Abstract
Substance addiction is a chronic and relapsing brain disorder characterized by compulsive seeking and continued substance use, despite adverse consequences. The high prevalence and social burden of addiction are indisputable; however, the available intervention is insufficient. The modulation of gene expression and aberrant adaptation of neural networks are attributed to the changes in brain functions under repeated exposure to addictive substances. Considerable studies have demonstrated that miRNAs are strong modulators of post-transcriptional gene expression in substance addiction. The emerging role of microRNA (miRNA) provides new insights into many biological and pathological processes in the central nervous system: their variable expression in different regions of the brain and tissues may play a key role in regulating the pathophysiological events of addiction. This work provides an overview of the current literature on miRNAs involved in addiction, evaluating their impaired expression and regulatory role in neuroadaptation and synaptic plasticity. Clinical implications of such modulatory capacities will be estimated. Specifically, it will evaluate the potential diagnostic role of miRNAs in the various stages of drug and substance addiction. Future perspectives about miRNAs as potential novel therapeutic targets for substance addiction and abuse will also be provided.
1. Introduction
Drug addiction is a significant public health issue worldwide. The term drug addiction comprehends a series of actions culminating in loss of control over drug intake, craving, and withdrawal. Another expression used in this regard is substance use disorder (SUD), and it refers to different types of behaviors varying from sporadic use of substances of abuse, abuse, dependence, or addiction.
The Diagnostic and Statistical Manual of Mental Disorders (DSM) provides a differential clinical diagnosis of these behaviors [1]. Repeated exposure to drugs of abuse frequently leads to a change in feeling, alarming behavior, temporary satiation, loss of control and negative consequences, and clear symptoms of addiction [2]. The evolution from casual drug use to compulsive drug seeking seems to be strictly related to the reward and motivation brain circuitry [3], an adaptive response to long-term drug use. One of the hallmarks of addiction is represented by the change from a positive reward, usually called a “high”, to negative stimuli during abstinence. Different brain regions are involved in these processes. In particular, acute rewarding effects are connected to an increase in dopamine release in the Nucleus Accumbens (NAc), while long-term plasticity in corticostriatal circuits are linked to compulsive drug seeking and susceptibility to relapse [4].
This behavior can be analyzed in animal models through Pavlovian-style conditioning such as operant conditioned lever-pressing self-administration (SA) or conditioned placement preference (CPP). Additionally, according to the Centers for Disease Control and Prevention, in the United States, deaths from drug overdoses topped 100,000 in 2022 [5].
There is, however, a lack of understanding of the underlying molecular mechanisms of addiction. Drug screening and confirmation of dependence or use disorder are mainly conducted by analyzing drugs and their metabolites in blood samples, which are not stable with levels constantly decreasing [6]. Furthermore, diagnoses of the severity of drug use disorders are mainly based on subjective reports and there are no objective biomarkers available. Therefore, the identification of stable and objective biomarkers for drug abuse is crucial.
Indeed, epigenetic, and social factors are involved in the development of addiction. On one hand, social and psychological factors predict a drug abuser profile; on the other hand, molecular processes are involved [7,8]. Although clinical studies have previously shown the impact of social factors on drug use [9], it is only recently that neuroscience has devoted careful attention to examining the influence of social factors on drug use and addiction in animal models [10]. Further specific attention should be given to the development of new disorders triggered by adaptive responses to drug use and those relapsed thereafter [11,12,13,14].
It is recognized that drugs of abuse operate by modulating gene expression and producing their rewarding effects of euphoria or pleasure through the interaction with the mesolimbic dopaminergic system, leading to persistent alterations in the reward-related and memory-related brain centers. These interactions translate into the alteration of proteins and gene expression by regulating transcription factors, chromatin structures, and small non-coding RNA (ncRNA) [2]. One of the most studied classes of ncRNA in drug addiction is that of microRNAs (miRNA) [15].
miRNAs are small non-coding RNA transcripts, comprising 19–25 ribonucleotides, that are not translated into proteins but are capable of regulating gene expression at a post-transcriptional level [16]. They have the ability to destabilize and cleave mRNA, therefore actively regulating their transcription. Moreover, the silencing effect of miRNAs on their target gene expression appears to be highly specific allowing a precise regulation of physiological responses [15]. Given their role in epigenetic regulation and synaptic plasticity, miRNAs have the potential to be used as markers of drug abuse both on brain tissue samples, but also on blood vessels and bodily fluids such as blood and saliva. By understanding which phases of addiction are regulated by specific miRNAs and which miRNAs are altered by different types of drugs, new tailored and ultra-selective therapies can be developed to aid in the battle against drug addiction. For this reason, a review of the currently available data on miRNAs involved in the main classes of drugs of abuse (alcohol, cocaine, methamphetamine, and opiates) has been carried out.
2. Materials and Methods
2.1. Eligibility Criteria
The present systematic review was carried out according to the Preferred Reporting Items for Systematic Review (PRISMA) standards [17]. We used an evidence-based model for framing a PICO.
2.2. Search Criteria and Critical Appraisal
A systematic literature search and a critical appraisal of the collected studies were conducted. An electronic search of PubMed, Science Direct Scopus, and Excerpta Medica Database (EMBASE) from the inception of these databases to September 2023 was performed. The search terms were “miRNA expression in addiction”; “miRNAs during alcohol addiction”; “miRNAs during cocaine addiction”; “miRNAs during methamphetamine addiction”; “miRNAs during opioids addiction” in the title, abstract, and keywords. The bibliographies of all identified documents were reviewed and cross-checked for the other relevant literature. Methodological assessment of each study was conducted according to PRISMA standards, including assessment of bias. Data collection included study selection and data extraction. Three researchers (C.O., J.L, N.I.) independently reviewed those documents whose title or abstract seemed relevant and selected those that analyzed miRNAs. The question of the suitability of miRNAs was resolved by consensus among the researchers. The unpublished or grey literature was not searched. The extraction of data was performed by three investigators (E.T., A.M., A.C.) and reviewed by two other researchers (V.F., P.F.). Only English-language papers or abstracts were included in the search.
2.3. Search Results and Included Studies
An appraisal based on titles and abstracts, as well as a hand search of reference lists, were carried out. The reference lists of all located articles were reviewed to detect the still unidentified literature. The resulting reference lists were then screened for title and abstract, leaving 216 articles, for further consideration, that were then screened based on their abstract to identify their relevance in respect to the following: Estimate diagnosis process; Clinical features analyzed; Circumstantial data evaluation; Study design.
The methodology of our search strategy is represented in Figure 1.
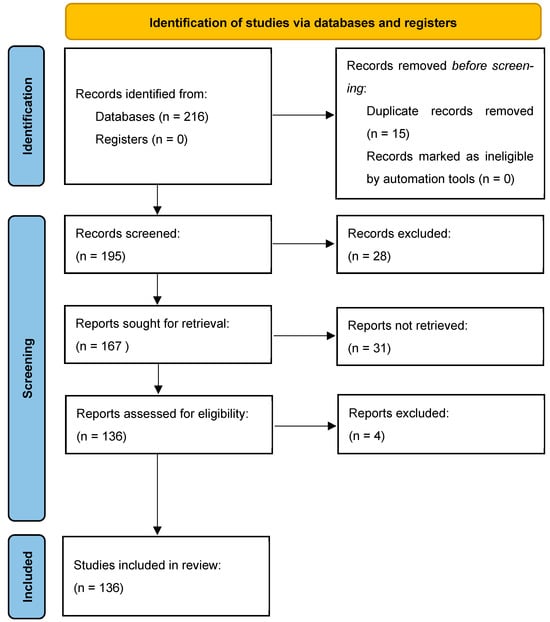
Figure 1.
The selection of appropriate scientific papers was performed; articles met the inclusion criteria and were included.
A further categorization of the articles included in this study was made based on the main parameter under study, as shown in the following table (Figure 1).
Non-English articles were excluded. The inclusion criteria were as follows: (1) original research articles; (2) case reports/series. Reviews and meta-analyses were excluded. Studies conducted on cultured cells, animals (mainly rats and mice), and humans that described the role of miRNAs in different stages of addiction on four types of substances of abuse (alcohol, cocaine, opioids, and methamphetamine) were selected. Among them, studies measuring miRNAs in CNS and some body fluids (mainly blood and saliva) from animals and humans were included, as also were studies with a control group. These publications were carefully assessed, considering the main objectives of this review. After this evaluation, 132 scientific papers remained.
2.4. Risk of Bias
This systematic review has several strengths that include the amount and breadth of the studies, which span the globe; the hand search and scan of reference lists for the identification of all relevant and significant studies; and a flowchart that describes in detail the study selection process. Included studies were evaluated according to the Quality Assessment of Diagnostic Accuracy Studies 2 (QUADAS-2) tool to assess the risk of bias. This review includes studies that were published in a time frame of 13 years (from 2010 to 2023); thus, despite our efforts to fairly evaluate the existing literature, study results should be interpreted taking into account that the accuracy of the clinical procedures, here reported, has changed over the years.
3. Results
3.1. miRNAs Expression in Alcohol Abuse
Alcohol is a neuro-depressant substance, able to determine the occurrence of short and long-term adverse effects. An unregulated consumption of alcohol may lead to a condition named “alcohol abuse”: it represents a spectrum of different unhealthy behaviors related to alcohol intake, ranging from binge drinking up to alcohol withdrawal, that comprises also entities such as alcohol use disorder (AUD) and alcohol dependence (AD). It has been widely demonstrated how alcohol abuse, in its various stages, regulates and is in turn regulated by miRNAs expression (see Table 1). Numerous are the miRNAs regulated by the consumption of alcohol, just as numerous are the targets on which they act. In a work by Lewohl and colleagues, it emerged that a typical pattern of interaction between miRNAs and their targets provides for an inverse correlation; it is usual to have an upregulation of miRNAs alongside a downregulation of their targets, or vice versa [18].

Table 1.
miRNAs expression in Alcohol abuse.
One of the most targeted molecules by miRNAs is the endogenous protective factor BDNF, which is usually found in the human brain. It works as a regulator of the transcriptional and translational mechanism when it is activated through the TrkB receptor. The BDNF/TrkB pathway has a key role in the development of the central nervous system (CNS) as it is involved in the formation of synapses, differentiation of neural cells, and synaptic plasticity.
Ehinger et al. [19] found out that moderate use of alcohol determines an increase of BDNF levels in the dorsolateral striatum (DLS), thus activating the TrkB/ERK signaling and inducing the dopamine D3 receptor, so as to maintain a moderate intake of the substance. Moreover, they also discovered that heavy use of alcohol is instead associated with a reduction of BDNF levels in the prefrontal cortex (PFC) and in serum 8 weeks after the intake of the substance, suggesting that after a past heavy alcohol intake, low BDNF protein levels may be measured.
In the study, it was found that BDNF is the target of numerous miRNAs, such as miR30a-5p, miR195, miR191, and miR206, whose levels are consistently increased after alcohol exposure even in serum. They hypothesized that such miRNAs are firstly produced in mPFC, then transported in the blood, thus acting as potential diagnostic tools in order to understand alcohol’s role in the brain. Even other studies reported similar results [2,4,43,44,45].
In addition, some of those studies further found out that miR-195 and miR-30a-5p are also upregulated after episodes of binge drinking and after one day of abstinence, whereas miR206 was upregulated even after three weeks of alcohol exposure, during the phase of withdrawal [20,22].
According to the studies conducted by Gowen [2], Heyer [4], Zhang [45], and Zhao [43], it seems that BDNF is also regulated by miR124a, which appears to have a key role in the mechanism of addiction to substances. In particular, miR124a levels were decreased in the DLS of rats after 15 days of ethanol intaking, and such featuring was associated with increased levels of BDNF [21]. On a practical level, the reduction of miR124a expression was related to a decrease of alcohol intake and, as a consequence, they suggest that it may be used to counteract vulnerability to alcohol dependence. Moreover, Mizuo et al. [26] found out that miR124 levels, along with miR132, remained upregulated 12 h after alcohol assumption, and also in the phase of abstinence, which means that it may play a significant role even in the relapse phase.
Another miRNA whose expression is modified subsequently to alcohol exposure is miR9. Several works highlighted how its levels tend to increase in the cells of the hypophysis, the supraoptic nucleus, and the striatum following acute intake of alcohol [4,25,43,45]. The target of miR9 is the BK channel, an ionic channel whose expression is usually reduced by alcohol.
By binding the BK channel, miR9 induces post-transcriptional modifications resulting in the downregulation of a specific subunit therefore making the BK channel less sensitive to alcohol. As a consequence, miR9 can determine a long-term adaptation that induces an increase in tolerance to ethanol [23].
Gu et al. [24] also identified HDA5 as a target of miR9. It is part of the histone deacetylases family and is negatively regulated by miR9. Its decreased expression in primary cortical neurons seemed to be responsible for behavioral responses to chronic drug exposure.
A further target of miR9 is the dopaminergic receptor DR2, where its lower levels have been associated with alcohol abuse. Such a result suggests that miR9 also affects the alcohol-related reward effect and can take part in the development of addiction 27 [25]. Even DR1 represents a target of miRNAs following alcohol exposure. In particular, this dopaminergic receptor is targeted by miR-382: miR-382 is downregulated after repeated alcohol administration whereas DR1 expression is increased. It was further demonstrated that inducing expression of miR-382 determines inhibition of DR1 and, consequently, reduction of alcohol consumption [27].
Moreover, overexpression of let-7d determines the downregulation of DR3 in the nucleus accumbens of rats, thus reducing alcohol consumption [21].
