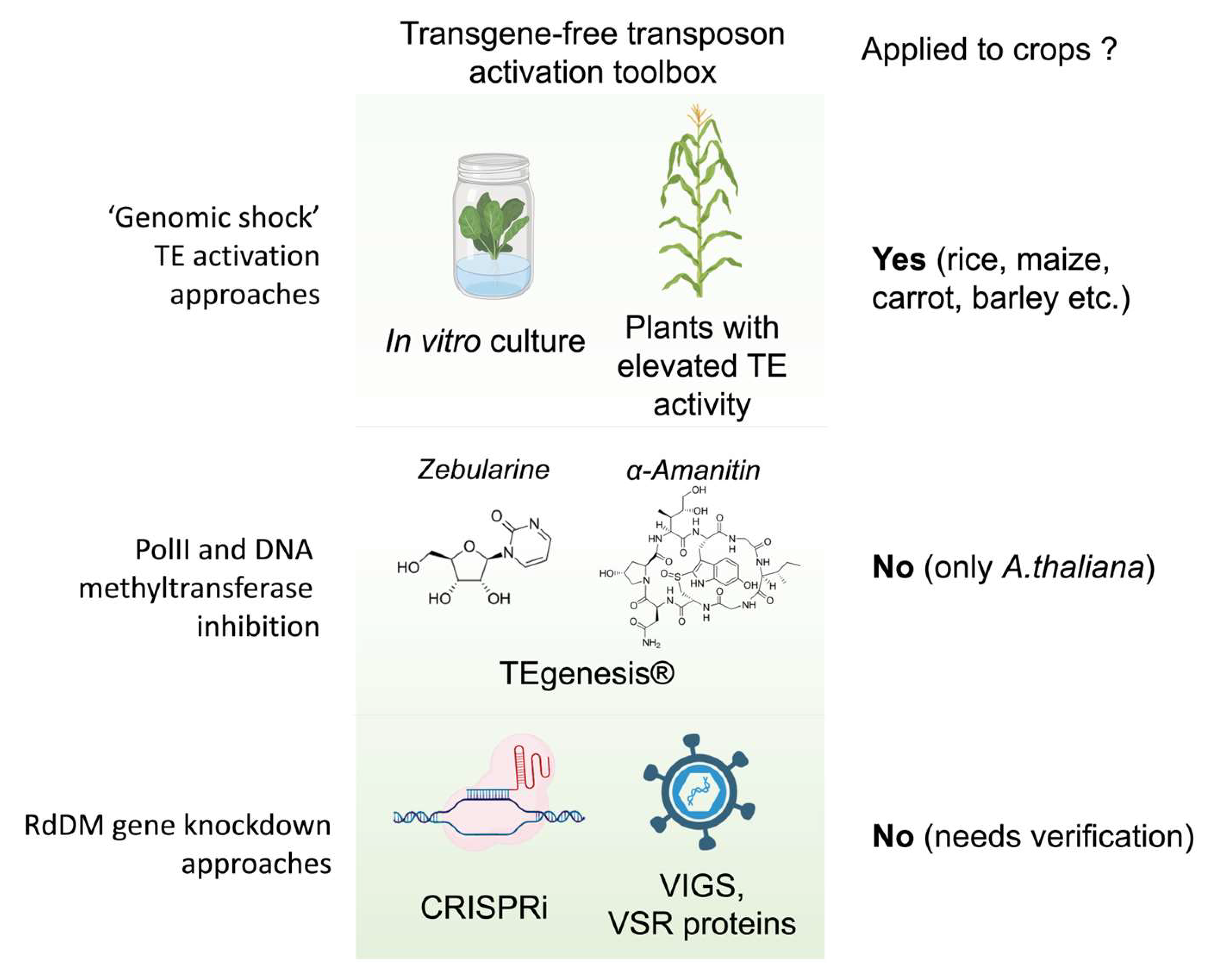
3. Controlled Activation of TEs via PolII and DNA Methylation Inhibition Using Toxins
4. Stress-Mediated TE Transposition Activation
5. Ensuring of Transgenerational Inheritance of Novel TE Insertions
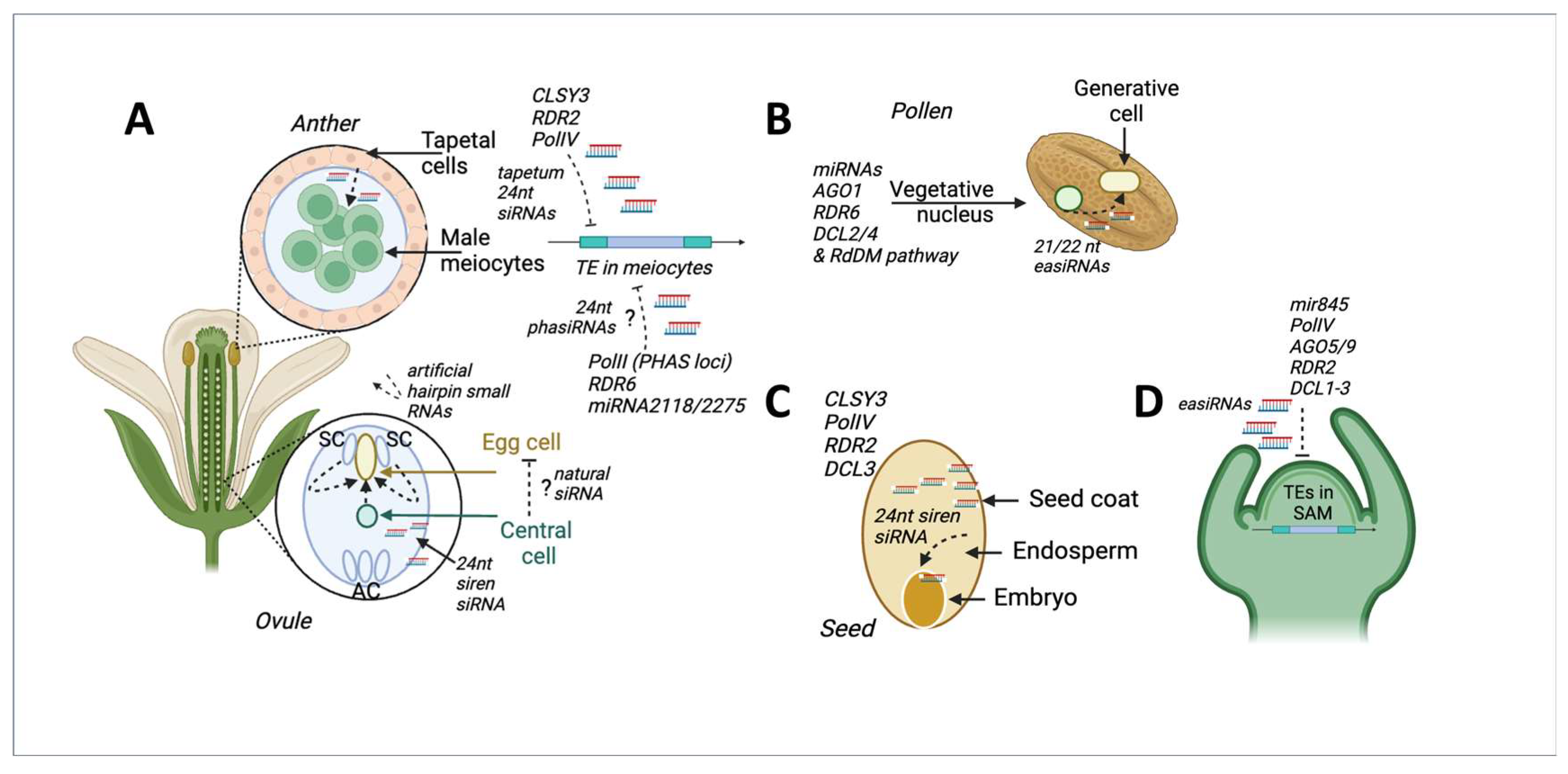
5.1. Tapetum-Derived Small RNAs Silence TEs in Meiocytes
5.2. TE Silencing in Pollen Generative Cell via easiRNAs Movement from Vegetative Cell
5.3. Putative Movement of Small RNAs to Silence TEs in the Egg Cell and Embryo
5.4. RdDM-Based TE Transposition Silencing in the Shoot Apical Meristem
6. Semi-Random Integration of TEs in Genome
7. Alternative Methods of DNA Methylation Reduction for Future Use in TE Activation
8. Challenges and Emerging Perspectives of TE-Mediated Biological Mutagenesis
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, L.; Kong, F.; Sun, K.; Wang, T.; Guo, T. From Classical Radiation to Modern Radiation: Past, Present, and Future of Radiation Mutation Breeding. Front. Public Health 2021, 9, 768071. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.R.; Zhang, X.; Rathjen, J.P. Precision Genome Editing of Crops for Improved Disease Resistance. Curr. Biol. 2023, 33, R650–R657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Massel, K.; Godwin, I.D.; Gao, C. Applications and Potential of Genome Editing in Crop Improvement. Genome Biol. 2018, 19, 210. [Google Scholar] [CrossRef] [PubMed]
- Belfield, E.J.; Brown, C.; Ding, Z.J.; Chapman, L.; Luo, M.; Hinde, E.; Es, S.W.; van Johnson, S.; Ning, Y.; Zheng, S.J.; et al. Thermal Stress Accelerates Arabidopsis Thaliana Mutation Rate. Genome Res. 2021, 31, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Cui, J.; Wang, L.; Teng, N.; Zhang, S.; Lam, H.-M.; Zhu, Y.; Xiao, S.; Ke, W.; Lin, J.; et al. Genome-Wide DNA Mutations in Arabidopsis Plants after Multigenerational Exposure to High Temperatures. Genome Biol. 2021, 22, 160. [Google Scholar] [CrossRef] [PubMed]
- Baduel, P.; Leduque, B.; Ignace, A.; Gy, I.; Gil, J.; Loudet, O.; Colot, V.; Quadrana, L. Genetic and Environmental Modulation of Transposition Shapes the Evolutionary Potential of Arabidopsis Thaliana. Genome Biol 2021, 22, 138. [Google Scholar] [CrossRef]
- Hämälä, T.; Ning, W.; Kuittinen, H.; Aryamanesh, N.; Savolainen, O. Environmental Response in Gene Expression and DNA Methylation Reveals Factors Influencing the Adaptive Potential of Arabidopsis Lyrata. eLife 2022, 11, e83115. [Google Scholar] [CrossRef]
- Quadrana, L.; Silveira, A.B.; Mayhew, G.F.; LeBlanc, C.; Martienssen, R.A.; Jeddeloh, J.A.; Colot, V. The Arabidopsis Thaliana Mobilome and Its Impact at the Species Level. eLife 2016, 5, e15716. [Google Scholar] [CrossRef]
- Merkulov, P.; Gvaramiya, S.; Dudnikov, M.; Komakhin, R.; Omarov, M.; Kocheshkova, A.; Konstantinov, Z.; Soloviev, A.; Karlov, G.; Divashuk, M.; et al. Cas9-targeted Nanopore Sequencing Rapidly Elucidates the Transposition Preferences and DNA Methylation Profiles of Mobile Elements in Plants. J. Integr. Plant Biol. 2023, 65, 2242–2261. [Google Scholar] [CrossRef]
- Quadrana, L.; Etcheverry, M.; Gilly, A.; Caillieux, E.; Madoui, M.-A.; Guy, J.; Silveira, A.B.; Engelen, S.; Baillet, V.; Wincker, P.; et al. Transposition Favors the Generation of Large Effect Mutations That May Facilitate Rapid Adaption. Nat. Commun. 2019, 10, 3421. [Google Scholar] [CrossRef]
- Paszkowski, J. Controlled Activation of Retrotransposition for Plant Breeding. Curr. Opin. Biotechnol. 2015, 32, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Dubin, M.J.; Scheid, O.M.; Becker, C. Transposons: A Blessing Curse. Curr. Opin. Plant Biol. 2018, 42, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, M.; Dugas, E.; Benchouaia, M.; Leduque, B.; Jiménez-Gómez, J.M.; Colot, V.; Quadrana, L. The Impact of Transposable Elements on Tomato Diversity. Nat. Commun. 2020, 11, 4058. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Lin, R.; Liang, J.; King, G.J.; Wu, J.; Wang, X. Transposable Element Insertion: A Hidden Major Source of Domesticated Phenotypic Variation in Brassica Rapa. Plant Biotechnol. J. 2022, 20, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Studer, A.; Zhao, Q.; Ross-Ibarra, J.; Doebley, J. Identification of a Functional Transposon Insertion in the Maize Domestication Gene Tb1. Nat. Genet. 2011, 43, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Cao, X. The Effect of Transposable Elements on Phenotypic Variation: Insights from Plants to Humans. Sci. China Life Sci. 2016, 59, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. How Important Are Transposons for Plant Evolution? Nat. Rev. Genet. 2013, 14, 49–61. [Google Scholar] [CrossRef]
