
Neuroprotection and Mechanism of Gas-miR36-5p from Gastrodia elata in an Alzheimer’s Disease Model by Regulating Glycogen Synthase Kinase-3β
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Gas-miR-36-5p Regulates GSK-3β Expression by Directly Binding to the 3′-Untranslated Regions
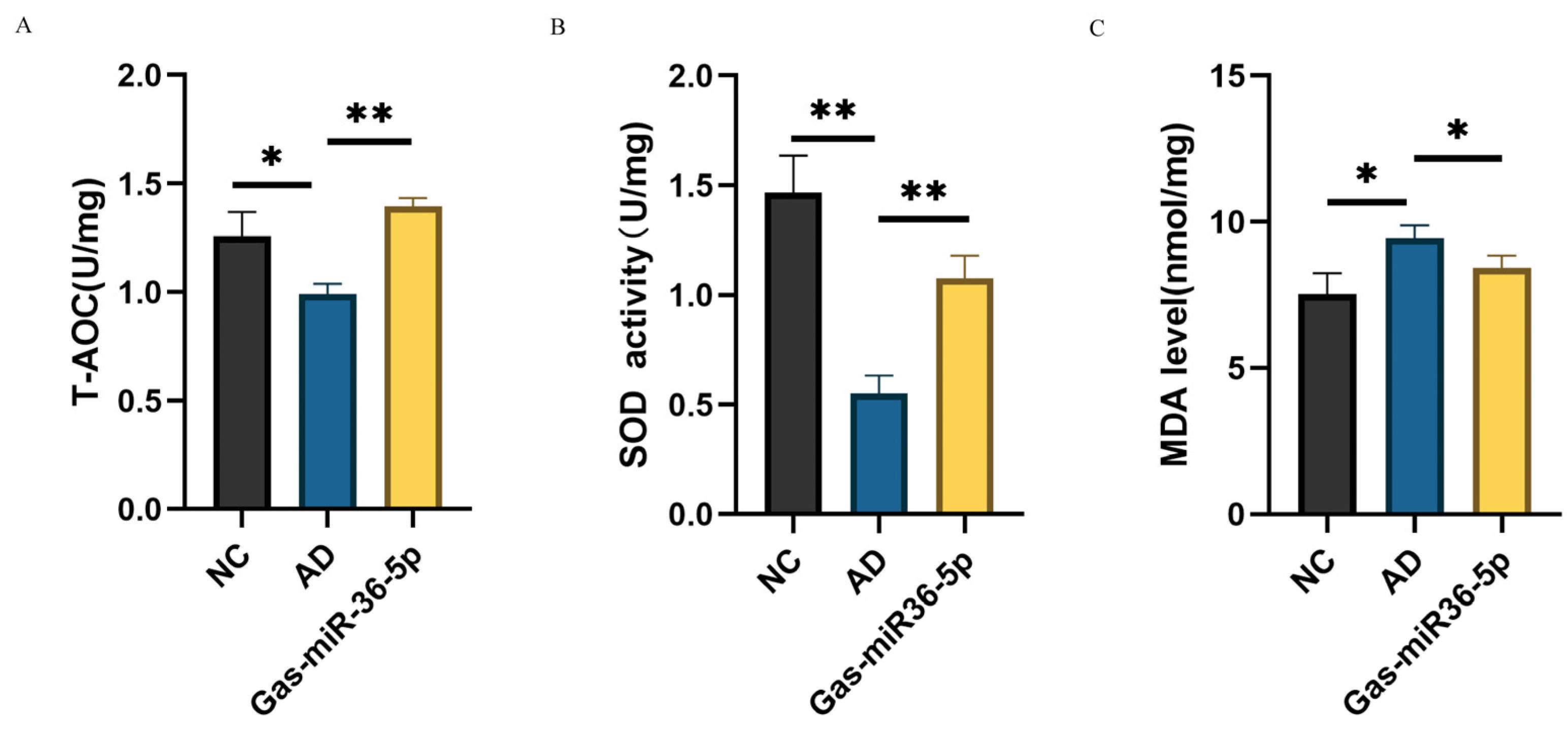
2.2. Oxidative Stress Indexes Show That Gas-miR36-5p Can Exert Neuroprotection in SH-SY5Y
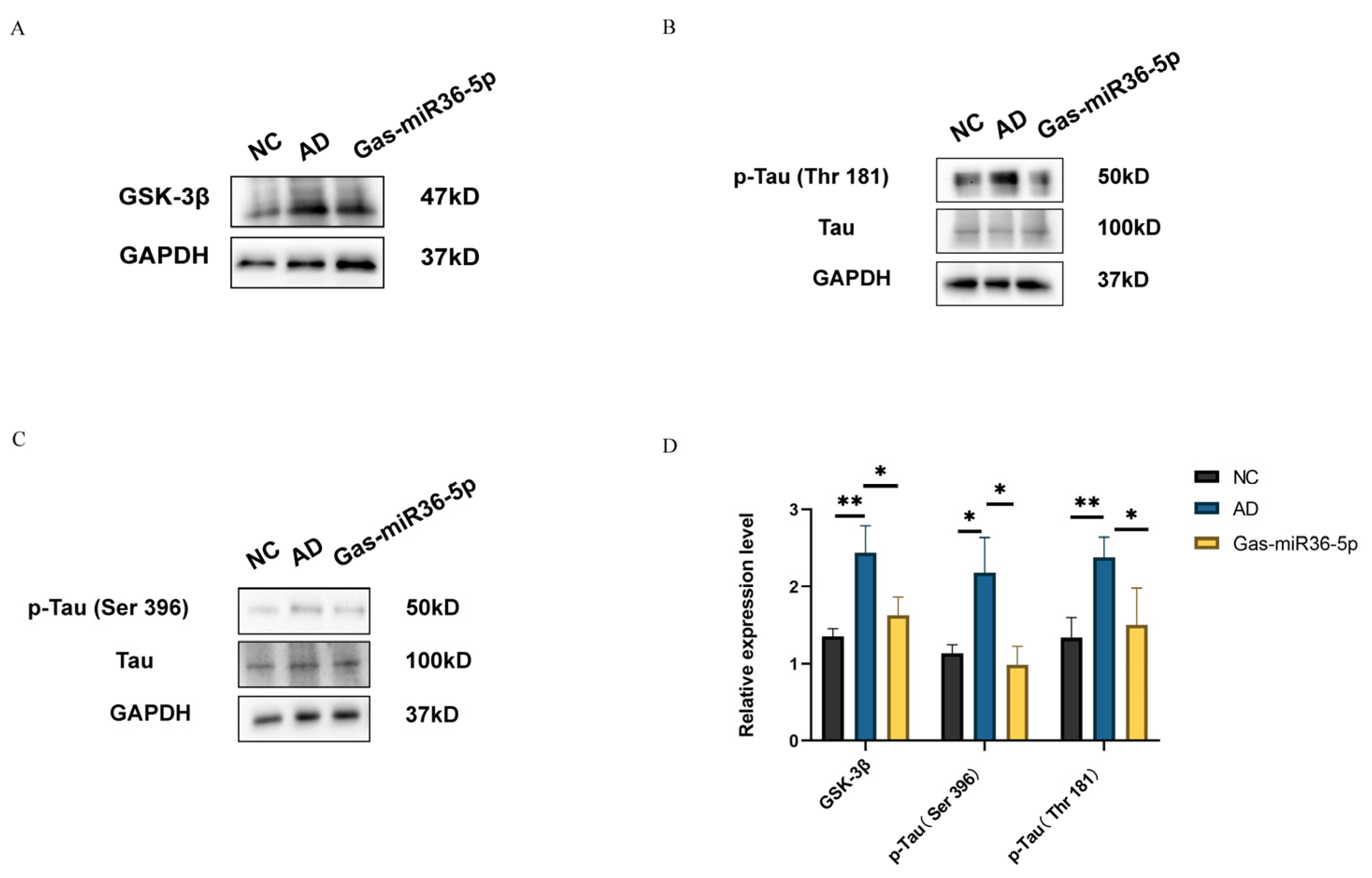
2.3. Gas-miR36-5p Suppresses Tau Phosphorylation in an AD Cell Model by Reducing the Activation of GSK-3β
2.4. Effect of Total RNA Extract from G. elata on Behavior in an AD Mouse Model
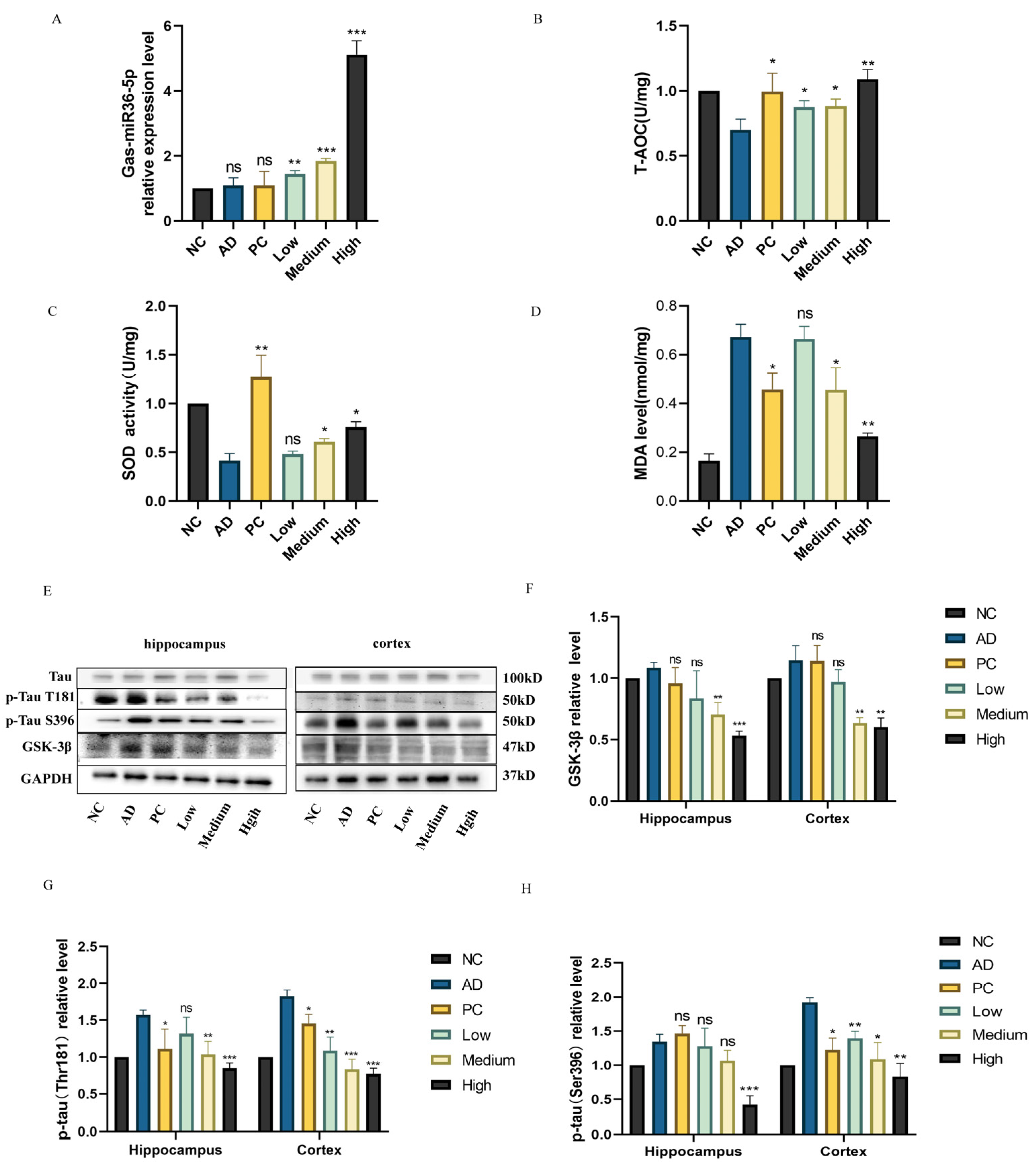
2.5. Regulation of G. elata-specific Gas-miR36-5p on the GSK-3β Signaling Pathway in an AD Mouse Model
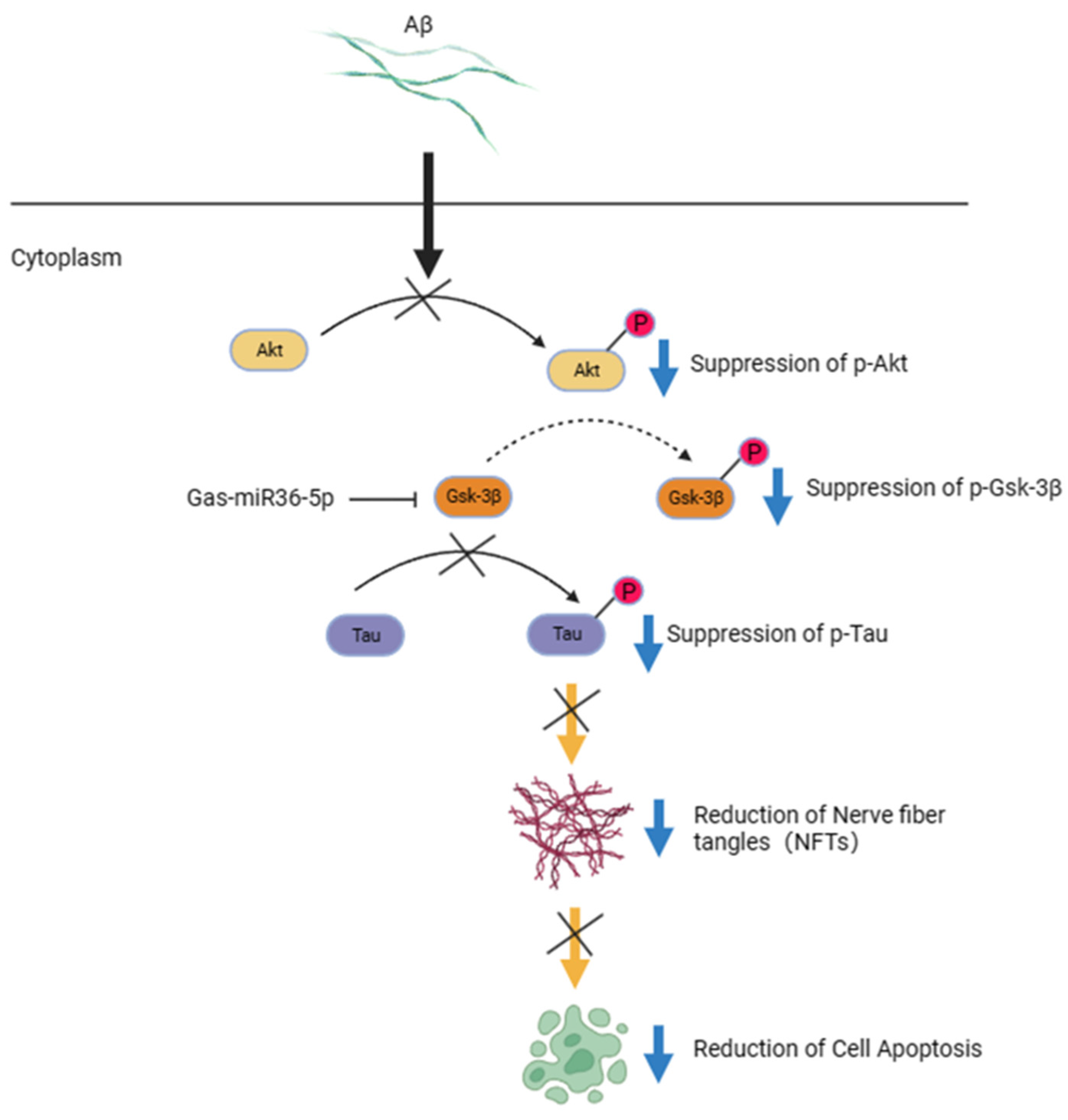
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. RNA Extraction and Real-Time Quantitative PCR
4.3. Target Prediction and Dual-Luciferase Reporter Assay
4.4. Protein Extraction and Western Blot
4.5. Animals and Treatments
4.6. Morris Water Maze Experiment
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qiu, C.; Kivipelto, M.; Von Strauss, E. Epidemiology of alzheimer’s disease: Occurrence, determinants, and strategies toward intervention. Dialogues Clin. Neurosci. 2009, 11, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Richard, H. Alzheimer’s disease. Nature 2018, 559, S1. [Google Scholar]
- Guzman-Martinez, L.; Calfío, C.; Farias, G.A.; Vilches, C.; Prieto, R.; Maccioni, R.B. New frontiers in the prevention, diagnosis, and treatment of alzheimer’s disease. J. Alzheimer’s Dis. 2021, 82, S51–S63. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.D.; Simms, G.A.; Weaver, D.F. The development of new therapeutics for alzheimer’s disease. Clin. Pharmacol. Ther. 2010, 88, 475–486. [Google Scholar] [CrossRef]