According to Most’s work, it seems that miRNAs affect and regulate other neurotransmission systems, like the glutamate one: for example, one of the targets of miR-411 is GluR2. miR-411 is reduced after chronic alcohol intake and knockdown of miR-411 reduces alcohol intake besides inducing expression of GluR2 [46].
Another circuit influenced by alcohol and miRNA expression is, clearly, the GABAergic one. Bekdash et al. [38] observed that during alcohol withdrawal, subunit Gabra4 of the GABAA receptor was down-expressed. They demonstrated that knockdown of Gabra4 was inducted by transfection of miR-186, miR-24, miR-375, and miR-155 mimics. Conversely, transfection of inhibitors of such miRNAs determined restoration of the expression of Gabra4, thus suggesting that Gabra4 represents an additional target.
The serotoninergic circuit is also influenced by miRNA expression. Another receptor targeted by miRNAs is the glucocorticoid one (GR). It was observed that long-term alcohol exposure determined a reduction in GRα’s expression in the presence of raised levels of miR-124. Such a phenomenon may be implicated in alcohol addiction [31].
Interestingly, some studies focused on the capacity of miRNAs regulated by alcohol exposure, in controlling apoptotic processes. Among these miRNAs, there are miR21 and miR335, which act as functional antagonists [35], and miR29b, which exerts a protective role against ethanol-induced neurotoxicity [36].
MiRNAs are also expressed in synapses and their regulation can be altered by chronic alcohol consumption. In their experiments, conducted on murine models, Most et al. demonstrated that alcohol actually alters miRNA expression in glutamate synapses. Some of those miRNAs, such as miR-203, miR-18 a, and miR-374, were also identified in human brains [28].
miRNAs altered by alcohol intake could also be found within some microvesicles released by glial cells. This is the case of let-7b, whose target is represented by TLR7 and activates the nuclear factor kβ (NF-kβ), thus determining neurodegeneration. Some studies evidenced that in alcoholic humans and rats, there were increased levels of TLR7. Such a result was the effect of the increased release of let-7b in microvesicles, that contributed to ethanol-induced neurotoxicity mediated through TLR7 [29]
An interesting work conducted by Asquith et al. demonstrated that miR-181 and miR-221 were overexpressed not only in the brain’s tissues but also in the peripheral blood mononuclear cells (PBMC) following chronic assumption of ethanol in those cells [37].
Choi et al. [39] evaluated the sex-dependent effects of alcohol on miRNA expression. They found that different patterns of miRNAs were up or downregulated depending on whether they were evaluated in males or females. Even neurogenesis was differently affected by alcohol depending on sex: this was evaluated by measuring DCX, demonstrating that males were more sensitive to alcohol than females.
Whereas most of the studies are based on experiments on rats, Lim et al.’s results are based on studies conducted on postmortem brain tissue collected from humans affected by AUD. They examined eight different brain regions, where the region with the greatest number of miRNAs associated with AUD was the PFC. The main targets of the different miRNAs were represented by CREB signaling, IL-8 signaling, and Axonal Guidance Signaling, thus suggesting the involvement of miRNAs in regulating anxiety-like and excessive alcohol-drinking behaviors [32]. Moreover, Liu and Zhang demonstrated, on postmortem brain tissue, that chronic alcohol consumption may determine hypermethylation of both miRNAs and mRNAs in the region of NAc [47].
Other studies conducted on humans by Manzardo et al. [42] examined miRNA expression in the medial frontal cortex of alcoholic subjects. They found that twelve miRNAs were upregulated (including miR-3065-5p, miR-299-5p, miR-767-5p, miR-375, miR-29b, miR-377, miR-399) and two were downregulated (miR-572 and miR-3162). These miRNAs targeted genes implicated in cellular adhesion (as THBS2), differentiation of tissues (as CHN2), migration (as NDE1), and maturation of neurons and oligodendrocytes.
Furthermore, it has been demonstrated that miRNAs could also cross the placenta and potentially exert some teratogenic effects. It was observed that the expression of some of these miRNAs is influenced by exposition to alcohol in a way that they could be possibly used as biomarkers for forecasting the effects on infants with prenatal alcohol exposure [30].
Similar results were found by Tseng et al. who focused attention on miR-140, a molecule contained in microvesicles released from fetal neural stem cells, whose levels were increased after 3 days of alcohol exposure [34]. Other miRNAs that have been observed in the serum of mothers with alcohol abuse were miR122, miR126, miR216b, miR221, miR3119, miR3942, miR4704, miR4743, miR514, and miR602.
Finally, an interesting work was presented by Rosato et al. [48] Through an experiment conducted on 110 participants, they demonstrated that miRNAs measured in saliva could be used as biomarkers in order to obtain a non-invasive prediction for alcohol dependence. In particular, they identified a cluster of five salivary miRNAs with an accuracy for prediction of over 70%.
3.2. miRNAs Expression in Cocaine Abuse
Cocaine is one of the most consumed stimulants in the world. It is a natural sympathomimetic tropane alkaloid that can be extracted from the leaves of Erythroxylon coca, by either chewing the leaves or brewing tea. In the mid-1800s, cocaine was considered safe and used in toothache drops, nausea pills, energy tonics, and also, in the original recipe of the ‘Coca-Cola’ beverage. Nowadays, cocaine is usually found in two forms: cocaine hydrochloride, commonly known as ‘coke’, ‘blow’, or ‘snow’, a fine crystalline white powder, soluble in water and mainly consumed intranasally (‘sniffing’ or ‘snorting’), orally, or intravenously; and as ‘crack’ cocaine, a free base resulting from the reaction of cocaine hydrochloride and ammonium or baking soda, and typically consumed via inhalation. Cocaine has singular pharmacodynamic properties that allow its use both as a sympathomimetic stimulant of the central nervous system and as a local anesthetic.
The sympathomimetic and psychoactive effects are linked to the blockade of presynaptic transporters responsible for the reuptake of serotonin, noradrenaline, and dopamine. The anesthetic properties are related, on the other hand, to its capacity to block voltage-gated sodium channels through their stabilization in an inactive state. When cocaine is consumed, dopaminergic activity along the mesocorticolimbic pathways is intensified. This substance induces euphoria, improves concentration and alertness, increases libido, promotes a general sensation of well-being, and reduces appetite and fatigue. Nonetheless, its use can also determine insomnia, irritability, anxiety, impulsive behavior, and dysphoria, particularly after continued drug use. Moreover, it is correlated to cardiovascular effects such as tachycardia, vasoconstriction, and hypertension. Cocaine has an elevated abuse potential, by increasing dopamine concentration in the nucleus accumbens, the dorsal caudate nucleus, and the striatum [49]. MiRNAs expression in cocaine abuse are shown in Table 2.
Table 2.
miRNAs Expression in Cocaine abuse.
Barreto-Valer et al. [50] evaluated the effects of cocaine on the expression of dopamine receptors and miR-133b. They demonstrated that cocaine alters dopaminergic receptor expression in various ways depending on both the receptor and the embryo’s developmental stage. Cocaine reduced the expression of miR-133 at 24 and 48 hpf and, conversely, increased pitx3 at 24 hpf. They then focused on the pitx3 target genes TH, DAT, and DRD2 expression, showing they were upregulated at 24 hpf. TH and DAT decreased at 48 hpf, whereas DRD2A increased and DRD2B was downregulated at both 24 and 48 hpf. Finally, cocaine decreased the expression of miR-133b in the central nervous system and at the periphery, particularly on somites. Therefore, the authors hypothesized that it could be involved in skeletal muscle development.
Bastle et al. [51] evaluated the direct binding of miR-495 to target genes showing that it significantly reduced the activity of BDNF, Camk2a, and Arc in vitro. NAc miR-495 was downregulated between 1 and 4 h post-injection in a region-specific manner. Moreover, BDNF and Camk2a were upregulated 2 h after cocaine injection in the NAc.
Cabana-Dominguez et al. [52] found that miR-9-5p, miR-101-3p, miR-124-3p, miR-124-5p, miR-137, miR-153-3p, and miR- 369-3p where down-egulated after acute exposure to cocaine. Moreover, they correlated these downregulated miRNAs to the upregulation of several networks associated with cocaine dependences comprehensive of PKC and JUN. Additionally, the study showed an interaction between miR-124-3p and TEAD1.
Another study, conducted by Chandrasekar et al. [53], observed significant downregulation of miR-124 in the caudate putamen region after chronic cocaine intake whilst let-7d was downregulated in the VTA, the CPU, HIP, PFC, and ROB regions. miR-181a was upregulated after chronic cocaine treatment in NAc, CPU, VTA, and piriform cortex. In a subsequent study, Chandrasekar et al. [54] evaluated the effect of miR-124 during the cocaine CPP acquisition period, suggesting its expression could be crucial for suppressing the reinstating properties of cocaine. Instead, once the CPP has been established, the overexpression of miR-124 did not affect the reinstatement of cocaine CPP. They found a correlation between miR-124, miR-181, and let-7d with the observed behavioral changes after cocaine CPP, showing that these miRNAs are modulated by cocaine CPP.
A study by Chen et al. [55] explored the changes in miRNA expression in the hippocampus of rats during the acquisition and extinction of cocaine-induced CPP. These miRNAs regulate the expression of target genes that act in many cellular processes most of which are involved in metabolic process and biological regulation. Some of these miRNAs were also implicated in brain disorders and drug abuse.
Interestingly, Chivero et al. [56] used EV loaded with miR-124 to restore miR-124 levels and thus function as a potential therapeutic approach for cocaine-mediated neuroinflammation. EV-loaded miR-124 inhibits cocaine-mediated microglial activation. In fact, cocaine determines the downregulation of miR-124 and, therefore, upregulates the expression of TLR4 and STAT3. On the other hand, Dash et al. [58] observed that miR-124 regulated PARP-1 by binding its 3′-UTR. Using a vector containing the 3′-UTR of PARP-1 in the presence or absence of miR-124, they found that knockdown of miR-124 resulted in a significant induction of PARP-1. The study suggested that PARP-1 3′-UTR activity is regulated by miR-124 expression in neuronal cells. Moreover, they found that miR-124 had a dominant regulatory effect compared to miR-125b. Conjointly downregulation of miR-124 is suggested in multiple neuroinflammatory diseases comprehensive of amyotrophic lateral sclerosis, PD, while its overexpression could mitigate the pathogenesis [75]. Guo et al. [66] observed that cocaine significantly decreased miR-124 expression in microglial cells by exposing BV-2 cells to cocaine for 24 h. Conversely, cocaine exposure increased levels of DNMT in rats’ primary microglia, and, in particular of DNMT1. Moreover, they discovered these alterations also in vivo. Then, they demonstrated that DNMT1 inhibitor 5-Azacytidine (5-AZA) reversed cocaine-mediated effects on miR-124 and DNMTs. These findings demonstrate a direct link between cocaine and the expression of miR-124 and DNMTs, therefore indicating that changes in the promoter of DNA methylation could be crucial in the cocaine-mediated downregulation of miR-124.
Dash et al. [57] evaluated the expression of miR-125b in a dopaminergic neuronal cell model and in the NAc of mice, finding it downregulated in both models. They also investigated PARP-1 expression finding that miR-125b negatively regulates PARP-1 acting on a post-transcriptional level. Moreover, it seems that miR-125b downregulation is dependent on DAT. Chronic cocaine exposure activated the miR-125b/PARP-1 axis, therefore, reducing its action on cellular damage repairs.
Doke et al. [60] evaluated miRNA variations in both HIV Tat 1 and cocaine-treated human primary astrocytes in order to understand if exposure to cocaine could dysregulate miRNA expression. They established that 8 miRNAs were significantly upregulated and 13 were downregulated when exposed to HIV Tat 1, while cocaine exposure altered the expression of 40 miRNAs, out of which 6 were downregulated while 8 were upregulated. Finally, they estimated how combined exposure altered miRNA expression, finding that they predominantly affected miR-155, miR-5580, miR-551a, miR-5090, miR-7155, miR-1255b2, miR-3133, and miR-6780a. miRNAs targeted gene sets associated with non-neoplastic disorder, unipolar disorder, and major depressive disorder and revealed they impacted most on pathways involved in metabolic pathways, fatty acid metabolism, and glycolysis/gluconeogenesis pathways. The correlation between cocaine exposure and HIV was also investigated by Napuri et al. [73] who evaluated the effect of cocaine on miR-155 and miR-20a. Both miRNAs are involved in the regulation of immune cell functions and are suppressed by HIV. They observed a significant downregulation of miR-155 and miR-20a after cocaine exposure. Then they focused on whether there was a synergic effect between cocaine and HIV on those miRNAs. Their findings suggested that cocaine exposure could enhance HIV-1 transcription and increase viral production in association with miR155 and miR-20a downregulation.
Sadakierska-Chudy et al. [78] assessed whether cocaine could induce long-lasting alterations of miRNA expression crucial for synaptic plasticity of the striatal region. They found miR-132 and miR-212 meaningfully upregulated after active cocaine intake. Their role is supposed to be crucial to synaptic plasticity and/or learning adaptation in the dorsal striatal region of the brain.
Schaefer et al. [79] found altered levels of 23 miRNAs. Moreover, they observed that Ago2 had a role in cocaine addiction due to ago2-dependent miRNAs and, specifically miRNAs with a potential role in neural plasticity and motivation to consume cocaine.
Tobon et al. [81] demonstrated that the D1 receptor is regulated in the caudate-putamen region of the brain but not in the nucleus accumbens and that D1 receptor levels increase within 30 min of cocaine administration in cocaine-sensitized mice. Within 5 min of cocaine administration, they found that levels of miR-142-3p and miR-382 were significantly decreased and that their downregulation in the caudate-putamen post-transcriptionally regulates D1 receptors.