- Latzel, V.; Puy, J.; Thieme, M.; Bucher, E.; Götzenberger, L.; Bello, F. Phenotypic Diversity Influenced by a Transposable Element Increases Productivity and Resistance to Competitors in Plant Populations. J. Ecol. 2023, 111, 2376–2387. [Google Scholar] [CrossRef]
- Lieberman-Lazarovich, M.; Kaiserli, E.; Bucher, E.; Mladenov, V. Natural and Induced Epigenetic Variation for Crop Improvement. Curr. Opin. Plant Biol. 2022, 70, 102297. [Google Scholar] [CrossRef]
- Thieme, M.; Lanciano, S.; Balzergue, S.; Daccord, N.; Mirouze, M.; Bucher, E. Inhibition of RNA Polymerase II Allows Controlled Mobilisation of Retrotransposons for Plant Breeding. Genome Biol. 2017, 18, 134. [Google Scholar] [CrossRef]
- Thieme, M.; Brêchet, A.; Bourgeois, Y.; Keller, B.; Bucher, E.; Roulin, A.C. Experimentally Heat-induced Transposition Increases Drought Tolerance in Arabidopsis Thaliana. New Phytol. 2022, 236, 182–194. [Google Scholar] [CrossRef]
- Roquis, D.; Robertson, M.; Yu, L.; Thieme, M.; Julkowska, M.; Bucher, E. Genomic Impact of Stress-Induced Transposable Element Mobility in Arabidopsis. Nucleic Acids Res. 2021, 49, gkab828. [Google Scholar] [CrossRef]
- Fedoroff, N.; Wessler, S.; Shure, M. Isolation of the Transposable Maize Controlling Elements Ac and Ds. Cell 1983, 35, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Schell, J.; Lörz, H.; Fedoroff, N. Transposition of the Maize Controlling Element “Activator” in Tobacco. Proc. Natl. Acad. Sci. USA 1986, 83, 4844–4848. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Tsugawa, H.; Miyao, A.; Yano, M.; Wu, J.; Yamamoto, S.; Matsumoto, T.; Sasaki, T.; Hirochika, H. The Rice Retrotransposon Tos17 Prefers Low-Copy-Number Sequences as Integration Targets. Mol. Genet. Genom. 2001, 265, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Hirochika, H. Efficient Insertion Mutagenesis of Arabidopsis by Tissue Culture-induced Activation of the Tobacco Retrotransposon Tto1. Plant J. 2000, 23, 291–304. [Google Scholar] [CrossRef]
- Courtial, B.; Feuerbach, F.; Eberhard, S.; Rohmer, L.; Chiapello, H.; Camilleri, C.; Lucas, H. Tnt1 Transposition Events Are Induced by in Vitro Transformation of Arabidopsis Thaliana, and Transposed Copies Integrate into Genes. Mol. Genet. Genom. 2001, 265, 32–42. [Google Scholar] [CrossRef]
- D’Erfurth, I.; Cosson, V.; Eschstruth, A.; Lucas, H.; Kondorosi, A.; Ratet, P. Efficient Transposition of the Tnt1 Tobacco Retrotransposon in the Model Legume Medicago Truncatula. Plant J. 2003, 34, 95–106. [Google Scholar] [CrossRef]
- Kim, E.Y.; Wang, L.; Lei, Z.; Li, H.; Fan, W.; Cho, J. Ribosome Stalling and SGS3 Phase Separation Prime the Epigenetic Silencing of Transposons. Nat. Plants 2021, 7, 303–309. [Google Scholar] [CrossRef]
- Nicolau, M.; Picault, N.; Moissiard, G. The Evolutionary Volte-Face of Transposable Elements: From Harmful Jumping Genes to Major Drivers of Genetic Innovation. Cells 2021, 10, 2952. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, M. How Transposable Elements Are Recognized and Epigenetically Silenced in Plants? Curr. Opin. Plant Biol. 2023, 75, 102428. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cuerda-Gil, D.; Shahid, S.; Slotkin, R.K. The Epigenetic Control of the Transposable Element Life Cycle in Plant Genomes and Beyond. Annu. Rev. Genet. 2022, 56, 63–87. [Google Scholar] [CrossRef]
- Panda, K.; Ji, L.; Neumann, D.A.; Daron, J.; Schmitz, R.J.; Slotkin, R.K. Full-Length Autonomous Transposable Elements Are Preferentially Targeted by Expression-Dependent Forms of RNA-Directed DNA Methylation. Genome Biol. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed]
- Catoni, M.; Cortijo, S. Chapter Four EpiRILs Lessons From Arabidopsis. Adv. Bot. Res. 2018, 88, 87–116. [Google Scholar] [CrossRef]
- Kooke, R.; Johannes, F.; Wardenaar, R.; Becker, F.; Etcheverry, M.; Colot, V.; Vreugdenhil, D.; Keurentjes, J.J.B. Epigenetic Basis of Morphological Variation and Phenotypic Plasticity in Arabidopsis Thaliana. Plant Cell 2015, 27, 337–348. [Google Scholar] [CrossRef]