- Thal, D.R.; Walter, J.; Saido, T.C.; Fändrich, M. Neuropathology and biochemistry of aβ and its aggregates in alzheimer’s disease. Acta Neuropathol. 2015, 129, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.D.C.; Grundke-Iqbal, I.; Iqbal, K. Alzheimer’s disease hyperphosphorylated tau sequesters normal tau into tangles of filaments and disassembles microtubules. Nat. Med. 1996, 2, 783. [Google Scholar] [CrossRef]
- Wen, Y.; Planel, E.; Herman, M.; Figueroa, H.Y.; Wang, L.; Liu, L.; Lau, L.F.; Yu, W.H.; Duff, K.E. Duff. Interplay between cyclin-dependent kinase 5 and glycogen synthase kinase 3 beta mediated by neuregulin signaling leads to differential effects on tau phosphorylation and amyloid precursor protein processing. J. Neurosci. 2008, 28, 2624–2632. [Google Scholar] [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L.; Giménez-Llort, L. Emerging Translational Research in Neurological and Psychiatric Diseases: From In Vitro to In Vivo Models. Int. J. Mol. Sci. 2023, 24, 15739. [Google Scholar] [CrossRef]
- Busche, M.A.; Konnerth, A. Impairments of neural circuit function in Alzheimer’s disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150429. [Google Scholar] [CrossRef]
- Harris, S.S.; Wolf, F.; De Strooper, B.; Busche, M.A. Tipping the Scales: Peptide-Dependent Dysregulation of Neural Circuit Dynamics in Alzheimer’s Disease. Neuron 2020, 107, 417–435. [Google Scholar] [CrossRef]
- Ranasinghe, K.G.; Verma, P.; Cai, C.; Xie, X.; Kudo, K.; Gao, X.; Lerner, H.; Mizuiri, D.; Strom, A.; Iaccarino, L. Altered excitatory and inhibitory neuronal subpopulation parameters are distinctly associated with tau and amyloid in Alzheimer’s disease. eLife 2022, 11, e77850. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Latypova, X.; Wilson, C.M.; Magnaudeix, A.; Terro, F. Tau protein kinases: Involvement in alzheimer’s disease. Ageing Res. Rev. 2012, 12, 289–309. [Google Scholar] [CrossRef]