Vannan et al. [80] studied the differential expression of miRNAs in the NAc shell and their correlation to cocaine-seeking behavior. They observed that 8 miRNAs were downregulated while 25 were upregulated in animals with low cocaine-seeking behavior. These miRNAs regulated the expression of several miRNAs targets: Zbtb20, Nfat5, Ago1, Qui, Nfib, Pde3a, Tcf4, Klf7, and Rorb.
Viola et al. [82] observed downregulation of miR-212 after 2 h cocaine-induced CPP test whilst Mecp2 is upregulated in the PFC of normally reared mice. This effect is blunted in maternally separated mice. Moreover, cocaine exposure determined a decreased expression of Bdnf.
Viola et al. [83] evaluated the association between cocaine abuse and miRNA expression showing upregulation of miR-124 and miR-181. These miRNAs regulate pathways involved in biological regulation, metabolic process, biogenesis, developmental process, response to stimulus, adhesion, signaling, and immune system.
3.3. miRNAs Expression in Opioids Abuse
Heroin is a morphine derivative, belonging to the opioids class, and it is one of the most powerfully addictive drugs of abuse in the world. Heroin addiction is a relapsing and chronic brain disorder that causes persistent alterations in synaptic plasticity [86].
Recent studies have indicated that miRNAs play a pivotal role in addiction by directly controlling synaptic remodeling, dendritic spine morphogenesis, drug dependence, and drug-rewarding properties in substance use disorders such as alcohol, cocaine, amphetamine and, of course, opioids [87] see Table 3.
Table 3.
miRNAs Expression in Opioids abuse.
Chronic morphine treatment has been shown to regulate the levels of miRNA-23b and let-7, which subsequently repress the µ-opioid receptor (MOR1) at a post-transcriptional level [98].
miRNAs offer a promising biomarker for integrating gene expression and regulation in the processes of substance use disorders.
Current research indicates that the NAc plays a central role in the regulation of behavioral functions associated with depression, anxiety, and addiction due to its involvement in the brain reward circuit. Approximately 95% of cells within the NAc are GABAergic neurons that possess dopamine D1-and D2-like receptors. NAc function is influenced by dopaminergic projections from the ventral tegmental area and numerous studies have indicated that the high level of MOR distribution in the NAc might be related to the behavioral properties of opioid addiction, including reward and withdrawal symptom [103].
The agonist of the µ-opioid receptor (OPRM1) elicits extracellular signal-regulated kinase (ERK) phosphorylation through different pathways: morphine utilizes the protein kinase C (PKC) pathway, whilst fentanyl operates in an arrestin2-dependent method.
Zheng and other authors, in their study, examined the effect of two pathways on miRNA expression. The first study [104] analyzed two pathways that led to distinct cellular localization of phosphorylated ERK and activated disparate sets of transcriptional factors. After treating the primary culture of rat hippocampal neurons and the mouse hippocampus with morphine or fentanyl for three days, they identified seven miRNAs regulated by one or both agonists. MiR-190 was downregulated by fentanyl but not by morphine. It was observed that the downregulation was attenuated by 1,4-diamino-2,3-dicyano-1,4-bis(methylthio)butadiene (U0126), a blocking agent of ERK phosphorylation. The observed effect was fentanyl-induced, but not morphine-induced. Downregulation of miR-190 was a consequence of selective ERK phosphorylation by the agonist. Additionally, the alterations in the expression of one of the miR-190 targets, NeuroD1, were found to correlate with changes in miR-190 expression. This suggests that the OPRM1 gene may regulate the NeuroD pathways by controlling the expression levels of miR-190. This aspect was expanded in a new study [105], in which it was shown that the cellular level of NeuroD1 is modulated differently: fentanyl increases NeuroD level by reducing the amount of miR-190, while morphine does not alter NeuroD level. After 3 days of treatment, morphine and fentanyl decreased the activity of Ca2/calmodulin-dependent protein kinase II (CaMKII), which phosphorylates and activates NeuroD. Zheng et al. [103] demonstrated that fentanyl decreases the miR-190 level by inhibiting the transcription of Talin2. Fentanyl-induced β-arrestin2-mediated ERK phosphorylation led to the phosphorylation of YinYang1 (YY1). In addition, YY1 phosphorylation impaired the association of YY1 with some regions on the Talin2 promoter, and this association was essential for YY1 to stimulate the transcription of the gene. Thus, fentanyl decreased the transcription of Talin2 and subsequently, the cellular level of miR-190 by inducing YY1 phosphorylation. In contrast, because morphine induces ERK phosphorylation via the protein kinase C pathway, morphine did not induce YY1 phosphorylation and had no effect on the transcription of Talin2 and the cellular content of miR-190.
Wang et al. [99] investigated the impact of opioids on the expression of anti-HIV miRNA in monocytes. Morphine-treated monocytes exhibited decreased levels of cellular anti-HIV miRNAs in comparison to untreated cells. Furthermore, morphine treatment of monocytes impeded the anti-HIV miRNA expression, induced by type I interferon (IFN). These findings align with the observation that morphine treatment of monocytes increases HIV replication. The action of morphine on the anti-HIV miRNAs and HIV could be countered by the opioid receptor antagonists (naltrexone or Cys2, Tyr3, Arg5, Pen7-amide). Additionally, the effect of morphine on miRNA expression in vitro was supported by the finding that anti-HIV miRNA levels were significantly reduced in heroin-dependent individuals in vivo (miRNA-28, 125b, 150, and 382) in peripheral blood mononuclear cells than the healthy subjects. These in vitro and in vivo findings indicate that opioid use impairs intracellular innate anti-HIV mechanisms in monocytes, contributing to cell susceptibility to HIV infection.
Another study [100] demonstrated that morphine treatment leads to differential miRNA and protein expression that have a potential impact on inflammation and oxidative stress processes that lead to the expansion of the HIV-1 viral reservoir in the CNS. Notably, hsa-miR-15b is likely to target FGF-2, resulting in the downregulation of FGF-2 secretion caused by morphine, and hsa-miR-15b was upregulated under similar treatment conditions. The induction of MCP-2 and IL-6 secretion following morphine treatment has been observed to elicit a pro-inflammatory response. Additionally, the unique aspects of morphine-induced metabolic changes in the CNS are represented by the induction of mitochondrial superoxide dismutase.
He et al. [106] examined the role of let-7 miRNAs in cellular and animal models of opioid tolerance. Inhibiting let-7 expression increased MOR protein expression in SH-SY5Y cells and the level of let-7 appeared to be inversely correlated with that of the receptor. On the other hand, let-7 expression was regulated by chronic treatment with morphine in SH-SY5Y cells and in mice that were tolerant to opioids. Moreover, knocking down let-7 partially attenuated the antinociceptive tolerance to morphine.
Morphine also decreases miR-133b expression, hence increasing the expression of its target, Pitx3, a transcription factor that activates tyrosine hydroxylase and dopamine transporter [93,107].
Yan et al. [95], revealed a functional role of miR-218 and its target, MeCP2, in the regulation of heroin-induced behavioral plasticity. MiR-218 was downregulated by chronic heroin use. MiR-218 is encoded by an intron of the Slit gene and inhibits the expression of Robo1 which play important roles in axonal growth, in motor neuron differentiation, and its loss of miR-218 cause systemic neuromuscular failure.
Jia et al. 100 [88] noted that repeated morphine exposure has been shown to induce neuronal plasticity in reward-related areas of the brain, particularly the dentate gyrus. miR-132, a CREB-induced and activation-dependent microRNA, has been suggested to be involved in neuronal plasticity by increasing neuronal dendritic branches and spinogenesis in rats treated with increasing doses of morphine injection for six consecutive days to develop morphine dependence. From their results, it is still unclear whether miR-132 is related to morphine dependence. However, they demonstrated that morphine treatment (24 h) promotes the differentiation of N2a cells stably expressing μ-opioid receptor by upregulating miR-132 expression. Moreover, inhibiting miR-132 3p (but not 5p) of the DG neurons can reverse the structural plasticity and disrupt the formation of morphine dependence in rats.
Chronic morphine intake induces a sustained increase in D1 receptor expression in glutamatergic terminals of medial prefrontal cortex (mPFC) projection neurons to the BLA but has no effect on D1 receptor expression in projection neurons from the hippocampus or thalamus to the BLA. This adaptation to chronic morphine is mediated by reduced expression of miR-105 in the mPFC, resulting in increased D1 receptor expression in glutamatergic terminals of projection neurons from the mPFC to the BLA. Projection neurons from the mPFC to the BLA are activated by abstinence-associated environmental cues morphine and overexpression of miR-105 in the mPFC leading to a reduction in D1 receptor induction [96].
Xie et al. [94], demonstrated that miR-592-3p, in the nucleus accumbens (NAc), can improve the incubation of morphine craving by targeting TMEFF1, and thus, it holds a therapeutic potential to inhibit opioid craving. Indeed, the miR-592-3p observed to be downregulated in the NAc core was linked to the incubation of morphine craving. Also, gain- and loss-of-function analyses revealed that the inhibition of miR-592-3p expression in the NAc core negatively regulated TMEFF1 expression, however, overexpression of miR-592-3p in the NAc nucleus resulted in decreased expression of TMEFF1, thereby reducing the incubation of morphine craving.
Michelhaugh et al. [101] described that also NEAT 1, NEAT 2, EMX2OS, MIAT, and MEG were upregulated in heroin abusers.
Different studies demonstrated how chronic morphine treatment increases let-7 and miR23b expression in a time- and dose-dependent manner and suppresses the association of MOR1 mRNA with polysomes. Expression of miR-24, miR-127, miR-186, and miR-222 was upregulated in the prefrontal cortex region of rats with escalated methamphetamine use. These findings suggested chronic exposure to addictive drugs may significantly alter brain miRNA profiles [90,106].
Liao et al. [92], examined the proinflammatory cytokine IL6/TNFα mRNA levels in isolated adult microglia from the brains of mice administered either morphine or saline, as an indicator of microglial activation. They demonstrate that miR-138 released in morphine-stimulated astrocyte EVs was taken up by the microglia, resulting in the activation of the TLR7/8 signaling pathway via the interaction of miR138 with TLR7.
Gu et al. [89] recruited 42 subjects who were addicted to heroin and from each subject was collected a total of 3 mL of venous blood. Microarray analysis identified 116 significantly altered miRNAs in heroin abusers; 3 miRNAs, including let-7b-5p, miR-206, and miR-486-5p, were verified to be significantly and steadily increased in heroin abusers, compared with normal controls.
3.4. miRNAs Expression in Methamphetamine Abuse
Methamphetamine (MA) is a synthetic drug of abuse with psychostimulant effects that is widely used because of its relatively low cost of manufacture. The chemical structure of methamphetamine differs from that of amphetamine by the addition of a single methyl group. This simple structural change gives the molecule enhanced neurostimulant potency, with effects that can last up to 8 h [108]. Like amphetamine, it increases motor activity, reduces appetite, and induces a general sense of well-being. The initial euphoria is followed by a marked state of agitation, which, in some individuals, may lead to violent behavior. In addition, like all substances of abuse, methamphetamine can easily lead to addiction, which, if prolonged, is typically associated with a spectrum of cognitive changes, particularly in learning and memory processes [109]. From a neurophysiological perspective, methamphetamine primarily affects the mesolimbic dopaminergic system, which is associated with motivation and reward pathways. Methamphetamine modulates the release of the neurotransmitter involved in these systems, dopamine, initially leading to increased release in certain brain areas such as the nucleus accumbens. However, during addiction, this sustained stimulation leads to chronic dopamine depletion over time, which appears to provide a neurophysiological explanation for the effects of dependence on this substance [110]. Indeed, several studies have shown that, methamphetamine dependence is associated with morphological and molecular changes in the brain, including changes in gene expression, as well as behavioral changes [111,112]. Recently, many scientists have focused on miRNAs to study their behavior and their effects on gene regulation in cases of methamphetamine abuse see Table 4.
Table 4.
miRNAs Expression in Methamphetamine abuse.
Chand et al. [112] evaluated the role of an extracellular vesicle (EV) associated with microRNA-29a-3p in chronic methamphetamine use disorder. They demonstrated that EV-miR29a-3p elicited both pro-inflammatory and injurious responses via the TLR7 pathway. The authors also provided evidence of the therapeutic relevance of the anti-inflammatory drug Ibudilast, which was shown to rescue EV biogenesis, reduce the secretion of miR-29a, and rescue synaptodendritic injury. These data support the use of Ibudilast as a therapeutic intervention for methamphetamine use disorder. Finally, they showed that miR-29a is upregulated in plasma obtained from male participants identified with methamphetamine use disorder, indicating a possible role as a potential biomarker for therapeutic interventions.
Du and colleagues [113] screened 28 differentially expressed microRNAs (miRNAs) in controlled or escalated methamphetamine use in rats. They found that miR-186 increased and miR-195 and miR-329 decreased in the prefrontal cortex of rats that self-administered methamphetamine for 1 h per session (controlled group). In contrast, miR-127, miR-186, miR-222, and miR-24 increased and miR-329 decreased in rats that self-administered methamphetamine for 6 h per session (escalated group). The levels of miR-329 decreased similarly in both groups, while the increase in miR-186 was higher in the escalated group. Bioinformatic analysis indicated that the predicted targets of these verified miRNAs are involved in the processes of neuronal apoptosis and synaptic plasticity.