- Reinders, J.; Wulff, B.B.H.; Mirouze, M.; Marí-Ordóñez, A.; Dapp, M.; Rozhon, W.; Bucher, E.; Theiler, G.; Paszkowski, J. Compromised Stability of DNA Methylation and Transposon Immobilization in Mosaic Arabidopsis Epigenomes. Genes Dev. 2009, 23, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Johannes, F.; Porcher, E.; Teixeira, F.K.; Saliba-Colombani, V.; Simon, M.; Agier, N.; Bulski, A.; Albuisson, J.; Heredia, F.; Audigier, P.; et al. Assessing the Impact of Transgenerational Epigenetic Variation on Complex Traits. PLoS Genet. 2009, 5, e1000530. [Google Scholar] [CrossRef]
- Tsukahara, S.; Kobayashi, A.; Kawabe, A.; Mathieu, O.; Miura, A.; Kakutani, T. Bursts of Retrotransposition Reproduced in Arabidopsis. Nature 2009, 461, 423–426. [Google Scholar] [CrossRef]
- Mirouze, M.; Reinders, J.; Bucher, E.; Nishimura, T.; Schneeberger, K.; Ossowski, S.; Cao, J.; Weigel, D.; Paszkowski, J.; Mathieu, O. Selective Epigenetic Control of Retrotransposition in Arabidopsis. Nature 2009, 461, 427–430. [Google Scholar] [CrossRef]
- Robertson, D.S. Characterization of a Mutator System in Maize. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1978, 51, 21–28. [Google Scholar] [CrossRef]
- McClintock, B. The Significance of Responses of the Genome to Challenge. Science 1984, 226, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Hake, S. The Power of Classic Maize Mutants: Driving Forward Our Fundamental Understanding of Plants. Plant Cell 2021, 34, 2505–2517. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. Mutator Transposons. Trends Plant Sci. 2002, 7, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kato, K.; Hosaka, A.; Fu, Y.; Toyoda, A.; Fujiyama, A.; Tarutani, Y.; Kakutani, T. Arms Race between Anti-silencing and RdDM in Noncoding Regions of Transposable Elements. EMBO Rep. 2023, 24, e56678. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.F.; Thompson, D.V.; Talbot, D.R.; Swanson, J.; Bennetzen, J.L. Nucleotide Sequence of the Maize Transposable Element Mul. Nucleic Acids Res. 1984, 12, 6924. [Google Scholar] [CrossRef]
- Alleman, M.; Freeling, M. The Mu Transposable Elements Of Maize: Evidence For Transposition And Copy Number Regulation During Development. Genetics 1986, 112, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Freeling, M.; Lisch, D. Mu Killer Causes the Heritable Inactivation of the Mutator Family of Transposable Elements in Zea Mays. Genetics 2003, 165, 781–797. [Google Scholar] [CrossRef]
- Burgess, D.; Li, H.; Zhao, M.; Kim, S.Y.; Lisch, D. Silencing of Mutator Elements in Maize Involves Distinct Populations of Small RNAs and Distinct Patterns of DNA Methylation. Genetics 2020, 215, 379–391. [Google Scholar] [CrossRef]
- Kolkman, J.M.; Conrad, L.J.; Farmer, P.R.; Hardeman, K.; Ahern, K.R.; Lewis, P.E.; Sawers, R.J.H.; Lebejko, S.; Chomet, P.; Brutnell, T.P. Distribution of Activator (Ac) Throughout the Maize Genome for Use in Regional Mutagenesis. Genetics 2005, 169, 981–995. [Google Scholar] [CrossRef]
- Bai, L.; Singh, M.; Pitt, L.; Sweeney, M.; Brutnell, T.P. Generating Novel Allelic Variation Through Activator Insertional Mutagenesis in Maize. Genetics 2007, 175, 981–992. [Google Scholar] [CrossRef]
- Bensen, R.J.; Johal, G.S.; Crane, V.C.; Tossberg, J.T.; Schnable, P.S.; Meeley, R.B.; Briggs, S.P. Cloning and Characterization of the Maize An1 Gene. Plant Cell 1995, 7, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Naito, K.; Cho, E.; Yang, G.; Campbell, M.A.; Yano, K.; Okumoto, Y.; Tanisaka, T.; Wessler, S.R. Dramatic Amplification of a Rice Transposable Element during Recent Domestication. Proc. Natl. Acad. Sci. USA 2006, 103, 17620–17625. [Google Scholar] [CrossRef]
- Simmons, C.R.; Lafitte, H.R.; Reimann, K.S.; Brugière, N.; Roesler, K.; Albertsen, M.C.; Greene, T.W.; Habben, J.E. Successes and Insights of an Industry Biotech Program to Enhance Maize Agronomic Traits. Plant Sci. 2021, 307, 110899. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Ito, M.; Sugita, T.; Tsukiyama, T.; Saito, H.; Naito, K.; Teraishi, M.; Tanisaka, T.; Okumoto, Y. Utilization of Transposable Element MPing as a Novel Genetic Tool for Modification of the Stress Response in Rice. Mol. Breed. 2013, 32, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Kirov, I.; Merkulov, P.; Polkhovskaya, E.; Konstantinov, Z.; Kazancev, M.; Saenko, K.; Polkhovskiy, A.; Dudnikov, M.; Garibyan, T.; Demurin, Y.; et al. Epigenetic Stress and Long-Read CDNA Sequencing of Sunflower (Helianthus annuus L.) Revealed the Origin of the Plant Retrotranscriptome. Plants 2022, 11, 3579. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Huang, H.; Bradai, M.; Zhao, C.; You, Y.; Ma, J.; Zhao, L.; Lozano-Durán, R.; Zhu, J.-K. DNA Methylation-Free Arabidopsis Reveals Crucial Roles of DNA Methylation in Regulating Gene Expression and Development. Nat. Commun. 2022, 13, 1335. [Google Scholar] [CrossRef]