- Laurent, C.; Buée, L.; Blum, D. Tau and neuroinflammation: What impact for alzheimer’s disease and tauopathies? Biomed. J. 2018, 41, 21–33. [Google Scholar] [CrossRef]
- Liu, S.J.; Zhang, J.Y.; Li, H.L.; Fang, Z.Y.; Wang, Q.; Deng, H.M.; Gong, C.X.; Grundke-Iqbal, I.; Iqbal, K.; Wang, J.Z. Tau becomes a more favorable substrate for GSK-3 when it is prephosphorylated by PKA in rat brain. J. Biol. Chem. 2004, 279, 50078–50088. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, M.; Du, Y.; Zhang, W.; Bai, M.; Zhang, Z.; Li, Z.; Miao, J. Inhibition of c-Jun N-terminal kinase activation reverses Alzheimer disease phenotypes in APPswe/PS1dE9 mice. Ann. Neurol. 2015, 77, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.; Ammit, A.J. Targeting p38 MAPK pathway for the treatment of Alzheimer’s disease. Neuropharmacology 2010, 58, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Lan, G.; Li, R.; Mei, Y.; Shui, X.; Gu, X.; Wang, L.; Zhang, T.; Gan, C.L.; Xia, Y.; et al. Melatonin ameliorates tau-related pathology via the miR-504-3p and CDK5 axis in Alzheimer’s disease. Transl. Neurodegener. 2022, 11, 27. [Google Scholar] [CrossRef]
- Martinez, A.; Perez, D.I. GSK-3 inhibitors: A ray of hope for the treatment of Alzheimer’s disease? J. Alzheimer’s Dis. 2008, 15, 181–191. [Google Scholar] [CrossRef]
- Duda, P.; Akula, S.M.; Abrams, S.L.; Steelman, L.S.; Mccubrey, J.A. Targeting gsk3 and associated signaling pathways involved in cancer. Cells 2020, 9, 1110. [Google Scholar] [CrossRef]
- Bai, F.; Shi, Y.; Yuan, Y.; Yue, C.; Zhuang, L.; Xu, X.; Liu, X.; Zhang, Z. Association of a GSK-3β polymorphism with brain resting-state function in amnestic-type mild cognitive impairment. J. Alzheimer’s Dis. 2012, 32, 387–396. [Google Scholar] [CrossRef]