Sun et al. [114] conducted a case-control association study in 400 methamphetamine users with psychotic symptoms and 448 controls. They found that miR-181a expression is increased in methamphetamine use disorder with psychosis, suppressing the expression of GluA2 on hippocampal neurons.
Zhu [115] identified two novel miRNA candidates, novel-m002C and novel-m009C, which were upregulated in the nucleus accumbens of mice exposed to daily injections of methamphetamine. Additionally, scientists observed changes in two predicted gene targets, upregulation of Arc and downregulation of Pde4c, which have been implicated in addiction and brain plasticity after drug exposure.
Kim et al. [116] analyzed the expression of miR-137 in the circulating extracellular vesicles (EVs) of patients with methamphetamine abstinence. They observed a stable reduction in circulating miR-137 across more than 20 years of methamphetamine abstinence, with a diagnostic accuracy of up to 97.7%. However, the functional roles of circulating miR-137 in animals remain unexplored to date.
Zhao et al. [117] investigated the differential expression of plasma microRNAs (miRNAs) in 124 patients with methamphetamine use disorders. They found that the expression of miR-181a, miR-15b, miR-let-7e, and miR-let-7d in plasma was downregulated compared with normal controls. Additionally, they observed a negative correlation between the expression level of these miRNAs and drug use days in the past months, suggesting that more severe drug use was associated with more aberrant miRNA expression.
Demirel et al. [118] studied samples of the ventral tegmental area and nucleus accumbens in postmortem human brain tissue from methamphetamine users. Through quantitative reverse transcription PCR, they found overexpression of miRNA let-7b-3p, which could be used as a diagnostic and therapeutic marker in methamphetamine addiction.
Li [119] compared the expression of miRNAs in the serum exosomes of methamphetamine-dependent and ketamine-dependent rats and identified ten differentially expressed co-miRNAs in the two model groups. miR-128-3p, miR-133a-3p, miR-152-3p, and miR-181a-5p are overexpressed in methamphetamine-dependent rats. From KEGG pathway analysis, they found that these target genes are mainly located in SNARE interactions in vesicular transport, amphetamine addiction, cGMP-PKG signaling pathway, dopaminergic synapse, and GABAergic synapse.
Shang and his group [120] demonstrated that changes in Ppp3r1, Cdkn1c, Fmr1, and PPARGC1A expression were negatively correlated with miR-222-3p expression in methamphetamine-induced conditioned place preference. Their findings suggest that these miRNAs may be involved in nervous system development and in the regulation of methamphetamine-induced reward-related changes in the brain.
Yang and colleagues [108] developed a rat model of methamphetamine addiction and analyzed the expression profile alterations of miRNA in the nucleus accumbens of the methamphetamine-addicted rats. A total of 40 differential miRNAs were identified: 17 upregulated and 23 downregulated that responded to methamphetamine. They performed a bioinformatic analysis of the differential miRNAs that suggested that these miRNAs may contribute to methamphetamine-induced alterations in the nervous system through the Wnt signaling pathway, axon guidance, and lysosome pathways.
Sim et al. [121] determined the global miRNA expression profiling in Nucleus Accumbens of methamphetamine-addicted rats. MiR-496-3p, miR-194-5p, miR-200b-3p, and miR-181a-5p, were found to be significantly associated with addiction. Canonical pathway analysis revealed that a high number of methamphetamine addiction-related miRNAs play important roles in cell apoptosis, cellular defense, and neurogenesis pathways.
Moradali and colleagues [122] conducted a case-control study in Iran to investigate miR-127 and miR-132 expression in methamphetamine abusers. They observed the downregulation of miR-127 and upregulation of miR-132 in patients with methamphetamine abuse compared with the control group. Authors hypothesize a correlation with CREB, GnRH, and MAPK signaling pathways involved in synaptic transmission, angiogenesis, and inflammation.
The study conducted by Wang and his team [123] revealed that miR-181a may be indirectly responsible for methamphetamine addiction and downregulation of GABAAα1 through the regulation of ERAD. In fact, methamphetamine may activate the ERAD pathway through the downregulation of miR-181 and may induce the ubiquitination degradation of GABAAα1, which is involved in the formation of addiction.
Deng et al. [136] conducted a review to summarize the role of miRNAs in the context of methamphetamine use, particularly their involvement in the reward effect and neurotoxic sequelae. They reported that several miRNAs actively participate in neuroinflammation, including miR-155-5p [125] and miR-143 [124], which can inhibit specific proinflammatory gene pathways, such as Peli1, PUMA, and NLRP3. Furthermore, they highlighted that methamphetamine could induce changes in miRNA expression in specific brain sub-regions. One of the most extensively studied regions is the Nucleus Accumbens, as it is one of the primary areas involved in regulating reward pathways. In this region, they observed relevant changes in miRNA expression during methamphetamine sensitization in mice: downregulation of miR-124 [126] and miR-3068-5p [129], associated, respectively, with the overexpression of Dicer1 and Grin1 genes, both of which are involved in synaptic plasticity. Conversely, the upregulation of miR-29c [127], miR-204-3p [128], and miR-9-5p [121] can lead to differential expression of genes involved in brain physiology, structural plasticity, and behavioral sensitization, such as Dnmt3a, Dnmt3b, Meg3, Sema3A, Plxna4, and BDNF. Similar changes were observed in other brain regions, such as the Ventral Tegmental Area [130], Dorsolateral Striatum [131], and Hippocampus [132,134], where the downregulation of miR-145, miR-129, and miR-29, and the upregulation of miR-134, miR-31-3p, and miR-183-5p were associated with alterations in the expression of HDAC, LIMK1, RhoA, and NRG1 genes.
Zhang et al. [134] explored the relationship between miR-181a and the GRIA2 gene in chronic and acute methamphetamine users. They discovered that miR-181a significantly downregulates the expression of GRIA2, a gene involved in enhancing the effects of addictive drugs. The researchers observed that chronic methamphetamine use leads to a downregulation of miR-181a expression in human serum, resulting in an overexpression of GRIA2 proteins.
Kim and colleagues [135] investigated the bidirectional miR-137/SYNCRIP pathway during methamphetamine abstinence in animal models. They observed the upregulation of miR-137 in the brain (Dorsal Striatum) and a concurrent downregulation of the same miRNA in the blood. They demonstrated that the upregulation of miR-137 during abstinence triggers the inhibition of the SYNCRIP gene, which is associated with the cognitive symptoms of methamphetamine abstinence. However, the inhibition of this gene activates intracellular miR-137 expression and exosome release, leading to an overexpression of this miRNA in the blood.
4. Discussion
Addiction to drugs and alcohol represents a complex mental health disorder, and it has become a worldwide public health burden. This is why a thorough understanding of the molecular mechanisms that regulate such a disorder could be fundamental for its treatment.
A review of the current literature revealed that miRNAs play a key role in the modulation of genes, pathways, and neural circuits that control addiction and its various stages.
This work shows how miRNAs are deeply involved in the neuroadaptive responses induced by exposure to substances of abuse; in particular, studies on changes in expression of miRNAs determined by four substances were evaluated: alcohol, cocaine, methamphetamine, and opioids.
Hundreds of miRNAs were found to be dysregulated following the intake of such substances but, most importantly, it is interesting to notice how the expression of some miRNAs turned out to be phase-specific: some miRNAs have shown to be upregulated after an acute intake of substances—such as miR-124 or miR-132 that were upregulated after one hour of alcohol intake [26]—whereas other miRNAs’ expression appeared to be modified after chronic assumption of substances, or even in the withdrawal phase [40,44,46,69,88,116].
This work also highlighted how the expression of some miRNAs seems to be linked to the intake of specific substances Table 5, Table 6, Table 7 and Table 8: for example, the expression of miRNA 146a appeared to be upregulated only after alcohol intake [18], miR150 was altered after opioids’ consumption, overexpression of miR29c was only found after methamphetamine intake [137]. On the other hand, the altered expression of some other miRNAs (in particular, miR-206a-3p, miR-124a, miR-9, miR-132, miR-382, let-7d, let-7b, miR-153, miR-133b, miR-29b, miR-181, miR-186, and miR-152) was common subsequently to the intake of all four—or at least three substances.
Table 5.
miRNAs involved in Alcohol addiction and their effects.
Table 6.
miRNAs involved in Cocaine addiction and their effects.
Table 7.
miRNAs involved in Opioid addiction and their effects.
Table 8.
miRNAs involved in Methamphetamine addiction and their effects.
Even if the processes by which miRNAs regulate and are regulated by such phenomenon remain difficult to understand, the literature showed that the most frequently mentioned targets were BDNF [4], CREB [67], and MeCP2 [68]. These pathways are known to play a crucial role in the development and maturation of the central nervous system, as also in neuronal differentiation and in formation of synapses. They are also involved in processes of plasticity and remodeling of the synapses.
Other molecules frequently targeted by miRNAs during addiction are Pitx3 [32], MAPK signaling pathway [2], and other genes involved in the processes of cell repair, neuroinflammation, apoptosis, and even in metabolic processes such as glycolysis and fatty acid metabolism [60,100].
Such pleiotropy could partly explain how altered miRNAs’ expression induced by intake of substances of abuse can determine both short-term and long-term effects on nerve functions of attention, learning, memory, behavior, and motivation. Not surprisingly, the brain regions in which miRNAs were most frequently isolated were the Nucleus Accumbens (Nac), the dorsolateral striatum (DLS), and the Ventral Tegmental Area (VTA)—some of the primary areas involved in regulating reward pathways. They were also found in the prefrontal cortex and in the hippocampus.
Interestingly, miRNAs altered in addiction were also found dysregulated in psychiatric and neurodegenerative disorders, such as schizophrenia, major depression, Alzheimer’s, and Parkinson’s disease, as such conditions determine the involvement of the same circuits and processes of addiction [22,25].
Most of the studies evaluated were conducted on animal models but, thanks to the high stability of miRNAs and the fact that they are highly conserved in human species, it has been demonstrated that they can be considered as potential biomarkers for the diagnosis of addiction. This is made possible by the fact that some miRNAs altered in addiction, could be easily extracted and measured in body fluids such as blood (as miR-181 and 221, or miR-124) [31,82], and in saliva [48].
Finally, studies conducted by Bahi et al. [21] and Bekdash et al. [38] also demonstrated that, by modulating the expression of certain miRNAs, it is possible to reduce the consumption of the substances, or even turn down the effects of withdrawal, thus suggesting a role for these little molecules as potential therapeutic tools [2].
5. Conclusions
Drugs and alcohol addiction represent a complex mental health disorder, and it has also become a worldwide public health burden. However, its diagnosis is still based on clinical criteria. With this work, we aimed to collect and unify the current knowledge on the role of miRNAs in addiction, in order to elucidate the biological processes that regulate this disorder. In recent years, miRNAs have increasingly attracted the attention of scientists and researchers due to their cardinal role in regulating biological processes, including neurogenesis, synaptic plasticity, and even neuroinflammation and apoptosis. Our study has shown that the major targets of miRNAs during addiction appear to be those involved in the development and maturation of the central nervous system, but also those related to cell cycle regulation and metabolism. It seems that the areas most frequently involved in such processes, are those of the reward circuit, and also those involved in regulating functions such as behavior, motivation, attention, and memory. In many cases, it was observed that certain miRNAs can be used as diagnostic markers for subsequent clinical outcomes. Moreover, it was observed how their pharmacological regulation could lead to tangible therapeutic benefits. However, this work shows that our biological and molecular understanding of addiction is not yet fully known. Therefore, further studies on this topic will be needed with the aim of the development of new diagnostic and effective therapeutic methods.