- Ito, H. Environmental Stress and Transposons in Plants. Genes Genet. Syst. 2022, 97, 169–175. [Google Scholar] [CrossRef]
- Lanciano, S.; Mirouze, M. Transposable Elements: All Mobile, All Different, Some Stress Responsive, Some Adaptive? Curr. Opin. Genet. Dev. 2018, 49, 106–114. [Google Scholar] [CrossRef]
- Sun, L.; Jing, Y.; Liu, X.; Li, Q.; Xue, Z.; Cheng, Z.; Wang, D.; He, H.; Qian, W. Heat Stress-Induced Transposon Activation Correlates with 3D Chromatin Organization Rearrangement in Arabidopsis. Nat. Commun. 2020, 11, 1886. [Google Scholar] [CrossRef]
- Kwolek, K.; Kędzierska, P.; Hankiewicz, M.; Mirouze, M.; Panaud, O.; Grzebelus, D.; Macko-Podgórni, A. Diverse and Mobile: EccDNA-based Identification of Carrot Low-copy-number LTR Retrotransposons Active in Callus Cultures. Plant J. 2022, 110, 1811–1828. [Google Scholar] [CrossRef]
- Hirochika, H. Activation of Tobacco Retrotransposons during Tissue Culture. EMBO J. 1993, 12, 2521–2528. [Google Scholar] [CrossRef] [PubMed]
- Hirochika, H.; Sugimoto, K.; Otsuki, Y.; Tsugawa, H.; Kanda, M. Retrotransposons of Rice Involved in Mutations Induced by Tissue Culture. Proc. Natl. Acad. Sci. USA 1996, 93, 7783–7788. [Google Scholar] [CrossRef] [PubMed]
- Bayram, E.; Yilmaz, S.; Hamat-Mecbur, H.; Kartal-Alacam, G.; Gozukirmizi, N. “Nikita” Retrotransposon Movements in Callus Cultures of Barley (Hordeum vulgare L.). Plant Omics 2012, 5, 211–215. [Google Scholar]
- Masuta, Y.; Nozawa, K.; Takagi, H.; Yaegashi, H.; Tanaka, K.; Ito, T.; Saito, H.; Kobayashi, H.; Matsunaga, W.; Masuda, S.; et al. Inducible Transposition of a Heat-Activated Retrotransposon in Tissue Culture. Plant Cell Physiol. 2016, 58, pcw202. [Google Scholar] [CrossRef]
- Miyao, A.; Tanaka, K.; Murata, K.; Sawaki, H.; Takeda, S.; Abe, K.; Shinozuka, Y.; Onosato, K.; Hirochika, H. Target Site Specificity of the Tos17 Retrotransposon Shows a Preference for Insertion within Genes and against Insertion in Retrotransposon-Rich Regions of the Genome. Plant Cell 2003, 15, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Zervudacki, J.; Yu, A.; Amesefe, D.; Wang, J.; Drouaud, J.; Navarro, L.; Deleris, A. Transcriptional Control and Exploitation of an Immune-responsive Family of Plant Retrotransposons. EMBO J. 2018, 37, e98482. [Google Scholar] [CrossRef]
- Grandbastien, M.-A.; Lucas, H.; Morel, J.-B.; Mhiri, C.; Vernhettes, S.; Casacuberta, J.M. The Expression of the Tobacco Tnt1 Retrotransposon is Linked to Plant Defense Responses. In Contemporary Issues in Genetics and Evolution; Springer: Dordrecht, The Netherlands, 1997; pp. 241–252. [Google Scholar] [CrossRef]
- Merkulov, P.; Egorova, E.; Kirov, I. Composition and Structure of Arabidopsis Thaliana Extrachromosomal Circular DNAs Revealed by Nanopore Sequencing. Plants 2023, 12, 2178. [Google Scholar] [CrossRef]
- Mehta, D.; Cornet, L.; Hirsch-Hoffmann, M.; Zaidi, S.S.-A.; Vanderschuren, H. Full-Length Sequencing of Circular DNA Viruses and Extrachromosomal Circular DNA Using CIDER-Seq. Nat. Protoc. 2020, 15, 1673–1689. [Google Scholar] [CrossRef]
- Wang, K.; Tian, H.; Wang, L.; Wang, L.; Tan, Y.; Zhang, Z.; Sun, K.; Yin, M.; Wei, Q.; Guo, B.; et al. Deciphering Extrachromosomal Circular DNA in Arabidopsis. Comput. Struct. Biotechnol. J. 2021, 19, 1176–1183. [Google Scholar] [CrossRef]
- Lanciano, S.; Carpentier, M.-C.; Llauro, C.; Jobet, E.; Robakowska-Hyzorek, D.; Lasserre, E.; Ghesquière, A.; Panaud, O.; Mirouze, M. Sequencing the Extrachromosomal Circular Mobilome Reveals Retrotransposon Activity in Plants. PLoS Genet. 2017, 13, e1006630. [Google Scholar] [CrossRef]
- Zhang, P.; Mbodj, A.; Soundiramourtty, A.; Llauro, C.; Ghesquière, A.; Ingouff, M.; Slotkin, R.K.; Pontvianne, F.; Catoni, M.; Mirouze, M. Extrachromosomal Circular DNA and Structural Variants Highlight Genome Instability in Arabidopsis Epigenetic Mutants. Nat. Commun. 2023, 14, 5236. [Google Scholar] [CrossRef]