- Ishiguro, K.; Shiratsuchi, A.; Sato, S.; Omori, A.; Imahori, K. Glycogen synthase kinase 3 beta is identical to tau protein kinase i generating several epitopes of paired helical filaments. FEBS Lett. 1993, 325, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Zhou, W.; Liu, S.; Deng, Y.; Cai, F.; Tone, M.; Tone, Y.; Tong, Y.; Song, W. Increased nf-kb signalling up-regulates bace1 expression and its therapeutic potential in alzheimer’s disease. Int. J. Neuropsychopharmacol. 2012, 15, 77–90. [Google Scholar] [CrossRef]
- Liu, F.; Liang, Z.; Shi, J.; Yin, D.; El-Akkad, E.; Grundke-Iqbal, I.; Iqbal, K.; Gong, C.X. Pka modulates gsk-3beta- and cdk5-catalyzed phosphorylation of tau in site- and kinase-specific manners. FEBS. Lett. 2006, 580, 6269–6274. [Google Scholar] [CrossRef]
- Qi, Z.; Zhang, Y.; Yao, K.; Zhang, M.; Xu, Y.; Zhang, J.; Bai, X.; Zu, H. DHCR24 Knockdown Lead to Hyperphosphorylation of Tau at Thr181, Thr231, Ser262, Ser396, and Ser422 Sites by Membrane Lipid-Raft Dependent PP2A Signaling in SH-SY5Y Cells. Neurochem. Res. 2021, 46, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Sayas, C.L.; Ávila, J. GSK-3 and Tau: A Key Duet in Alzheimer’s Disease. Cells 2021, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.J.; Hernández, F.; Gómez-Ramos, P.; Morán, M.A.; Hen, R.; Avila, J. Decreased nuclear beta-catenin, tau hyperphosphorylation and neurodegeneration in gsk-3beta conditional transgenic mice. EMBO J. 2001, 20, 27–39. [Google Scholar] [CrossRef]
- Qu, C.; Li, Q.P.; Su, Z.R.; Ip, S.P.; Yuan, Q.J.; Xie, Y.L.; Xu, Q.Q.; Yang, W.; Huang, Y.F.; Xian, Y.F.; et al. Nano-honokiol ameliorates the cognitive deficits in tgcrnd8 mice of alzheimer’s disease via inhibiting neuropathology and modulating gut microbiota. J. Adv. Res. 2022, 35, 231–243. [Google Scholar] [CrossRef]
- Kaidanovich Beilin, O.; Lipina, T.V.; Takao, K.; van Eede, M.; Hattori, S.; Laliberté, C.; Khan, M.; Okamoto, K.; Chambers, J.W.; Fletcher, P.J.; et al. Abnormalities in brain structure and behavior in gsk-3alpha mutant mice. Mol. Brain 2009, 2, 35. [Google Scholar] [CrossRef]