Author Contributions
Conceptualization, A.M. and V.F.; methodology, R.L.R.; software, A.C.; validation, J.L., A.M., and N.D.F.; formal analysis, A.C.; investigation, C.O.; resources, F.D.D.; data curation, F.D.D.; writing—original draft preparation, J.L.; writing—review and editing, F.D.D.; visualization, N.I.; supervision, A.M.; project administration, A.M.; funding acquisition, V.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AD: alcohol dependence; Ago2: Human Argonaute2; ARC: Activity-regulated cytoskeleton-associated protein; ARNT: aryl hydrocarbon receptor nuclear translocator; AUD: Alcohol use disorder; Bcl2: B-cell lymphoma 2; BDNF: brain-derived neurotropic factor; BLA: basolateral amygdala; Cacna2d2: Calcium Voltage-Gated Channel Auxiliary Subunit Alpha 2 delta 2; CaMKII: Ca2/calmodulin-dependent protein kinase II; CaMK2a: Calcium/Calmodulin Dependent Protein Kinase II Alpha; Cdkn1c: cyclin dependent kinase inhibitor 1C; cGMP-PKG: Cyclic guanosine monophosphate-dependent protein kinase or protein kinase G Dicer1; CGN: Cerebellar Granule Neurons; CHN2: chimerin 2; CLCA4: chloride channel accessory 4; CNP: 2′,3′-cyclic nucleotide 3′phosphodiesterase; CNS: central nervous system; CPP: conditioned place preference; CPU: caudate putamen; CREB: cAMP response element-binding protein; CTNNb: catenin beta; DA D3R: dopamine D3 receptor; DAT: dopamine transporters; DCX: Doublecortin; DG: dentate gyrus; DNMT: DNA Methyltransferase; DNMT3a: DNA methyltransferase 3 alpha; DNMT3b: DNA methyltransferase 3 beta; DR1: D1-like receptor; DR2: D2-like receptor; DR3: D3-like receptor; DLS: dorsolateral striatum; ELMO1: engulfment and cell motility 1; EMSX2OS: empty spiracles homeobox 2 opposite strand; ERAD: Endoplasmic Reticulum-Associated protein Degradation; ERK: extracellular signal-regulated kinase; EVs: extracellular vesicles; FGF-2: Fibroblast Growth Factors; Fmr1: Fragile X Messenger Ribonucleoprotein 1; FosB: FBJ murine osteosarcoma viral oncogene homolog B; GABAAα1: Gamma-aminobutyric acidA α1; GABAA: GABA type A receptor; GNAI3: Guanine nucleotide-binding protein G(k) subunit alpha; GluR2: glutamatic receptor 2; GRα: glucocorticoid receptor α; GRIA2: Glutamate ionotropic receptor AMPA type subunit 2; Grin1: Glutamate Ionotropic Receptor NMDA Type Subunit 1; GSTT2: Glutathione S-Transferase Theta 2; HDAC2: Histone Deacetylase 2; HDAC4: histone deacetylase 4; HDA5: histone deacetylase 5; HIP: hippocampus; HKDC1: Hexokinase Domain Containing 1; Hpf: hours post fertilization; IL-6: interleukin 6; KLF: Kruppel like factor; KTF: KLF transcription factor; KYNU: Kynureninase; LIMK1: Lin-11, Isl-1 and Mec-3 Domain Kinase 1; LIMK1: Lin-11, Isl-1 and Mec-3 Domain Kinase 1; LIPG: Lipase G; MAPK: mitogen-activated protein kinase; MCP-2: Monocyte Chemotactic Protein 2; Me-1: Malik enzyme 1; MeCP2: methyl CpG binding protein 2; Meg3: Maternally Expressed Gene 3; MEIS1: Meis Homeobox 1; MIAT: myocardial infarction associated transcript; MMT: methadone maintenance treatment; MOR: Opioid Receptor Mu; mPFC: medial prefrontal cortex; MYD88: Myeloid Differentiation Primary Response Protein; N2a: neuro 2A cell; NAc: Nucleus Accumbens; NDE1: neurodevelopment protein 1; NDUFA9: NADH: Ubiquinone Oxidoreductase Subunit A9; NEAT1: nuclear enriched abundant transcript 1; NEAT2: nuclear enriched abundant transcript 2; NFAT: Nuclear Factor of Activated B cells; NFIB: Nuclear Factor I B; NF-kβ: nuclear factor kβ; NLRP3: NLR Family Pyrin Domain Containing 3; Npas: Neuronal PAS Domain Protein; NRG1: Neuregulin 1; NZF-2b: neural zinc finger transcription factor 2b; OUD: Opioids use disorder; p53: Protein 53; PARP-1: poly (ADP-ribose) polymerase-1; PBMC: peripheral blood mononuclear cells; PD: Parkinson’s Disease; Pde: Phosphodeisterase; Pde4c: Phosphodiesterase 4C; Peli1: Pellino E3 Ubiquitin Protein Ligase 1; Per2: Period Circadian Regulator 2; PFC: prefrontal cortex; Pitx3: Paired Like Homeodomain 3; PKC: protein kinase C; PKR: double-stranded RNA-sctivated protein kinase; PLP1: proteolipid protein 1; Plxna4: Plexin A4; PPARGC1A: Peroxisome Proliferator Activated Receptor Gamma Coactivator 1 Alpha; Ppp3r1: Protein Phosphatase 3 Regulatory Subunit B, Alpha; PRTG: Protogenin; Pum2: Pumilio RNA Binding Family Member 2; PUMA: p53 upregulated modulator of apoptosis; QKI: protein quaking; RAX: PKR activator X; REST: RE1 Silencing Transcription Factor; RhoA: Ras Homolog Family Member A; ROB: rest of the brain; Rorb: Retinoic acid receptor-related orphan receptor B; ROS: reactive oxygen species; Sema3A: Semaphorin 3A; SEMA4D1: semaphorin 4D1; SH-SY5Y: human neuroblastoma cell; SNARE: Soluble N-ethylmaleimide-sensitive factor-attachment protein receptor; SSH2: Slingshot Protein Phosphatase 2; SYNCRIP: Synaptotagmin Binding Cytoplasmic RNA Interacting Protein; SP1: Specificity protein 1; STAT: Signal transducer and activator of transcription; STAT3: signal transducer and activator of transcription 3; TAOK1: TAO Kinase 1; Tcf: Transcription factor; TEAD1: TEA Domain Transcription Factor 1; TH: Tyrosine hydroxylases; THBS2: thromspondin 2; TLR: toll-like receptor; TLR7: Toll-like receptor 7; TMCC3: transmembrane and coiled-coil domain family 3; TMEFF1: Tomoregulin-1; TMTC2: transmembrane and tetratricopeptide repeat containing 2; TrkB: tropomyosin-related kinase B; UGT8: UDP glycosyltransferase 8; uPA: Urokinase-type Plasminogen Activator; USF3: Upstream Transcription Factor Family Member 3; VAMP: Vesicle Associated Membrane Protein; VTA: ventral tegmental area; YY1: YinYang1; ZBTB20: zinc finger and BTB domain containing 20; ZC3H6: Zinc Finger CCCH-Type Containing 6; ZFDOR: zebrafish µ-opioid receptor; ZO-1: tight junction protein.
References
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 Criteria for Substance Use Disorders: Recommendations and Rationale. Am. J. Pathol. 2013, 170, 834–851. [Google Scholar] [CrossRef] [PubMed]
- Gowen, A.M.; Odegaard, K.E.; Hernandez, J.; Chand, S.; Koul, S.; Pendyala, G.; Yelamanchili, S.V. Role of microRNAs in the Pathophysiology of Addiction. Wiley Interdiscip. Rev. RNA 2021, 12, e1637. [Google Scholar] [CrossRef] [PubMed]
- Bardo, M.T.; Donohew, R.L.; Harrington, N.G. Psychobiology of Novelty Seeking and Drug Seeking Behavior. Behav. Brain Res. 1996, 77, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Heyer, M.P.; Kenny, P.J. Corticostriatal microRNAs in Addiction. Brain Res. 2015, 1628, 2–16. [Google Scholar] [CrossRef] [PubMed]
- NCHS. Provisional Data Shows U.S. Drug Overdose Deaths Top 100,000 in 2022. Available online: https://blogs.cdc.gov/nchs/2023/05/18/7365/ (accessed on 17 October 2023).
- Mancinelli, R.; Guiducci, M.S. Procedural aspects and interpretation problems in the analysis of drugs of abuse. Ann. Ist. Super. Sanita 2002, 38, 305–313. [Google Scholar] [PubMed]
- Ajonijebu, D.; Abboussi, O.; Russell, V.; Mabandla, M.; Daniels, W. Epigenetics: A Link between Addiction and Social Environment. Cell. Mol. Life Sci. 2017, 74, 2735–2747. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.A.; Utrankar, A.; Reyes, J.A.; Simons, D.D.; Kosten, T.R. Epigenetics of Drug Abuse: Predisposition or Response. Pharmacogenomics 2012, 13, 1149–1160. [Google Scholar] [CrossRef]
- Giannouli, V.; Ivanova, D. Codependency in Mothers of Addicted Persons: Cross-Cultural Differences between Greece and Bulgaria. Eur. Psychiatry 2016, 33, S622–S623. [Google Scholar] [CrossRef]
- Whitesell, M.; Bachand, A.; Peel, J.; Brown, M. Familial, Social, and Individual Factors Contributing to Risk for Adolescent Substance Use. J. Addict. 2013, 2013, 579310. [Google Scholar] [CrossRef]
- Brown, B.S. Drug Use--Chronic and Relapsing or a Treatable Condition? Subst. Use Misuse 1998, 33, 2515–2520. [Google Scholar] [CrossRef]
- O’Brien, C.P.; McLellan, A.T. Myths about the Treatment of Addiction. Lancet 1996, 347, 237–240. [Google Scholar] [CrossRef] [PubMed]
- McLellan, A.T. Have We Evaluated Addiction Treatment Correctly? Implications from a Chronic Care Perspective. Addiction 2002, 97, 249–252. [Google Scholar] [CrossRef] [PubMed]
- McLellan, A.T.; Lewis, D.C.; O’Brien, C.P.; Kleber, H.D. Drug Dependence, a Chronic Medical Illness: Implications for Treatment, Insurance, and Outcomes Evaluation. JAMA 2000, 284, 1689–1695. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.W.; Kenny, P.J. MicroRNAs Regulate Synaptic Plasticity Underlying Drug Addiction: MicroRNAs Regulate Synaptic Plasticity Underlying Drug Addiction. Genes. Brain Behav. 2018, 17, e12424. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.M.; Gururajan, A.; Dinan, T.G.; Kenny, P.J.; Cryan, J.F. All Roads Lead to the miRNome: miRNAs Have a Central Role in the Molecular Pathophysiology of Psychiatric Disorders. Trends Pharmacol. Sci. 2016, 37, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Health Care Interventions: Explanation and Elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef] [PubMed]
- Lewohl, J.M.; Nunez, Y.O.; Dodd, P.R.; Tiwari, G.R.; Harris, R.A.; Mayfield, R.D. Up-Regulation of MicroRNAs in Brain of Human Alcoholics: MICRORNAS IN HUMAN ALCOHOLICS. Alcohol. Clin. Exp. Res. 2011, 35, 1928–1937. [Google Scholar] [CrossRef]
- Ehinger, Y.; Phamluong, K.; Darevesky, D.; Welman, M.; Moffat, J.J.; Sakhai, S.A.; Whiteley, E.L.; Berger, A.L.; Laguesse, S.; Farokhnia, M.; et al. Differential Correlation of Serum BDNF and microRNA Content in Rats with Rapid or Late Onset of Heavy Alcohol Use. Addiction Biology 2021, 26, e12890. [Google Scholar] [CrossRef]
- Tapocik, J.D.; Solomon, M.; Flanigan, M.; Meinhardt, M.; Barbier, E.; Schank, J.R.; Schwandt, M.; Sommer, W.H.; Heilig, M. Coordinated Dysregulation of mRNAs and microRNAs in the Rat Medial Prefrontal Cortex Following a History of Alcohol Dependence. Pharmacogenomics J. 2013, 13, 286–296. [Google Scholar] [CrossRef]
- Bahi, A.; Dreyer, J.-L. Striatal Modulation of BDNF Expression Using microRNA124a-Expressing Lentiviral Vectors Impairs Ethanol-Induced Conditioned-Place Preference and Voluntary Alcohol Consumption. Eur. J. Neurosci. 2013, 38, 2328–2337. [Google Scholar] [CrossRef]