- Lee, S.C.; Ernst, E.; Berube, B.; Borges, F.; Parent, J.-S.; Ledon, P.; Schorn, A.; Martienssen, R.A. Arabidopsis Retrotransposon Virus-like Particles and Their Regulation by Epigenetically Activated Small RNA. Genome Res. 2020, 30, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.; Catoni, M.; Iwasaki, M.; Paszkowski, J. Sequence-Independent Identification of Active LTR Retrotransposons in Arabidopsis. Mol. Plant 2018, 11, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Benoit, M.; Catoni, M.; Drost, H.-G.; Brestovitsky, A.; Oosterbeek, M.; Paszkowski, J. Sensitive Detection of Pre-Integration Intermediates of Long Terminal Repeat Retrotransposons in Crop Plants. Nat. Plants 2019, 5, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, V.; Fan, W.; Chu, J.; Cho, J. Recent Advancement of NGS Technologies to Detect Active Transposable Elements in Plants. Genes Genom. 2021, 43, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Bajus, M.; Macko-Podgórni, A.; Grzebelus, D.; Baránek, M. A Review of Strategies Used to Identify Transposition Events in Plant Genomes. Front. Plant Sci. 2022, 13, 1080993. [Google Scholar] [CrossRef] [PubMed]
- Thieme, M.; Minadakis, N.; Himber, C.; Keller, B.; Xu, W.; Rutowicz, K.; Matteoli, C.; Böhrer, M.; Rymen, B.; Laudencia-Chingcuanco, D.; et al. Transposition of HOPPLA in SiRNA-Deficient Plants Suggests a Limited Effect of the Environment on Retrotransposon Mobility in Brachypodium Distachyon. bioRxiv 2023. [Google Scholar] [CrossRef]
- Erdmann, R.M.; Picard, C.L. RNA-Directed DNA Methylation. PLoS Genet. 2020, 16, e1009034. [Google Scholar] [CrossRef]
- Long, J.; Walker, J.; She, W.; Aldridge, B.; Gao, H.; Deans, S.; Vickers, M.; Feng, X. Nurse Cell–Derived Small RNAs Define Paternal Epigenetic Inheritance in Arabidopsis. Science 2021, 373, eabh0556. [Google Scholar] [CrossRef]
- Carvalheira, G.M.G. Plant Polytene Chromosomes. Genet. Mol. Biol. 2000, 23, 1043–1050. [Google Scholar] [CrossRef]
- Madlung, A.; Tyagi, A.P.; Watson, B.; Jiang, H.; Kagochi, T.; Doerge, R.W.; Martienssen, R.; Comai, L. Genomic Changes in Synthetic Arabidopsis Polyploids. Plant J. 2005, 41, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Kasprzewska, A.; Tennessen, K.; Fernandes, J.; Nan, G.-L.; Walbot, V.; Sundaresan, V.; Vance, V.; Bowman, L.H. Clusters and Superclusters of Phased Small RNAs in the Developing Inflorescence of Rice. Genome Res. 2009, 19, 1429–1440. [Google Scholar] [CrossRef]
- Zhai, J.; Zhang, H.; Arikit, S.; Huang, K.; Nan, G.-L.; Walbot, V.; Meyers, B.C. Spatiotemporally Dynamic, Cell-Type–Dependent Premeiotic and Meiotic PhasiRNAs in Maize Anthers. Proc. Natl. Acad. Sci. USA 2015, 112, 3146–3151. [Google Scholar] [CrossRef]
- Araki, S.; Le, N.T.; Koizumi, K.; Villar-Briones, A.; Nonomura, K.-I.; Endo, M.; Inoue, H.; Saze, H.; Komiya, R. MiR2118-Dependent U-Rich PhasiRNA Production in Rice Anther Wall Development. Nat. Commun. 2020, 11, 3115. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.T.; Mosher, R.A. Small RNA-Mediated DNA Methylation during Plant Reproduction. Plant Cell 2023, 35, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, X.; Wang, C.; Li, Q.; Meyers, B.C.; Springer, N.M.; Walbot, V. CHH DNA Methylation Increases at 24-PHAS Loci Depend on 24-nt Phased Small Interfering RNAs in Maize Meiotic Anthers. New Phytol. 2021, 229, 2984–2997. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Teng, C.; Xia, R.; Meyers, B.C. PhasiRNAs in Plants: Their Biogenesis, Genic Sources, and Roles in Stress Responses, Development, and Reproduction. Plant Cell 2020, 32, 3059–3080. [Google Scholar] [CrossRef]
- Teng, C.; Zhang, H.; Hammond, R.; Huang, K.; Meyers, B.C.; Walbot, V. Dicer-like 5 Deficiency Confers Temperature-Sensitive Male Sterility in Maize. bioRxiv 2020, 498410. [Google Scholar] [CrossRef]
- Patel, P.; Mathioni, S.; Kakrana, A.; Shatkay, H.; Meyers, B.C. Reproductive PhasiRNAs in Grasses Are Compositionally Distinct from Other Classes of Small RNAs. bioRxiv 2018, 242727. [Google Scholar] [CrossRef]
- Pachamuthu, K.; Borges, F. Epigenetic Control of Transposons during Plant Reproduction: From Meiosis to Hybrid Seeds. Curr. Opin. Plant Biol. 2023, 75, 102419. [Google Scholar] [CrossRef]
- Wang, Z.; Butel, N.; Santos-González, J.; Borges, F.; Yi, J.; Martienssen, R.A.; Martinez, G.; Köhler, C. Polymerase IV Plays a Crucial Role in Pollen Development in Capsella. Plant Cell 2020, 32, 950–966. [Google Scholar] [CrossRef]
- Slotkin, R.K.; Vaughn, M.; Borges, F.; Tanurdžić, M.; Becker, J.D.; Feijó, J.A.; Martienssen, R.A. Epigenetic Reprogramming and Small RNA Silencing of Transposable Elements in Pollen. Cell 2009, 136, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; Panda, K.; Köhler, C.; Slotkin, R.K. Silencing in Sperm Cells Is Directed by RNA Movement from the Surrounding Nurse Cell. Nat. Plants 2016, 2, 16030. [Google Scholar] [CrossRef] [PubMed]