- Liu, J.; Zuo, X.; Han, J.; Dai, Q.; Xu, H.; Liu, Y.; Cui, S. Mir-9-5p inhibits mitochondrial damage and oxidative stress in ad cell models by targeting gsk-3β. Biosci. Biotechnol. Biochem. 2020, 84, 2273–2280. [Google Scholar] [CrossRef]
- Jiang, H.; Liu, J.; Guo, S.; Zeng, L.; Cai, Z.; Zhang, J.; Wang, L.; Li, Z.; Liu, R. MiR-23b-3p rescues cognition in Alzheimer’s disease by reducing tau phosphorylation and apoptosis via GSK-3β signaling pathways. Mol. Ther.-Nucleic Acids 2022, 28, 539–557. [Google Scholar] [CrossRef]
- Kou, X.; Chen, D.; Chen, N. The Regulation of microRNAs in Alzheimer’s Disease. Front. Neurol. 2020, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Avila, J.; Hernández, F. Gsk-3 inhibitors for alzheimer’s disease. Expert Rev. Neurother. 2007, 7, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, H.; Zhang, J.G. Role and Biological Significance of GSK-3β Signal Pathway in Alzheimer’s Disease. Her. Med. 2020, 39, 1716–1720. (In Chinese) [Google Scholar]
- Rippin, I. Mechanisms and therapeutic implications of gsk-3 in treating neurodegeneration. Cells 2021, 10, 262. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Q.; Pan, X.; Zhang, B.; Wang, Q.; Pan, X. Micrornas and their regulatory roles in animals and plants. J. Cell. Physiol. 2007, 210, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Filipowicz, W.; Jaskiewicz, L.; Kolb, F.A.; Pillai, R.S. Post-transcriptional gene silencing by siRNAs and miRNAs. Curr. Opin. Struct. Biol. 2005, 15, 331–341. [Google Scholar] [CrossRef]
- Sun, M.; Xu, S.; Mei, Y.; Li, J.; Gu, Y.; Zhang, W.; Wang, J. MicroRNAs in medicinal plants. Int. J. Mol. Sci. 2022, 23, 10477. [Google Scholar] [CrossRef]
- Wang, W.; Liu, D.; Zhang, X.; Chen, D.; Cheng, Y.; Shen, F. Plant microRNAs in cross-kingdom regulation of gene expression. Int. J. Mol. Sci. 2018, 19, 2007. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D. Honeysuckle-encoded atypical microrna2911 directly targets influenza a virus. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef]