- Darcq, E.; Warnault, V.; Phamluong, K.; Besserer, G.M.; Liu, F.; Ron, D. MicroRNA-30a-5p in the Prefrontal Cortex Controls the Transition from Moderate to Excessive Alcohol Consumption. Mol. Psychiatry 2015, 20, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Pietrzykowski, A.Z. Coinciding Revolutions: How Discovery of Non-Coding DNA and RNA Can Change Our Understanding of Addiction. Front. Gene. 2012, 3, 271. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Fu, C.; Lin, L.; Liu, S.; Su, X.; Li, A.; Wu, Q.; Jia, C.; Zhang, P.; Chen, L.; et al. miR-124 and miR-9 Mediated Downregulation of HDAC5 Promotes Neurite Development through Activating MEF2C-GPM6A Pathway. J. Cell Physiol. 2018, 233, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Li, M.D.; Van Der Vaart, A.D. MicroRNAs in Addiction: Adaptation’s Middlemen? Mol. Psychiatry 2011, 16, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Mizuo, K.; Okazaki, S. Acute Ethanol Administration Increases Mir-124 Expression via Histone Acetylation in the Brain. J. Alcohol. Drug Depend. 2016, 4, 2. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Liu, X.; Qin, S.; Guan, Y.; Liu, Y.; Cheng, Y.; Chen, X.; Li, W.; Wang, S.; et al. Micro RNA Expression Profile and Functional Analysis Reveal That Mi R -382 Is a Critical Novel Gene of Alcohol Addiction. EMBO Mol. Med. 2013, 5, 1402–1414. [Google Scholar] [CrossRef] [PubMed]
- Most, D.; Leiter, C.; Blednov, Y.A.; Harris, R.A.; Mayfield, R.D. Synaptic microRNAs Coordinately Regulate Synaptic mRNAs: Perturbation by Chronic Alcohol Consumption. Neuropsychopharmacology 2016, 41, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Coleman, L.G.; Zou, J.; Crews, F.T. Microglial-Derived miRNA Let-7 and HMGB1 Contribute to Ethanol-Induced Neurotoxicity via TLR7. J. Neuroinflammation 2017, 14, 22. [Google Scholar] [CrossRef]
- Balaraman, S.; Winzer-Serhan, U.H.; Miranda, R.C. Opposing Actions of Ethanol and Nicotine on MicroRNAs Are Mediated by Nicotinic Acetylcholine Receptors in Fetal Cerebral Cortical-Derived Neural Progenitor Cells. Alcohol. Clin. Exp. Res. 2012, 36, 1669–1677. [Google Scholar] [CrossRef]
- Alhaddad, H.; Gordon, D.M.; Bell, R.L.; Jarvis, E.E.; Kipp, Z.A.; Hinds, T.D.; Sari, Y. Chronic Ethanol Consumption Alters Glucocorticoid Receptor Isoform Expression in Stress Neurocircuits and Mesocorticolimbic Brain Regions of Alcohol-Preferring Rats. Neuroscience 2020, 437, 107–116. [Google Scholar] [CrossRef]
- Lim, Y.; Beane-Ebel, J.E.; Tanaka, Y.; Ning, B.; Husted, C.R.; Henderson, D.C.; Xiang, Y.; Park, I.-H.; Farrer, L.A.; Zhang, H. Exploration of Alcohol Use Disorder-Associated Brain miRNA–mRNA Regulatory Networks. Transl. Psychiatry 2021, 11, 504. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Banan, A.; Forsyth, C.B.; Fields, J.Z.; Lau, C.K.; Zhang, L.J.; Keshavarzian, A. Effect of Alcohol on miR-212 Expression in Intestinal Epithelial Cells and Its Potential Role in Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 2008, 32, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.M.; Chung, D.D.; Pinson, M.R.; Salem, N.A.; Eaves, S.E.; Miranda, R.C. Ethanol Exposure Increases miR-140 in Extracellular Vesicles: Implications for Fetal Neural Stem Cell Proliferation and Maturation. Alcohol. Clin. Exp. Res. 2019, 43, 1414–1426. [Google Scholar] [CrossRef] [PubMed]
- Sathyan, P.; Golden, H.B.; Miranda, R.C. Competing Interactions between Micro-RNAs Determine Neural Progenitor Survival and Proliferation after Ethanol Exposure: Evidence from an Ex Vivo Model of the Fetal Cerebral Cortical Neuroepithelium. J. Neurosci. 2007, 27, 8546–8557. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, M.; Li, H.; Frank, J.A.; Dai, L.; Liu, H.; Chen, G. MicroRNA-29b Regulates Ethanol-Induced Neuronal Apoptosis in the Developing Cerebellum through SP1/RAX/PKR Cascade. J. Biol. Chem. 2014, 289, 10201–10210. [Google Scholar] [CrossRef] [PubMed]
- Asquith, M.; Pasala, S.; Engelmann, F.; Haberthur, K.; Meyer, C.; Park, B.; Grant, K.A.; Messaoudi, I. Chronic Ethanol Consumption Modulates Growth Factor Release, Mucosal Cytokine Production, and MicroRNA Expression in Nonhuman Primates. Alcohol. Clin. Exp. Res. 2014, 38, 980–993. [Google Scholar] [CrossRef]
- Bekdash, R.A.; Harrison, N.L. Downregulation of Gabra4 Expression during Alcohol withdrawal Is Mediated by Specific Micro RNA s in Cultured Mouse Cortical Neurons. Brain Behav. 2015, 5, e00355. [Google Scholar] [CrossRef]
- Choi, M.R.; Han, J.S.; Jin, Y.-B.; Lee, S.-R.; Choi, I.Y.; Lee, H.; Cho, H.; Kim, D.-J. Differential Expression of microRNAs in the Hippocampi of Male and Female Rodents after Chronic Alcohol Administration. Biol. Sex. Differ. 2020, 11, 65. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Y.; Carreon, S.; Qiang, M. Chronic Intermittent Ethanol Exposure and Its Removal Induce a Different miRNA Expression Pattern in Primary Cortical Neuronal Cultures: CIE EXPOSURE ALTERS miRNA EXPRESSION. Alcohol. Clin. Exp. Res. 2012, 36, 1058–1066. [Google Scholar] [CrossRef]
- Santos-Bezerra, D.P.; Cavaleiro, A.M.; Santos, A.S.; Suemoto, C.K.; Pasqualucci, C.A.; Jacob-Filho, W.; Leite, R.E.P.; Passarelli, M.; Marie, S.K.N.; Machado, U.F.; et al. Alcohol Use Disorder Is Associated with Upregulation of MicroRNA-34a and MicroRNA-34c in Hippocampal Postmortem Tissue. Alcohol. Clin. Exp. Res. 2021, 45, 64–68. [Google Scholar] [CrossRef]
- Manzardo, A.M.; Gunewardena, S.; Butler, M.G. Over-Expression of the miRNA Cluster at Chromosome 14q32 in the Alcoholic Brain Correlates with Suppression of Predicted Target mRNA Required for Oligodendrocyte Proliferation. Gene 2013, 526, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qin, F.; Han, S.; Li, S.; Zhao, Y.; Wang, H.; Tian, J.; Cen, X. MicroRNAs in Drug Addiction: Current Status and Future Perspectives. Pharmacol. Ther. 2022, 236, 108215. [Google Scholar] [CrossRef] [PubMed]
- Most, D.; Workman, E.; Harris, R.A. Synaptic Adaptations by Alcohol and Drugs of Abuse: Changes in microRNA Expression and mRNA Regulation. Front. Mol. Neurosci. 2014, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jing, X.; Wang, G. MicroRNAs as Regulators of Drug Abuse and Immunity. Cent. Eur. J. Immunol. 2016, 4, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Most, D.; Salem, N.A.; Tiwari, G.R.; Blednov, Y.A.; Mayfield, R.D.; Harris, R.A. Silencing Synaptic MicroRNA-411 Reduces Voluntary Alcohol Consumption in Mice. Addict. Biol. 2019, 24, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, H. RNA m6A Modification Changes in Postmortem Nucleus Accumbens of Subjects with Alcohol Use Disorder: A Pilot Study. Genes 2022, 13, 958. [Google Scholar] [CrossRef]
- Rosato, A.J.; Chen, X.; Tanaka, Y.; Farrer, L.A.; Kranzler, H.R.; Nunez, Y.Z.; Henderson, D.C.; Gelernter, J.; Zhang, H. Salivary microRNAs Identified by Small RNA Sequencing and Machine Learning as Potential Biomarkers of Alcohol Dependence. Epigenomics 2019, 11, 739–749. [Google Scholar] [CrossRef]
- Roque Bravo, R.; Faria, A.C.; Brito-da-Costa, A.M.; Carmo, H.; Mladěnka, P.; Dias Da Silva, D.; Remião, F.; on behalf of The OEMONOM Researchers. Cocaine: An Updated Overview on Chemistry, Detection, Biokinetics, and Pharmacotoxicological Aspects Including Abuse Pattern. Toxins 2022, 14, 278. [Google Scholar] [CrossRef]
- Barreto-Valer, K.; López-Bellido, R.; Macho Sánchez-Simón, F.; Rodríguez, R.E. Modulation by Cocaine of Dopamine Receptors through miRNA-133b in Zebrafish Embryos. PLoS ONE 2012, 7, e52701. [Google Scholar] [CrossRef]
- Bastle, R.M.; Oliver, R.J.; Gardiner, A.S.; Pentkowski, N.S.; Bolognani, F.; Allan, A.M.; Chaudhury, T.; St. Peter, M.; Galles, N.; Smith, C.; et al. In Silico Identification and in Vivo Validation of miR-495 as a Novel Regulator of Motivation for Cocaine That Targets Multiple Addiction-Related Networks in the Nucleus Accumbens. Mol. Psychiatry 2018, 23, 434–443. [Google Scholar] [CrossRef]
- Cabana-Domínguez, J.; Arenas, C.; Cormand, B.; Fernàndez-Castillo, N. MiR-9, miR-153 and miR-124 Are down-Regulated by Acute Exposure to Cocaine in a Dopaminergic Cell Model and May Contribute to Cocaine Dependence. Transl. Psychiatry 2018, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Dreyer, J.-L. microRNAs miR-124, Let-7d and miR-181a Regulate Cocaine-Induced Plasticity. Mol. Cell. Neurosci. 2009, 42, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Dreyer, J.-L. Regulation of MiR-124, Let-7d, and MiR-181a in the Accumbens Affects the Expression, Extinction, and Reinstatement of Cocaine-Induced Conditioned Place Preference. Neuropsychopharmacology 2011, 36, 1149–1164. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-L.; Liu, H.; Guan, X. Changes in microRNA Expression Profile in Hippocampus during the Acquisition and Extinction of Cocaine-Induced Conditioned Place Preference in Rats. J. Biomed. Sci. 2013, 20, 96. [Google Scholar] [CrossRef] [PubMed]
- Chivero, E.T.; Liao, K.; Niu, F.; Tripathi, A.; Tian, C.; Buch, S.; Hu, G. Engineered Extracellular Vesicles Loaded With miR-124 Attenuate Cocaine-Mediated Activation of Microglia. Front. Cell Dev. Biol. 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Balasubramaniam, M.; Rana, T.; Godino, A.; Peck, E.G.; Goodwin, J.S.; Villalta, F.; Calipari, E.S.; Nestler, E.J.; Dash, C.; et al. Poly (ADP-Ribose) Polymerase-1 (PARP-1) Induction by Cocaine Is Post-Transcriptionally Regulated by miR-125b. eNeuro 2017, 4, ENEURO.0089-17.2017. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Balasubramaniam, M.; Martínez-Rivera, F.J.; Godino, A.; Peck, E.G.; Patnaik, S.; Suar, M.; Calipari, E.S.; Nestler, E.J.; Villalta, F.; et al. Cocaine-Regulated microRNA miR-124 Controls Poly (ADP-Ribose) Polymerase-1 Expression in Neuronal Cells. Sci. Rep. 2020, 10, 11197. [Google Scholar] [CrossRef]
- Doke, M.; Kashanchi, F.; Khan, M.A.; Samikkannu, T. HIV-1 Tat and Cocaine Impact Astrocytic Energy Reservoir Influence on miRNA Epigenetic Regulation. Genomics 2021, 113, 3461–3475. [Google Scholar] [CrossRef]
- Doke, M.; McLaughlin, J.P.; Cai, J.J.; Pendyala, G.; Kashanchi, F.; Khan, M.A.; Samikkannu, T. HIV-1 Tat and Cocaine Impact Astrocytic Energy Reservoirs and Epigenetic Regulation by Influencing the LINC01133-Hsa-miR-4726-5p-NDUFA9 Axis. Mol. Ther.-Nucleic Acids 2022, 29, 243–258. [Google Scholar] [CrossRef]
- Domingo-Rodriguez, L.; Cabana-Domínguez, J.; Fernàndez-Castillo, N.; Cormand, B.; Martín-García, E.; Maldonado, R. Differential Expression of miR-1249-3p and miR-34b-5p between Vulnerable and Resilient Phenotypes of Cocaine Addiction. Addict. Biol. 2022, 27, e13201. [Google Scholar] [CrossRef]