- Creasey, K.M.; Zhai, J.; Borges, F.; Ex, F.V.; Regulski, M.; Meyers, B.C.; Martienssen, R.A. MiRNAs Trigger Widespread Epigenetically Activated SiRNAs from Transposons in Arabidopsis. Nature 2014, 508, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Parent, J.-S.; van Ex, F.; Wolff, P.; Martínez, G.; Köhler, C.; Martienssen, R.A. Transposon-Derived Small RNAs Triggered by MiR845 Mediate Genome Dosage Response in Arabidopsis. Nat. Genet. 2018, 50, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.A.; Bonnet, D.M.V.; Jullien, P.E. Non-Cell-Autonomous Small RNA Silencing in Arabidopsis Female Gametes. Curr. Biol. 2023, 33, 183–188.e3. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.W.; Burgess, D.; Kendall, T.; Baten, A.; Pokhrel, S.; King, G.J.; Meyers, B.C.; Freeling, M.; Mosher, R.A. Abundant Expression of Maternal SiRNAs Is a Conserved Feature of Seed Development. Proc. Natl. Acad. Sci. USA 2020, 117, 15305–15315. [Google Scholar] [CrossRef]
- Kitagawa, M.; Jackson, D. Plasmodesmata-Mediated Cell-to-Cell Communication in the Shoot Apical Meristem: How Stem Cells Talk. Plants 2017, 6, 12. [Google Scholar] [CrossRef]
- Gutzat, R.; Rembart, K.; Nussbaumer, T.; Hofmann, F.; Pisupati, R.; Bradamante, G.; Daubel, N.; Gaidora, A.; Lettner, N.; Donà, M.; et al. Arabidopsis Shoot Stem Cells Display Dynamic Transcription and DNA Methylation Patterns. EMBO J. 2020, 39, e103667. [Google Scholar] [CrossRef]
- Bradamante, G.; Scheid, O.M.; Incarbone, M. Under Siege: Virus Control in Plant Meristems and Progeny. Plant Cell 2021, 33, koab140. [Google Scholar] [CrossRef]
- Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. An SiRNA Pathway Prevents Transgenerational Retrotransposition in Plants Subjected to Stress. Nature 2011, 472, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, Z.; Zhang, Z.; Cheng, L.; Zhang, X.; Li, T. Small RNA-Sequencing Links Physiological Changes and RdDM Process to Vegetative-to-Floral Transition in Apple. Front. Plant Sci. 2017, 8, 873. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, W.; Ohama, N.; Tanabe, N.; Masuta, Y.; Masuda, S.; Mitani, N.; Yamaguchi-Shinozaki, K.; Ma, J.F.; Kato, A.; Ito, H. A Small RNA Mediated Regulation of a Stress-Activated Retrotransposon and the Tissue Specific Transposition during the Reproductive Period in Arabidopsis. Front. Plant Sci. 2015, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Scheid, O.M.; Gutzat, R. Heat Stress Response and Transposon Control in Plant Shoot Stem Cells. bioRxiv 2023. [Google Scholar] [CrossRef]
- Baubec, T.; Finke, A.; Scheid, O.M.; Pecinka, A. Meristem-specific Expression of Epigenetic Regulators Safeguards Transposon Silencing in Arabidopsis. EMBO Rep. 2014, 15, 446–452. [Google Scholar] [CrossRef]
- Yu, D.; Fan, B.; MacFarlane, S.A.; Chen, Z. Analysis of the Involvement of an Inducible Arabidopsis RNA-Dependent RNA Polymerase in Antiviral Defense. Mol. Plant-Microbe Interact. MPMI 2003, 16, 206–216. [Google Scholar] [CrossRef]
- Bradamante, G.; Nguyen, V.H.; Incarbone, M.; Meir, Z.; Bente, H.; Donà, M.; Lettner, N.; Scheid, O.M.; Gutzat, R. Two AGO Proteins with Transposon-Derived SRNA Cargo Mark the Germline in Arabidopsis. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bredthauer, C.; Fischer, A.; Ahari, A.J.; Cao, X.; Weber, J.; Rad, L.; Rad, R.; Wachutka, L.; Gagneur, J. Transmicron: Accurate Prediction of Insertion Probabilities Improves Detection of Cancer Driver Genes from Transposon Mutagenesis Screens. Nucleic Acids Res. 2023, 51, e21. [Google Scholar] [CrossRef]
- Merenciano, M.; Ullastres, A.; de Cara, M.A.R.; Barrón, M.G.; González, J. Multiple Independent Retroelement Insertions in the Promoter of a Stress Response Gene Have Variable Molecular and Functional Effects in Drosophila. PLoS Genet. 2016, 12, e1006249. [Google Scholar] [CrossRef]
- Siudeja, K.; Beek, M.; Riddiford, N.; Boumard, B.; Wurmser, A.; Stefanutti, M.; Lameiras, S.; Bardin, A.J. Unraveling the Features of Somatic Transposition in the Drosophila Intestine. EMBO J. 2021, 40, e106388. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, M.; McCarty, D.R.; Lisch, D. Transposable Elements Employ Distinct Integration Strategies with Respect to Transcriptional Landscapes in Eukaryotic Genomes. Nucleic Acids Res. 2020, 48, gkaa370. [Google Scholar] [CrossRef]