- Yuan, L.; Jialiang, G.; Min, P.; Hongyan, M.; Hongbo, M.; Pingping, C.; Yuan, X.; Qiong, Z.; Guomin, S. A review on central nervous system effects of gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar]
- Ojemann, L.M.; Nelson, W.L.; Shin, D.S.; Rowe, A.O.; Buchanan, R.A. Tian ma, an ancient chinese herb, offers new options for the treatment of epilepsy and other conditions. Epilepsy Behav. 2006, 8, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zhou, H.; Xu, X.; Jiang, T.; Li, S.; Wang, D.; Nie, Z.; Sheng, Q. Identification and investigation of miRNAs from Gastrodia elata blume and their potential function. Front. Pharmacol. 2020, 11, 542405. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Qin, L.; Tang, B. Micrornas in alzheimer’s disease. Front. Genet. 2019, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Deng, Y.; Ge, P.; Peng, Q.; Soufiany, I.; Zhu, L.; Duan, R. Diminazene ameliorates neuroinflammation by suppression of astrocytic mirna-224-5p/nlrp3 axis in alzheimer’s disease model. J. Inflamm. Res. 2023, 16, 1639–1652. [Google Scholar] [CrossRef]
- Xu, J.; Wu, K.J.; Jia, Q.J.; Ding, X.F. Roles of miRNA and lncRNA in triple-negative breast cancer. J. Zhejiang Univ. Sci. B 2020, 21, 673–689. [Google Scholar] [CrossRef]
- Xuan, C.; Yang, E.; Zhao, S.; Xu, J.; Li, P.; Zhang, Y.; Jiang, Z.; Ding, X. Regulation of LncRNAs and microRNAs in neuronal development and disease. PeerJ. 2023, 11, e15197. [Google Scholar] [CrossRef]
- Wei, B.; Huang, B.; Zhao, X. An overview of biochemical technologies for the cancer biomarker miR-21 detection. Anal. Sci. 2023, 39, 815–827. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, B.; Gao, C.; Zhang, H.; Wen, F.; Tao, L.; Fu, G.; Xiong, J. The Evolution and Functional Roles of miR408 and Its Targets in Plants. Int. J. Mol. Sci. 2022, 23, 530. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Wu, Y.; Niu, Q.; Miao, M.; Zhang, D.; Zhao, Y.; Cai, F.; Yu, D.; Ke, L.; Feng, H.; et al. Integrative Analysis of Expression Profiles of mRNA and MicroRNA Provides Insights of Cotton Response to Verticillium dahliae. Int. J. Mol. Sci. 2022, 23, 4702. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fan, H.; Pei, X.; Hua, X.; Xu, T.; He, Q. mRNA-Seq and miRNA-Seq Analyses Provide Insights into the Mechanism of Pinellia ternata Bulbil Initiation Induced by Phytohormones. Genes 2023, 14, 1727. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Wang, Z.; Wang, Y.; Liu, Z.; Wang, H.; Xu, B. Cross-kingdom regulation of plant-derived mirnas in modulating insect development. Int. J. Mol. Sci. 2023, 24, 7978. [Google Scholar] [CrossRef] [PubMed]
- Rabuma, T.; Gupta, O.P.; Chhokar, V. Recent advances and potential applications of cross-kingdom movement of mirnas in modulating plant’s disease response. RNA Biol. 2022, 19, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, S.; Ayachit, G.; Bhairappanavar, S.B.; Ansari, A.; Sharma, P.; Soni, S.; Das, J. Cross-kingdom regulation of putative mirnas derived from happy tree in cancer pathway: A systems biology approach. Int. J. Mol. Sci. 2017, 18, 1191. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hotz, T.; Broadnax, L.; Yarmarkovich, M.; Elbaz-Younes, I.; Hirschi, K.D. Anomalous uptake and circulatory characteristics of the plant-based small RNA MIR2911. Sci. Rep. 2016, 6, 26834. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Bloom, G.S. Amyloid-β and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef]
- Li, S.; Zhang, B. Traditional Chinese medicine network pharmacology: Theory, methodology and application. Chin. J. Nat. Med. 2013, 11, 110–120. [Google Scholar] [CrossRef]
- Zhan, H.D.; Zhou, H.Y.; Sui, Y.P.; Du, X.L.; Wang, W.H.; Dai, L.; Sui, F.; Huo, H.R.; Jiang, T.L. The rhizome of Gastrodia elata Blume—An ethnopharmacological review. J. Ethnopharmacol. 2016, 189, 361–385. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Application | |