- Dykxhoorn, D.M.; Wang, H.; Da Fonseca Ferreira, A.; Wei, J.; Dong, C. MicroRNA-423-5p Mediates Cocaine-Induced Smooth Muscle Cell Contraction by Targeting Cacna2d2. Int. J. Mol. Sci. 2023, 24, 6584. [Google Scholar] [CrossRef] [PubMed]
- Forget, B.; Garcia, E.M.; Godino, A.; Rodriguez, L.D.; Kappes, V.; Poirier, P.; Andrianarivelo, A.; Marchan, E.S.; Allichon, M.-C.; Marias, M.; et al. Cell-Type- and Region-Specific Modulation of Cocaine Seeking by Micro-RNA-1 in Striatal Projection Neurons. Mol. Psychiatry 2022, 27, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Gawliński, D.; Gawlińska, K.; Frankowska, M.; Filip, M. Cocaine and Its Abstinence Condition Modulate Striatal and Hippocampal Wnt Signaling in a Male Rat Model of Drug Self-Administration. Int. J. Mol. Sci. 2022, 23, 14011. [Google Scholar] [CrossRef] [PubMed]
- Giannotti, G.; Caffino, L.; Calabrese, F.; Racagni, G.; Riva, M.A.; Fumagalli, F. Prolonged Abstinence from Developmental Cocaine Exposure Dysregulates BDNF and Its Signaling Network in the Medial Prefrontal Cortex of Adult Rats. Int. J. Neuropsychopharm. 2014, 17, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.-L.; Periyasamy, P.; Liao, K.; Kook, Y.H.; Niu, F.; Callen, S.E.; Buch, S. Cocaine-Mediated Downregulation of Microglial miR-124 Expression Involves Promoter DNA Methylation. Epigenetics 2016, 11, 819–830. [Google Scholar] [CrossRef]
- Hollander, J.A.; Im, H.-I.; Amelio, A.L.; Kocerha, J.; Bali, P.; Lu, Q.; Willoughby, D.; Wahlestedt, C.; Conkright, M.D.; Kenny, P.J. Striatal microRNA Controls Cocaine Intake through CREB Signalling. Nature 2010, 466, 197–202. [Google Scholar] [CrossRef]
- Im, H.-I.; Hollander, J.A.; Bali, P.; Kenny, P.J. MeCP2 Controls BDNF Expression and Cocaine Intake through Homeostatic Interactions with microRNA-212. Nat. Neurosci. 2010, 13, 1120–1127. [Google Scholar] [CrossRef]
- Kumaresan, V.; Lim, Y.; Juneja, P.; Tipton, A.E.; de Guglielmo, G.; Carrette, L.L.G.; Kallupi, M.; Maturin, L.; Liu, Y.; George, O.; et al. Abstinence from Escalation of Cocaine Intake Changes the microRNA Landscape in the Cortico-Accumbal Pathway. Biomedicines 2023, 11, 1368. [Google Scholar] [CrossRef]
- Li, Y.; Lu, X.; Nie, J.; Hu, P.; Ge, F.; Yuan, T.-F.; Guan, X. MicroRNA134 of Ventral Hippocampus Is Involved in Cocaine Extinction-Induced Anxiety-like and Depression-like Behaviors in Mice. Mol. Ther.-Nucleic Acids 2020, 19, 937–950. [Google Scholar] [CrossRef]
- López-Bellido, R.; Barreto-Valer, K.; Sánchez-Simón, F.M.; Rodríguez, R.E. Cocaine Modulates the Expression of Opioid Receptors and miR-Let-7d in Zebrafish Embryos. PLoS ONE 2012, 7, e50885. [Google Scholar] [CrossRef]
- Mantri, C.K.; Pandhare Dash, J.; Mantri, J.V.; Dash, C.C.V. Cocaine Enhances HIV-1 Replication in CD4+ T Cells by Down-Regulating MiR-125b. PLoS ONE 2012, 7, e51387. [Google Scholar] [CrossRef] [PubMed]
- Napuri, J.; Pilakka-Kanthikeel, S.; Raymond, A.; Agudelo, M.; Yndart-Arias, A.; Saxena, S.K.; Nair, M. Cocaine Enhances HIV-1 Infectivity in Monocyte Derived Dendritic Cells by Suppressing microRNA-155. PLoS ONE 2013, 8, e83682. [Google Scholar] [CrossRef] [PubMed]
- Nudelman, A.S.; DiRocco, D.P.; Lambert, T.J.; Garelick, M.G.; Le, J.; Nathanson, N.M.; Storm, D.R. Neuronal Activity Rapidly Induces Transcription of the CREB-Regulated microRNA-132, in Vivo. Hippocampus 2009, 20, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy, P.; Liao, K.; Kook, Y.H.; Niu, F.; Callen, S.E.; Guo, M.-L.; Buch, S. Cocaine-Mediated Downregulation of miR-124 Activates Microglia by Targeting KLF4 and TLR4 Signaling. Mol. Neurobiol. 2018, 55, 3196–3210. [Google Scholar] [CrossRef] [PubMed]
- Quinn, R.K.; Brown, A.L.; Goldie, B.J.; Levi, E.M.; Dickson, P.W.; Smith, D.W.; Cairns, M.J.; Dayas, C.V. Distinct miRNA Expression in Dorsal Striatal Subregions Is Associated with Risk for Addiction in Rats. Transl. Psychiatry 2015, 5, e503. [Google Scholar] [CrossRef]
- Quinn, R.K.; James, M.H.; Hawkins, G.E.; Brown, A.L.; Heathcote, A.; Smith, D.W.; Cairns, M.J.; Dayas, C.V. Temporally Specific miRNA Expression Patterns in the Dorsal and Ventral Striatum of Addiction-Prone Rats: miRNA Expression in Addiction. Addict. Biol. 2018, 23, 631–642. [Google Scholar] [CrossRef]
- Sadakierska-Chudy, A.; Frankowska, M.; Miszkiel, J.; Wydra, K.; Jastrzębska, J.; Filip, M. Prolonged Induction of miR-212/132 and REST Expression in Rat Striatum Following Cocaine Self-Administration. Mol. Neurobiol. 2017, 54, 2241–2254. [Google Scholar] [CrossRef]
- Schaefer, A.; Im, H.-I.; Venø, M.T.; Fowler, C.D.; Min, A.; Intrator, A.; Kjems, J.; Kenny, P.J.; O’Carroll, D.; Greengard, P. Argonaute 2 in Dopamine 2 Receptor–Expressing Neurons Regulates Cocaine Addiction. J. Exp. Med. 2010, 207, 1843–1851. [Google Scholar] [CrossRef]
- Vannan, A.; Powell, G.L.; Dell’Orco, M.; Wilson, M.A.; Perrone-Bizzozero, N.I.; Neisewander, J.L. microRNA Regulation Related to the Protective Effects of Environmental Enrichment against Cocaine-Seeking Behavior. Drug Alcohol. Depend. 2021, 221, 108585. [Google Scholar] [CrossRef]
- Tobón, K.E.; Catuzzi, J.E.; Cote, S.R.; Sonaike, A.; Kuzhikandathil, E.V. Post-Transcriptional Regulation of Dopamine D1 Receptor Expression in Caudate-Putamen of Cocaine-Sensitized Mice. Eur. J. Neurosci. 2015, 42, 1849–1857. [Google Scholar] [CrossRef]
- Viola, T.W.; Wearick-Silva, L.E.; De Azeredo, L.A.; Centeno-Silva, A.; Murphy, C.; Marshall, P.; Li, X.; Singewald, N.; Garcia, F.; Bredy, T.W.; et al. Increased Cocaine-Induced Conditioned Place Preference during Periadolescence in Maternally Separated Male BALB/c Mice: The Role of Cortical BDNF, microRNA-212, and MeCP2. Psychopharmacology 2016, 233, 3279–3288. [Google Scholar] [CrossRef] [PubMed]
- Viola, T.W.; Heberle, B.A.; Zaparte, A.; Sanvicente-Vieira, B.; Wainer, L.M.; Fries, G.R.; Walss-Bass, C.; Grassi-Oliveira, R. Peripheral Blood microRNA Levels in Females with Cocaine Use Disorder. J. Psychiatr. Res. 2019, 114, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-F.; Wang, J.; Lv, F.B.; Song, Q. Functions of microRNA in Response to Cocaine Stimulation. Genet. Mol. Res. 2013, 12, 6160–6167. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wang, H.; Wei, J.; Sartor, G.C.; Bao, M.M.; Pierce, C.T.; Wahlestedt, C.R.; Dykxhoorn, D.M.; Dong, C. Cocaine Exposure Increases Blood Pressure and Aortic Stiffness via the miR-30c-5p–Malic Enzyme 1–Reactive Oxygen Species Pathway. Hypertension 2018, 71, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.W.; O’Brien, C. Drug Addiction as a Pathology of Staged Neuroplasticity. Neuropsychopharmacol. 2008, 33, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, J.-L. New Insights into the Roles of microRNAs in Drug Addiction and Neuroplasticity. Genome Med. 2010, 2, 92. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Wang, X.; Zhang, H.; Wang, X.; Ma, H.; Yang, M.; Li, Y.; Cui, C. MicroRNA-132 Is Involved in Morphine Dependence via Modifying the Structural Plasticity of the Dentate Gyrus Neurons in Rats. Addict. Biol. 2022, 27, e13086. [Google Scholar] [CrossRef]
- Gu, W.-J.; Zhang, C.; Zhong, Y.; Luo, J.; Zhang, C.-Y.; Zhang, C.; Wang, C. Altered Serum microRNA Expression Profile in Subjects with Heroin and Methamphetamine Use Disorder. Biomed. Pharmacother. 2020, 125, 109918. [Google Scholar] [CrossRef]
- Liu, H.; Xu, W.; Feng, J.; Ma, H.; Zhang, J.; Xie, X.; Zhuang, D.; Shen, W.; Liu, H.; Zhou, W. Increased Expression of Plasma miRNA-320a and Let-7b-5p in Heroin-Dependent Patients and Its Clinical Significance. Front. Psychiatry 2021, 12, 679206. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, M.; Lin, Z.; Liu, H.; Ma, H.; Hong, Q.; Gui, D.; Feng, J.; Liu, Y.; Zhou, W.; et al. Increased Expression of Plasma hsa-miR-181a in Male Patients with Heroin Addiction Use Disorder. J. Clin. Lab. Anal. 2020, 34, e23486. [Google Scholar] [CrossRef]
- Liao, K.; Niu, F.; Hu, G.; Yang, L.; Dallon, B.; Villarreal, D.; Buch, S. Morphine-mediated Release of miR-138 in Astrocyte-derived Extracellular Vesicles Promotes Microglial Activation. J. Extracell. Vesicle 2020, 10, e12027. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.W.; Huang, T.L.; Tsai, M.C. Decreased Level of Blood MicroRNA-133b in Men with Opioid Use Disorder on Methadone Maintenance Therapy. J. Clin. Med. 2019, 8, 1105. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Zhang, J.; Ma, C.; Yu, H.; Ni, Z.; Cong, B.; Wen, D. Roles of miR-592-3p and Its Target Gene, TMEFF1, in the Nucleus Accumbens During Incubation of Morphine Craving. Int. J. Neuropsychopharmacol. 2022, 25, 412–424. [Google Scholar] [CrossRef]
- Yan, B.; Hu, Z.; Yao, W.; Le, Q.; Xu, B.; Liu, X.; Ma, L. MiR-218 Targets MeCP2 and Inhibits Heroin Seeking Behavior. Sci. Rep. 2017, 7, 40413. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, J.; Yang, H.; Cui, D.; Song, J.; Ma, Q.; Luan, W.; Lai, B.; Ma, L.; Chen, M.; et al. Memory Retrieval in Addiction: A Role for miR-105-Mediated Regulation of D1 Receptors in mPFC Neurons Projecting to the Basolateral Amygdala. BMC Biol. 2017, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Im, H.-I.; Moon, C. Intravenous Morphine Self-Administration Alters Accumbal microRNA Profiles in the Mouse Brain. Neural Regen. Res. 2018, 13, 77. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, L.; Law, P.-Y.; Wei, L.-N.; Loh, H.H. Long-Term Morphine Treatment Decreases the Association of μ-Opioid Receptor (MOR1) mRNA with Polysomes through miRNA23b. Mol. Pharmacol. 2009, 75, 744–750. [Google Scholar] [CrossRef]
- Wang, X.; Ye, L.; Zhou, Y.; Liu, M.-Q.; Zhou, D.-J.; Ho, W.-Z. Inhibition of Anti-HIV MicroRNA Expression. Am. J. Pathol. 2011, 178, 41–47. [Google Scholar] [CrossRef]
- Dave, R.S.; Khalili, K. Morphine Treatment of Human Monocyte-Derived Macrophages Induces Differential miRNA and Protein Expression: Impact on Inflammation and Oxidative Stress in the Central Nervous System. J. Cell. Biochem. 2010, 110, 834–845. [Google Scholar] [CrossRef]
- Michelhaugh, S.K.; Lipovich, L.; Blythe, J.; Jia, H.; Kapatos, G.; Bannon, M.J. Mining Affymetrix Microarray Data for Long Non-Coding RNAs: Altered Expression in the Nucleus Accumbens of Heroin Abusers: Long Non-Coding RNAs in Heroin Abusers. J. Neurochem. 2011, 116, 459–466. [Google Scholar] [CrossRef]