- Sultana, T.; Zamborlini, A.; Cristofari, G.; Lesage, P. Integration Site Selection by Retroviruses and Transposable Elements in Eukaryotes. Nat. Rev. Genet. 2017, 18, 292–308. [Google Scholar] [CrossRef] [PubMed]
- Sandmeyer, S.B.; Hansen, L.J.; Chalker, D.L. Integration Specificity of Retrotransposons and Retroviruses. Annu. Rev. Genet. 1990, 24, 491–518. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hou, Y.; Ebina, H.; Levin, H.L.; Voytas, D.F. Chromodomains Direct Integration of Retrotransposons to Heterochromatin. Genome Res. 2008, 18, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Arbuckle, J.; Wessler, S.R. Recent, Extensive, and Preferential Insertion of Members of the Miniature Inverted-Repeat Transposable Element Family Heartbreaker into Genic Regions of Maize. Proc. Natl. Acad. Sci. USA 2000, 97, 1160–1165. [Google Scholar] [CrossRef]
- Zhou, J.; Lei, X.; Shafiq, S.; Zhang, W.; Li, Q.; Li, K.; Zhu, J.; Dong, Z.; He, X.; Sun, Q. DDM1-Mediated R-Loop Resolution and H2A.Z Exclusion Facilitates Heterochromatin Formation in Arabidopsis. Sci. Adv. 2023, 9, eadg2699. [Google Scholar] [CrossRef]
- Yelagandula, R.; Stroud, H.; Holec, S.; Zhou, K.; Feng, S.; Zhong, X.; Muthurajan, U.M.; Nie, X.; Kawashima, T.; Groth, M.; et al. The Histone Variant H2A.W Defines Heterochromatin and Promotes Chromatin Condensation in Arabidopsis. Cell 2014, 158, 98–109. [Google Scholar] [CrossRef]
- Naish, M.; Alonge, M.; Wlodzimierz, P.; Tock, A.J.; Abramson, B.W.; Schmücker, A.; Mandáková, T.; Jamge, B.; Lambing, C.; Kuo, P.; et al. The Genetic and Epigenetic Landscape of the Arabidopsis Centromeres. Science 2021, 374, eabi7489. [Google Scholar] [CrossRef]
- Bourguet, P.; Picard, C.L.; Yelagandula, R.; Pélissier, T.; Lorković, Z.J.; Feng, S.; Pouch-Pélissier, M.-N.; Schmucker, A.; Jacobsen, S.E.; Berger, F.; et al. The Histone Variant H2A.W and Linker Histone H1 Co-Regulate Heterochromatin Accessibility and DNA Methylation. Nat. Commun. 2021, 12, 2683. [Google Scholar] [CrossRef]
- Alonge, M.; Shumate, A.; Puiu, D.; Zimin, A.V.; Salzberg, S.L. Chromosome-Scale Assembly of the Bread Wheat Genome Reveals Thousands of Additional Gene Copies. Genetics 2020, 216, 599–608. [Google Scholar] [CrossRef]
- Aury, J.-M.; Engelen, S.; Istace, B.; Monat, C.; Lasserre-Zuber, P.; Belser, C.; Cruaud, C.; Rimbert, H.; Leroy, P.; Arribat, S.; et al. Long-Read and Chromosome-Scale Assembly of the Hexaploid Wheat Genome Achieves High Resolution for Research and Breeding. GigaScience 2022, 11, giac034. [Google Scholar] [CrossRef] [PubMed]
- Rabanus-Wallace, M.T.; Hackauf, B.; Mascher, M.; Lux, T.; Wicker, T.; Gundlach, H.; Baez, M.; Houben, A.; Mayer, K.F.X.; Guo, L.; et al. Chromosome-Scale Genome Assembly Provides Insights into Rye Biology, Evolution and Agronomic Potential. Nat. Genet. 2021, 53, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Liao, N.; Hu, Z.; Miao, J.; Hu, X.; Lyu, X.; Fang, H.; Zhou, Y.-M.; Mahmoud, A.; Deng, G.; Meng, Y.-Q.; et al. Chromosome-Level Genome Assembly of Bunching Onion Illuminates Genome Evolution and Flavor Formation in Allium Crops. Nat. Commun. 2022, 13, 6690. [Google Scholar] [CrossRef] [PubMed]
- Coletta, R.D.; Qiu, Y.; Ou, S.; Hufford, M.B.; Hirsch, C.N. How the Pan-Genome Is Changing Crop Genomics and Improvement. Genome Biol. 2021, 22, 3. [Google Scholar] [CrossRef]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.L.; Papadopoulou, K. Genetically Modified Organism-Free RNA Interference: Exogenous Application of RNA Molecules in Plants. Plant Physiol. 2019, 182, 38–50. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and Advantages of Virus-induced Gene Silencing for Gene Function Studies in Plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Kiani, S.; Chavez, A.; Tuttle, M.; Hall, R.N.; Chari, R.; Ter-Ovanesyan, D.; Qian, J.; Pruitt, B.W.; Beal, J.; Vora, S.; et al. Cas9 GRNA Engineering for Genome Editing, Activation and Repression. Nat. Methods 2015, 12, 1051–1054. [Google Scholar] [CrossRef]
- Tsanova, T.; Stefanova, L.; Topalova, L.; Atanasov, A.; Pantchev, I. DNA-Free Gene Editing in Plants: A Brief Overview. Biotechnol. Biotechnol. Equip. 2021, 35, 131–138. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Wei, H.-M.; Lu, H.-Y.; Lee, Y.-I.; Fu, S.-F. Developmental- and Tissue-Specific Expression of NbCMT3-2 Encoding a Chromomethylase in Nicotiana Benthamiana. Plant Cell Physiol. 2015, 56, 1124–1143. [Google Scholar] [CrossRef]