|---|---|---|---|
| Gas-miR36-5p | Loop-stem primer | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGAACAAAAT | Reverse transcription |
| Forward | GCCGAGTGCAGATGACTTG | qRT-PCR | |
| Reverse | CTCAACTGGTGTCGTGGA | ||
| 5.8 s rRNA | Forward | ATCACTCGGCTCGTGCGTC | qRT-PCR |
| Reverse | CAAGTGCGTTCGAAGTGTCG | ||
| GSK-3β | Forward | AGGAGAACCCAATGTTTCGTAT | qRT-PCR |
| Reverse | ATCCCCTGGAAATATTGGTTGT | ||
| GAPDH | Forward | AATGGATTTGGACGCATTGGT | qRT-PCR |
| Reverse | TTTGCACTGGTACGTGTTGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Fu, J.; Wu, G.; Yang, Z.; Wu, X.; Wang, D.; You, Z.; Nie, Z.; Sheng, Q. Neuroprotection and Mechanism of Gas-miR36-5p from Gastrodia elata in an Alzheimer’s Disease Model by Regulating Glycogen Synthase Kinase-3β. Int. J. Mol. Sci. 2023, 24, 17295. https://doi.org/10.3390/ijms242417295
Lu Z, Fu J, Wu G, Yang Z, Wu X, Wang D, You Z, Nie Z, Sheng Q. Neuroprotection and Mechanism of Gas-miR36-5p from Gastrodia elata in an Alzheimer’s Disease Model by Regulating Glycogen Synthase Kinase-3β. International Journal of Molecular Sciences. 2023; 24(24):17295. https://doi.org/10.3390/ijms242417295
Chicago/Turabian StyleLu, Zhongteng, Jianyuan Fu, Guang Wu, Zhecheng Yang, Xiaoqi Wu, Dan Wang, Zhengying You, Zuoming Nie, and Qing Sheng. 2023. "Neuroprotection and Mechanism of Gas-miR36-5p from Gastrodia elata in an Alzheimer’s Disease Model by Regulating Glycogen Synthase Kinase-3β" International Journal of Molecular Sciences 24, no. 24: 17295. https://doi.org/10.3390/ijms242417295
APA StyleLu, Z., Fu, J., Wu, G., Yang, Z., Wu, X., Wang, D., You, Z., Nie, Z., & Sheng, Q. (2023). Neuroprotection and Mechanism of Gas-miR36-5p from Gastrodia elata in an Alzheimer’s Disease Model by Regulating Glycogen Synthase Kinase-3β. International Journal of Molecular Sciences, 24(24), 17295. https://doi.org/10.3390/ijms242417295