- Wu, Q.; Law, P.-Y.; Wei, L.-N.; Loh, H.H. Post-Transcriptional Regulation of Mouse μ Opioid Receptor (MOR1) via Its 3′ Untranslated Region: A Role for microRNA23b. FASEB J. 2008, 22, 4085–4095. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, Y.; Zhang, X.; Chu, J.; Loh, H.H.; Law, P.-Y. μ-Opioid Receptor Agonists Differentially Regulate the Expression of miR-190 and NeuroD. Mol. Pharmacol. 2010, 77, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, Y.; Chu, J.; Kam, A.Y.; Loh, H.H.; Law, P.-Y. Modulations of NeuroD Activity Contribute to the Differential Effects of Morphine and Fentanyl on Dendritic Spine Stability. J. Neurosci. 2010, 30, 8102–8110. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chu, J.; Zeng, Y.; Loh, H.H.; Law, P.-Y. Yin Yang 1 Phosphorylation Contributes to the Differential Effects of μ-Opioid Receptor Agonists on MicroRNA-190 Expression. J. Biol. Chem. 2010, 285, 21994–22002. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, C.; Kirkmire, C.M.; Wang, Z.J. Regulation of Opioid Tolerance by Let-7 Family MicroRNA Targeting the μ Opioid Receptor. J. Neurosci. 2010, 30, 10251–10258. [Google Scholar] [CrossRef]
- Sanchez-Simon, F.M.; Zhang, X.X.; Loh, H.H.; Law, P.-Y.; Rodriguez, R.E. Morphine Regulates Dopaminergic Neuron Differentiation via miR-133b. Mol. Pharmacol. 2010, 78, 935–942. [Google Scholar] [CrossRef]
- Yang, J.; Li, L.; Hong, S.; Zhang, D.; Zhou, Y. Methamphetamine Leads to the Alterations of microRNA Profiles in the Nucleus Accumbens of Rats. Pharm. Biol. 2020, 58, 797–805. [Google Scholar] [CrossRef]
- Cadet, J.L.; Patel, R.; Jayanthi, S. Compulsive Methamphetamine Taking and Abstinence in the Presence of Adverse Consequences: Epigenetic and Transcriptional Consequences in the Rat Brain. Pharmacol. Biochem. Behav. 2019, 179, 98–108. [Google Scholar] [CrossRef]
- Jan, R.K.; Kydd, R.R.; Russell, B.R. Functional and Structural Brain Changes Associated with Methamphetamine Abuse. Brain Sci. 2012, 2, 434–482. [Google Scholar] [CrossRef]
- Xie, T.; Tong, L.; Barrett, T.; Yuan, J.; Hatzidimitriou, G.; McCann, U.D.; Becker, K.G.; Donovan, D.M.; Ricaurte, G.A. Changes in Gene Expression Linked to Methamphetamine-Induced Dopaminergic Neurotoxicity. J. Neurosci. 2002, 22, 274–283. [Google Scholar] [CrossRef]
- Chand, S.; Gowen, A.; Savine, M.; Moore, D.; Clark, A.; Huynh, W.; Wu, N.; Odegaard, K.; Weyrich, L.; Bevins, R.A.; et al. A Comprehensive Study to Delineate the Role of an Extracellular Vesicle-associated microRNA-29a in Chronic Methamphetamine Use Disorder. J. Extracell. Vesicle 2021, 10, e12177. [Google Scholar] [CrossRef] [PubMed]
- Du, H.-Y.; Cao, D.-N.; Chen, Y.; Wang, L.; Wu, N.; Li, J. Alterations of Prefrontal Cortical microRNAs in Methamphetamine Self-Administering Rats: From Controlled Drug Intake to Escalated Drug Intake. Neurosci. Lett. 2016, 611, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhao, Y.; Zhang, K.; Su, H.; Chen, T.; Jiang, H.; Du, J.; Zhong, N.; Yu, S.; Zhao, M. An Association Study between Methamphetamine Use Disorder with Psychosis and Polymorphisms in MiRNA. Neurosci. Lett. 2020, 717, 134725. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhu, J.; Liu, Y.; Chen, Y.; Li, Y.; Chen, S.; Li, T.; Dang, Y.; Chen, T. Chronic Methamphetamine Regulates the Expression of MicroRNAs and Putative Target Genes in the Nucleus Accumbens of Mice: METH-Regulated miRNAs and Putative Targets. J. Neurosci. Res. 2015, 93, 1600–1610. [Google Scholar] [CrossRef]
- Kim, B.; Tag, S.H.; Kim, Y.S.; Cho, S.N.; Im, H.-I. Circulating microRNA miR-137 as a Stable Biomarker for Methamphetamine Abstinence. Psychopharmacology 2022, 239, 831–840. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, K.; Jiang, H.; Du, J.; Na, Z.; Hao, W.; Yu, S.; Zhao, M. Decreased Expression of Plasma MicroRNA in Patients with Methamphetamine (MA) Use Disorder. J. Neuroimmune Pharmacol. 2016, 11, 542–548. [Google Scholar] [CrossRef]
- Demirel, G.; Tanoglu, E.G.; Aslıyuksek, H. Evaluation of microRNA Let-7b-3p Expression Levels in Methamphetamine Abuse. Rev. Assoc. Med. Bras. 2023, 69, e20221391. [Google Scholar] [CrossRef]
- Li, H.; Li, C.; Zhou, Y.; Luo, C.; Ou, J.; Li, J.; Mo, Z. Expression of microRNAs in the Serum Exosomes of Methamphetamine-Dependent Rats vs. Ketamine-Dependent Rats. Exp. Ther. Med. 2018, 15, 3369–3375. [Google Scholar] [CrossRef]
- Shang, Q.; Wang, J.; Xi, Z.; Gao, B.; Qian, H.; An, R.; Shao, G.; Liu, H.; Li, T.; Liu, X. Mechanisms Underlying microRNA-222-3p Modulation of Methamphetamine-Induced Conditioned Place Preference in the Nucleus Accumbens in Mice. Psychopharmacology 2022, 239, 2997–3008. [Google Scholar] [CrossRef]
- Sim, M.S.; Soga, T.; Pandy, V.; Wu, Y.S.; Parhar, I.S.; Mohamed, Z. MicroRNA Expression Signature of Methamphetamine Use and Addiction in the Rat Nucleus Accumbens. Metab. Brain Dis. 2017, 32, 1767–1783. [Google Scholar] [CrossRef]
- Rezai Moradali, S.; Soltanzadeh, H.; Montazam, H.; Asadi, Z.; Fathi, S. MicroRNA-127 and MicroRNA-132 Expression in Patients with Methamphetamine Abuse in East Azerbaijan, Iran: A Case-Control Study. Addict. Health 2022, 14, 214–217. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, T.; Zhao, W.; Ren, Z.; Wang, Y.; Zhou, Y.; Song, X.; Zhou, R.; Zhang, X.; Jiao, D. MicroRNA-181a Is Involved in Methamphetamine Addiction Through the ERAD Pathway. Front. Mol. Neurosci. 2021, 14, 667725. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Shen, K.; Bai, Y.; Chao, J.; Hu, G.; Zhang, Y.; Yao, H. Involvement of NLRP3 Inflammasome in Methamphetamine-Induced Microglial Activation through miR-143/PUMA Axis. Toxicol. Lett. 2019, 301, 53–63. [Google Scholar] [CrossRef]
- Yu, G.; Song, Y.; Xie, C.; Tao, L.; Wan, F.; Jiang, L.; Wang, J.; Tang, J. MiR-142a-3p and miR-155-5p Reduce Methamphetamine-Induced Inflammation: Role of the Target Protein Peli1. Toxicol. Appl. Pharmacol. 2019, 370, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhu, L.; Ni, T.; Guan, F.; Chen, Y.; Ma, D.; Goh, E.L.K.; Chen, T. Ago2 and Dicer1 Are Involved in METH-induced Locomotor Sensitization in Mice via Biogenesis of miRNA. Addict. Biol. 2019, 24, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Zhu, L.; Li, J.; Wang, R.; Liu, D.; Han, W.; Cadet, J.L.; Chen, T. Regulation of microRNA-29c in the Nucleus Accumbens Modulates Methamphetamine -Induced Locomotor Sensitization in Mice. Neuropharmacology 2019, 148, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Ni, T.; Li, Y.; Wang, R.; Hu, T.; Guan, F.; Zhu, L.; Han, W.; Chen, T. The Potential Involvement of miR-204-3p-Axon Guidance Network in Methamphetamine-Induced Locomotor Sensitization of Mice. Neurosci. Lett. 2019, 707, 134303. [Google Scholar] [CrossRef]
- Liu, D.; Liang, M.; Zhu, L.; Zhou, T.; Wang, Y.; Wang, R.; Wu, F.; Goh, E.L.K.; Chen, T. Potential Ago2/miR-3068-5p Cascades in the Nucleus Accumbens Contribute to Methamphetamine-Induced Locomotor Sensitization of Mice. Front. Pharmacol. 2021, 12, 708034. [Google Scholar] [CrossRef]
- Bosch, P.J.; Benton, M.C.; Macartney-Coxson, D.; Kivell, B.M. mRNA and microRNA Analysis Reveals Modulation of Biochemical Pathways Related to Addiction in the Ventral Tegmental Area of Methamphetamine Self-Administering Rats. BMC Neurosci. 2015, 16, 43. [Google Scholar] [CrossRef]
- Shi, J.-J.; Cao, D.-N.; Liu, H.-F.; Wang, Z.-Y.; Lu, G.-Y.; Wu, N.; Zhou, W.-H.; Li, J. Dorsolateral Striatal miR-134 Modulates Excessive Methamphetamine Intake in Self-Administering Rats. Metab. Brain Dis. 2019, 34, 1029–1041. [Google Scholar] [CrossRef]
- Qian, H.; Shang, Q.; Liang, M.; Gao, B.; Xiao, J.; Wang, J.; Li, A.; Yang, C.; Yin, J.; Chen, G.; et al. MicroRNA-31-3p/RhoA Signaling in the Dorsal Hippocampus Modulates Methamphetamine-Induced Conditioned Place Preference in Mice. Psychopharmacology 2021, 238, 3207–3219. [Google Scholar] [CrossRef]
- Zhou, Y.; Xiao, S.; Li, C.; Chen, Z.; Zhu, C.; Zhou, Q.; Ou, J.; Li, J.; Chen, Y.; Luo, C.; et al. Extracellular Vesicle-Encapsulated miR-183-5p from Rhynchophylline-Treated H9c2 Cells Protect against Methamphetamine-Induced Dependence in Mouse Brain by Targeting NRG1. Evid.-Based Complement. Altern. Med. 2021, 2021, 2136076. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, Q.; Jing, X.; Zhao, Y.; Jiang, H.; Du, J.; Yu, S.; Zhao, M. miR-181a Is a Negative Regulator of GRIA2 in Methamphetamine-Use Disorder. Sci. Rep. 2016, 6, 35691. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Tag, S.H.; Nam, E.; Ham, S.; Ahn, S.; Kim, J.; Cho, D.-W.; Lee, S.; Yang, Y.-S.; Lee, S.E.; et al. SYNCRIP Controls miR-137 and Striatal Learning in Animal Models of Methamphetamine Abstinence. Acta Pharm. Sin. B 2022, 12, 3281–3297. [Google Scholar] [CrossRef]
- Deng, B.; Zhang, Z.; Zhou, H.; Zhang, X.; Niu, S.; Yan, X.; Yan, J. MicroRNAs in Methamphetamine-Induced Neurotoxicity and Addiction. Front. Pharmacol. 2022, 13, 875666. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Inoue, K.; Ishii, J.; Vanti, W.B.; Voronov, S.V.; Murchison, E.; Hannon, G.; Abeliovich, A. A MicroRNA Feedback Circuit in Midbrain Dopamine Neurons. Science 2007, 317, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, W.-Y.; Mao, Y.-W.; Gräff, J.; Guan, J.-S.; Pan, L.; Mak, G.; Kim, D.; Su, S.C.; Tsai, L.-H. A Novel Pathway Regulates Memory and Plasticity via SIRT1 and miR-134. Nature 2010, 466, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Zovoilis, A.; Agbemenyah, H.Y.; Agis-Balboa, R.C.; Stilling, R.M.; Edbauer, D.; Rao, P.; Farinelli, L.; Delalle, I.; Schmitt, A.; Falkai, P.; et al. microRNA-34c Is a Novel Target to Treat Dementias: MicroRNA-34c Is a Novel Target to Treat Dementias. EMBO J. 2011, 30, 4299–4308. [Google Scholar] [CrossRef]
- Gorini, G.; Nunez, Y.O.; Mayfield, R.D. Integration of miRNA and Protein Profiling Reveals Coordinated Neuroadaptations in the Alcohol-Dependent Mouse Brain. PLoS ONE 2013, 8, e82565. [Google Scholar] [CrossRef]
- Nunez, Y.O.; Truitt, J.M.; Gorini, G.; Ponomareva, O.N.; Blednov, Y.A.; Harris, R.A.; Mayfield, R.D. Positively Correlated miRNA-mRNA Regulatory Networks in Mouse Frontal Cortex during Early Stages of Alcohol Dependence. BMC Genom. 2013, 14, 725. [Google Scholar] [CrossRef]
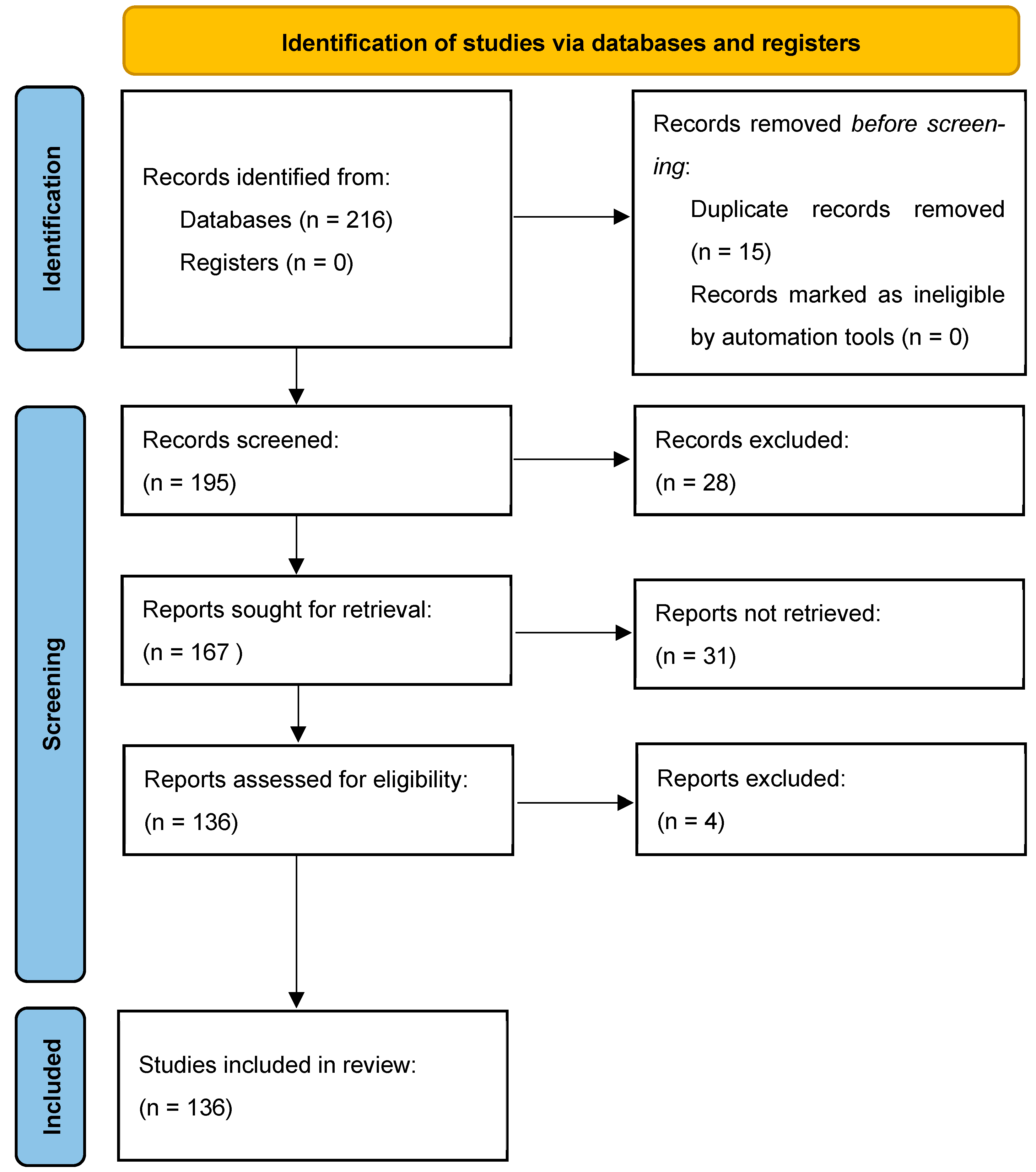
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).