- Hou, P.; Lee, Y.; Hsu, K.; Lin, Y.; Wu, W.; Lin, J.; Nam, T.N.; Fu, S. Functional Characterization of Nicotiana Benthamiana Chromomethylase 3 in Developmental Programs by Virus-induced Gene Silencing. Physiol. Plant. 2014, 150, 119–132. [Google Scholar] [CrossRef]
- Raz, A.; Dahan-Meir, T.; Melamed-Bessudo, C.; Leshkowitz, D.; Levy, A.A. Redistribution of Meiotic Crossovers Along Wheat Chromosomes by Virus-Induced Gene Silencing. Front. Plant Sci. 2021, 11, 635139. [Google Scholar] [CrossRef] [PubMed]
- Moissiard, G.; Voinnet, O. Viral Suppression of RNA Silencing in Plants. Mol. Plant Pathol. 2004, 5, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Pumplin, N.; Voinnet, O. RNA Silencing Suppression by Plant Pathogens: Defence, Counter-Defence and Counter-Counter-Defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Kontra, L.; Csorba, T.; Tavazza, M.; Lucioli, A.; Tavazza, R.; Moxon, S.; Tisza, V.; Medzihradszky, A.; Turina, M.; Burgyán, J. Distinct Effects of P19 RNA Silencing Suppressor on Small RNA Mediated Pathways in Plants. PLoS Pathog. 2016, 12, e1005935. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, Y.-R.; Pei, Y.; Lin, S.-S.; Tuschl, T.; Patel, D.J.; Chua, N.-H. Cucumber Mosaic Virus-Encoded 2b Suppressor Inhibits Arabidopsis Argonaute1 Cleavage Activity to Counter Plant Defense. Genes Dev. 2006, 20, 3255–3268. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Martínez, L.; Rakitina, D.V.; Lewsey, M.G.; Atencio, F.A.; Llave, C.; Kalinina, N.O.; Carr, J.P.; Palukaitis, P.; Canto, T. Cucumber Mosaic Virus 2b Protein Subcellular Targets and Interactions: Their Significance to RNA Silencing Suppressor Activity. Mol. Plant-Microbe Interact. 2010, 23, 294–303. [Google Scholar] [CrossRef]
- Yu, A.; Lepère, G.; Jay, F.; Wang, J.; Bapaume, L.; Wang, Y.; Abraham, A.-L.; Penterman, J.; Fischer, R.L.; Voinnet, O.; et al. Dynamics and Biological Relevance of DNA Demethylation in Arabidopsis Antibacterial Defense. Proc. Natl. Acad. Sci. USA 2013, 110, 2389–2394. [Google Scholar] [CrossRef]
- Hamera, S.; Yan, Y.; Song, X.; Chaudhary, S.U.; Murtaza, I.; Su, L.; Tariq, M.; Chen, X.; Fang, R. Expression of Cucumber Mosaic Virus Suppressor 2b Alters FWA Methylation and Its SiRNA Accumulation in Arabidopsis Thaliana. Biol. Open 2016, 5, 1727–1734. [Google Scholar] [CrossRef]
- Shamandi, N.; Zytnicki, M.; Charbonnel, C.; Elvira-Matelot, E.; Bochnakian, A.; Comella, P.; Mallory, A.C.; Lepère, G.; Sáez-Vásquez, J.; Vaucheret, H. Plants Encode a General SiRNA Suppressor That Is Induced and Suppressed by Viruses. PLoS Biol. 2015, 13, e1002326. [Google Scholar] [CrossRef]
- Baduel, P.; Quadrana, L.; Colot, V. Plant Transposable Elements, Methods and Protocols. Methods Mol. Biol. 2021, 2250, 157–169. [Google Scholar] [CrossRef]
- McDonald, T.L.; Zhou, W.; Castro, C.P.; Mumm, C.; Switzenberg, J.A.; Mills, R.E.; Boyle, A.P. Cas9 Targeted Enrichment of Mobile Elements Using Nanopore Sequencing. Nat. Commun. 2021, 12, 3586. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.B.; Höijer, I.; Kvist, T.; Ameur, A.; Mikkelsen, M.J. Xdrop: Targeted Sequencing of Long DNA Molecules from Low Input Samples Using Droplet Sorting. Hum. Mutat. 2020, 41, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Gilpatrick, T.; Lee, I.; Graham, J.E.; Raimondeau, E.; Bowen, R.; Heron, A.; Downs, B.; Sukumar, S.; Sedlazeck, F.J.; Timp, W. Targeted Nanopore Sequencing with Cas9-Guided Adapter Ligation. Nat. Biotechnol. 2020, 38, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Vives-Vallés, J.A.; Collonnier, C. The Judgment of the CJEU of 25 July 2018 on Mutagenesis: Interpretation and Interim Legislative Proposal. Front. Plant Sci. 2020, 10, 1813. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirov, I. Toward Transgene-Free Transposon-Mediated Biological Mutagenesis for Plant Breeding. Int. J. Mol. Sci. 2023, 24, 17054. https://doi.org/10.3390/ijms242317054
Kirov I. Toward Transgene-Free Transposon-Mediated Biological Mutagenesis for Plant Breeding. International Journal of Molecular Sciences. 2023; 24(23):17054. https://doi.org/10.3390/ijms242317054
Chicago/Turabian StyleKirov, Ilya. 2023. "Toward Transgene-Free Transposon-Mediated Biological Mutagenesis for Plant Breeding" International Journal of Molecular Sciences 24, no. 23: 17054. https://doi.org/10.3390/ijms242317054
APA StyleKirov, I. (2023). Toward Transgene-Free Transposon-Mediated Biological Mutagenesis for Plant Breeding. International Journal of Molecular Sciences, 24(23), 17054. https://doi.org/10.3390/ijms242317054